



Introduction
The record of millipede body fossils extends from the lower Paleozoic into the Holocene. Fossil millipedes are particularly well known from Carboniferous strata, but two key reviews of fossil millipedes have noted Permian occurrences as being extremely rare (Shear et al., Reference Shear, Seldon and Gall2009, p. 9; Shear and Edgecombe, Reference Shear and Edgecombe2010, p. 186). Permian occurrences are rare, but a number of forms have been described to date. The record is complex, however. Some taxa that were described as being Permian, including those from the Gaskohle of the “Permformation” of Bohemia (Fritsch, Reference Fritsch1901), are now known to be late Carboniferous. Thus, most of the specimens noted in Branson's Bibliographic Index of Permian Invertebrates (Reference Branson1948, p. 984–988) are Carboniferous. Some other supposed Permian millipedes have been determined not to be millipedes. Sterzel (Reference Sterzel1878) determined that the supposed millipede Palaeojulus dyadicus Geinitz, Reference Geinitz1872, described from the ‘Dyas’ (Permian) of Saxony, was half of a fern leaf. And Hannibal et al. (Reference Hannibal, Rindsberg, Lerner, Lucas, Lucas and Zeigler2005) reidentified a supposed Permian millipede from New Mexico as an ichnofossil without any millipede affinities.
There are authentic Permian millipedes, however. These include Arthropleura, whose range extends into the Permian (Kraus, Reference Kraus2005; Schneider et al., Reference Schneider, Lucas, Werneburg, Rößler, Lucas, Schneider and Spielmann2010). Not all authors have agreed, however, on that taxon being a millipede. Several moderate-sized millipede species have been named from the lower Permian of central Europe. These include Archiulus brassii (Dohrn, Reference Dohrn1868) from Lebach-Saar, Xyloiulus permicus (Beurlen, Reference Beurlen1925) from Plauenschen Grund near Dresden, and Pleurojulus steuri Schneider and Werneburg, Reference Schneider and Werneburg1998 from Manebach, Thuringia.
Guthörl (Reference Guthörl1934) redescribed and illustrated A. brassii in his classic paper on the Permo-Carboniferous fossil arthropods from the Saar-Nahe-Pfalz region of Germany. This has resulted in other specimens from Central Europe being either assigned to, or compared with, this species. Three specimens from the lower Permian Rotliegend of the Palatinate have been identified as Archiulus cf. A. brassii (and one millipede specimen left indeterminate) by Heidtke (Reference Heidtke1983). Goretzki (Reference Goretzki1990) compared a millipede from the Permian Oberhöfer Schichten of the Rotliegendes (Permian) in Thuringia with A. brassii. More recently, Poschmann and Schindler (Reference Poschmann and Schindler2004, p. 303, figs. 2G, 3H) have referred millipedes from the Permo-Carboniferous sites of Sitters and Grügelborn to ‘Archiulus’ brassii. Their assignment was based on the presence of fine longitudinal grooves that are continuous across the prozonite and metazonite, and so they assigned this taxon to the Xyloiulidae Cook, 1895. The genus Archiulus Scudder in Dawson, Reference Dawson1868 (p. 496) is also in need of revision. Not all of the specimens referred to this genus are even millipedes (Hannibal, Reference Hannibal2001).
Schneider and Werneburg (Reference Schneider and Werneburg1998, p. 30) noted that X. permicus could belong to the Xyloiulidae or the Nyraniidae Hoffman, Reference Hoffman and Moore1969. Beurlen's (Reference Beurlen1925, p. 183) figures show X. permicus to have pleurotergites whose metazonites are marked with horizontal striae, and his figures show a very close similarity to the diagrammatic figures of Xyloiulus (Xylobius) made by Scudder (in Dawson, Reference Dawson1878, p. 56). On the basis of this comparison, X. permicus does belong to the genus Xyloiulus and the family Xyloiulidae.
Pleurojulus steuri is an elongate millipede with paranota, ozopores, and pleurotergites whose dorsal sides are marked with coarse longitudinal ornamentation. That species, however, does not belong in the genus Pleurojulus Fritsch, Reference Fritsch1901 as what were interpreted as pleurites by Schneider and Werneburg (Reference Schneider and Werneburg1998, figs. 11, 13, 14) are paranota (Wilson and Hannibal, Reference Wilson and Hannibal2005, p. 1106). It is closer to the Carboniferous Hexecontasoma Hannibal, Reference Hannibal, Wytwer and Golovatch2000 in having both distinct paranota and longitudinal ornamentation on its dorsum.
Sharov (Reference Sharov and Rohdendorf1962) referred the late Permian (formerly considered Triassic) form Tomiulus angulatus Martynov, Reference Martynov1936 from southwestern Siberia to the Archiulidae Scudder, Reference Scudder1873. Dzik (Reference Dzik1981) referred this species to the Xyloiulida, but Hannibal and Shcherbakov (Reference Hannibal, Shcherbakov, Dányi, Korsós and Lazányi2019) have suggested a nematophoran affinity instead. They also noted that late Permian millipedes have recently been found in Russia.
There are also several occurrences of unnamed fossil millipedes known from the Permian of Central Europe, the United States, and Africa. These include millipedes from the Permian Tambach Formation of the Rotliegend of the Thuringian Forest of Germany (Martens et al., Reference Martens, Schneider and Walter1981, p. 82, pl. 3, figs. 5–7). Poschmann (Reference Poschmann2007), in a review that discussed various taxa of Permo-Carboniferous millipedes (and other fauna), noted a possible oniscomorph and xyloiulids from the Meisenheim Formation (lower Permian). Poschman et al. (Reference Poschmann, Schindler and Emrich2018) have also identified a specimen from the lower Permian Rotliegend strata of Pfeffelbach in the Saar-Nahe Basin of the Rhineland-Palatinate as a xyloiulid. Xyloiulid millipedes from the lower Permian of Texas have been briefly described by Baird (Reference Baird1958) and noted by Mamay (Reference Mamay1966, p. E11), but neither author illustrated their material from their Texas sites. Fossils identified as millipede-like arthropods have been figured (Reisz and Laurin, Reference Reisz and Laurin1991, fig. 1; Reisz and Scott, Reference Reisz and Scott2002, fig. 1) but not described from the late Permian Cistecephalus Assemblage Zone of South Africa (the locality was originally reported as Triassic). No centipedes have been described from the Permian, but there has been a mention by Wilson (Reference Wilson2006, fig. 4) of a scolopendromorph centipede from the Permian.
It is clear that all of the previously described or noted Permian millipedes, with the possible exception of Arthropleura, are in need of restudy.
The purpose of this paper is to describe a millipede fauna, including three new genera of millipedes, found at the Dolese Quarry, a Permian fissure-fill locality near Richards Spur, Oklahoma, to compare these with other fossil millipedes and to bring to notice other occurrences of fossil myriapods (millipedes and centipedes) preserved in karst deposits. Specimens figured from this fauna are cited with Sam Noble Oklahoma Museum of Natural History numbers (OU). An additional Carboniferous specimen (Hexecontasoma carinatum Hannibal, Reference Hannibal, Wytwer and Golovatch2000) illustrated is cited by its Field Museum of Natural History number (FMNH PE).
