UPLC-ESI-Q-TOF-MS-Based Metabolite Profiling, Antioxidant and Anti-Inflammatory Properties of Different Organ Extracts of Abeliophyllum distichum


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and UPLC-ESI-Q-TOF-MS-Based Metabolomic Analysis of A. distichum Organ Extracts
2.2. Determination of Antioxidant Properties by Chemical-Based Assays
2.3. Analysis of Nitric Oxide (NO) Production and Cell Viability
2.4. DCFH-DA (Dichlorofluorescein Diacetate) Assay
2.5. Western Blot
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Statistical Analyses
3. Results and Discussion
3.1. Identification of the Chemical Composition of A. distichum Organ Extracts Using UPLC-ESI-Q-TOF-MS
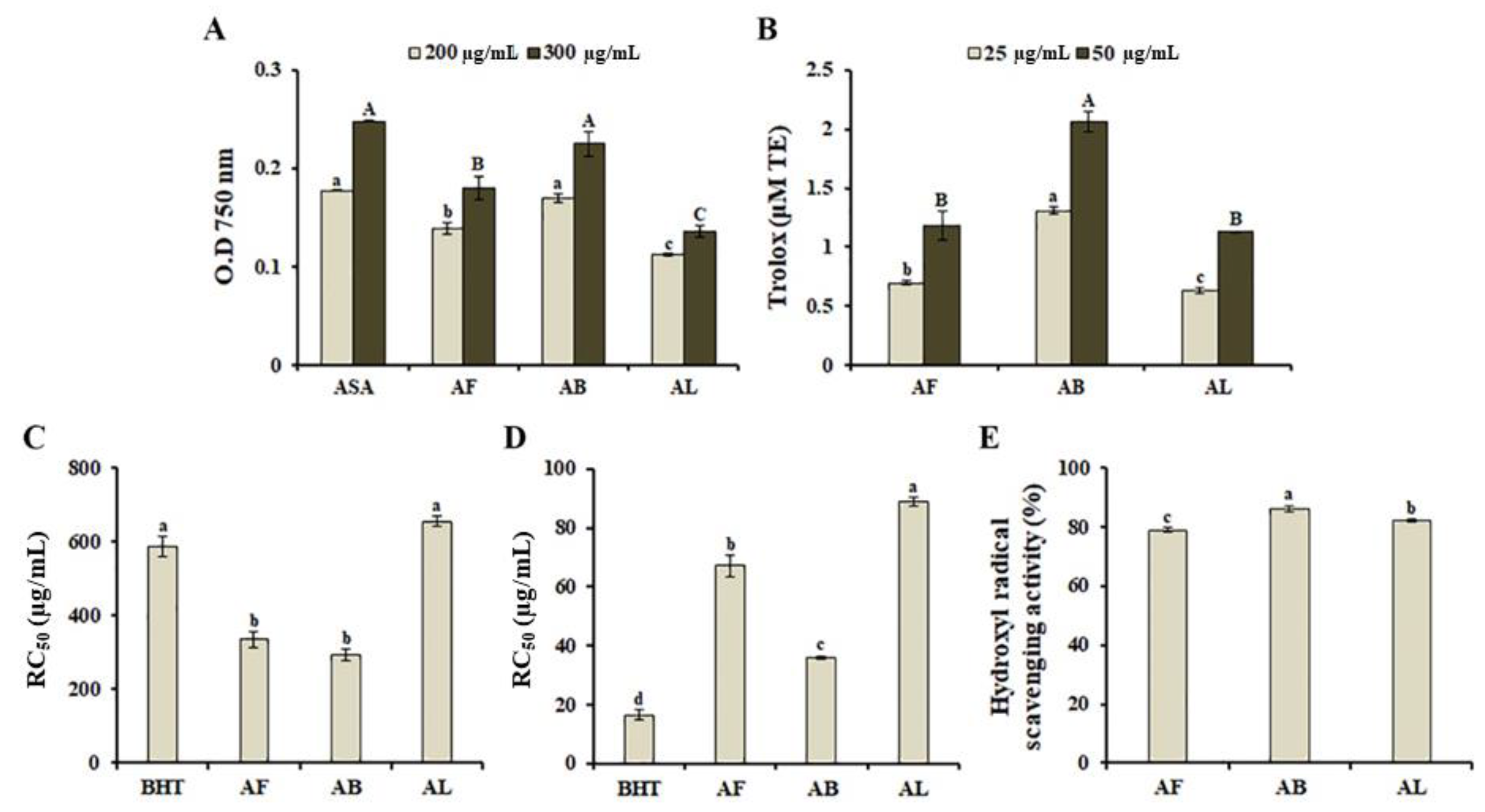
3.2. In-Vitro Antioxidant Properties of A. distichum Organ Extracts
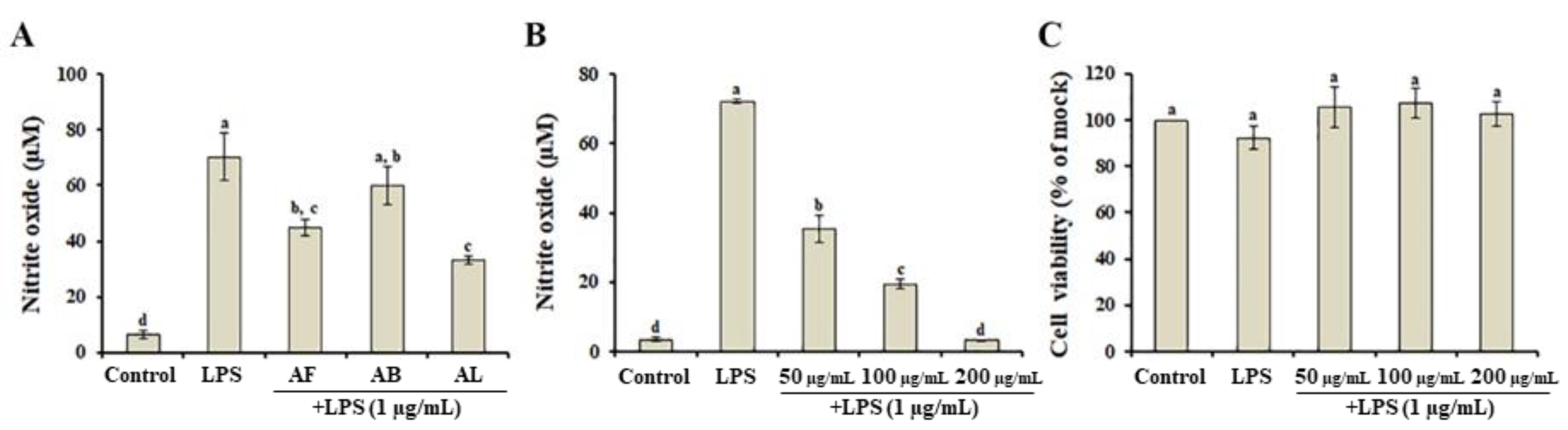
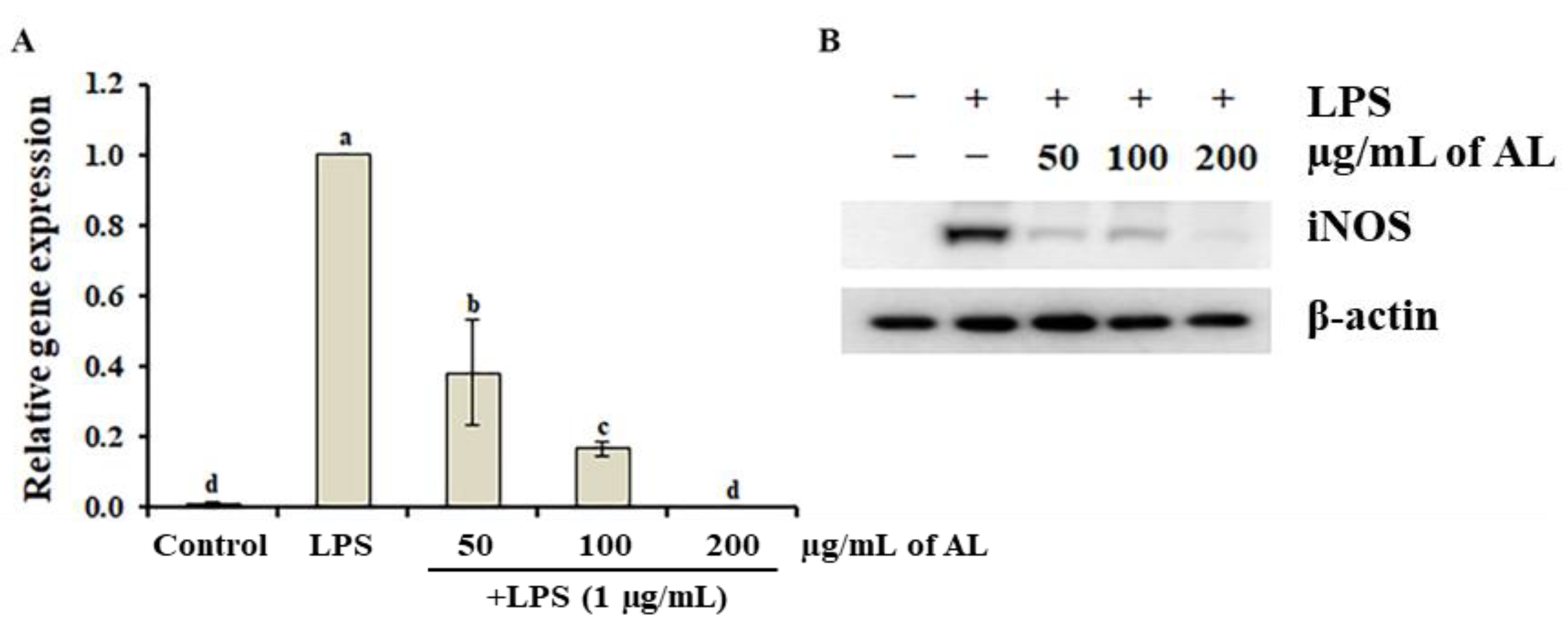
3.3. Effects of A. distichum Organ Extracts on LPS-Induced NO Production
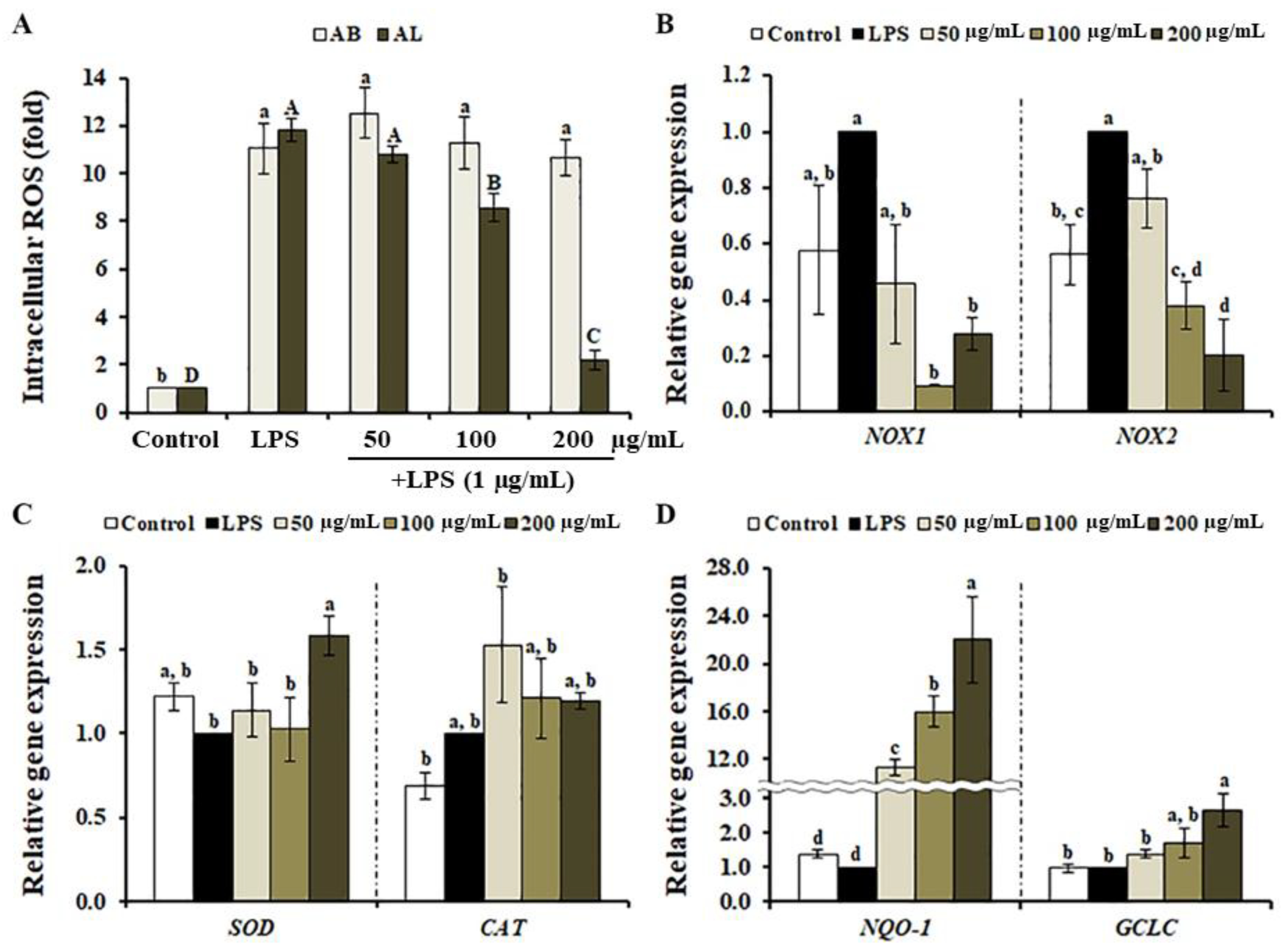
3.4. Effects of A. distichum Leaf Extract on LPS-Induced ROS Production in RAW264.7 Cells
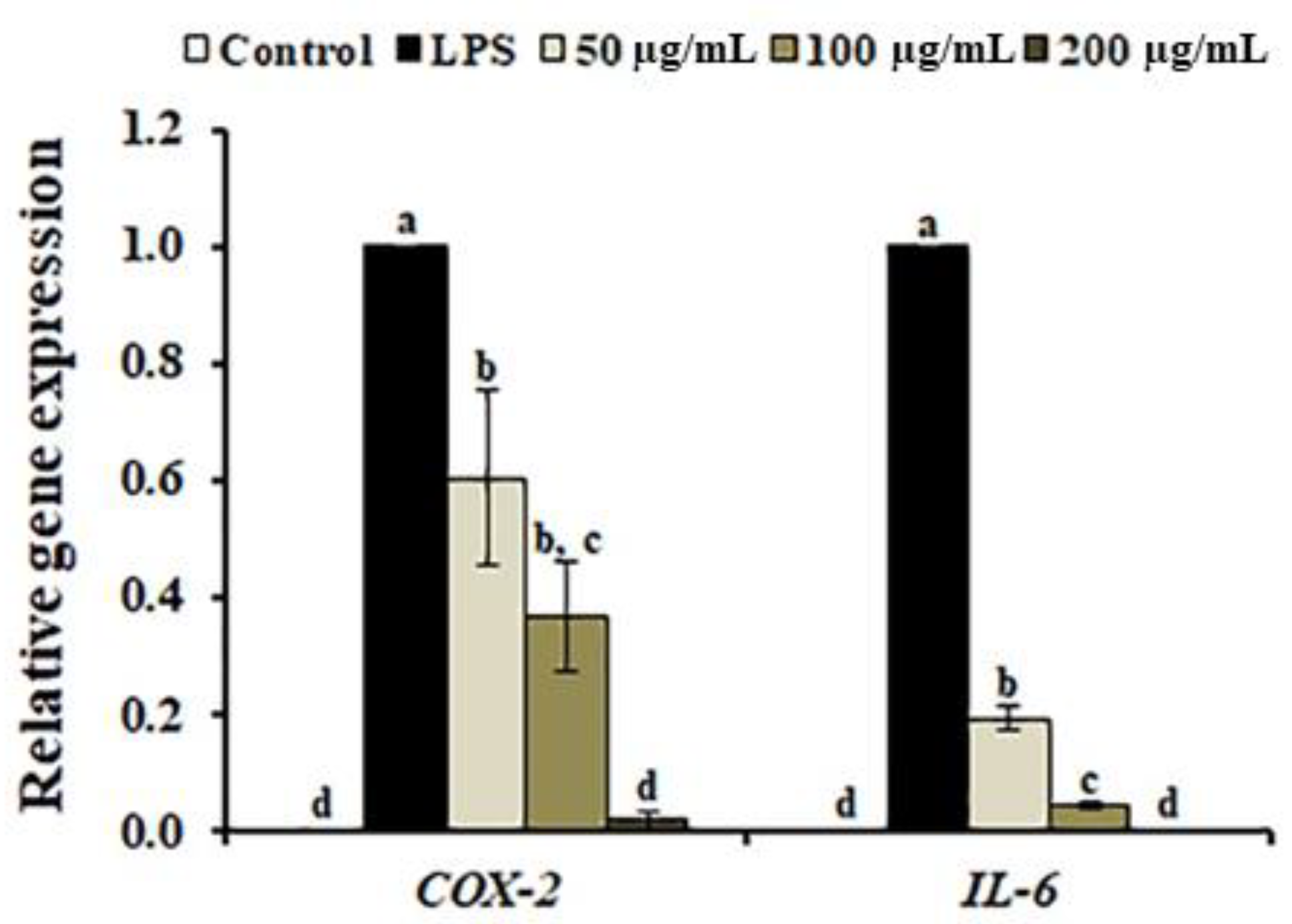
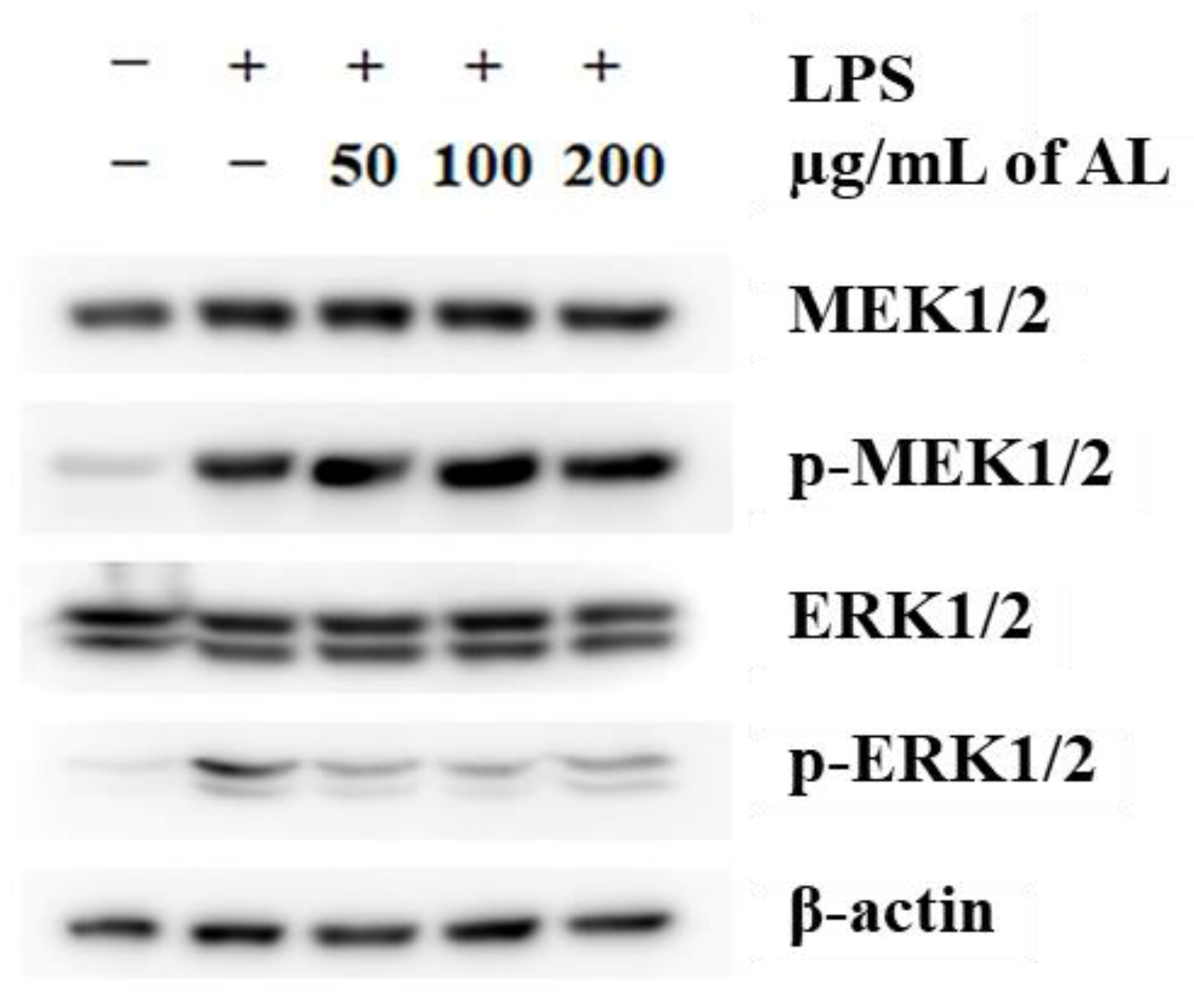
3.5. Effects of A. distichum Leaf Extract on the LPS-Induced MEK-ERK1/2 Pathway in RAW264.7 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids. Mechanisms of action in health and disease. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Süleyman, H.; Demircan, B.; Karagöz, Y. Anti-inflammatory and side effects of cyclooxygenase inhibitors. Pharmacol. Rep. 2007, 59, 247–258. [Google Scholar] [PubMed]
- Wongrakpanich, S.; Wongrakpanich, A.; Melhado, K.; Rangaswami, J. A comprehensive review of non-steroidal anti-inflammatory drug use in the elderly. Aging Dis. 2018, 9, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Gupta, E.; Mishra, N.; Mishra, P. Shikimic acid as intermediary model for the production of drugs effective against influenza virus. Phytochem. Lead Compd. New Drug Discov. 2020, 245–256. [Google Scholar]
