Can Differences in Symbiont Transmission Mode Explain the Abundance and Distribution of Fungus-Growing Termites in West Africa?
 Judith Korb
Judith Korb Simon Kolotchèlèma Silué1,2
Simon Kolotchèlèma Silué1,2  N'golo Abdoulaye Koné
N'golo Abdoulaye Koné- 1Evolutionary Biology and Ecology, Institute of Biology i (Zoology), University of Freiburg, Freiburg im Breisgau, Germany
- 2Natural Sciences Department (UFR-SN), Nangui Abrogoua University, Abidjan, Côte d'Ivoire
Fungus-growing termites (Isoptera: Macrotermitinae) dominate African savannah ecosystems where they play important roles in ecosystem functioning. Their ecological dominance in these ecosystems has been attributed to living in an ectosymbiosis with fungi of the genus Termitomyces (Lyophyllaceae). Evolutionary theory predicts that the transmission mode of a symbiont determines cooperation and conflict between host and symbiont with vertical transmission (co-transmission of host and symbiont offspring to the next generation) leading to less conflict than horizontal transmission (symbionts are acquired by the host from the environment). Thus, one can hypothesize associations with vertical transmission to be ecological more successful than those with horizontal transmission. We tested this by analyzing whether there is an association between transmission mode and fungus-growing termite species abundance and distribution in West-African savannah and forest ecosystems. We used data from a total of 78 study sites comprising protected National Parks as well as anthropogenically disturbed ecosystems, covering Benin, Côte d'Ivoire, and Togo. Our results showed that, in contrast to expectation, species with horizontal symbiont transmission were more common. We encountered more often species with horizontal than vertical transmission. This result might be due to the fact that only five out of the 25 identified fungus-growing termite species had vertical transmission. Yet, species with horizontal transmission also had higher relative abundances within study sites than those with vertical transmission. Thus, transmission mode is unlikely to explain abundance differences between fungus-growing termite species.
Introduction
Fungus-growing termites (Isoptera: Macrotermitinae) thrive in African savannah ecosystems where they often make up more than 50% of termites' species diversity and the large majority of termites (e.g., Hausberger et al., 2011; Hausberger and Korb, 2015, 2016; Schyra et al., 2019a; and reference therein) and where they play pivotal roles in ecosystem functioning (e.g., Bignell and Eggleton, 2000; and references therein). Within the ecological food web, they are main macro-detrivores, essential for decomposition of dead plant material, and are prey for a wealth of animals from invertebrates (such as ants) to vertebrates (such as birds and mammals). Some species are even specialized on termites as prey, like the ant Megaponera analis or aardwarks (Orycteropus afer), aardwolfs (Proteles cristata) or pangolins. Termites also provide important ecosystem services as their activity enhances soil quality, for instance by increasing soil fertility and improving water infiltration rates (Lee and Wood, 1971; Holt and Lepage, 2000; and references therein). Additionally, the “termitaria” of mound-building species present new habitats for plants and animals which are partly obligatorily dependent on them (e.g., Darlington, 1989; Erpenbach et al., 2017). Thus, fungus-growing termites foster biodiversity and are essential for its maintenance, especially in savannah ecosystems.
The ecological dominance of fungus-growing termites in savannah ecosystems, which are characterized by dry seasons, has been attributed to their ectosymbiosis with Termitomyces fungi (reviewed in Korb, 2020). The termites cultivate these fungal symbionts within their nests where they provide constant optimal conditions for fungal growth, including high humidity. This allows the fungal symbiont to be active all year round and relatively independent of environmental conditions, while other decomposing fungi and microbes are mainly restricted to the rainy season. Thus, Termitomyces symbionts flourish together with their termite hosts which benefit from reliable and more efficient plant decomposition compared to other termites (e.g., Rouland et al., 1991; Rouland-Lefèvre, 2000; Poulsen, 2015; da Costa et al., 2019). Yet, species of fungus-growing termites differ in distribution and abundance (e.g., Pomeroy et al., 1991; Bagine et al., 1994; Lepage and Darlington, 2000; Schyra et al., 2019a), and thus potentially in ecological importance. In this study, we tested whether the fungal transmission mode (i.e., how the symbiont is transmitted to the next generation) may contribute to explaining the ecological success of different fungus-growing termite species.
Evolutionary theory predicts that the transmission mode of a symbiont can be an important factor in explaining the degree of cooperation and conflict in symbiotic associations, and thus their “success” (Frank, 1994, 1996, 1997, 1998; Foster and Wenseleers, 2006; Leeks et al., 2019). Associations in which symbiont offspring are strictly co-transmitted with host offspring to the next generation (vertical transmission) are predicted to have less conflict than associations in which the symbiont can disperse independently of their host and infect new hosts (horizontal transmission). In the former, the fitness interests of host and symbiont are largely aligned and symbionts can mainly increase their fitness by enhancing host fitness. This leads to less potential conflict between host and symbiont. Additionally, evolutionary theory predicts that vertical transmission of symbionts, especially if uniparental (i.e., via a single host parent), further reduces potential conflict because it results in increased genetic homogeneity (i.e., increased relatedness) of symbionts within hosts. In the case of strict uniparental, vertical transmission, symbionts are expected to be clonally propagated. Thus, conflict among different symbiont strains within a host is prevented which would, for instance, reduce availability of resources to hosts.
In fungus-growing ants, fungal symbiont transmission is generally vertical and per default uniparental as colonies are founded by a single female (the queen) (Nobre et al., 2011c). This contrasts with fungus-growing termites (e.g., Korb and Aanen, 2003; Nobre et al., 2011c). Most association are characterized by horizontal transmission with Termitomyces fungi producing fruiting bodies (mushrooms) which release spores that are picked up by foraging termite workers (Johnson et al., 1981; Koné et al., 2011). The few exceptions are Macrotermes bellicosus and Microtermes species (Grasse and Noirot, 1955; Johnson, 1981; Johnson et al., 1981; Korb and Aanen, 2003; Nobre et al., 2011a, c). Strikingly, in both taxa vertical transmission is uniparental (in M. bellicosus the winged males carry fungal spores, in Microtermes the winged females) although the default option would be biparental as colonies are founded by a male (king) and a female (queen). This variation in transmission mode in fungus-growing termites offers the unique possibility to test for an effect of transmission mode on the ecological success of fungus-growing termites.
