Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Phytochemical and Antioxidant Characterization
2.1.1. HPLC-ESI-qTOF-MS/MS Characterization
Phenolic Acids
Flavonols and Flavone
Flavan-3-Ols
2.1.2. TPC, PAC and Antioxidant Activity
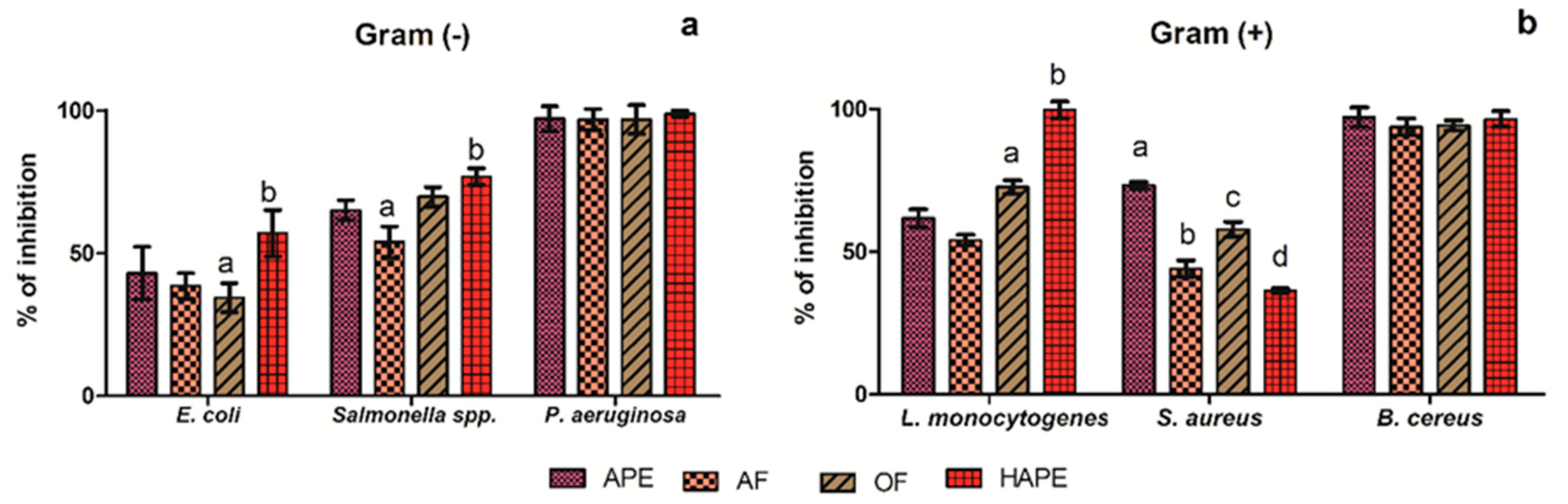
2.2. Antibacterial Activity
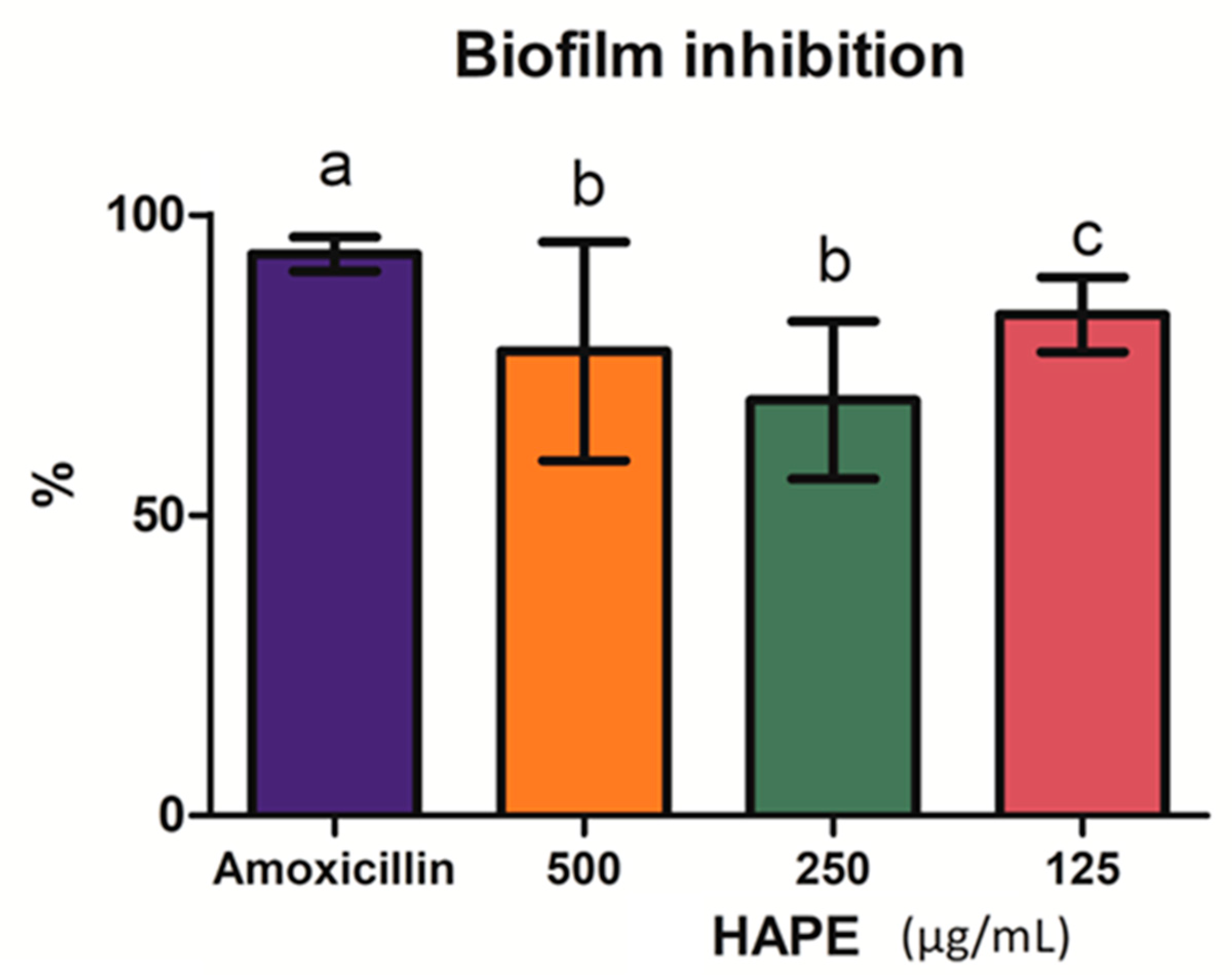
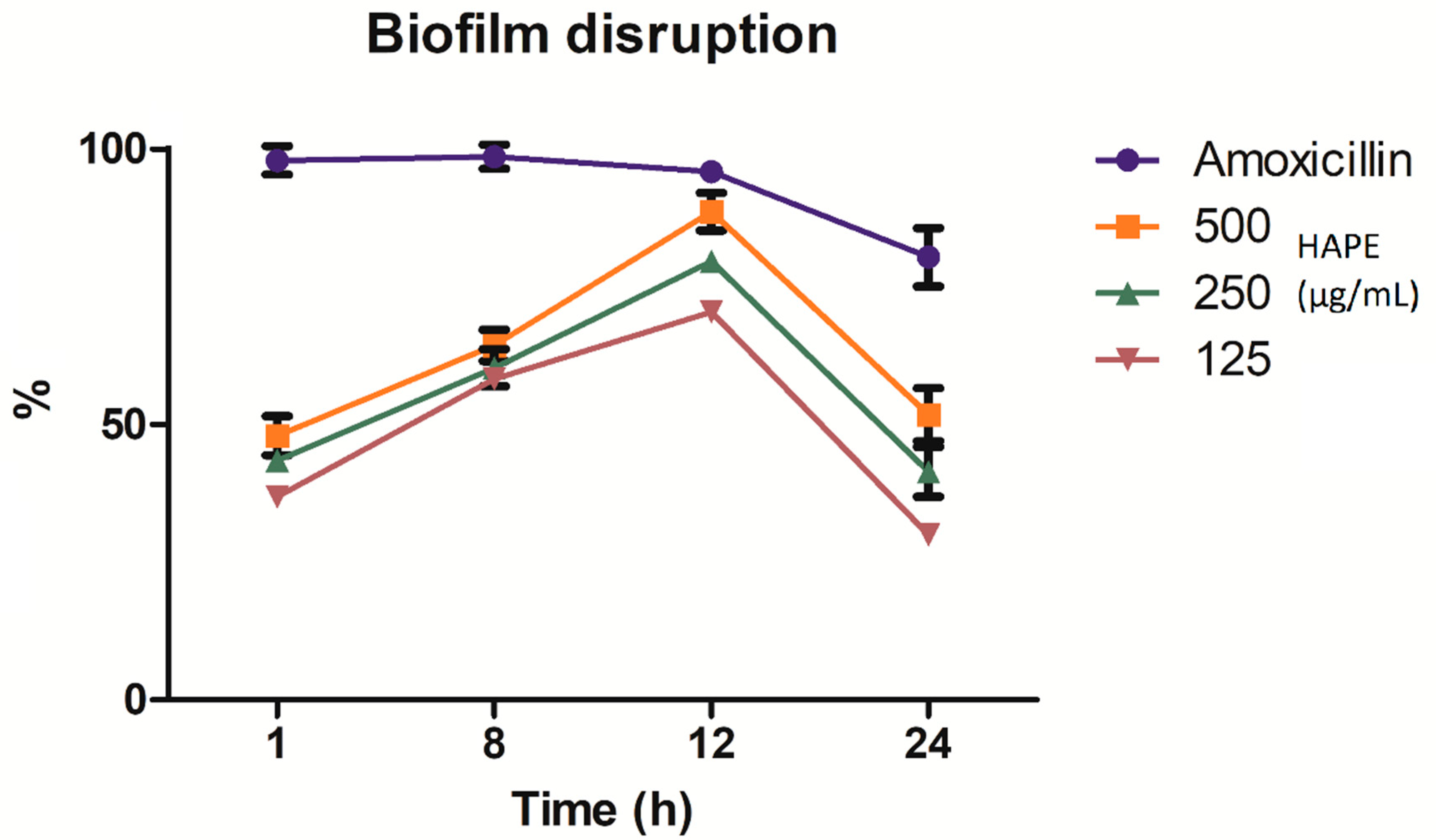
2.3. Listeria Monocytogenes Biofilm Inhibition and Disruption
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents
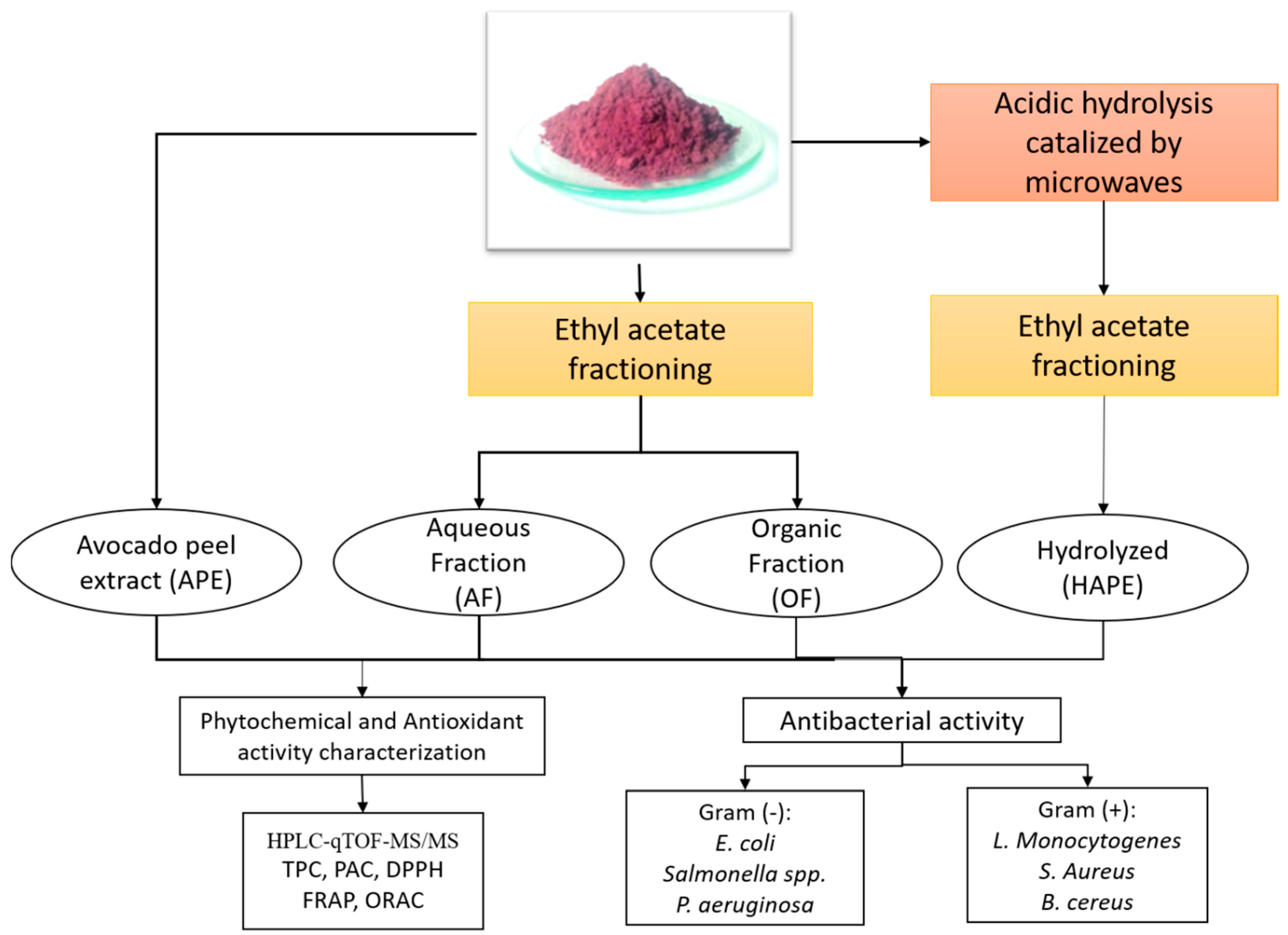
4.2. Avocado Peel Extract Obtainment, Fractionation, and Hydrolysis
4.2.1. Organic and Aqueous Fraction Obtainment
4.2.2. Microwave-assisted Hydrolysis:
4.3. Phytochemical Analysis:
Chromatographical Analysis
4.4. Antioxidant Assays
4.4.1. DPPH Scavenging Capacity
4.4.2. FRAP
4.4.3. ORAC
4.5. Antibacterial Bioassay
4.5.1. Culture Media and Microbial Identification
4.5.2. Broth Microdilution Assay (Planktonic)
4.5.3. L. monocytogenes Biofilm Inhibition and Disruption
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Alarcón-Enos, J.; Céspedes-Acuña, C.; Silva, F.L. Improvement of the polyphenol extraction from avocado peel by assisted ultrasound and microwaves. Food Process Eng. 2019, 1–11. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, J.; Xiao, A.; Liu, L. Antibacterial activity of polyphenols: Structure-activity relationship and influence of hyperglycemic condition. Molecules 2017, 22, 1913. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-del-Río, I.; Fernández, J.; Lombó, F. Plant nutraceuticals as antimicrobial agents in food preservation: Terpenoids, polyphenols and thiols. Int. J. Antimicrob. Agents 2018, 52, 309–315. [Google Scholar] [CrossRef]
- Orhan, D.D.; Özçelik, B.; Özgen, S.; Ergun, F. Antibacterial, antifungal, and antiviral activities of some flavonoids. Microbiol. Res. 2010, 165, 496–504. [Google Scholar] [CrossRef]
- Taylor, P.W. Alternative natural sources for a new generation of antibacterial agents. Int. J. Antimicrob. Agents 2013, 42, 195–201. [Google Scholar] [CrossRef]
- Mostafa, A.A.; Al-Askar, A.A.; Almaary, K.S.; Dawoud, T.M.; Sholkamy, E.N.; Bakri, M.M. Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi J. Biol. Sci. 2018, 25, 361–366. [Google Scholar] [CrossRef]
- Rempe, C.S.; Burris, K.P.; Lenaghan, S.C.; Stewart, C.N. The potential of systems biology to discover antibacterial mechanisms of plant phenolics. Front. Microbiol. 2017, 8, 422. [Google Scholar] [CrossRef] [Green Version]
- Murphy Cowan, M. Plant Products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Kim, K.H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Ramos, L.; Moreno, C.; Zúñiga-Paredes, J.C.; Carlosama-Yepez, M.; Ruales, P. Antimicrobial activity of plant-food by-products: A review focusing on the tropics. Livest. Sci. 2016, 189, 32–49. [Google Scholar] [CrossRef]
- Ribeiro, D.A.; Damasceno, S.S.; Boligon, A.A.; de Menezes, I.R.A.; de Almeida Souza, M.M.; da Costa, J.G.M. Chemical profile and antimicrobial activity of Secondatia floribunda A. DC (Apocynaceae). Asian Pac. J. Trop. Biomed. 2017, 7, 739–749. [Google Scholar] [CrossRef]
- Raymond Chia, T.W.; Dykes, G.A. Antimicrobial activity of crude epicarp and seed extracts from mature avocado fruit (Persea americana) of three cultivars. Pharm. Biol. 2010, 48, 753–756. [Google Scholar] [CrossRef]
- Liu, H.; Zou, T.; Gao, J.M.; Gu, L. Depolymerization of cranberry procyanidins using (+)-catechin, (−)-epicatechin, and (−)-epigallocatechin gallate as chain breakers. Food Chem. 2013, 141, 488–494. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; De Camargo, A.C.; Shahidi, F. Phenolic compounds of pomegranate byproducts (Outer Skin, mesocarp, divider membrane) and their antioxidant activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef]