Geologic setting
The Richards Spur locality is at the Dolese Quarry, a limestone quarry located near the small town of Richards Spur, Comanche County, in southwest Oklahoma. The locality is north of Fort Sill Army Post, and so Olson (Reference Olson1967, p. 34) referred to this site as the “Richards Spur (Ft. Sill) site.” It is located at 34°46′31.33′′N and 98°24′22.00′′W, which is about 5 miles (8 km) west of the center of the city of Elgin, Oklahoma. The Richards Spur locality has been documented in a number of publications, beginning with Gregory et al. (Reference Gregory, Peabody and Price1956) and continuing to the present (e.g., Olson, Reference Olson1967; May and Cifelli, Reference May and Cifelli1998; Sullivan and Reisz, Reference Sullivan and Reisz1999; Burkhalter and May, Reference Burkhalter, May, Burkhalter, Czaplewski and Lupia2002; Woodhead et al., Reference Woodhead, Reisz, Fox, Drysdale, Hellstrom, Maas, Cheng and Edwards2010; deBraga et al., Reference deBraga, Bevitt, Joseph and Reisz2019). A fissure and cave complex has developed here on the north and south flanks of a small mountain in the Slick Hills in Ordovician bedrock belonging to the Arbuckle Formation. Millipedes described here were found at two localities, about 1 mile apart, at about the same elevation in the quarry.
The fillings of the cave systems at the location are highly mineralized. There are numerous stalactites and typical cave formations including stalagmites, flowstone, and a few cave popcorn and soda-straw structures. Large calcite crystals have been found measuring up to 30 cm (12 inches) long. The fissure and cave infill deposits are considered equivalent with the Arroyo Formation, lower Clear Fork Group, lower Permian (Woodhead et al., Reference Woodhead, Reisz, Fox, Drysdale, Hellstrom, Maas, Cheng and Edwards2010). The formation belongs to the Cisuralian Series (MacDougall et al., Reference MacDougall, Tabor, Woodhead, Daoust and Reisz2017) in International Union of Geological Sciences (IUGS) stratigraphic terms. Tabor and Yapp (Reference Tabor, Yapp, Mora and Surge2005) and MacDougall et al. (Reference MacDougall, Tabor, Woodhead, Daoust and Reisz2017) have described and illustrated aspects of the setting and nature of the fissure-fill material at Richards Spur. They determined that the fill material included sparry calcite, iron sulfides, and goethite. They, and previous studies cited by them, noted that the calcite mineralization occurred or was initialized in the Permian. Woodhead et al. (Reference Woodhead, Reisz, Fox, Drysdale, Hellstrom, Maas, Cheng and Edwards2010) dated a stalagmite from Richards Spur at 289 ± 0.68 Ma and analyzed speleothems from the caves/fissures as bearers of climatic information.
Varying concentrations of disarticulated and articulated skeletal remains are found within the soft clay and in some of the calcite-cemented rocks that have been deposited in the caves/fissures at Richards Spur. With more than 40 described species, Richards Spur is one of the richest known sources of early Permian, upland terrestrial tetrapod skeletal remains in the world. The site is unique in the early Permian because of the preservation of the vertebrate assemblages in undistorted condition. Most are disarticulate elements, however. Many bones have smaller calcite and pyrite crystals on and inside them. Many of the skulls that have been found are complete and cemented together with calcite. The primary tetrapod found at the site (May and Cifelli, Reference May and Cifelli1998) is a reptile, Captorhinus aguti (Cope, Reference Cope1882) that is believed to have been an insectivore and so was a potential predator of millipedes. An acleistorhinid parareptile from Richards Spur has been reported (Modesto et al., Reference Modesto, Scott and Reisz2009) as having indeterminate arthropod cuticle preserved in its oral cavity. That material is composed of an antenna and an elongate, slightly curved “presumed cercal element.” These fragments are relatively large, several millimeters in length. The illustrations of these fragments are not sufficiently detailed to allow assignment to a particular arthropod group (although the elongate element certainly does not belong to a millipede; the antenna is in need of a closer examination). Neither plants nor pollen have been recovered from this site.
Most of the fossils are found in the greenish clay within the caves/fissures. But there are also large pieces of calcite-cemented rock that contain hundreds of miscellaneous bones. The millipedes were recovered by wet-screening. Specimens are associated with calcite. Some specimens are infilled with calcite. Others are composed of a brown-colored material that overlays the calcite.
Materials and methods
Materials
The fossil millipedes were found in the greenish-gray clay infill of the Permian cave/fissure system at the Dolese Richards Spur quarry. This clay also contains bones of fossil vertebrates. The millipedes were discovered during the recovery of the fossil bones by WJM.
Methods
Collected clay was wet-screened through a 30-mesh screen. The screened residue was examined using a Bausch and Lomb BVB-73 dissecting microscope to recover the fossil vertebrate bones. Other than screen washing, no preparation was done on the recovered millipedes. All microphotographs except Figure 1.1 were taken with Scanning Electron Microscopes at the Sam Noble Oklahoma Museum of Natural History and the University of Akron Environmental Scanning Microscope Laboratory. Figure 1.1 was made using a camera through a light microscope. Photoshop was used to obtain optimal lighting levels and contrast, and burning and dodging was used to bring out details, of the photomicrographs.