For a long time, research on fungus-growing termites was hampered by taxonomical problems. Except for a few, often iconic mound building species such as Macrotermes bellicosus, species could not be identified reliably using morphological means (Korb et al., 2019). Especially for the potentially species-rich genera Microtermes and Odontotermes this is very difficult (Korb et al., 2019). Morphological species identification is generally difficult in termites as species-specific markers are rare (e.g., Hausberger et al., 2011; Korb et al., 2019; and references therein), but it is even more problematic in Macrotermitinae. They have simplified guts which prevents using gut traits for identification, as successfully done in some other termites (Sands, 1998). The problem of species identification has been overcome by applying genetic markers (Korb et al., 2019 and references therein), so that we now have a good species list for West African termites (Hausberger et al., 2011; Schyra et al., 2019a). This allows us to do comparative analyses and test for an effect of transmission mode on the ecological success of fungus-growing termite species across West African savannah and forest regions.
Materials and Methods
Data Sets
We used data on termite communities that we had published for disturbed and protected savannah ecosystems in Benin (Hausberger et al., 2011; Hausberger and Korb, 2015, 2016) and Togo (Schyra and Korb, 2019; Schyra et al., 2019a,b). They were supplemented with data from Côte d'Ivoire collected in protected as well as disturbed areas belonging to different phytogeographical zones (i.e., from evergreen forest to Guinean and Sudano-Guinean savannah). In line with the published studies (Hausberger and Korb, 2016), we had three disturbance regimes: (1) protected (i.e., well protected National Parks); (2) intermediate anthropogenic disturbance (e.g., old fallows, National Parks with cattle grazing, low protection status); (3) strong anthropogenic disturbance (e.g., young fallows, plantations, fields).
In all studies, sampling was done using the standardized belt transect protocol, first developed for sampling termites in forests (Jones and Eggleton, 2000) and then adapted to savannahs (Hausberger et al., 2011). In short, a transect is established of 50–100 m lengths and 2 m width which is subdivided into 5 m × 2 m sections. Within each section, a thorough systematic search for termites of dead plant material on the ground, on and in trees and mounds is done for a standardized period of time by a trained person. This search is supplemented by soil scrapes measuring around 15 cm × 15 cm × 10 cm to specifically collect termites in the soil. Whenever we found/encountered termites during the search within a transect section, we collected a few specimens in a vial (5–10 individuals; mainly soldiers). Then we continued searching within the section and when we encountered termites again they were placed in a separate vial. The number of resulting vials for a study site (i.e., the sum over all transect sections for all replicate within a site) was used as encounter rate. This is used as a surrogate of species abundance (Davies, 2002). Samples were stored in 99% ethanol for species identification and subsequent analysis.
Some details in sampling effort differed between the published data and the newly included data for Côte d'Ivoire. Transect lengths in Togo and Benin was 50 m (i.e., 10 sections) while it was 100 m (i.e., 20 sections) for Côte d'Ivoire. The time that one person spent searching for termites in a section was 15 min for Togo and Benin, while it was four persons each 10 min for Ivorian savannah sites and 15 min for Ivorian forest sites. The number of soil scrapes was eight in the former studies and 12 in Côte d'Ivoire. To characterize a study site, one transect was done in Benin, three transects in Togo, and five in Côte d'Ivoire. Sampling effort differed to adjust for varying termite abundance between countries/regions and habitats. Varying sampling effort, however, did not bias this study because we do not compare termite composition across study sites. This study is a global analysis that uses all data combined to compare the encounter rates and relative abundance (see below) of fungus-growing termites with horizontal vs. vertical transmission across all regions.
Species Identification
All Macrotermitinae species were identified morphologically using keys for African termites by Bouillon and Mathot (1965, 1966, 1971) and Webb (1961), illustrations by Josens (1972) and descriptions by Grassé (1984, 1986). Additionally, samples were genetically identified as described elsewhere (Hausberger et al., 2011; Schyra and Korb, 2019), using particularly a fragment of cytochrome oxidase II (COII) as “barcode,” which turned out to be especially suitable for West African termite species identification. Species names were assigned consistently across all studies.
Statistical Analyses
We analyzed the total number of encounters of all fungus-growing termite species as well as their relative abundance among the Macrotermitinae within a study site to compare species with horizontal vs. vertical transmission. As additional variables, we tested for effects of “habitat” (savannah vs. forest) and “disturbance” (protected, intermediate disturbance, strong disturbance) on the occurrence of fungus-growing termites. All analyses were done with IBM SPSS 26. All tests were two-tailed with an alpha-value of 0.05 which was adjusted for multiple testing using the false discovery rate (FDR) approach (Benjamini and Hochberg, 1995), whenever necessary. For more details on specific tests, see Results.
Results
Total Encounters Across All Sites
In total, we analyzed the occurrence of Macrotermitinae in 78 study sites distributed across three countries. We found 25 species, belonging to seven genera (Allodontotermes, Ancistrotermes, Macrotermes, Microtermes, Odontotermes, Protermes, and Pseudacanthotermes), in 3,812 encounters with Macrotermitinae (Figure 1; for more details see Supplementary Table 1). Five species (Macrotermes bellicosus, Microtermes subhyalinus, Microtermes osborni, Microtermes sp. 2, Microtermes sp. 4) had vertical symbiont transmission (Figure 1, Supplementary Table 1).