- Nuutila, A.M.; Kammiovirta, K.; Oksman-Caldentey, K.M. Comparison of methods for the hydrolysis of flavonoids and phenolic acids from onion and spinach for HPLC analysis. Food Chem. 2002, 76, 519–525. [Google Scholar] [CrossRef]
- Escalona-Arranz, J.C.; Pérez-Rosés, R.; Urdaneta-Laffita, I.; Rodríguez-Amado, J.; Morris-Quevedo, H.; Camacho-Pozo, M.I.; Sierra González, G. Role of polyphenols in the antimicrobial activity of ethanol Tamarindus indica L leaves fluid extract. Bol. Latinoam. Caribe Plantas Med. Aromat. 2013, 12, 516–522. [Google Scholar]
- Tian, F.; Li, B.; Ji, B.; Yang, J.; Zhang, G.; Chen, Y.; Luo, Y. Antioxidant and antimicrobial activities of consecutive extracts from Galla chinensis: The polarity affects the bioactivities. Food Chem. 2009, 113, 173–179. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Glibota, N.; Frías, M.P.; Altarejos, J.; Gálvez, A.; Ortega-Morente, E.; Salido, S. Antimicrobial and antibiofilm activities of procyanidins extracted from laurel wood against a selection of foodborne microorganisms. Int. J. Food Sci. Technol. 2017, 52, 679–686. [Google Scholar] [CrossRef]
- Torres, E.; García, A.; Aranda, M.; Saéz, V.; Zúñiga, F.; Alarcón, J.; Avello, M.; Pastene, E. One-step purification of two semi-synthetic epicatechin adducts prepared from avocado peels procyanidins by centrifugal partition chromatography and evaluation of their anti-inflammatory effects on adenocarcinoma gastric cells infected with Helicobacter pylori. J. Chil.Chem. Soc. 2018, 4, 4222–4228. [Google Scholar] [CrossRef] [Green Version]
- Simirgiotis, M.J.; Benites, J.; Areche, C.; Sepulveda, B. Antioxidant capacities and analysis of phenolic compounds in three endemic nolana species by HPLC-PDA-ESI-MS. Molecules 2015, 20, 11490–11507. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Magariño, S.; Revilla, I.; González-Sanjosé, M.L.; Beltrán, S. Various applications of liquid chromatography-mass spectrometry to the analysis of phenolic compounds. J. Chromatogr. A 1999, 847, 75–81. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Herranz-López, M.; Barrajón-Catalán, E.; Arráez-Román, D.; González-Álvarez, I.; Bermejo, M.; Gutiérrez, A.F.; Micol, V.; Segura-Carretero, A. Permeability study of polyphenols derived from a phenolic-enriched Hibiscus sabdariffa extract by UHPLC-ESI-UHR-Qq-TOF-MS. Int. J. Mol. Sci. 2015, 16, 18396–18411. [Google Scholar] [CrossRef]
- Li, S.; Lin, Z.; Jiang, H.; Tong, L.; Wang, H.; Chen, S. Rapid identification and assignation of the active ingredients in Fufang Banbianlian injection using HPLC-DAD-ESI-IT-TOF-MS. J. Chromatogr. Sci. 2016, 54, 1225–1237. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, J.G.; Borrás-Linares, I.; Lozano-Sánchez, J.; Segura-Carretero, A. Comprehensive identification of bioactive compounds of avocado peel by liquid chromatography coupled to ultra-high-definition accurate-mass Q-TOF. Food Chem. 2018, 245, 707–716. [Google Scholar] [CrossRef]