- Bochkov, D.V.; Sysolyatin, S.V.; Kalashnikov, A.I.; Surmacheva, I.A. Shikimic acid: Review of its analytical, isolation, and purification techniques from plant and microbial sources. J. Chem. Biol. 2012, 5, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Perini, J.A.; Angeli-Gamba, T.; Alessandra-Perini, J.; Ferreira, L.C.; Nasciutti, L.E.; Machado, D.E. Topical application of Acheflan on rat skin injury accelerates wound healing: A histopathological, immunohistochemical and biochemical study. BMC Complement. Altern. Med. 2015, 30, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-inflammatory activity of natural products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Huang, Y.L.; Oppong, M.B.; Guo, Y.; Wang, L.Z.; Fang, S.M.; Deng, Y.R.; Gao, X.M. The Oleaceae family: A source of secoiridoids with multiple biological activities. Fitoterapia 2019, 136, 104155. [Google Scholar] [CrossRef]
- Choi, J.H.; Seo, E.J.; Sung, J.; Choi, K.M.; Kim, H.; Kim, J.S.; Lee, J.; Efferth, T.; Hyun, T.K. Polyphenolic compounds, antioxidant and anti-inflammatory effects of Abeliophyllum distichum Nakai extract. J. Appl. Bot. Food Qual. 2017, 90, 266–273. [Google Scholar]
- Jang, T.; Choi, J.; Kim, H.; Lee, E.; Han, M.; Lee, K.; Kim, D.; Park, J.-H. Whitening activity of Abeliophyllum distichum Nakai leaves according to the ratio of prethanol A in the extracts. Korean J. Plant Reources. 2018, 31, 667–674. [Google Scholar]
- Li, H.M.; Kim, J.K.; Jang, J.M.; Cui, C.B.; Lim, S.S. Analysis of the inhibitory activity of Abeliophyllum distichum leaf constituents against aldose reductase by using high-speed counter current chromatography. Arch. Pharm. Res. 2013, 36, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Yoo, T.K.; Kim, J.S.; Hyun, T.K. Polyphenolic composition and anti-melanoma activity of white forsythia (Abeliophyllum distichum Nakai) organ extracts. Plants 2020, 9, 757. [Google Scholar] [CrossRef]