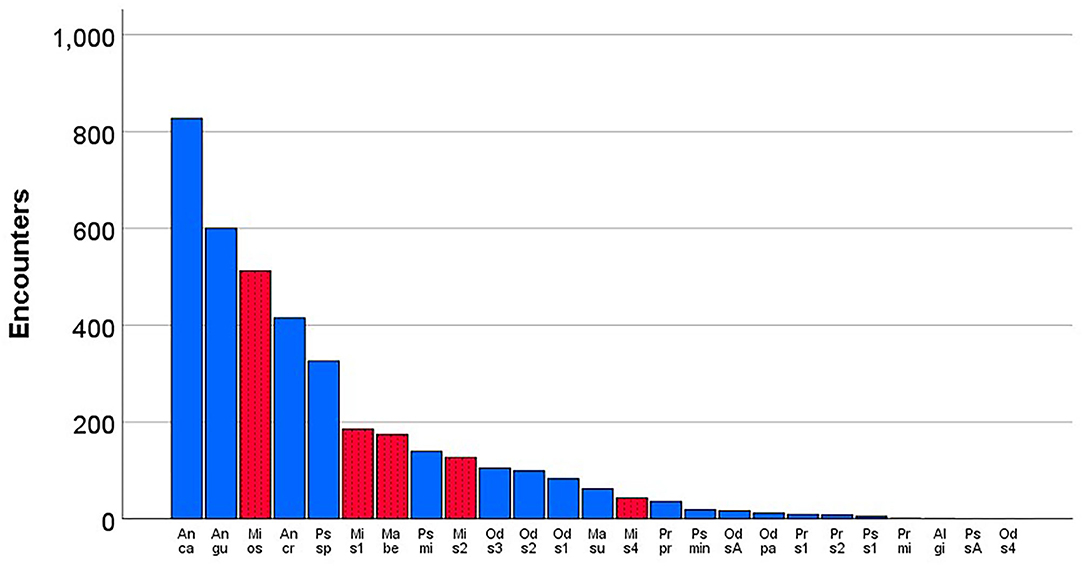
Figure 1. Total number of encounters of different fungus-growing termite species studied in 78 savannah and forest study sites from Benin, Côte d'Ivoire, and Togo. Species with horizontal transmission are indicated in blue and those with vertical transmission in red (with dots). Al gi, Allodontotermes giffardi; An ca, Ancistrotermes cavithorax; An cr, Ancistrotermes crucifer; An gu, Ancistrotermes guineensis; Ma be, Macrotermes bellicosus; Ma su, Macrotermes subhyalinus; Mi os, Microtermes osborni; Mi su, Microtermes subhyalinus; Mi s2, Microtermes sp. 2; Mi s4, Microtermes sp. 4; Od sA, Odontotermes sp.A; Od pa, Odontotermes sp. (aff. pauperans); Od s1, Odontotermes sp. 1; Od s2, Odontotermes sp. 2; Od s3, Odontotermes sp. 3; Od s4, Odontotermes sp. 4; Pr mi, Protermes minutus; Pr pr, Protermes prorepens; Pr s1, Protermes sp. 1; Pr s2, Protermes sp. 2; Ps mi, Pseudacanthotermes militaris; Ps min, Pseudacanthotermes minor; Ps sp, Pseudacanthotermes spiniger; Ps sA, Pseudacanthotermes sp.A; Ps s1, Pseudacanthotermes sp. 1.
The three most common species (with more than 500 encounters each) were Ancistrotermes cavithorax, Ancistrotermes guineensis and Microtermes osborni with 21.7, 15.7, and 13.4% of all samples, respectively (Figure 1). The former two species are supposed to transmit their fungal symbiont horizontally, while the last species has supposed vertical transmission (Grasse and Noirot, 1955; Johnson, 1981; Johnson et al., 1981; Korb and Aanen, 2003). Two species were encountered only once: Pseudacanthotermes sp. A and Odontotermes sp. 4 (Figure 1, Supplementary Table 1).
In contrast to expectation, when a termite was encountered it was significantly less likely to be a species with vertical than with horizontal transmission of the fungal symbiont (chi21 = 785.13, p < 0.001). It was almost three times more likely to encounter a species with horizontal (73%) than vertical (27%) transmission (Figure 2). Encounter frequencies were significantly affected by habitat (chi21 = 111.22, p < 0.001) (Figure 2). In forests, encounters of species with horizontal transmission were more than four times more likely than encounters with vertically transmitting species (80.6 vs. 19.4%), while it was only around twice as likely that an encounter in the savannah was with a horizontally vs. vertically transmitting species (65.4 vs. 34.6%).
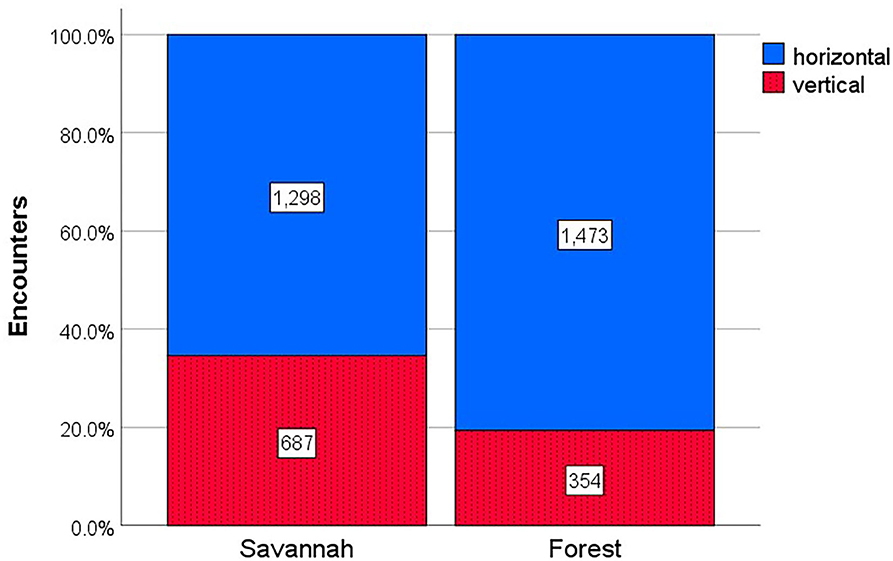
Figure 2. Encounter frequencies of fungus-growing termites with different symbiont transmission modes across habitats. Shown is the proportion of encounters (with absolute numbers inside the bars) of termites with horizontal (blue) and vertical transmission (red, dotted) in 78 savannah and forest study sites from Benin, Côte d'Ivoire, and Togo.
More detailed analyses within habitats showed that disturbance had an effect on the likelihood to encounter species with horizontal vs. vertical transmission in forests (chi22 = 28.55, p < 0.001), but not the savannah (chi22 = 0.46, p = 0.796) (Figure 3). In the forests, the proportion of species with horizontal transmission increased on disturbed sites while it did not change in the savannah (Figure 3).