- Jang, G.H.; Kim, H.W.; Lee, M.K.; Jeong, S.Y.; Bak, A.R.; Lee, D.J.; Kim, J.B. Characterization and quantification of flavonoid glycosides in the Prunus genus by UPLC-DAD-QTOF/MS. Saudi J. Biol. Sci. 2018, 25, 1622–1631. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, S.S.; Ibrahim, S.A.; Li, E.H.; Yang, H.; Huang, W. Identification and antioxidant properties of polyphenols in lotus seed epicarp at different ripening stages. Food Chem. 2015, 185, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J.G.; Borrás-Linares, I.; Lozano-Sánchez, J.; Quirantes-Piné, R.; Segura-Carretero, A. Optimization of drying process and pressurized liquid extraction for recovery of bioactive compounds from avocado peel by-product. Electrophoresis 2018, 39, 1908–1916. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Guo, H.; Xu, W.B.; Ge, J.; Li, X.; Alimu, M.; He, D.J. Rapid identification of flavonoid constituents directly from PTP1B inhibitive extract of Raspberry (Rubus idaeus L.) leaves by HPLC-ESI-QTOF-MS-MS. J. Chromatogr. Sci. 2016, 54, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Singh, A.; Kumar, B. Identification and characterization of phenolics and terpenoids from ethanolic extracts of Phyllanthus species by HPLC-ESI-QTOF-MS/MS. J. Pharm. Anal. 2017, 7, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Yuzuak, S.; Ballington, J.; Xie, D.Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 2018, 8, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Šuković, D.; Knežević, B.; Gašić, U.; Sredojević, M.; Ćirić, I.; Todić, S.; Mutić, J.; Tešić, Ž. Phenolic profiles of leaves, grapes and wine of grapevine variety vranac (Vitis vinifera L.) from Montenegro. Foods 2020, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- de la Luz Cádiz-Gurrea, M.; Fernández-Arroyo, S.; Segura-Carretero, A. Pine bark and green tea concentrated extracts: Antioxidant activity and comprehensive characterization of bioactive compounds by HPLC-ESI-QTOF-MS. Int. J. Mol. Sci. 2014, 15, 20382–20402. [Google Scholar] [CrossRef]
- De Souza, L.M.; Cipriani, T.R.; Iacomini, M.; Gorin, P.A.J.; Sassaki, G.L. HPLC/ESI-MS and NMR analysis of flavonoids and tannins in bioactive extract from leaves of Maytenus ilicifolia. J. Pharm. Biomed. Anal. 2008, 47, 59–67. [Google Scholar] [CrossRef]
- Zerbib, M.; Cazals, G.; Enjalbal, C.; Saucier, C. Identification and quantification of flavanol glycosides in vitis vinifera grape seeds and skins during ripening. Molecules 2018, 23, 2745. [Google Scholar] [CrossRef] [Green Version]
- Sobeh, M.; Mahmoud, M.F.; Sabry, O.M.; Adel, R.; Dmirieh, M.; El-Shazly, A.M.; Wink, M. HPLC-PDA-MS/MS characterization of bioactive secondary metabolites from Turraea fischeri bark extract and its antioxidant and hepatoprotective activities in vivo. Molecules 2017, 22, 2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spínola, V.; Pinto, J.; Castilho, P.C. Identification and quantification of phenolic compounds of selected fruits from Madeira Island by HPLC-DAD-ESI-MSn and screening for their antioxidant activity. Food Chem. 2015, 173, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.G.; Costa, H.B.; Ventura, J.A.; Kondratyuk, T.P.; Barroso, M.E.S.; Correia, R.M.; Pimentel, E.F.; Pinto, F.E.; Endringer, D.C.; Romão, W. Chemical profile of mango (Mangifera indica L.) using electrospray ionisation mass spectrometry (ESI-MS). Food Chem. 2016, 204, 37–45. [Google Scholar] [CrossRef]
- Bystrom, L.M.; Lewis, B.A.; Brown, D.L.; Rodriguez, E.; Obendorf, R.L. Characterisation of phenolics by LC-UV/Vis, LC-MS/MS and sugars by GC in Melicoccus bijugatus Jacq. “Montgomery” fruits. Food Chem. 2008, 111, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Fraternale, D.; Ricci, D.; Verardo, G.; Gorassini, A.; Stocchi, V.; Sestili, P. Activity of Vitis vinifera Tendrils extract against phytopathogenic fungi. Nat. Prod. Commun. 2015, 10, 1037–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueñas, M.; Mingo-Chornet, H.; Pérez-Alonso, J.J.; Di Paola-Naranjo, R.; González-Paramás, A.M.; Santos-Buelga, C. Preparation of quercetin glucuronides and characterization by HPLC-DAD-ESI/MS. Eur. Food Res. Technol. 2008, 227, 1069–1076. [Google Scholar] [CrossRef]