- Lee, H.D.; Kim, J.H.; Pang, Q.Q.; Jung, P.M.; Cho, E.J.; Lee, S. Antioxidant activity and acteoside analysis of Abeliophyllum distichum. Antioxidants 2020, 9, 1148. [Google Scholar] [CrossRef]
- Nam, S.Y.; Kim, H.Y.; Yoou, M.S.; Kim, A.H.; Park, B.J.; Jeong, H.J.; Kim, H.M. Anti-inflammatory effects of isoacteoside from Abeliophyllum distichum. Immunopharmacol. Immunotoxicol. 2015, 37, 258–264. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, H.; Hyun, T.K. Transcriptome analysis of Abeliophyllum distichum NAKAI reveals potential molecular markers and candidate genes involved in anthocyanin biosynthesis pathway. S. Afr. J. Bot. 2018, 116, 34–41. [Google Scholar] [CrossRef]
- Han, Y.X.; Wang, P.F.; Zhao, M.; Chen, L.M.; Wang, Z.M.; Liu, X.Q.; Gao, H.M.; Gong, M.X.; Li, H.; Zhu, J.Z.; et al. Chemical profiling of xueshuan xinmaining tablet by HPLC and UPLC-ESI-Q-TOF/MS. Evid. Based Complement. Altern. Med. 2018, 2018, 2781597. [Google Scholar] [CrossRef]
- Hyun, T.K.; Lee, S.; Rim, Y.; Kumar, R.; Han, X.; Lee, S.Y.; Lee, C.H.; Kim, J.Y. De-novo RNA sequencing and metabolite profiling to identify genes involved in anthocyanin biosynthesis in Korean black raspberry (Rubus coreanus Miquel). PLoS ONE 2014, 9, e88292. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yu, Y.; Shi, R.; Xie, G.; Zhu, Y.; Wu, G.; Qin, M. Organ-specific metabolic shifts of flavonoids in Scutellaria baicalensis at different growth and development stages. Molecules 2018, 23, 428. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hu, Z. Accumulation and dynamic trends of triterpenoid saponin in vegetative organs of Achyranthus bidentata. J. Integr. Plant Biol. 2009, 51, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Lee, J.K.; Kim, H.; Hyun, T.K. De novo transcriptomic analysis to reveal functional genes involved in triterpenoid saponin biosynthesis in Oplopanax elatus NAKAI. J. Appl. Bot. Food Qual. 2017, 90, 25–31. [Google Scholar]