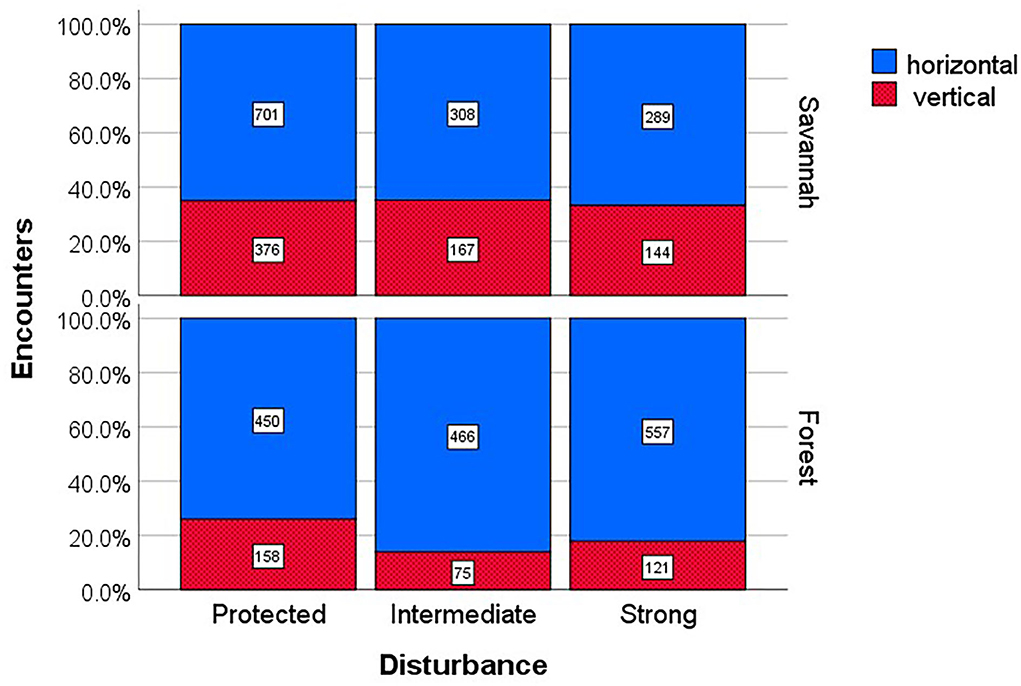
Figure 3. Encounter frequencies of fungus-growing termites with different symbiont transmission modes across disturbance regimes and habitats. Shown is the proportion of encounters (with absolute numbers inside the bars) of termites with horizontal (blue) and vertical transmission (red, dotted) across anthropogenic disturbance regimes for savannah and forest study sites.
Only five out of the 25 identified fungus-growing termite species had vertical transmission. This biases interpretation of the results on total encounter frequencies. Therefore, we did additional analyses in which we compared species-specific encounter frequencies (i.e., the proportion of a species among all encounters; hereafter “encounter proportions;” Figure 1) and tested whether these values differed between species with horizontal vs. vertical symbiont transmission. Across all study sites, encounter proportions did not differ between the five species with vertical transmission and the 20 species with horizontal transmission (Mann Whitney U-test: N1 = 20, N2 = 5, U = 24.00, ns after FDR). This shows that (absolute) encounters with horizontally transmitting species were more common mainly because we had more species with horizontal transmission.
Relative Abundance Within Sites
The results of the total encounter frequencies were biased due to the fact that there were more Macrotermitinae species with horizontal than vertical transmission (20 vs. 5). Thus, we did another set of analyses in which we determined the relative abundance (i.e., the number of encounters of a species within a study site divided by the total number of encounters with fungus-growing termites in this site; this measure only included species that occurred in a site) of each Macrotermitinae species in a study site and tested whether species with horizontal vs. vertical transmission differed in their relative abundance across all sites.
The species that occurred in the most study sites were Microtermes osborni (55 out of 78 study sites) and Microtermes subhyalinus (51 out of 78 study sites), both have vertical transmission. Rare species that only occurred in a single plot were Pseudacanthotermes sp. A, Odontotermes sp. 4 and Protermes sp., all with horizontal transmission.
Ancistrotermes cavithorax had the highest relative abundance within study sites (mean ± SE: 0.29 ± 0.027), followed by Pseudacanthotermes militaris (0.20 ± 0.049), Pseudacanthotermes spiniger (0.20 ± 0.047), and Pseudacanthotermes minor (0.19 ± 0.075). All four species have horizontal transmission. Species with the lowest relative abundances within plots were Odontotermes sp. 4 (0.01, N = 1) and Pseudacanthotermes sp. 1 (0.01 ± 0.002), both with horizontal transmission.
Similar as with total encounter frequencies (Figure 2) and in contrast to expectation, species with horizontal transmission had significantly higher relative abundances within study sites than species with vertical transmission (Mann-Whitney test: N = 439, U = 20676.5, P = 0.024) (Figure 4). This effect was less strong than when using encounter frequencies (Figure 2) because also some species with horizontal transmission had low relative abundances (e.g., Odontotermes sp. 4, Pseudacanthotermes sp. 1; Figure 4). The significant effect disappeared when we split our data set and analyzed it separately for forest and savannah (savannah: N = 260, U = 7494.0, P = 0.219; forest: N = 179, U = 2483.5, P = 0.389), supporting the notion that the effect size was small.

Figure 4. Relative abundances of fungus-growing termite species with horizontal (blue) and vertical (red) transmission. Shown are frequency distributions of relative abundances of species within study sites.
In the savannah as well as the forest, there was a trend that species with horizontal transmission had higher relative abundances than species with vertical transmission at intermediate disturbance levels (savannah: N = 60, U = 305.00, P = 0.067; forest: N = 54, U = 178.50, P = 0.075), while this was not the case in protected study sites (savannah: N = 145, U = 248.00, P = 0.601; forest: N = 78, U = 400.00, P = 0.763) and sites with strong anthropogenic disturbance (savannah: N = 55, U = 329.00, P = 0.818; forest: N = 47, U = 212.00, P = 0.830) (Figure 5). Combining both habitats, species with horizontal transmission had significantly higher relative abundances than species with vertical transmission at intermediate anthropogenic disturbance (N = 114, U = 1103.50, p = 0.005), but not in protected (N = 223, U = 5462.0, P = 0.251) or strongly disturbed study sites (N = 102, U = 1267.5, P = 0.835).