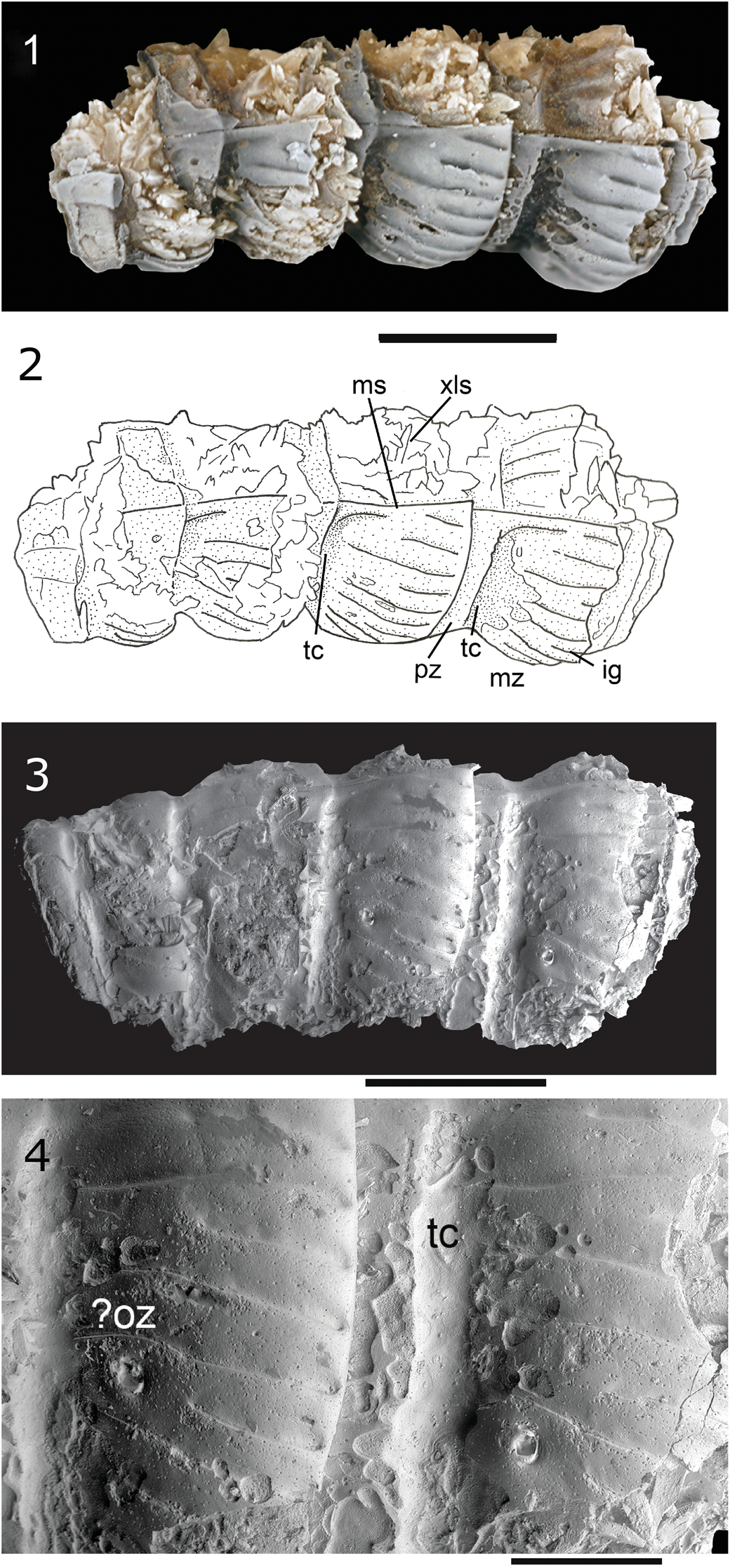
Figure 1. Karstiulus fortsillensis n. gen. n. sp., holotype (OU 12154), from the Fort Sill fissures. Scanning electron microscope (SEM) illustrations except where noted. (1) Dorsal view (light-microscope photograph); (2) interpretive drawing of (1) identifying features noted in text; (3) lateral view of left side; (4) detail of pleurotergites identifying possible ozopore and transverse constriction. ig = intercalary groove; ms = mid-dorsal suture; mz = metazonite; ?oz = possible ozopore; pz = prozonite; tc = transverse constriction; xls = crystals. (1–3) Scale bars = 1 mm; (4) scale bar = 500 μ.
Repositories and institutional abbreviations
The fossil millipedes from the Fort Sill fissures described here are deposited in the Sam Noble Oklahoma Museum of Natural History (OU 12150 through OU 12154 and OU 44526). The holotype of Hexecontasoma carinatum, illustrated in this paper, is deposited in the Field Museum of Natural History (FMNH PE 23487).
Systematic paleontology
Class Diplopoda Blainville in Gervais, Reference Gervais1844
Subclass Chilognatha Latreille, Reference Latreille1802–1803
Infraclass Helminthomorpha Pocock, Reference Pocock1887
Superorder Juliformia Attems, Reference Attems, Kükenthal and Krumbach1926
Order Incertae sedis
Superfamily ?Xyloiuloidea Attems, Reference Attems, Kükenthal and Krumbach1926
Remarks
The Xyloiuloidea, as used here (following Wilson, Reference Wilson2006), include cylindrical millipedes with ozopores and ornamentation consisting of horizontal striae or oblique ridges and grooves. Wilson (Reference Wilson2006) also diagnosed members of this superfamily as having fused tergites, pleurites, and sternites (that is, having their body segments fused into complete rings). Because of incomplete preservation, complete fusion—or lack of complete fusion—of body segments of the new species from the Fort Sill fissures cannot be determined, and so assignment to the superfamily Xyloiuloidea is tentative. The nature of the ring structure of other forms that have been called xyloiulids is also debatable.
The genus Xyloiulus Cook, Reference Cook1895 is in great need of revision. The type species of Xyloiulus, the Carboniferous Xylobius sigillariae Dawson, Reference Dawson1860a, is poorly known but appears to have strongly expressed horizontal ridges on its metazonites. The genus includes species such as the late Carboniferous Xyloiulus moniliformis (Woodward, Reference Woodward1905), which was described backward and is cambaloid-like in some ways (Hannibal, Reference Hannibal, Wytwer and Golovatch2000, p. 31). That species has a single transverse row of prominent longitudinal ridges on its metazonite.
Family incertae sedis
Genus Karstiulus new genus
Type species
Karstiulus fortsillensis new genus new species, by original designation and monotypy.
Diagnosis
As per species.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures. Collected on the north side of what was a small mountain (now quarried away). Cisuralian Series.
Etymology
The name (masculine) is derived from the German word Karst (masculine), which is widely used for carbonate terrain, combined with a common millipede ending for julimorph millipedes.
Remarks
This genus has deeper impressed and more varied striae than Xyloiulus Cook, Reference Cook1895, Nyranius Hoffman, Reference Hoffman1963, Plagiacetus Hoffman, Reference Hoffman1963, Blaniulus Langiaux and Sotty, Reference Langiaux and Sotty1976, Sigmastria Wilson, Reference Wilson2006, and Gaspestria Wilson, Reference Wilson2006. It also differs from the enigmatic genus Anthracojulus Fritsch, Reference Fritsch1901, which appears to have three pleurotergal sclerites (Hoffman, Reference Hoffman and Moore1969, p. R588), and Archiulus, which may not have striae or may have subdued striae.
Karstiulus fortsillensis new genus new species
Figure 1
Holotype
Holotype OU 12154, from the Fort Sill fissures, early Permian, Dolese Quarry, near Richards Spur, Oklahoma.
Diagnosis
Very small millipede with shorter prozonite separated by transverse constriction from longer metazonite, which is also larger in diameter than the prozonite. Midline suture deep. Metazonites with widely spaced, deeply incised, oblique grooves and shorter intercalary grooves. Dorsalmost grooves converge dorsally but do not touch midline suture. Ozopores probably present.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures. Collected on the north side of what was a small mountain (now quarried away). Cisuralian Series.
Description
Very small, cylindrical millipede. Preserved segments up to ~1.2 mm long. Prozonite shorter, poorly preserved, separated by transverse, dorsally shallowing constriction from longer metazonite. Midline suture distinct, deep. Metazonites inflated, marked by widely spaced, deeply incised, oblique grooves, the longer of which originate close to the anterior of the metazonite and terminate as a depression very close to the posterior of the metazonite. Shorter, intercalary grooves originate at about mid-metazonite. Dorsalmost grooves converge toward midline suture but do not connect at midline. Raised area between grooves generally convex dorsally. Probable rimmed (?)ozopores (Fig. 1.4) located close to anterior of metazonite. Groove posterior of (?)ozopore truncated, that is, an intercalary groove. Cuticle finely and irregularly punctate.
Etymology
The trivial species name is an adjective denoting the Fort Sill fissures in which the specimen was found. The gender is masculine.
Material
OU 12154, consisting of a somewhat laterally compressed specimen composed of cuticle over a crystallized internal filling. Specimen is composed of three better-preserved segments and two partial segments. The specimen is from a new fossil-producing pocket in the greenish-colored clay from the fissure-and-cave complex. This pocket has also produced five to six new genera of amphibians and reptiles that were unknown or had been represented only by isolated bones from the Richards Spur site.