- Sindhi, V.; Gupta, V.; Sharma, K.; Bhatnagar, S.; Kumari, R.; Dhaka, N. Potential applications of antioxidants—A review. J. Pharm. Res. 2013, 7, 828–835. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Hou, A.J.; Zhu, G.F.; Chen, D.F.; Sun, H.D. Cytotoxic and antifungal isoprenylated xanthones and flavonoids from Cudrania fruticosa. Planta Med. 2005, 71, 273–274. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuan, W.; Wang, P.; Grant, G.; Li, S. Flavonoids from Lupinus texensis and their free radical scavenging activity. Nat. Prod. Res. 2011, 25, 1641–1649. [Google Scholar] [CrossRef]
- Wei, S.; Wu, W.; Ji, Z. New antifungal pyranoisoflavone from Ficus tikoua bur. Int. J. Mol. Sci. 2012, 13, 7375–7382. [Google Scholar] [CrossRef] [Green Version]
- Akter, K.; Barnes, E.C.; Loa-Kum-Cheung, W.L.; Yin, P.; Kichu, M.; Brophy, J.J.; Barrow, R.A.; Imchen, I.; Vemulpad, S.R.; Jamie, J.F. Antimicrobial and antioxidant activity and chemical characterisation of Erythrina stricta Roxb. (Fabaceae). J. Ethnopharmacol. 2016, 185, 171–181. [Google Scholar] [CrossRef]
- Lowry, J.L.; Brovkovych, V.; Zhang, Y.; Skidgel, R.A. Endothelial nitric-oxide synthase activation generates an inducible nitric-oxide synthase-like output of nitric oxide in inflamed endothelium. J. Biol. Chem. 2013, 288, 4174–4193. [Google Scholar] [CrossRef] [Green Version]
- Rapovy, S.M.; Zhao, J.; Bricker, R.L.; Schmidt, S.M.; Setchell, K.D.R.; Qualls, J.E. Differential requirements for l-citrulline and l-arginine during antimycobacterial macrophage activity. J. Immunol. 2015, 195, 3293–3300. [Google Scholar] [CrossRef] [Green Version]
- Sarti, P.; Forte, E.; Mastronicola, D.; Giuffrè, A.; Arese, M. Cytochrome c oxidase and nitric oxide in action: Molecular mechanisms and pathophysiological implications. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Grottelli, S.; Amoroso, R.; Macchioni, L.; D’Onofrio, F.; Fettucciari, K.; Bellezza, I.; Maccallini, C. Acetamidine-based iNOS inhibitors as molecular tools to counteract inflammation in BV2 microglial cells. Molecules 2020, 25, 2646. [Google Scholar] [CrossRef]