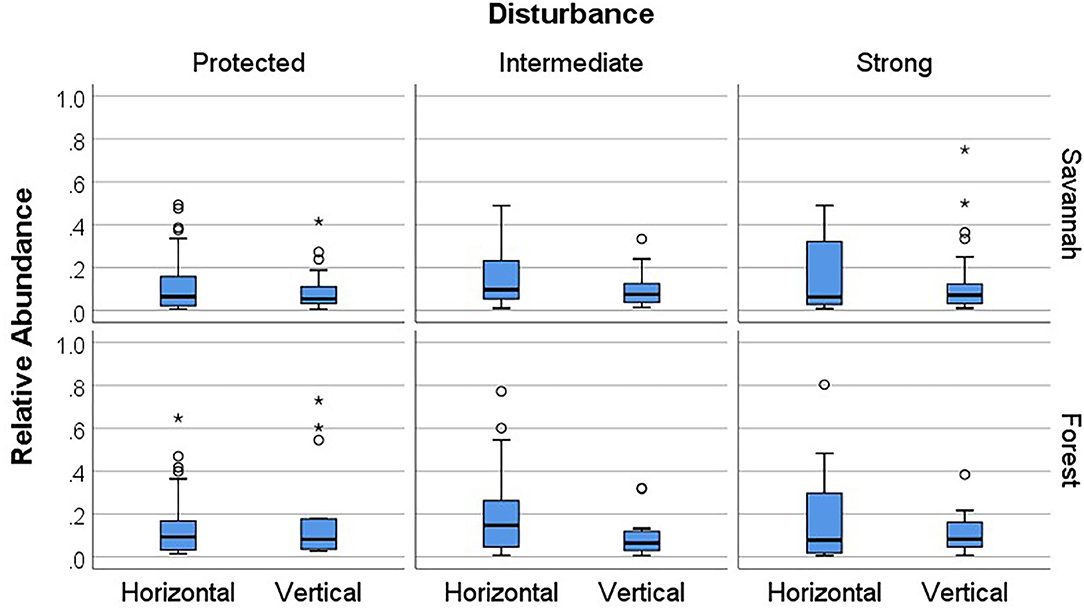
Figure 5. Boxplots of the relative abundances of fungus-growing termite species with horizontal vs. vertical transmission in the savannah and forest across different anthropogenic disturbance regimes. ° are outliers, defined as data that fall not within the whiskers. * are extreme outliers, defined as data that have values more than three times the height of the boxes.
Which Factors Influence the Relative Abundance of a Fungus-Growing Termite?
To analyse the global effects on the occurrence of fungus-growing termite species we run a generalized linear mixed model (GLMM) with gamma error distribution, using “relative abundance” as dependent variable and “transmission mode,” “habitat,” and “disturbance” as fixed factors and “species ID” as random factor. The analysis revealed that transmission mode had a strong and significant effect on the relative abundance of a species (Table 1) with species with horizontal transmission having higher abundances within study sites than species with vertical transmission (Figure 6). Noteworthy is also the trend for an interaction between transmission mode × disturbance (Table 1). The relative abundances of species with horizontal transmission increased under disturbed conditions. In contrast to the former analyses, this GLMM controlled for species identity by using it as random factor.
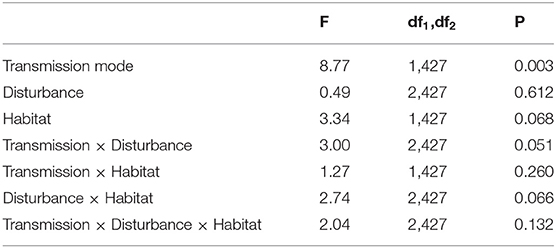
Table 1. Results of a GLMM analyzing the effect of transmission mode, disturbance, and habitat (all fixed factors) on the relative abundance of fungus-growing termites, using species ID as random factor.
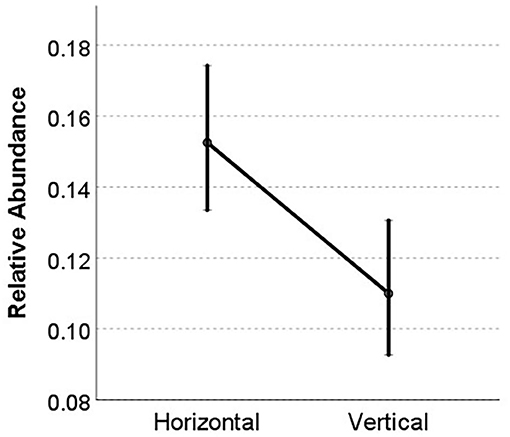
Figure 6. Relative abundances of fungus-growing termite species with horizontal vs. vertical transmission. Shown are estimated marginal means with standard errors as revealed by the GLMM.
Discussion
Ecological Dominance and Transmission Mode of Fungal Symbiont
In contrast to expectation, we did not find fungus-growing termites with vertical transmission to be more abundant. On the contrary, species with horizontal transmission were more likely to be encountered (Figure 2) and also had higher relative abundances (Figures 4, 6). While the results for the encounter frequencies have been biased by the fact that there were more species with horizontal than vertical transmission (which itself may tell something about the evolutionary success of species with horizontal transmission), this effect was accounted for in the relative abundance analyses across species (Figure 4) and also when including species identity in the GLMM (Figure 6). There was only one result which might indicate that species with vertical transmission are more common: the two species that occurred in most study sites, Microtermes subhyalinus and M. osborni, had vertical symbiont transmission. These two species were also among the three most common encountered species (Figure 1, Supplementary Table 1). This may imply that unknown species-specific traits (e.g., broad ecological niche, good dispersal abilities, or fungal identity) rather than transmission mode account for their wide-spread occurrence. Overall, however, our results suggest that species with horizontal transmission were more abundant.