Remarks
Much of the cuticle of the specimen is broken away at the midline suture. The metazonites of this species may have flared outward posteriad (see position of fourth segment in Fig. 1.1, 1.2). This, along with the general form of the pleurotergites, the oblique ornamentation of the metazonites, and the placement of the ozopores, would have given this millipede an appearance that was generally similar to the modern julid Unciger, which is in the extant tribe Brachyiulini. On the basis of its general similarities with julimorphs, this species was probably elongate. Xyloiulidans are known as far back as the Lower Devonian (Wilson, Reference Wilson2006). The specimen is, in general form and in the probable presence of ozopores, similar to the specimens of xyloiulidans illustrated in classic papers (Scudder, Reference Scudder1873, Reference Scudder1878) as well as the xyloiulid specimen figured by Baird et al. (Reference Baird, Shabica, Anderson and Richardson1985, fig. 5.3).
The ornamentation of the Richards Spur specimen, however, differs from that of those other specimens of xyloiulidans. The grooves of the Richards Spur specimen differ from those of the Xyloiulus as diagnosed by Hoffman (Reference Hoffman1963) in that the grooves of the Richards Spur specimen are not always continuous across a segment. In addition, at least some specimens referred to Xyloiulus (Hoffman, Reference Hoffman1963, text-fig. 2) have longitudinal grooves that meet at the midline, in contrast to the grooves of the Richards Spur specimen, which do not meet at the midline.
The distinct midline suture of the new form also differentiates it from most other xyloiulidans, including Gaspestria Wilson, Reference Wilson2006. The new form differs from other genera including Tomiulus angulatus and Sigmastria Wilson, Reference Wilson2006 in both tergite form and ornamentation.
The supposed xyloiulidan that Baird (Reference Baird1958) briefly described from Texas is coeval in age and proximity to the Richards Spur specimen. That specimen is from the Arroyo Formation of the Clear Fork Group (lower Permian, Leonardian) at Coffee Creek, Baylor County, Texas. Baird (Reference Baird1958) referred this specimen to the genus Xylobius (now Xyloiulus). The specimen is in need of a more detailed redescription.
Superorder and Order incertae sedis
Remarks
Oklahomasoma (Figs. 2, 3) shares a resemblance with a number of modern orders but most closely resembles the Carboniferous Hexecontasoma carinatum Hannibal, Reference Hannibal, Wytwer and Golovatch2000 (Fig. 4) from Mazon Creek, Illinois, which Hannibal (Reference Hannibal, Wytwer and Golovatch2000) did not place in an order. He suggested, however, that Hexecontasoma may be a callipodidan. Wilson (Reference Wilson2006, p. 643) placed Hexecontasoma as a basal nematophoran and agreed that it may be a callipodidan nematophoran. Shear and Edgecombe (Reference Shear and Edgecombe2010, p. 186) subsequently noted that Hexecontasoma lacks the derived characters that define the callipodidans, but also agreed (p. 182) that it may be a nematophoran. More recently, Edgecombe (Reference Edgecombe and Minelli2015, p. 347) noted that Hexecontasoma could not be reliably placed in an order due to the lack of critical characters. This is even more true for Oklahomasoma as the ventral side of O. fortsillensis is poorly preserved.

Figure 2. Oklahomasoma richardsspurense n. gen. n. sp., OU 44526, SEM microphotographs except for drawing. (1) Dorsal view; (2) close-up of dorsum; (3) interpretive drawing of (2) identifying features noted in text; (4) ventral view, slightly tilted to better see left side; (5) close-up of ventral side of two segments and one coxal segment. dn = dorsal node; cox = coxa; ln = lateral node; ma = mineralized area where cuticle is missing; ms = mid-dorsal suture; mz = metazonite; pn = paranota; pz = prozonite. (1, 4) Scale bars = 1 mm; (2, 3) scale bars = 500 μ; (5) scale bar = 200 μ.

Figure 3. Oklahomasoma richardspurense n. gen. n. sp., OU 44526. (1) SEM micrograph of right side; anterior is to the right; (2) interpretive drawing identifying parts noted in text. bt = broken-off terminations of paranota; ln = lateral node; lr = longitudinal ridges (elevated loaf-like features); ma = mineralized area where cuticle is missing; ms = mid-dorsal suture; mz = metazonite; pn = paranota; pz = prozonite; tc = transverse constriction. Scale bar = 1 mm.
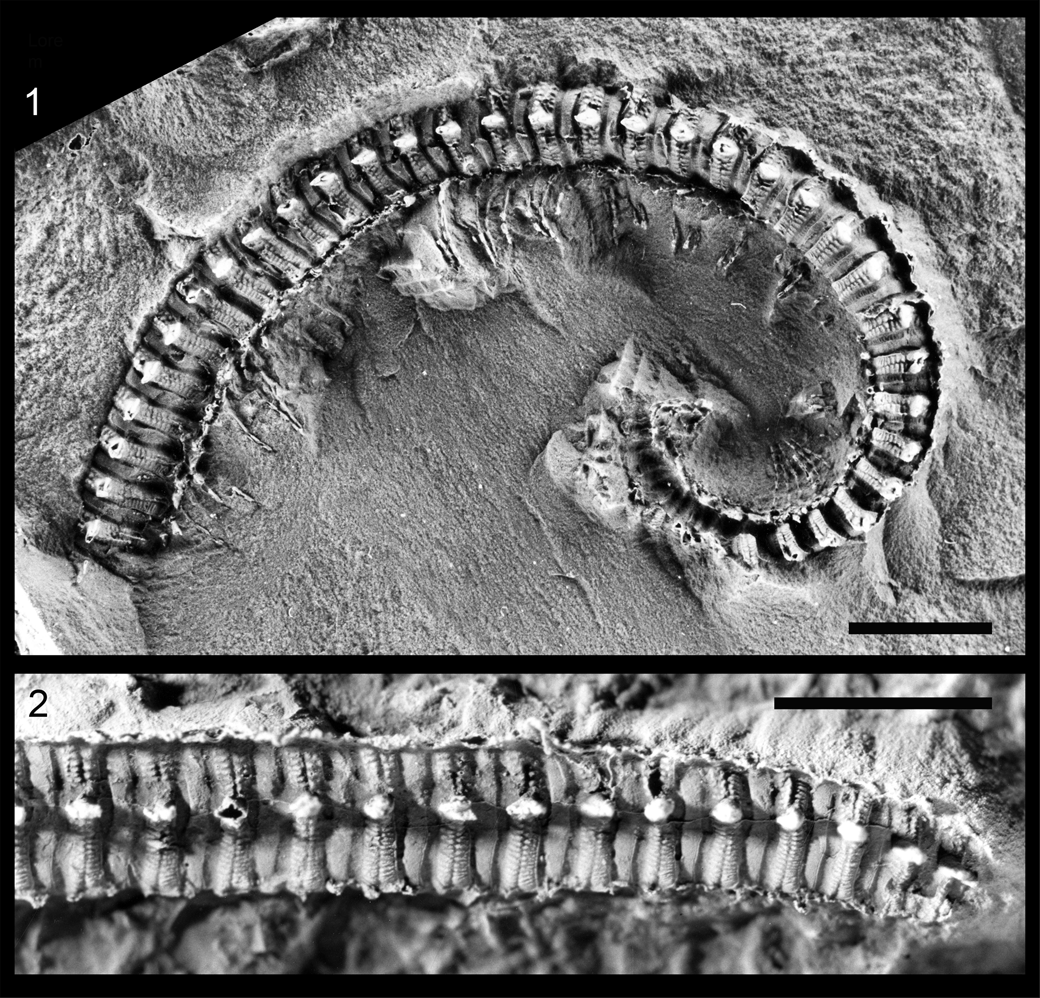
Figure 4. Hexecontasoma carinatum Hannibal, Reference Hannibal, Wytwer and Golovatch2000, whitened latex molds of holotype, FMNH PE 23487, Carboniferous of Mazon Creek, Illinois. (1) View of midpoint and anterior of right side (from Hannibal, Reference Hannibal, Wytwer and Golovatch2000); (2) posterior of left side. (1) Scale bar = 5 mm; (2) scale bar = 4 mm.