- Muniandy, K.; Gothai, S.; Badran, K.M.H.; Kumar, S.S.; Esa, N.M.; Arulselvan, P. Suppression of proinflammatory cytokines and mediators in LPS-Induced RAW 264.7 macrophages by stem extract of Alternanthera sessilis via the inhibition of the NF-κB pathway. J. Immunol. Res. 2018, 2018, 3430684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Chen, D.; Li, M.; Gao, X.; Shi, G.; Zhao, H. Plantamajoside inhibits lipopolysaccharide-induced epithelial-mesenchymal transition through suppressing the NF-κB/IL-6 signaling in esophageal squamous cell carcinoma cells. Biomed. Pharmacother. 2018, 102, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.G.; Kim, A.R.; Ko, H.J.; Lee, S.K.; Woo, E.R. Three new lignan glycosides with IL-6 inhibitory activity from Akebia quinata. Chem. Pharm. Bull. 2014, 62, 288–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, H.; Ku, S.K.; Baek, Y.D.; Bae, J.S. Anti-inflammatory effects of rutin on HMGB1-induced inflammatory responses in vitro and in vivo. Inflamm. Res. 2014, 63, 197–206. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Meng, T.; Yu, J.; Lei, Z.; Wu, J.; Wang, S.; Bo, Q.; Zhang, X.; Ma, Z.; Yu, J. Propofol reduces lipopolysaccharide-induced, NADPH oxidase (NOX 2) mediated TNF- α and IL-6 production in macrophages. Clin. Dev. Immunol. 2013, 2013, 325481. [Google Scholar] [CrossRef] [Green Version]
- Tucsek, Z.; Radnai, B.; Racz, B.; Debreceni, B.; Priber, J.K.; Dolowschiak, T.; Palkovics, T.; Gallyas, F.; Sumegi, B.; Veres, B. Suppressing LPS-induced early signal transduction in macrophages by a polyphenol degradation product: A critical role of MKP-1. J. Leukoc. Biol. 2011, 89, 105–111. [Google Scholar] [CrossRef]
- Ryan, K.A.; Smith, M.F.; Sanders, M.K.; Ernst, P.B. Reactive oxygen and nitrogen species differentially regulate toll-like receptor 4-mediated activation of NF-κB and interleukin-8 expression. Infect. Immun. 2004, 72, 2123–2130. [Google Scholar] [CrossRef] [Green Version]