Transmission Mode, Habitat Type, and Disturbance
We found some striking effects of disturbance on the occurrence of fungus-growing species with horizontal vs. vertical transmission mode. Species with horizontal transmission increased with disturbance. This was revealed for encounter rates (Figure 3) and also in the relative abundance analyses (Figure 5), the latter confirming that it is not only an effect of more species with horizontal than vertical transmission. Also the GLMM results, which included species identity, supports this notion with a trend for the interaction between transmission mode and disturbance to be significant (Supplementary Table 1). This implies that species with horizontal transmission thrive better under disturbed conditions than those with vertical transmission. An explanation for this may be that humans directly favor species with horizontal transmission, for instance, because they collect and eat mushrooms and spread spores. This is contrary to the common understanding that, at least nowadays, human consumption of Termitomyces mushrooms constrain the occurrence of Macrotermitinae (Koné et al., 2013). Humans may also indirectly favor horizontal transmitting species because they might be more resistant to disturbance, for example, as they can re-acquire fungal spores during colony establishment when they get lost due to disturbance. By contrast, species with vertical transmission are confined to the single inoculum that the king or queen carries at colony foundation.
Besides disturbance, habitat also seemed to have an effect on transmission mode. In the forest, encounter rates of species with horizontal transmission were around four times (80.6%) more common than those with vertical transmission, while they were only twice (65.4%) as likely in the savannah (Figure 2). However, this effect can be explained by the different proportions of species with horizontal vs. vertical transmission in both habitats. In the forest, 19 out of 22 species (86.3%) had horizontal transmission, while it were 11 out of 16 (68.7%) species in the savannah (Supplementary Table 1). In line with this interpretation, we did not find habitat effects in the relative abundance analyses. The occurrence of more species with horizontal transmission in the forest than in the savannah may be explained by the fact that—despite their strong prevalence in savannahs nowadays—fungus-growing termites evolved in the African rainforest (Aanen and Eggleton, 2005) and that horizontal transmission was most likely the ancestral mode of fungal transmission (Aanen et al., 2002; Nobre et al., 2011b). We also can hypothesize that species with vertical transmission especially evolved in the savannah. This derived trait is associated with species (M. bellicosus, Microtermes) that mainly or exclusively occur in the savannah. This hypothesis can easily be tested when considering data for whole Africa.
Why Are Species With Horizontal Transmission so Common?
Theory predicts that there are two main reasons why associations with uniparental, vertical symbiont transmission should have less conflict than those with horizontal transmission (Frank, 1994, 1995, 1996, 1997, 1998; Foster and Wenseleers, 2006; Leeks et al., 2019), and hence may be more successful. First, uniparental, vertical transmission results in symbionts being closely related (generally clones). Thus, ‘intra-symbiont’ conflict is reduced. Second, strict vertical transmission leads to an alignment of the fitness interests of symbiont and host because symbionts can only increase their fitness by enhancing host fitness. Thus symbiont-host conflict is reduced. A recent modeling and simulation study implies that both, relatedness and fitness alignment, influence the level of cooperation evolving between hosts and symbionts but that relatedness is more important than fitness alignment to explain symbiont cooperation (Leeks et al., 2019).
What is the relatedness of fungal symbionts within a colony? Studies for three common fungus-growing termite species from Southern Africa have shown that the fungus garden of established colonies consists always of a single fungal cultivar, despite horizontal transmission (Aanen et al., 2009). Aanen and colleagues (Aanen, 2006; Aanen et al., 2009) explained this by positive feedback mechanisms within colonies in which the most productive fungal cultivar is positively selected through re-current inoculation of new fungus combs and genetic bottlenecks, after a colony had been inoculated by different cultivars at the incipient stage. This mechanism is in line with lab-experiments that simulated with-nest propagation of the fungal cultivar (Aanen et al., 2009). Thus, intra-symbiont conflict is reduced within termite colonies by generating high relatedness within the fungal cultivar, which stabilizes the association.
However, there is still conflict between host and symbiont in associations with horizontal transmission, for instance, as the fungus is selected to spread independently from its host colony by producing mushrooms with sexual spores that can be picked up by other colonies (Vreeburg et al., 2020; Wisselink et al., 2020). These mushrooms have a high biomass (Yorou et al., 2014). Following three nests of a Pseudacanthotermes species during their fructification period, revealed a mean biomass of 27 kg/colony/year (N.A. Koné, unpublished data). This means a large amount of food lost for consumption by the termite colony. Therefore, there must be other costs associated with vertical transmission to account for its lack of ecological dominance.
What Remains to Be Done?
In order to understand the contribution of the fungal symbiont for the varying ecological success of different fungus-growing species, it will be essential to know which fungi associate with which termite species. This would allow us to test, for instance, whether specific 1:1 associations (“specialists”) are ecologically more successful due to co-evolutionary fine-tuning of plant degrading pathways. Alternatively, termites with a wide range of fungal symbionts may be more ecological dominant because this may allow their host termite species to occupy broader niches, depending on which symbiont they actually are associated with. Lastly, the success of the termite might depend only on symbiont identity when there are some Termitomyces species which are just more competitive or efficient in degrading plant material than others. The fungal symbiont might also shape whole termite community composition through niche differentiation as a recent study hypothesized (Schyra et al., 2019b).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author/s.
Author Contributions
JK conceptionalized the manuscript and did statistical analyses. SS, JK, and NAK collected samples. SS identified all new samples. JK and NAK wrote the manuscript. All authors have read and approved the manuscript.