Order incertae sedis
Family Oklahomasomatidae new family
Type genus
Oklahomasoma new genus.
Diagnosis
As for species.
Genus Oklahomasoma new genus
Type species
Oklahomasoma richardsspurense new genus new species, by original designation and monotypy.
Diagnosis
As for species.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures, a fissure on the south side of what was a small mountain (now quarried away). Cisuralian Series.
Etymology
The name combines the name of the state in which the fossil was found and the Greek word for body (soma, neuter), a common ending for millipede genera.
Remarks
The millipede is similar in a number of respects to Hexecontasoma from the upper Carboniferous of Mazon Creek, Illinois (Hannibal, Reference Hannibal2001; Fig. 4) in that both genera have prominent subrectangular paranota and two rows of transverse longitudinal ornamentation on their metazonites. Oklahomasoma differs from Hexecontasoma, however, in having different dorsal ornamentation and a different number of notches on the lateral edges of its paranota. The new form, like Hexecontasoma, bears some resemblance to extant cambaloid millipedes (see discussion in Hannibal, Reference Hannibal, Wytwer and Golovatch2000). The new form, however, has differently formed prozonites, strikingly different dorsal morphology, and differently shaped paranota from Hexecontasoma and extant cambaloids.
Holotype
OU 44526, composed of eight articulated anterior segments, the anteriormost of which is only partially preserved. From the Fort Sill fissures, early Permian, Dolese Quarry, near Richards Spur, Oklahoma.
Diagnosis
Very small millipede with prominent mid-dorsal suture. Metazonite with pair of wide, subovoid, dorsal nodes located toward anterior. Metazonite below paranota with two transverse rows of irregularly bulbous, loaflike, longitudinal ornamentation. Paranota prominent, subrectangular, with a prominent notch located near anterior of lateral edge.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures, a fissure on the south side of what was a small mountain (now quarried away). Cisuralian Series. This specimen was found in a fissure that also produced the captorhinid reptile Captorhinus magnus Kissel, Dilkes, and Reisz, Reference Kissel, Dilkes and Reisz2002.
Description
Very small, cylindrical millipede. Length of holotype and sole specimen (composed of part of eight anterior segments) about 7.2 mm. Preserved segments ~1 mm long. Mid-dorsal suture present. Prozonite relatively smooth to subtly striate, smaller in diameter than, and so transversely constricted compared with, metazonite. Metazonite with two anterior, laterally elongate, subovoid dorsal nodes that extend laterally from close to midline across most of metazonite. Indication of a less prominent lateral node located at intersection of posterolateral side of metazonite with the paranota. Paranota broad, elongate, subrectangular, with prominent notch located near anterior of lateral edge. Paranota horizontal or angled slightly upward from horizontal, subtriangular in transverse cross section. Ozopores not apparent. Metazonite below paranota with two transverse rows of longitudinal, bulbous, loaflike ridges. Sternites narrow. Coxal segments prominent (other leg segments not preserved).
Etymology
The species epithet refers to the town nearest to the locality at which this taxon was found.
Remarks
It is possible that the specimen was in a molting stage, accounting for the prominence of the mid-dorsal suture and separation along it. Both Oklahomasoma richardsspurense and Hexecontasoma carinatum (Fig. 4) bear some similarity to “Pleurojulus” steuri from the lower Permian of Germany (Schneider and Werneburg, Reference Schneider and Werneburg1998). “Pleurojulus” steuri differs from O. richardsspurense in having longitudinal ridges and ozopores on its dorsum. “Pleurojulus” steuri does resemble H. carinatum in having dorsal longitudinal ornamentation and paranota; “P.” steuri may belong to the Hexecontasomatidae. Additional millipedes with paranota have been described from the Carboniferous (see Wilson et al., Reference Wilson, Daeschler and Desbiens2005; Ross et al., Reference Ross, Edgecombe, Clark, Bennett, Carrió, Contreras Izquierdo and Crighton2018), but those have different paranotal configurations and been assigned to the Archipolypoda.
Like a number of other millipedes found in more recent caves and fissures (see Remarks that follow), O. richardsspurense is preserved in three dimensions.
Superorder, order, and family incertae sedis
Genus Dolesea new genus
Type species
Dolesea subtila new genus new species, by original designation and monotypy.
Diagnosis
As per species.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures, a fissure on the north side of what was a small mountain (now quarried away). Cisuralian Series.
Etymology
The name is derived from the Dolese Quarry in which the specimen was found. Its gender as used here is feminine.
Remarks
The general form of the pleurotergites of this genus resembles that of the unornamented pleurojulid Pleurojulus levis Fritsch, Reference Fritsch1901 (see Wilson and Hannibal, Reference Wilson and Hannibal2005). All Pleurojulus species, however, have distinctly separate pleurites. The relatively simple body structure of Dolesea also resembles that of the Triassic millipede genus Hannibaliulus Shear, Selden, and Gall, Reference Shear, Seldon and Gall2009, but Hannibaliulus has metazonites with prominent transverse depressions and a simpler termination of its pleurotergites.
Holotype and paratype
Holotype, OU 12152, mostly compressed, consisting of three segments. Another specimen (paratype, OU 12153) consisting of four more or less complete and one partial segment.

Figure 5. Dolesea subtila n. gen. n. sp. (1, 2) SEM microphotograph (1) and interpretive drawing (2) of anterior of holotype (OU 12152) mostly compressed, in dorsal view; specimen is tilted somewhat to the right; (3) detail of (1) showing cytoscutes in greater detail; (4–6) SEM microphotographs (4, 5) and interpretive drawing (6) of ventrolateral side of paratype (OU 12153). br = branching ridges; cp = cuticular pores; cr = crack; ms = mid-dorsal suture; mz = metazonite; oz = ozopore; pz = prozonite; tc = transverse constriction; vr = ventral ridges. (2) Scale bar = 500 μm; (3) scale bar = 200 μm; (4, 6) scale bars = approximately 100 μm.

Figure 6. Dolesea subtila n. gen. n. sp., SEM microphotograph showing detail of inwardly rolled ventral edges of both sides of millipede. Upper set is that of the left side; lower set is that of right side. Scale bar = 100 μm. br = branching ridge; vr = ventral ridges.
Diagnosis
Cylindrical millipedes with shorter prozonites and longer metazonites. Midline suture present. Prozonites separated from metazonites by a ridge or ridge and narrow trough. Ozopores on most segments. Ornamentation subtle. Pleurotergites with ridges of various configurations near terminus.
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures, a fissure on the north side of what was a small mountain (now quarried away). Cisuralian Series. These specimens were found in a fissure that also produced the captorhinid reptile Captorhinus magnus.