- Grauzdytė, D.; Pukalskas, A.; Viranaicken, W.; Kalamouni, C.E.; Venskutonis, P.R. Protective effects of Phyllanthus phillyreifolius extracts against hydrogen peroxide induced oxidative stress in HEK293 cells. PLoS ONE 2018, 13, e0207672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Zhou, J.; Lv, H.; Yu, Q.; Peng, L.; Hua, S. Betulin exhibits anti-inflammatory activity in LPS-stimulated macrophages and endotoxin-shocked mice through an AMPK/AKT/Nrf2-dependent mechanism. Cell Death Dis. 2017, 8, e2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guangwei, Z.; An, H.; Dong, F.; Wei, W.; Yang, H.; Cheng, L. Plantamajoside protects H9c2 cells against hypoxia/reoxygenation-induced injury through regulating the akt/Nrf2/HO-1 and NF-κB signaling pathways. J. Recept. Signal Transduct. Res. 2020, 22, 1–8. [Google Scholar]
- Shaul, Y.D.; Seger, R. The MEK/ERK cascade: From signaling specificity to diverse functions. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1213–1226. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, T.; Zhou, H.; Zhang, C.; Feng, Y.; Tang, F.; Hoi, M.P.M.; He, C.; Zheng, Y.; Lee, S.M.Y. Schisantherin a attenuates neuroinflammation in activated microglia: Role of Nrf2 activation through ERK phosphorylation. Cell. Physiol. Biochem. 2018, 47, 1769–1784. [Google Scholar] [CrossRef]
- Hua, K.F.; Chou, J.C.; Lam, Y.; Tasi, Y.L.; Chen, A.; Ka, S.M.; Fang, Z.; Liu, M.L.; Yang, F.L.; Yang, Y.L.; et al. Polyenylpyrrole Derivatives Inhibit NLRP3 Inflammasome Activation and Inflammatory Mediator Expression by Reducing Reactive Oxygen Species Production and Mitogen-Activated Protein Kinase Activation. PLoS ONE 2013, 8, e76754. [Google Scholar] [CrossRef] [Green Version]
- Scherle, P.A.; Jones, E.A.; Favata, M.F.; Daulerio, A.J.; Covington, M.B.; Nurnberg, S.A.; Magolda, R.L.; Trzaskos, J.M. Inhibition of MAP kinase kinase prevents cytokine and prostaglandin E2 production in lipopolysaccharide-stimulated monocytes. J. Immunol. 1998, 161, 5681–5686. [Google Scholar]