- Fasciotti, M.; Alberici, R.M.; Cabral, E.C.; Cunha, V.S.; Silva, P.R.; Daroda, R.; Eberlin, M. Wood Chemotaxonomy via ESI-MS profiles of phytochemical markers: The challenging case of African versus Brazilian Mahogany Woods. Anal. Methods 2015, 7, 8576–8583. [Google Scholar] [CrossRef] [Green Version]
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Antioxidants: Reviewing the chemistry, food applications, legislation and role as preservatives. Trends Food Sci. Technol. 2018, 71, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Hioki, H.; Harada, K.; Kubo, M.; Fukuyama, Y. Antioxidant phenylpropanoid-substituted epicatechins from Trichilia catigua. J. Nat. Prod. 2007, 70, 2010–2013. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Rommel, A.; Wrolstad, R.E. Influence of acid and base hydrolysis on the phenolic composition of red raspberry juice. J. Agric. Food Chem. 1993, 41, 1237–1241. [Google Scholar] [CrossRef]
- Pernin, A.; Dubois-Brissonnet, F.; Roux, S.; Masson, M.; Bosc, V.; Maillard, M.N. Phenolic compounds can delay the oxidation of polyunsaturated fatty acids and the growth of Listeria monocytogenes: Structure-activity relationships. J. Sci. Food Agric. 2018, 98, 5401–5408. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Georgescu, C.; Turcuş, V.; Olah, N.K.; Mathe, E. An overview of natural antimicrobials role in food. Eur. J. Med. Chem. 2018, 143, 922–935. [Google Scholar] [CrossRef]
- Gyawali, R.; Hayek, S.A.; Ibrahim, S.A. Plant extracts as antimicrobials in food products: Mechanisms of action, extraction methods, and applications. In Handbook of Natural Antimicrobials for Food Safety and Quality; Taylor, M., Ed.; Elsevier Ltd.: Raleigh, NC, USA, 2014; pp. 49–68. ISBN 9781782420422. [Google Scholar]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Rivas, C.L.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.M.; Nabavi, S.F.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Saavedra, M.; Simoes, M. Insights on antimicrobial resistance, biofilms and the use of phytochemicals as new antimicrobial agents. Curr. Med. Chem. 2015, 22, 2590–2614. [Google Scholar] [CrossRef] [Green Version]
- Mayer, R.; Stecher, G.; Wuerzner, R.; Silva, R.C.; Sultana, T.; Trojer, L.; Feuerstein, I.; Krieg, C.; Abel, G.; Popp, M.; et al. Proanthocyanidins: Target compounds as antibacterial agents. J. Agric. Food Chem. 2008, 56, 6959–6966. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Patra, A.K. An overview of antimicrobial properties of different classes of phytochemicals. In Dietary Phytochemicals and Microbes; Patra, A.K., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–32. ISBN 978-94-007-3926-0. [Google Scholar]
- Tapiero, H.; Tew, K.; Ba, G.N.; Mathé, G. Polyphenols: Do they play a role in the prevention of human pathologies? Biomed. Pharmacother. 2002, 56, 200–207. [Google Scholar] [CrossRef]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial activity of some flavonoids and organic acids widely distributed in plants. J. Clin. Med. 2019, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Kabir, F.; Katayama, S.; Tanji, N.; Nakamura, S. Antimicrobial effects of chlorogenic acid and related compounds. J. Kor. Soc. Appl. Biol. Chem. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Stojković, D.S.; Živković, J.; Soković, M.; Glamočlija, J.; Ferreira, I.C.F.R.; Janković, T.; Maksimović, Z. Antibacterial activity of Veronica montana L. extract and of protocatechuic acid incorporated in a food system. Food Chem. Toxicol. 2013, 55, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial efficacy of plant phenolic compounds against Salmonella and Escherichia Coli. Food Biosci. 2015, 11, 8–15. [Google Scholar] [CrossRef]