Description
Cylindrical millipede. Prozonites short. Ornamentation of prozonite consisting of pronounced scalelike cytoscutes (cuticular platelets). Prozonites distinctly separated from metazonites by a transverse constriction, the anterior of which distinctly drops in elevation and the posterior of which may or may not be distinctly expressed. Ornamentation of metazonite similar to that of prozonite, but scalelike forms less well defined. Ozopores located closer to anterior than posterior of metazonite. Small cuticular pits (possible sites of setal attachment) scattered in metazonite. Pits also in irregular rows at anterior of metazonites. Midline suture distinct. Pleurotergites with anteriorly bifurcating ridge located near lateral edge. Lateral edge with up to five or more closely spaced ventral ridges giving corrugated appearance.
Etymology
The trivial species name is from the Latin adjective subtilis, referring to its fine, delicate ornamentation.
Remarks
Presence of a midline suture is inferred on the basis of the overlap of the anteriormost segment preserved, where the segment is broken longitudinally. This longitudinal break is more or less straight. The bifurcating ridge on the pleurotergites of this species is similar to that seen in illustrations of Chordeuma sylvestre Koch, Reference Koch1847 (Blower, Reference Blower1985, fig. 26A). Polygonal cuticular structures (cytoscutes) seen on this taxon are known for species in various millipede orders, including the Julida, Polydesmida, and the Spirostreptida (e.g., Eisenbeis and Wichard, Reference Eisenbeis and Wichard1987, pl. 63; Adis et al., Reference Adis, Caoduro, Messner, Enghoff and Enghoff1997, figs. 4–6; Golovatch et al., Reference Golovatch, Geoffroy, Mauriès and Van den Spiegel2009). The cytoscutes of D. subtila preserve very fine microstructure, consisting of randomly oriented fine (?)pits located ~1 μ apart from each other.
This specimen bears an overall resemblance to Xyloiulus platti (Baldwin, Reference Baldwin1911). Xyloiulus platti, however, is not a xyloiulid as it lacks longitudinal striae (see illustrations in Baldwin, Reference Baldwin1911; see also Edgecombe, Reference Edgecombe and Minelli2015, fig. 14.2A). Both taxa have short prozonites and longer metazonites and have pleurotergites that lack prominent striae and grooves. The lateral edge of the pleurotergite of X. platti, however, ends with a rim (rebordering of Shear et al., Reference Shear, Seldon and Gall2009) but lacks the anteriorly bifurcating ridge located near lateral edge of the pleurotergite of D. subtila.
The ventral ridges of D. subtila are partly preserved in an inwardly rolled position, with the ridges of the right and left sides converging (Fig. 6). This must have been due to the disarticulation or disintegration of the sternal area, allowing for the distal edges of the pleurotergite to deform and coil inward.
Superorder, order, family, genus, and species incertae sedis
Figure 7
Occurrence
Dolese Quarry, near Richards Spur, Comanche County, southwest Oklahoma, from the Fort Sill fissures, a fissure on the south side of what was a small mountain (now quarried away). Cisuralian Series.

Figure 7. Left side of coiled helminthomorph millipede, OU 12150. Scale bar = 1 mm.
Descriptions
Two elongate millipedes, OU 12150 (Fig. 7) and OU 12151, with relatively unornamented pleurotergites. These two specimens, one preserved in a coiled position and the other outstretched, are briefly described for completeness.
OU 12150 (Fig. 7) is a coiled specimen consisting of at least 20 midbody segments. Prozonite shorter than metazonite. Segments about 0.8–1 mm wide. Pleurotergites wide; with rim on dorsal, anterodorsal, and posterodorsal sides of pleurite; the left side has microgranular ornamentation, but the other side is coarsely pitted. Prozonite shorter than metazonite. Sternites separate, narrow. OU 12151 (not illustrated) is an outstretched specimen 0.6 mm wide, consisting of about seven segments. It is so highly weathered that pleurotergal details are obscured. It appears to have narrow subtriangular sternites.
Remarks
These two additional specimens from the Fort Sill fissures are less well preserved than those we have assigned to new species. It was also difficult photographing key features of these specimens with the scanning electron microscopes available.
Discussion
Occurrence of fossil myriapods in karst
The Richards Spur locality is the site of one of the oldest occurrences of fossil myriapods (millipedes and centipedes) in karst deposits, including limestone cave deposits and fissure fills. However, there are many additional occurrences in karstic deposits. It is clear from the number of examples in Table 1 that the occurrence of millipedes in karst is one of the most important modes of preservation of millipedes in the fossil record, especially when the relative scarcity of millipedes in the fossil record is considered. Most reports of such occurrences in karstic deposits have not cited other such occurrences in karst, so the relatively large number of karstic records of fossil millipedes has not been properly noted previously, and very little has been done comparing various occurrences with each other.
Table 1. Examples of fossil myriapods found in karst deposits along with references describing these myriapods and/or the localities
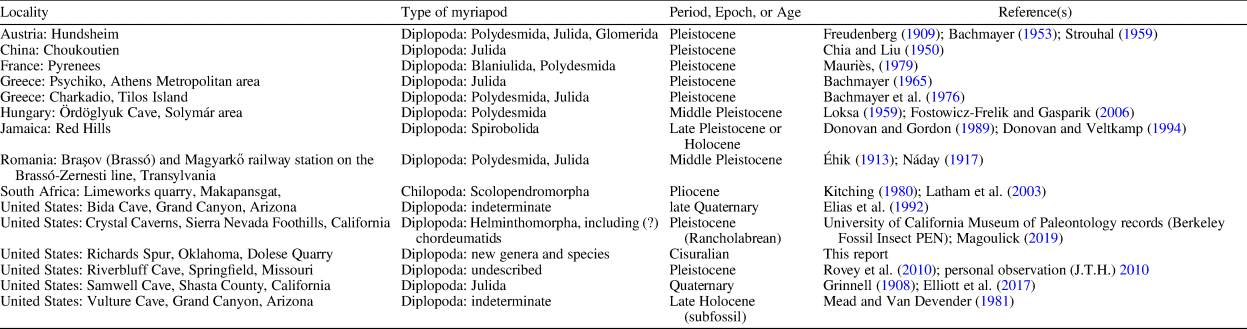
By contrast, occurrences of millipedes in amber and concretion faunas are widely known. A good number of diplopod taxa have been described from Carboniferous concretions from Europe (including Great Britain and France) and North America (the Mazon Creek deposits of Illinois, USA) and from Cretaceous, Cenozoic, and other amber faunas worldwide (notably amber from Myanmar [e.g., Stoev et al., Reference Stoev, Moritz and Wesener2019], the Baltic area [e.g., Haug et al., Reference Haug, Haug, Neumann, Sombke and Hörnig2018], and the Dominican Republic). Other occurrences of fossil millipedes have been described from coal (e.g., the Gaskohle of the Czech Republic), siltstone, limestone, calcitic onyx, asphalt, and other types of rock.