- Low, H.B.; Zhang, Y. Regulatory roles of MAPK phosphatases in cancer. Immune Netw. 2016, 16, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Bognar, E.; Sarszegi, Z.; Szabo, A.; Debreceni, B.; Kalman, N.; Tucsek, Z.; Sumegi, B.; Gallyas, F. Antioxidant and anti-Inflammatory effects in RAW264.7 macrophages of malvidin, a major red wine polyphenol. PLoS ONE 2013, 8, e65355. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Rt (min) | Neutral Mass (Da) | Observed m/z | Mass Error (mDa) | Formula | Proposed Molecule | Fragment Ions | Organ |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.52 | 192.0634 | 191.0555 | −0.6 | C7H12O6 | D-(−)-Quinic acid | 173.04, 149.04, 127.04, 109.03, 93.03, 85.03 | AF, AB, AL |
| 2 | 5.43 | 446.2152 | 445.2074 | −0.5 | C21H34O10 | Dendromoniliside D | 401.14, 269.10, 229.07, 185.04, 141.02, 103.04 | AL |
| 3 | 7.02 | 772.2426 | 771.2342 | −1.1 | C32H36O22 | α-Glucosyl hesperidin | 661.18, 607.19, 591.19, 229.07, 179.03, 161.02 | AB |
| 4 | 7.09 | 640.2003 | 639.1922 | −0.9 | C29H36O16 | Plantamajoside | 475.14, 185.04, 179.03, 161.02 | AL |
| 5 | 7.37 | 478.1475 | 477.1399 | −0.4 | C23H26O11 | Calceolarioside B | 221.04, 161.02 | AL |
| 6 | 7.50 | 786.2582 | 785.2512 | −0.3 | C35H46O20 | Echinacoside | 624.22, 429.15, 387.15, 305.09, 179.03, 161.02, 135.04 | AF |
| 7 | 8.01 | 610.1534 | 609.1459 | −0.2 | C27H30O16 | Rutin | 301.03, 300.03, 299.02 | AF, AB, AL |
| 8 | 8.14 | 488.3138 | 487.3055 | −1.0 | C24H40O10 | Senegenic acid | 367.27, 309.22, 290.22, 225.16 | AF, AL |
| 9 | 8.48 | 462.1526 | 461.1451 | −0.2 | C23H26O10 | 6′-coumaroyl-1′-O-[2-(3,4-dihydroxyphenyl)ethyl]-β-d-glucopyranoside | 265.07, 205.05, 163.04, 145.03 | AF, AL |
| 10 | 8.56 | 464.1682 | 463.1599 | −1.1 | C21H20O12 | Isomucronulatol 7-O-Glucoside | 341.06, 327.04, 222.09, 141.02, 137.02 | AL |
| 11 | 8.64 | 756.2477 | 755.2397 | −0.7 | C32H38O20 | Forsythoside B | 725.23, 593.21, 325.09, 275.08, 179.03, 161.03, 135.04 | AF, AB |