- Proestos, C.; Chorianopoulos, N.; Nychas, G.J.E.; Komaitis, M. RP-HPLC analysis of the phenolic compounds of plant extracts. Investigation of their antioxidant capacity and antimicrobial activity. J. Agric. Food Chem. 2005, 53, 1190–1195. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Burton, S.; Kim, C.; Sismour, E. Phenolic compounds, antioxidant, and antibacterial properties of pomace extracts from four Virginia-grown grape varieties. Food Sci. Nutr. 2016, 4, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Oliveira, D.F.; Silva, G.H.; Figueiredo, H.C.P.; Cavalheiro, A.J.; Carvalho, D.A.; Souza, L.P.; Chalfoun, S.M. Identification of the antimicrobial substances produced by Solanum palinacanthum (Solanaceae). An. Acad. Bras. Cienc. 2008, 80, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, C.; Soulti, K.; Roussis, I.G. Potential antimicrobial activity of red and white wine phenolic extracts against strains of Staphylococcus aureus, Escherichia coli and Candida albicans. Food Technol. Biotechnol. 2005, 43, 41–46. [Google Scholar]
- Melgar, B.; Dias, M.I.; Ciric, A.; Sokovic, M.; Garcia-Castello, E.M.; Rodriguez-Lopez, A.D.; Barros, L.; Ferreira, I.C.R.F. Bioactive characterization of Persea americana Mill. by-products: A rich source of inherent antioxidants. Ind. Crops Prod. 2018, 111, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.K.; Rashid, R.; Fatima, N.; Mahmood, S.; Mir, S.; Khan, S.; Jabeen, N.; Murtaza, G. Pharmacological activities of protocatechuic acid. Acta Pol. Pharm.—Drug Res. 2015, 72, 643–650. [Google Scholar]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinic acid: A spectroscopic study and biological screening for antimicrobial activity. LWT—Food Sci. Technol. 2016, 65, 471–479. [Google Scholar] [CrossRef]
- Pernin, A.; Guillier, L.; Dubois-Brissonnet, F. Inhibitory activity of phenolic acids against Listeria monocytogenes: Deciphering the mechanisms of action using three different models. Food Microbiol. 2019, 80, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Pizzolatti, M.G.; Venson, A.F.; Smânia Júnior, A.; Smânia, E.D.F.A.; Braz-Filho, R. Two epimeric flavalignans from Trichilia catigua (Meliaceae) with antimicrobial activity. Zeitschrift fur Naturforsch.—Sect. C J. Biosci. 2002, 57, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Zajdel, S.M.; Graikou, K.; Sotiroudis, G.; Głowniak, K.; Chinou, I. Two new iridoids from selected Penstemon species—Antimicrobial activity. Nat. Prod. Res. 2013, 27, 2263–2271. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wei, L.; Ge, Z.; Zhu, W.; Li, C. Comparison of the degradation kinetics of A-type and B-type proanthocyanidins dimers as a function of pH and temperature. Eur. Food Res. Technol. 2015, 240, 707–717. [Google Scholar] [CrossRef]
- Wood, J.E.; Senthilmohan, S.T.; Peskin, A.V. Antioxidant activity of procyanidin-containing plant extracts at different pHs. Food Chem. 2002, 77, 155–161. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Tapia-Rodriguez, M.R.; Gonzalez-Aguilar, G.A.; Lopez-Zavala, A.A.; Martinez-Tellez, M.A.; Hernandez-Oñate, M.A.; Ayala-Zavala, J.F. Quercetin reduces adhesion and inhibits biofilm development by Listeria monocytogenes by reducing the amount of extracellular proteins. Food Control 2018, 90, 266–273. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Lizardi-Mendoza, J.; Silva-Espinoza, B.A.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Nazzaro, F.; Fratianni, F.; Ayala-Zavala, J.F. Phenolic extracts from grape stems inhibit Listeria monocytogenes motility and adhesion to food contact surfaces. J. Adhes. Sci. Technol. 2018, 32, 889–907. [Google Scholar] [CrossRef]