The majority of the karstic sites are Pleistocene, reflecting the greater accessibility of cave entrances that have continued to be open since the Pleistocene. Some additional Pleistocene caves have been open for exploration only after accidental discovery during large-scale construction projects, the caves having been buried. These include Riverbluff Cave in Missouri, discovered while building a highway (Rovey et al., Reference Rovey, Forir, Balco, Gaunt, Evans and Aber2010). The Red Hills cave in Jamaica was probably exposed similarly (Donovan and Veltkamp, Reference Donovan and Veltkamp1994). Geologically older sites are rarer, doubtlessly due to their relative inaccessibility. The Fort Sill fissures, comprising filled-in karst that had been deeply buried, represent a deposit that would have never been found had the site not been quarried for carbonate rock. Likewise, a fossil chilopod is preserved in calcite at the Limeworks site of Makapanscat, South Africa (Kitching, Reference Kitching1980), where deposits containing fossils were first exposed by lime working.
Millipedes have relatively robust, calcium-impregnated exoskeletons. Various authors have independently commented on the preservation potential of millipedes in limestone crevices and caves. Náday (Reference Náday1917, p. 16) noted that thick but easily broken exoskeletons of myriapods are more easily preserved in cave fillings. Bachmayer (Reference Bachmayer1953, p. 27), Bachmayer et al. (1976, p. 133, 138), and Mauriès (Reference Mauriès1979) briefly noted the calcification/encrustation with calcite of fossil millipedes preserved in karstic, calcium-rich environments. Bachmayer (Reference Bachmayer1953, fig. 16) and Bachmayer et al. (1976, pl. 5) also illustrated calcite encrusted specimens in comparison with similar Recent taxa. Donovan and Veltkamp (Reference Donovan and Veltkamp1994) noted the preservation potential of millipedes in carbonate environments in their discussion of fossil millipedes found in a cave in Jamaica and convincingly postulated early diagenetic cementation of the material they studied. They also hypothesized that the millipedes at their Red Hills site were washed into the cave during tropical storms or hurricanes, drowned, and then coated with calcite, their calcareous skeleton acting as a depositional substrate in calcium-carbonate-rich water just before or during burial in sediment (Donovan and Veltkamp, Reference Donovan and Veltkamp1994, fig. 7). They hypothesized that the set of conditions leading to such preservation was more likely to occur in the tropics. This may have been the case with the Richards Spur fauna as the site was in the tropics in the Permian. Tabor and Yapp (Reference Tabor, Yapp, Mora and Surge2005, p. 68) indicated that the Richards Spur area would have been warm and dry in the Permian, with some seasonality that could have included monsoons. Monsoonal conditions have been increasingly accepted for Richards Spur (deBraga et al., Reference deBraga, Bevitt, Joseph and Reisz2019), so it is possible that the Richards Spur site would have experienced the conditions envisioned by Donovan and Veltkamp (Reference Donovan and Veltkamp1994). However, this is not entirely consistent with Pleistocene occurrences (Table 1) in other parts of the world. Náday (Reference Náday1917), for example, noted that the Braşov, Transylvania, millipede fauna was deposited when the Braşov area was experiencing a Mediterranean (not tropical) climate, and specimens have been found in Austria (Bachmayer, Reference Bachmayer1953).
Macdougal et al. (2017, fig. 3) have published a set of scenarios for a vertebrate at the Fort Hill fissures locality that is also applicable to millipedes but different from the scenario envisioned by Donovan and Veltkamp (Reference Donovan and Veltkamp1994, fig. 7). Macdougal et al. (Reference MacDougall, Tabor, Woodhead, Daoust and Reisz2017) included an animal dying and then disarticulating before being washed into a karst pit, an animal being washed into a pit after death but before disarticulation, and an animal falling in a pit and dying there.
To these scenarios can be added the possibility that millipedes found preserved in karst actually lived in the caves and fissures, or were occasional visitors to these sites, in which they are found. Millipedes are a common element of the modern cave fauna. Some caves have yielded both fossil and modern millipedes that might be conspecific. Mauriès (Reference Mauriès1979), for example, identified fossil millipede material in the genus Typhloblaniulus in the Grotte de la Carriére, in which the extant Typhloblaniulus troglobius (Latzel in Gadeau de Kerville and Latzel, Reference Gadeau de Kerville and Latzel1886) was also found (see also discussions by Elliott et al., Reference Elliott, Reddell, Rudolph, Graening, Briggs, Ubick, Aalbu, Krejca and Taylor2017).
In addition, various other animals may have transported them to these karstic environments. Mead and Van Devender (Reference Mead and Van Devender1981), for example, have reported millipedes in the feces of the extant ringtail Bassariscus astutus (Lichtenstein, Reference Lichtenstein1830) at California's Vulture Cave. In fact, diplopods were the most common arthropods found in dung (it is also possible that the millipedes were feeding on the dung) and loose within the analyzed cave sediment at Vulture Cave.
Many extant millipedes are known to favor carbonate terrains, and such terrains are likely to have karst features. Extant millipedes may be found living both outside and inside caves and fissure openings. Numerous millipedes are troglobitic (Hopkin and Read, Reference Hopkin and Read1992, p. 181). Living millipedes and millipede carcasses can also be carried into caves by streams and can be transported into fissures by sheet wash and floods. Millipedes can also be transported into caves by other animals.
Because of the variety of possibilities that could lead to a millipede being preserved as a fossil in a karst setting, any original habitat of the Richards Spur specimens cannot be determined, except that these millipedes were preserved in a carbonate terrain. A similar type of preservation of a Miocene millipede is known in lacustrine carbonates (Arp, Reference Arp1995). In that case, like at Richards Spur, microstructures are preserved.
Preservation
Some millipedes (e.g., Fig. 1) found at Richards Spur are preserved three dimensionally. This was facilitated by preservation in karstic environments that allowed for preservation before complete flattening. Such three-dimensionality is well known for specimens preserved within concretions, but it is also common in specimens preserved in karst (see illustrations in Náday, Reference Náday1917; Bachmayer, Reference Bachmayer1953, Reference Bachmayer1965; Loksa, Reference Loksa1959; Bachmayer et al., Reference Bachmayer, Symeonidis, Seemann and Zapfe1976; Donovan and Veltkamp, Reference Donovan and Veltkamp1994). Details preserved (Figs. 2–4), even when the specimens are not fully three-dimensionally preserved, can be extraordinary, including views of cytoscutes seen in some of the Richards Spur material (Figs. 2, 3). Various authors writing about fossil millipede faunas (Table 1) have commented on this mode of preservation. (Detailed, three-dimensional preservation is also found in some special nonkarst environments such as that described by Duncan et al., Reference Duncan, Briggs and Archer1998.) In many cases of preservation in karst, however, the specimens are also relatively fragile. Specimens may also be coated in part with fine calcitic material that strengthens the fossil but, because it adheres to the fossil cuticle, also obscures features. Specimens may have easily recognizable calcite crystals (e.g., Fig. 1) adhering to their inside or outside.
Co-occurrence with vertebrates and gastropods
The occurrence of millipedes at a site best known for vertebrates such as Richards Spur is unusual neither in general nor in the case of karstic localities. Co-occurrence of fossil millipedes and vertebrates has been known since the dawn of paleomyriapodological investigations, with the work of Dawson (Reference Dawson1860a, p. 36–37), who described the Carboniferous millipede Xyloiulus sigillariae (Dawson, Reference Dawson1860a) as being found along with the tetrapod Dendrepeton (and land snails) within a single hollow tree stump found in a sequence of shales, siltstone, and coal at Joggins, Nova Scotia. Dawson (Reference Dawson1860b, Reference Dawson1862) also elaborated on such co-occurrences, noting that segments of Xyloiulus were found in coprolites at Joggins. Xyloiulus permicus was found associated with the sphenacodon Pantelosaurus (Beurlen, Reference Beurlen1925, p. 182).