Funding
We thank the British Ecological Society (BES) for support of NAK within the framework of Ecologists in Africa (ID_4075/4956), the German Academic Exchange Service (DAAD) for a Short-Term Research grants fellowship (ID No. 57381332) to SS, and the German Science Foundation (DFG) for support to JK (KO1895/12-1, KO 1895/24-1).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Karen Meusemann for help with species delimitation, two reviewers for their helpful comments and the OIPR (Office Ivoirien des Parcs et Réserves) for research permits for work in protected areas.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.600318/full#supplementary-material
References
Aanen, D. K. (2006). As you reap, so shall you sow: coupling of harvesting and inoculating stabilizes the mutualism between termites and fungi. Biol. Lett. 2, 209–212. doi: 10.1098/rsbl.2005.0424
Aanen, D. K., de Fine Licht, H. H., Debets, A. J. M., Kerstes, N. A. G., Hoekstra, R. F., and Boomsma, J. J. (2009). High symbiont relatedness stabilizes mutualistic cooperation in fungus-growing termites. Science 326, 1103–1106. doi: 10.1126/science.1173462
Aanen, D. K., and Eggleton, P. (2005). Fungus-growing termites originated in African rain forest. Curr. Biol. 15, 851–855. doi: 10.1016/j.cub.2005.03.043
Aanen, D. K., Eggleton, P., Rouland-Lefevre, C., Guldberg-Froslev, T., Rosendahl, S., and Boomsma, J. J. (2002). The evolution of fungus-growing termites and their mutualistic fungal symbionts. Proc. Natl. Acad. Sci. U.S.A. 99, 14887–14892. doi: 10.1073/pnas.222313099
Bagine, R. K. N., Darlington, J. P. E. C., Brandl, R., and Kaib, M. (1994). “Biodiversity of the termite genus Macrotermes in Kenya: taxonomy, distribution, density patterns, and intraspecific variation,” in Les Insectes Sociaux, eds A. Lenoir, G. Arnold, and M. Lepage (Paris: IUSSI), 28.
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
Bignell, D. E., and Eggleton, P. (2000). “Termites in ecosystems,” in Termites: Evolution, Sociality, Symbiosis and Ecology, eds T. Abe, D. E. Bignell, and M. Higashi (Dordrecht: Kluwer Academic Publishers), 363–387. doi: 10.1007/978-94-017-3223-9_17
Bouillon, A., and Mathot, G. (1966). Quel est ce Termite Africain? (Suppl. 1). Kinshasa: Université Lovanium.
Bouillon, A., and Mathot, G. (1971). Quel est ce Termite Africain? (Suppl. 2). Kinshasa: Université Nationale du Zaire.
da Costa, R. R., Hu, H., Li, H., and Poulsen, M. (2019). Symbiotic plant biomass decomposition in fungus-growing termites. Insects 10:e87. doi: 10.3390/insects10040087
Darlington, J. P. E. C. (1989). The significance of termite mounds in landscapes. Sociobiology 15, 265–266.
Davies, R. G. (2002). Feeding group responses of a neotropical termite assemblage to rain forest fragmentation. Oecologica 133, 233–242. doi: 10.1007/s00442-002-1011-8
Erpenbach, A., Bernhardt-Römermann, M., Wittig, R., and Hahn, K. (2017). The contribution of termite mounds to lanscape-scale variation in vegetation in a West African national park. J. Vegetation Sci. 28, 105–116. doi: 10.1111/jvs.12463
Foster, K. R., and Wenseleers, T. (2006). A general model for the evolution of mutualisms. J. Evol. Biol. 19, 1283–1293. doi: 10.1111/j.1420-9101.2005.01073.x
Frank, S. A. (1994). Genetics of mutualism - the evolution of altruism between species. J. Theor. Biol. 170, 393–400. doi: 10.1006/jtbi.1994.1200
Frank, S. A. (1995). The origin of synergistic symbiosis. J. Theor. Biol. 176, 403–410. doi: 10.1006/jtbi.1995.0208
Grassé, P. P. (1986). Termitologia. Tome III Comportement, socialite, écologie, évolution, systématique. Paris: Masson.
Grasse, P. P., and Noirot, C. (1955). La fondation de nouvelles societes par Bellicositermes natalensis Hav. Insectes Soc. 2, 213–219. doi: 10.1007/BF02224382
Hausberger, B., and Korb, J. (2015). A phylogenetic community approach for studying termite communities in a West African savanna. Biol. Lett. 11:20150625. doi: 10.1098/rsbl.2015.0625
Hausberger, B., and Korb, J. (2016). The impact of anthropogenic disturbance on assembly patterns of termite communities. Biotropica 48, 356–364. doi: 10.1111/btp.12278
Hausberger, B., van Neer, A., Kimpel, D., and Korb, J. (2011). Uncovering cryptic species diversity of a termite community in a West African savanna. Mol. Phylogenet. Evol. 61, 964–969. doi: 10.1016/j.ympev.2011.08.015
Holt, J. A., and Lepage, M. (2000). “Termites and soil properties,” in Termites: Evolution, Sociality, Symbiosis and Ecology, eds T. Abe, D. E. Bignell, and M. Higashi (Dordrecht: Kluwer Academic Publishers), 389–407. doi: 10.1007/978-94-017-3223-9_18
Johnson, R. A. (1981). Colony development and establishment of the fungus comb in Microtermes sp. nr. umbaricus (Sjöstedt) (Isoptera: Macrotermitinae) from Nigeria. Insectes Soc. 28, 3–12. doi: 10.1007/BF02223617
Johnson, R. A., Thomas, R. J., Wood, T. G., and Swift, M. J. (1981). The inoculation of the fungus comb in newly founded colonies of some species of the Macrotermitinae (Isoptera) from Nigeria. J. Nat. Hist. 15, 751–756. doi: 10.1080/00222938100770541
Jones, D. T., and Eggleton, P. (2000). Sampling termite assemblages in tropical forests: testing a rapid biodiversity assessment protocol. J. Appl. Ecol. 37, 191–203. doi: 10.1046/j.1365-2664.2000.00464.x
Josens, G. (1972). Etudes biologique et écologique des termites (Isoptera) de la savane de Lamto-Pakobo (Côte d'Ivoire) (Ph.D. thesis). Université Libre de Bruxelles, Brussels, Belgium.
Koné, N. A., Dosso, K., Konate, S., Kouadio, J. Y., and Linsenmair, K. E. (2011). Environmental and biological determinants of Termitomyces species seasonal fructification in central and southern Cote d'Ivoire. Insectes Soc. 58, 371–382. doi: 10.1007/s00040-011-0154-1
Koné, N. A., Yéo, K., Konaté, S., and Linsenmair, K. E. (2013). Socio-economical aspects of the exploitation of Termitomyces fruit bodies in central and southern Côte d'Ivoire: raising awareness for their sustainable use. J. Appl. Biosci. 70, 5580–5590. doi: 10.4314/jab.v70i1.98759
Korb, J. (2020). “Fungus-growing termites: an eco-evolutionary perspective,” in The Convergent Evolution of Agriculture in Humans and Insects, eds T. R. Schultz, P. N. Peregrine, and R. Gawne (Cambridge, MA: MIT Press).