- Du, W.; Zhou, M.; Liu, Z.; Chen, Y.; Li, R. Inhibition effects of low concentrations of epigallocatechin gallate on the biofilm formation and hemolytic activity of Listeria monocytogenes. Food Control 2018, 85, 119–126. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Capita, R.; Rodríguez-Jerez, J.J.; Martínez-Suárez, J.V.; Alonso-Calleja, C. Effect of low doses of disinfectants on the biofilm-forming ability of Listeria monocytogenes. Foodborne Pathog. Dis. 2019, 16, 262–268. [Google Scholar] [CrossRef]
- Rhodes, P.L.; Mitchell, J.W.; Wilson, M.W.; Melton, L.D. Antilisterial activity of grape juice and grape extracts derived from Vitis vinifera variety Ribier. Int. J. Food Microbiol. 2006, 107, 281–286. [Google Scholar] [CrossRef]
- Murthy, P.S.; Manonmani, H.K. Physico-chemical, antioxidant and antimicrobial properties of Indian monsooned coffee. Eur. Food Res. Technol. 2009, 229, 645–650. [Google Scholar] [CrossRef]
- Ahmad, J.; Langrish, T.A.G. Optimisation of total phenolic acids extraction from mandarin peels using microwave energy: The importance of the Maillard reaction. J. Food Eng. 2012, 109, 162–174. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Badillo-Muñoz, G.; Céspedes-Acuña, C.; Alarcón-Enos, J. The Relationship between fruit size and phenolic and enzymatic composition of avocado by products (Persea americana Mill.): The importance for biorefinery applications. Horticulturae 2020, 6, 91. [Google Scholar] [CrossRef]
- Franco, S.D.; Ferreira, M.J.P.; de Melo, F.H.M.; Soares, J.P.M.; Torres, L.M.B. In Vitro evaluation of the potential antioxidant of Bidens segetum Mart. ex Colla (Asteraceae) in melanocyte and melanoma cells. J. Biosci. Med. 2019, 07, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | tR (min) | Molecular Formula | [M-H]− (m/z) | Δ ppm | MS2 Fragments m/z (% Base Peak) | Proposed Compound | Sample | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| APE | AF | OF | HAPE | ||||||||
| Phenolic acids | |||||||||||
| 1 | 10.0 | C32H36O18 | 707.1864 | 6.51 | 353 (97), 191 (100) | chlorogenic acid dimer | + | − | − | − | [25] |
| 2 | 10.5 | C7H6O4 | 153.0191 | 5.65 | 153 (32), 109 (100) | protocatechuic acid | − | − | − | + | [26] |
| 3 | 16.4 | C18H22O9 | 381.1185 | 1.29 | 381 (43), 179 (51), 161 (22), 135 (100) | ethyl chlorogenate | − | − | + | − | [27] |
| 4 | 17.3 | C16H18O9 | 353.0864 | −0.87 | 191 (100) | 5-O-caffeoylquinic acid | − | + | + | − | [28] |
| Flavonols | |||||||||||
| 5 | 11.9 | C27H30O17 | 625.1398 | −0.20 | 301 (19), 300 (100), 271 (58), 255 (31), 243 (40), 151 (21) | quercetin diglucoside | + | + | − | − | [29] |
| 6 | 12.7 | C26H28O16 | 595.1285 | 0.57 | 301 (22), 300 (100), 271 (55), 255 (35), 243 (21), 151 (12) | quercetin 3-O-arabinoglucoside | + | + | − | − | [30] |
| 7 | 14.0 | C21H20O12 | 463.0888 | 3.66 | 463 (47), 300 (100), 301 (64), 271 (85) | quercetin 3-O-galactoside | + | − | − | − | [31] |
| 8 | 14.1 | C21H18O13 | 477.0686 | 4.68 | 477 (11), 301 (100), 255 (6), 179 (5) | quercetin 3-O-glucuronide | + | − | − | − | [29] |
| 9 | 14.2 | C21H20O12 | 463.0882 | 2.37 | 463 (45), 301 (97), 300 (100), 271 (83) | quercetin 3-O-glucoside | + | − | − | − | [31] |