Fossil myriapods found in cave and fissure sites (Table 1) are typically dominated by vertebrate remains (e.g., Donovan and Veltkamp, Reference Donovan and Veltkamp1994). Several cave and fissure sites at which Cenozoic (mostly Pleistocene) fossil myriapods have been found are best known for vertebrates. These include the Austrian site of Hundsheim (Freudenberg, Reference Freudenberg1909; Bachmayer, Reference Bachmayer1953), the Greek site of Charkadio on Tilos Island (Bachmayer et al., Reference Bachmayer, Symeonidis, Seemann and Zapfe1976), and Braşov, Transylvania (Náday, Reference Náday1917).
Interestingly, Reisz and Laurin (Reference Reisz and Laurin1991) hypothesized that a close co-occurrence of what are probably millipedes and procolophonid reptiles in a Permian (nonkarstic) fossil deposit in South Africa was due to the scavenging of reptiles by the millipede-like arthropods. This is a possibility as the probable millipedes are closely associated with the procolophonids, as seen in Reisz and Laurin (Reference Reisz and Laurin1991, fig. 1) and in Reisz and Scott (Reference Reisz and Scott2002, fig. 1) (both figures of the same material). Abdala et al. (Reference Abdala, Cisneros and Smith2006) have proposed an alternative scenario, postulating that a similar co-occurrence of tetrapods and millipedes in Lower Triassic rocks of South Africa was due to reptiles and millipedes sharing a burrow. When such previous accounts of finds of millipedes associated with reptiles are taken into account, it seems that the co-occurrences of millipedes and vertebrates may simply be due to the occurrence of both taxa in similar habitats.
Although gastropods have not been found in the Fort Sill fissures, the co-occurrence of millipedes and gastropods has been documented from other localities, karstic and nonkarstic, beginning with the well-known association of the Carboniferous X. sigillariae with land snails documented by Dawson (Reference Dawson1860a, b). In a report of taxa found at Hundsheim, Freudenberg (Reference Freudenberg1909, p. 199–200) specifically noted two groups of fossil invertebrates: gastropods and millipedes. Gastropods are a prominent part of the fossil-invertebrate fauna found at Braşov (Brassó) (Soós, Reference Soós1916) and the Jamaican site of Red Hills (Donovan and Veltkamp, Reference Donovan and Veltkamp1994), as are millipedes. This co-occurrence must be based on the relative abundances of snails and millipedes, the mineralogy of their shells and exoskeletons, and the high-calcium content of the material filling fissures and caves in karst terrain. No gastropods have been found to date at Richards Spur, however.
Millipedes are not the only arthropods found in karst. Moldovan et al. (Reference Moldovan, Mihevc, Miko, Constantin, Meleg, Petculescu and Bosák2011), for example, reported oribatid mites, sparse cladocerids, and insects from a Pliocene–Pleistocene cave in Slovenia, and some deposits noted in Table 1, for example Makapansgat, contain a variety of insects and other animals, most famously hominids. Arthropods, other than the indeterminate elements noted under Geologic setting, and millipedes have yet to be identified at Richards Spur, however.
Comparison of early Permian and Carboniferous millipedes
The three new genera described here, plus occurrences at other sites noted in the introduction, hint at the existence of a diverse millipede fauna of small millipedes in the Permian, although not as diverse as the fauna described from the Carboniferous. As the preceding individual comparisons indicate, the species of the Richards Spur fauna do bear some similarity to forms from the Carboniferous. The overall paleogeographical location of the major Carboniferous millipede-bearing sites of Mazon Creek and Nýřany was similar to that of the Richards Spur locality in the early Permian. The three sites were all located in the low latitudes that would have had considerable precipitation (Wilson and Hannibal, Reference Wilson and Hannibal2005, fig. 1; Woodhead et al., Reference Woodhead, Reisz, Fox, Drysdale, Hellstrom, Maas, Cheng and Edwards2010, p. 456). On the basis of the vertebrate fauna, Richards Spur has been interpreted as more of an upland fauna (Woodhead et al., Reference Woodhead, Reisz, Fox, Drysdale, Hellstrom, Maas, Cheng and Edwards2010), however, and this might have made a difference in habitats, with the Richards Spur locality being somewhat more xeric than most Pennsylvanian coal-forest faunas that include millipedes. To date, the Permian worldwide lacks any representative of the euphoberiidan archipolypods, the prominent spined millipedes of the Carboniferous Euroamerican coal belt (Hannibal, Reference Hannibal, Shabica and Hay1997).
Conclusions
The millipede fauna described here is important for a number of reasons: (1) it is composed of new Permian millipede taxa; (2) it includes morphologically diverse taxa; (3) details of the millipede exoskeletons are preserved, some in three dimensions; and (4) it is one of the oldest examples of a myriapod fauna preserved in karst. Karst faunas should be ranked with concretion faunas, cannel coals, and amber faunas as a major source of fossil myriapods. In very general form (that is, in being composed of juliform millipeds, forms that resemble nematophorans, and millipedes with prominent paranota), the millipedes from Richards Spur resemble those from Pleistocene cave faunas, but the taxa at Richards Spur appear to be more closely related to Carboniferous forms than to Pleistocene or extant taxa.
Acknowledgments
We thank the Dolese Richards Spur Quarry for allowing and facilitating quarrying of this material. Environmental Scanning Electron Micrographs were taken by R. Burkhalter at the Sam Noble Oklahoma Museum of Natural History and L. Park at the University of Akron's Environmental Scanning Electron Microscope Laboratory, funded by NSF grant EAR-MRI 0320898. Funds in support of this article were supplied by J. B. Kitto. The Museum and Institute of Zoology of the Polish Academy of Sciences gave permission to reprint Fig. 4.1. For access to specimens and/or photos and literature, we thank The Paleontology Institute, Moscow; J. Almond, Cape Town, South Africa; J. Cundiff, Harvard Museum of Comparative Anatomy; C. Neumann, Museum für Naturkunde, Berlin; the Magyar Állami Földtani Intézet, Budapest; and M. Forir and others at the Missouri Institute of Natural Science, Springfield, Missouri. We thank W. Wasman, Cleveland Museum of Natural History; A. Lerner, New Mexico Museum of Natural History & Science; P. Rachebouf, Université Claude Bernard-Lyon; U.H.J. Heidtke, Pfalzmuseum für Naturkunde (Pollichia-Museum), Bad Dürkheim, Germany; and G. Edgecombe, Natural History Museum, London, for help with references. We also thank H. Enghoff, University of Copenhagen, Museum of Comparative Anatomy, for comments on the microsculpture of the specimens. D. Dunn and A. Sanford, Cleveland Museum of Natural History, and K. Farago proofread versions of this paper. S.I. Golovatch, Institute for Problems of Ecology & Evolution, Russian Academy of Sciences, Moscow, kindly provided a number of helpful corrections to a much earlier draft of this manuscript. Finally, we thank S. Golovatch and an anonymous reviewer for their helpful peer reviews of this manuscript,









 Open access
Open access