| 12 | 8.96 | 624.2054 | 623.1979 | −0.2 | C29H36O15 | Isoacteoside | 461.16, 179.03, 161.02 | AF, AB, AL |
| 13 | 9.25 | 520.1945 | 519.1869 | −0.3 | C23H22O13 | Brusatol | 465.18, 447.09, 357.13, 342.11, 209.08, 103.04 | AB |
| 14 | 9.94 | 374.1366 | 373.1292 | −0.0 | C20H22O7 | (−)-Nortrachelogenin | 343.12, 313.11, 207.07, 181.05, 145.03 | AB, AL |
| 15 | 12.42 | 358.1416 | 357.1343 | −0.1 | C19H18O7 | Miroestrol | 342.11, 299.09, 195.07, 151.04, 122.04 | AB |
| 16 | 12.72 | 338.1154 | 337.1081 | −0.0 | C20H18O5 | Wighteone | 295.06, 267.07, 190.06, 175.04, 151.04 | AB |
| 17 | 13.09 | 534.2465 | 533.2393 | −0.1 | C24H22O14 | Bulleyanin | 383.11, 343.16, 341.14, 163.04, 145.03, 121.03 | AL |
| 18 | 13.98 | 436.2097 | 435.2027 | −0.2 | C21H24O10 | Polhovolide | 229.07, 227.11, 185.04, 163.00, 141.02 | AF, AL |
| 19 | 20.65 | 488.3502 | 487.3425 | −0.4 | C30H48O5 | Asiatic acid | 455.32, 409.31 | AF, AB, AL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, T.-K.; Jeong, W.T.; Kim, J.G.; Ji, H.S.; Ahn, M.-A.; Chung, J.-W.; Lim, H.B.; Hyun, T.K. UPLC-ESI-Q-TOF-MS-Based Metabolite Profiling, Antioxidant and Anti-Inflammatory Properties of Different Organ Extracts of Abeliophyllum distichum. Antioxidants 2021, 10, 70. https://doi.org/10.3390/antiox10010070
Yoo T-K, Jeong WT, Kim JG, Ji HS, Ahn M-A, Chung J-W, Lim HB, Hyun TK. UPLC-ESI-Q-TOF-MS-Based Metabolite Profiling, Antioxidant and Anti-Inflammatory Properties of Different Organ Extracts of Abeliophyllum distichum. Antioxidants. 2021; 10(1):70. https://doi.org/10.3390/antiox10010070
Chicago/Turabian StyleYoo, Tong-Kewn, Won Tae Jeong, Jun Gu Kim, Hyo Seong Ji, Min-A Ahn, Jong-Wook Chung, Heung Bin Lim, and Tae Kyung Hyun. 2021. "UPLC-ESI-Q-TOF-MS-Based Metabolite Profiling, Antioxidant and Anti-Inflammatory Properties of Different Organ Extracts of Abeliophyllum distichum" Antioxidants 10, no. 1: 70. https://doi.org/10.3390/antiox10010070