| 10 | 14.5 | C27H30O16 | 609.1448 | −0.34 | 300 (100), 271 (66), 255 (40), 243 (36) | quercetin 3-O-rutinoside | + | + | − | − | [29] |
| 11 | 15.2 | C26H28O15 | 579.1316 | −4.91 | 579 (34), 301 (33), 300 (100), 271 (35), 255 (11) | quercetin xylosyl-rhamnoside | + | − | − | − | [32] |
| 12 | 15.6 | .. | 561.0736 | .. | 447 (79), 301 (79), 300 (100), 271 (21), 255 (26) | quercetin derivative | + | − | − | − | [32] |
| 13 | 15.7 | C27H30O15 | 593.1469 | 5.39 | 593 (23), 285 (37), 284 (100), 255 (33) | kaempferol O-hexosyl-deoxyhexose | + | − | − | − | [33] |
| 14 | 20.6 | C15H10O7 | 301.0343 | 0.07 | 301 (31), 178 (34), 151 (100) | Quercetin | − | − | + | + | [29] |
| 15 | 23.4 | C15H10O6 | 285.0400 | 2.23 | 285 (100), 151 (10) | Kaempferol | − | − | + | − | [34] |
| 16 | 28.1 | C21H18O13 | 477.0665 | 2.79 | 301 (39), 179 (52), 151 (100), 121 (56) | quercetin derivative | − | − | + | − | [32] |
| Flavan-3-ols | |||||||||||
| 17 | 9.3 | C30H26O12 | 577.1337 | −0.61 | 407 (8), 289 (9), 245 (10), 161 (23), 151 (27), 125 (100), 109 (21) | procyanidin dimer B a | − | − | + | − | [29,35,36] |
| 18 | 9.5 | C30H26O12 | 577.1372 | 5.45 | 577 (33), 425(37), 407 (66), 289 (100), 243 (17) | procyanidin dimer B a | + | − | − | − | [29,35,36] |
| 19 | 10.3 | C30H26O12 | 577.1366 | 4.41 | 577 (33), 425(60), 407 (52), 289 (100), 245 (16) | procyanidin dimer B a | + | − | − | − | [29,35,36] |
| 20 | 10.6 | C30H26O12 | 577.1352 | 1.99 | 577 (58), 451 (9), 425(84), 407 (66), 289 (100), 245 (17) | procyanidin dimer B a | + | − | − | − | [29,35,36] |
| 21 | 11.3 | C30H26O12 | 577.1339 | −0.26 | 407 (12), 289 (14), 245 (17), 161 (30), 151 (21), 125 (100), 109 (39) | procyanidin dimer B a | − | + | + | − | [29,35,36] |
| 22 | 11.5 | C30H28O12 | 579.1512 | 2.58 | 289 (100), 245 (11), 203 (6) | chalcan-flavan-3-ol dimer b | + | − | − | − | [37,38] |
| 23 | 11.7 | C45H38O18 | 865.2008 | 4.0 | 865 (4), 577 (9), 451 (9), 425 (14), 407 (52), 289 (100), 287 (82), 261 (19), 243 (32) | procyanidin trimer B c | + | − | − | − | [29,35,36] |
| 24 | 11.9 | C30H28O12 | 579.1498 | 0.01 | 289 (22), 245 (27), 203 (27), 151 (36), 137 (48), 125 (69), 123 (80), 109 (100) | chalcan-flavan-3-ol dimer b | − | − | + | − | [37,38] |
| 25 | 12.1 | C30H26O11 | 561.1348 | −7.73 | 561 (15), 407 (6), 289 (100), 245 (8) | (epi)afzelechin–(epi)catechin | + | − | − | − | [39] |
| 26 | 12.2 | C21H24O11 | 451.1231 | −0.86 | 289 (8) 245 (23), 123 (83), 109 (100) | catechin-3-O-glucoside | − | − | + | − | [35] |
| 27 | 12.3 | C15H14O6 | 289.0712 | 1.85 | 289 (23), 245 (18), 203 (32), 161 (42), 123 (80), 109 (100) | (epi)catechin | − | + | − | − | [29,35] |
| 28 | 12.4 | C45H38O18 | 865.1954 | −2.35 | 407 (27), 289 (18), 243 (8), 161 (39), 137 (33), 125(100) | procyanidin trimer B c | − | − | + | − | [29,35,36] |
| 29 | 13.7 | C30H26O12 | 577.1333 | −1.30 | 407 (17), 289 (10), 245 (10), 161 (9), 137 (37), 125 (100), 123 (28), 109 (25) | procyanidin dimer B a | − | − | + | − | [29,35,36] |
| 30 | 14.1 | C39H32O15 | 739.1642 | −2.09 | 289 (21), 245 (12), 177 (100), 161 (31), 137 (34), 125 (35), 109 (60) | procyanidin dimer monoglycoside | − | − | + | − | [40] |
| 31 | 14.4 | C30H26O12 | 577.1343 | 0.43 | 407 (11), 289 (13), 245 (10), 161 (35), 137 (32), 125 (100), 123 (24), 109 (26) | procyanidin dimer B a | − | − | + | − | [29,35,36] |
| 32 | 14.4 | C24H20O9 | 451.1024 | 0.09 | 341 (13), 217 (57), 189 (85), 177 (34), 161 (35), 133 (35), 123 (33), 109 (100) | cinchonain I d | − | − | + | − | [29,41] |
| 33 | 15.2 | C24H20O9 | 451.1031 | 1.64 | 341 (9), 217 (26), 189 (100), 177 (38), 161 (36), 133 (25), 123 (34), 109 (56) | cinchonain I d | − | − | + | − | [29,41] |