Korb, J., and Aanen, D. K. (2003). The evolution of uniparental transmission of fungal symbionts in fungus-growing termites (Macrotermitinae). Behav. Ecol. Sociobiol. 53, 65–71. doi: 10.1007/s00265-002-0559-y
Korb, J., Kasseney, B. D., Cakpo, Y. T., Casalla, R. H., Gbenyedji, J. N. B. K., Ilboudo, M. E., et al. (2019). Termite taxonomy, challenges and prospects: West Africa, a case example. Insects 10:e32. doi: 10.3390/insects10010032
Leeks, A., dos Santos, M., and West, S. A. (2019). Transmission, relatedness, and the evolution of cooperative symbionts. J. Evol. Biol. 32, 1036–1045. doi: 10.1111/jeb.13505
Lepage, M., and Darlington, J. P. E. C. (2000). “Population dynamics of termites,” in Termites: Evolution, Sociality, Symbiosis, and Ecology, eds T. Abe, D. E. Bignell, and M. Higashi (Dordrecht: Klumer Academic Publishers), 333–361. doi: 10.1007/978-94-017-3223-9_16
Nobre, T., Fernandes, C., Boomsma, J. J., Korb, J., and Aanen, D. K. (2011a). Farming termites determine the genetic population structure of Termitomyces fungal symbionts. Mol. Ecol. 20, 2023–2033. doi: 10.1111/j.1365-294X.2011.05064.x
Nobre, T., Kone, N. A., Konate, S., Linsenmair, K. E., and Aanen, D. K. (2011b). Dating the fungus-growing termites' mutualism shows a mixture between ancient codiversification and recent symbiont dispersal across divergent hosts. Mol. Ecol. 20, 2619–2627. doi: 10.1111/j.1365-294X.2011.05090.x
Nobre, T., Rouland-Lefevre, C., and Aanen, D. K. (2011c). “Comparative biology of fungus cultivation in termites and ants,” in Biology of Termites: A Modern Synthesis, eds D. E. Bignell, Y. Roisin, and N. Lo (Dordrecht; Heidelberg; London; New York, NY: Springer), 193–210. doi: 10.1007/978-90-481-3977-4_8
Pomeroy, D. E., Bagine, R. K., and Darlington, J. P. E. C. (1991). Fungus-growing termites in East African savannas. Afr. Wildlife Res. Manag. 1991, 41–50.
Poulsen, M. (2015). Towards an integrated understanding of the consequences of fungus domestication on the fungus growing gut microbiota. Environ. Microbiol. 17, 2562–2572. doi: 10.1111/1462-2920.12765
Rouland, C., Lenoir, F., and Lepage, M. (1991). The role of the symbiotic fungus in the digestive metabolism of several species of fungus-growing termites. Comp. Biochem. Physiol. 99, 657–663. doi: 10.1016/0300-9629(91)90146-4
Rouland-Lefèvre, C. (2000). “Symbiosis with fungi,” in Termites: Evolution, sociality, symbioses, ecology, eds T. Abe, D. E. Bignell, and M. Higashi (Dordrecht: Kluwer Academic Publishers), 289–306. doi: 10.1007/978-94-017-3223-9_14
Sands, W. A. (1998). The Identification of Worker Castes of Termite Genera From Soils of Africa and the Middle East. Wallingford: CAB International.
Schyra, J., Gbenyedji, J. N. B. K., and Korb, J. (2019a). A comparison of termite assemblages from West African savannah and forest ecosystems using morphological and molecular markers. PLoS ONE 14:e0216986. doi: 10.1371/journal.pone.0216986
Schyra, J., and Korb, J. (2019). Termite communities along a disturbance gradient in a West African Savanna. Insects 10:e17. doi: 10.3390/insects10010017
Schyra, J., Scheu, S., and Korb, J. (2019b). Cryptic niche differentiation in West African savannah termites as indicated by stable isotopes. Ecol. Entomol. 44, 190–196. doi: 10.1111/een.12687
Vreeburg, S. M. E., de Ruijter, N. C. A., Zwaan, B. J., da Costa, R. R., Poulsen, M., and Aanen, D. K. (2020). Asexual and sexual reproduction are two separate developmental pathways in a Termitomyces species. Biol. Lett. 16:20200394. doi: 10.1098/rsbl.2020.0394
Wisselink, M., Aanen, D. K., and van‘t Padje, A. (2020). The longevity of colonies of fungus-growing termites and the stability of the symbiosis. Insects 11:527. doi: 10.3390/insects11080527
Yorou, N. S., Koné, N. A. K., Guissou, M.-L., Guelly, A. K., De Kesel, A., and Ekue, M. R. (2014). “Biodiversity and sustainable use of wild edible fungi in the Soudanian centre of endemism: a plea for valorisation,” in Ectomycorrhizal Symbioses in Tropical and Neotropical Forests, eds A. M. Bâ, K. L. McGuire, and A. G. Diédhiou (London; New York, NY: CRC Press), 241.
Keywords: cooperation, conflict, host, symbiosis, symbiont, termites, transmission mode
Citation: Korb J, Silué SK and Koné NA (2020) Can Differences in Symbiont Transmission Mode Explain the Abundance and Distribution of Fungus-Growing Termites in West Africa? Front. Ecol. Evol. 8:600318. doi: 10.3389/fevo.2020.600318
Received: 29 August 2020; Accepted: 20 November 2020;
Published: 18 December 2020.
Edited by:
Alberto Arab, Federal University of ABC, BrazilReviewed by:
Aram Mikaelyan, North Carolina State University, United StatesDuur Kornelis Aanen, Wageningen University and Research, Netherlands
Copyright © 2020 Korb, Silué and Koné. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Judith Korb, judith.korb@biologie.uni-freiburg.de