| 34 | 15.8 | C24H20O9 | 451.1029 | 1.20 | 407 (10), 341 (16), 255 (34), 217 (25), 189 (65), 177 (54), 161 (27), 123 (30), 109 (100) | cinchonain I d | − | − | + | − | [29,41] |
| 35 | 17.5 | C24H22O16 | 565.0834 | 1.7 | 451 (100), 341 (87), 217 (40), 189 (34) | cinchonain I derivative d | + | − | − | − | [29,41] |
| 36 | 17.8 | C24H20O9 | 451.0997 | −5.89 | 451 (10), 341 (100), 189 (9) | cinchonain I d | + | − | − | − | [29,41] |
| 37 | 18.2 | C24H20O9 | 451.1030 | 1.42 | 341 (13), 217 (40), 189 (100), 177 (41), 161 (34), 151 (17), 133 (27), 123 (33), 109 (85) | cinchonain I d | − | − | + | − | [29,41] |
| Other compounds | |||||||||||
| 38 | 2.5 | .. | 533.1737 | .. | 533 (2), 191 (100) | quinic acid derivative | + | − | − | − | [42] |
| 39 | 2.5 | .. | 383.1180 | .. | 341 (11), 191 (100) | quinic acid derivative | + | − | − | − | [42] |
| 40 | 2.5 | .. | 305.0483 | .. | 191 (100) | quinic acid derivative | + | − | − | − | [42] |
| 41 | 2.7 | .. | 249.0145 | .. | 249 (3), 211 (14), 191 (100), 171 (11), 127 (18), 101 (10) | quinic acid derivative | − | + | − | − | [42] |
| 42 | 3.0 | C12H22O11Cl | 377.0853 | 2.07 | 377 (45), 341 (100), 215 (20) | sucrose | − | + | − | − | [43] |
| 43 | 7.7 | C14H24O12 | 383.1217 | 1.56 | 191 (100) | quinic acid dimer | − | − | + | − | [43] |
| 44 | 9.6 | C7H12O6 | 191.0561 | 5.02 | 191 (100), 171 (10), 127 (18), 111 (8), 109 (18) | quinic acid | − | − | − | + | [29] |
| 45 | 10.1 | .. | 467.0790 | .. | 353(49), 191 (100) | quinic acid derivative | + | − | − | − | [29] |
| 46 | 10.5 | C21H32O10 | 443.1919 | 1.64 | 443 (19), 119 (51), 113 (40), 101 (100) | penstemide | − | + | − | − | [29] |
| 47 | 11.4 | C18H26O10HCOOH | 401.1466 | 2.38 | 401 (100), 269 (28) | benzyl alcohol hexose pentose | + | − | − | − | [44] |
| 48 | 22.5 | C8H12O7 | 219.0505 | 2.61 | 219 (2), 111 (100) | ethyl citrate | − | − | − | − | [45] |
| Sample | TPC | PAC | DPPH | FRAP | ORAC | |
|---|---|---|---|---|---|---|
| Assay | mg GAE/g DE | mg CaE/g DE | µM TEAC/g DE | |||
| APE | 297.42 ± 10.7 b | 2535.43 ± 65.56 a | 900.4 ± 8.8 d | 4954.2 ± 100.8 d | 12541.2 ± 574.8 d | |
| OF | 282.98 ± 12.87 b | 4708.39 ± 177.96 c | 706.4 ± 29.3 c | 7176.5 ± 142.9 c | 15584.6 ± 268.1 c | |
| AF | 244.45 ± 8.65 a | 456.13 ± 9.37 b | 502.8 ± 8.9 ba | 2160.6 ± 137.7 a | 6252.3 ± 267.8 a | |
| HAPE | 252.12 ± 10.79 a | ND | 596.5 ± 9.1 b | 2935.2 ± 124.9 b | 8930.3 ± 409.4 b | |
| Extract | MIC µg/mL | ||||||
|---|---|---|---|---|---|---|---|
| Strain | E. coli | Salmonella spp. | P. aeruginosa | L. monocytogenes | S. aureus | B. cereus | |
| Amoxicillin | ≥10 | ≥1000 | ≥125 | ≥10 | ≥25 | ≥10 | |
| APE | ≥1000 | ≥1000 | ≥500 | ≥750 | ≥750 | ≥500 | |
| AF | ≥1000 | ≥1000 | ≥500 | ≥1000 | ≥1000 | ≥500 | |
| OF | ≥1000 | ≥750 | ≥500 | ≥750 | ≥1000 | ≥500 | |
| HAPE | ≥750 | ≥750 | ≥500 | ≥125 | ≥1000 | ≥500 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillo-Mayol, I.; Casas-Forero, N.; Pastene-Navarrete, E.; Lima Silva, F.; Alarcón-Enos, J. Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity. Antibiotics 2021, 10, 23. https://doi.org/10.3390/antibiotics10010023
Trujillo-Mayol I, Casas-Forero N, Pastene-Navarrete E, Lima Silva F, Alarcón-Enos J. Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity. Antibiotics. 2021; 10(1):23. https://doi.org/10.3390/antibiotics10010023
Chicago/Turabian StyleTrujillo-Mayol, Igor, Nidia Casas-Forero, Edgar Pastene-Navarrete, Fabiana Lima Silva, and Julio Alarcón-Enos. 2021. "Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity" Antibiotics 10, no. 1: 23. https://doi.org/10.3390/antibiotics10010023