Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling Strategy
2.2. Soil Chemical Analyses
2.3. Calculations of the Effective Cation Exchange Capacity, Base Saturation%, Al:Ca Ratio, and Effective Acidity
2.4. Statistical Analysis
3. Results
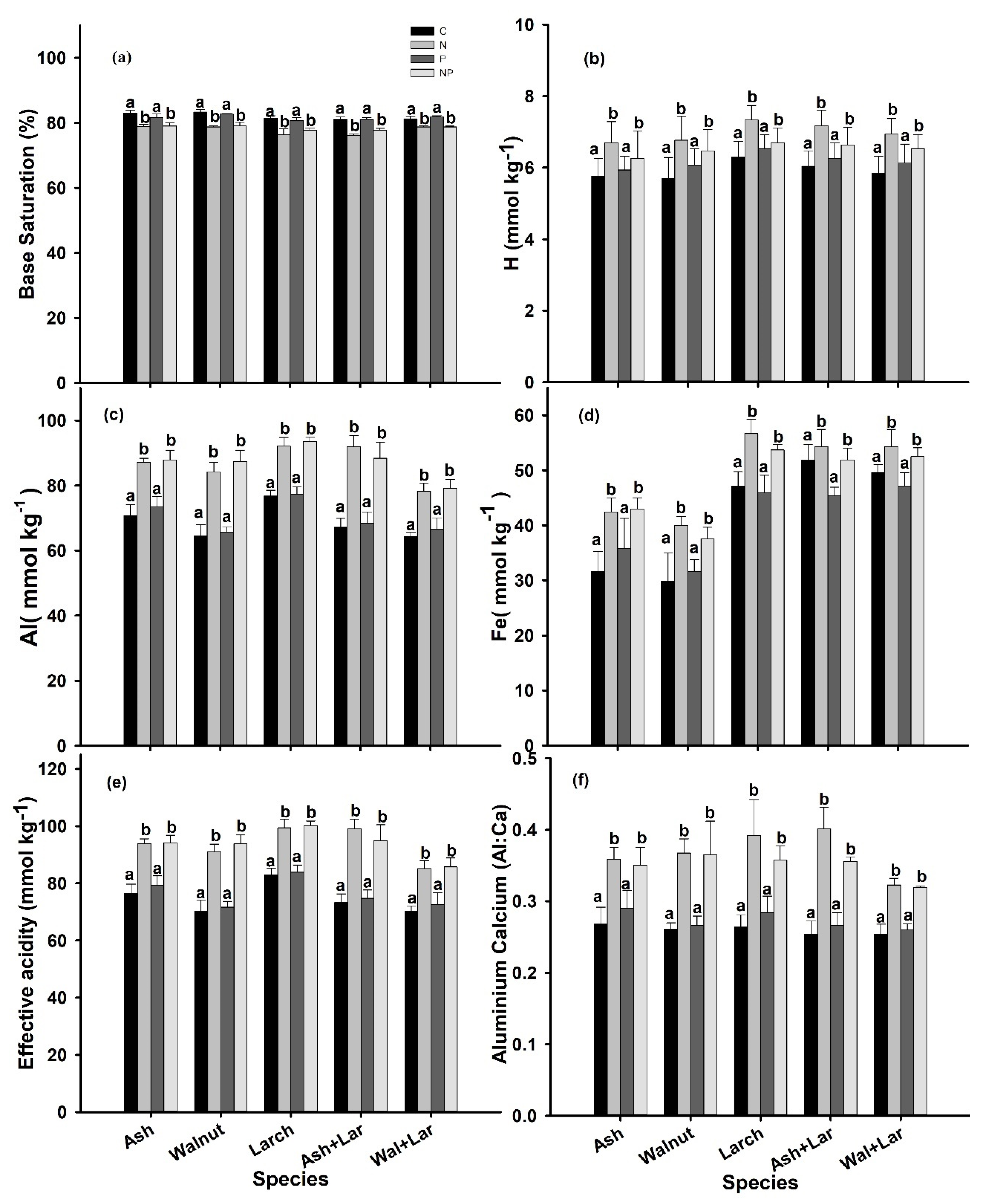
3.1. Soil Chemical Properties
3.2. Soil P Fractions
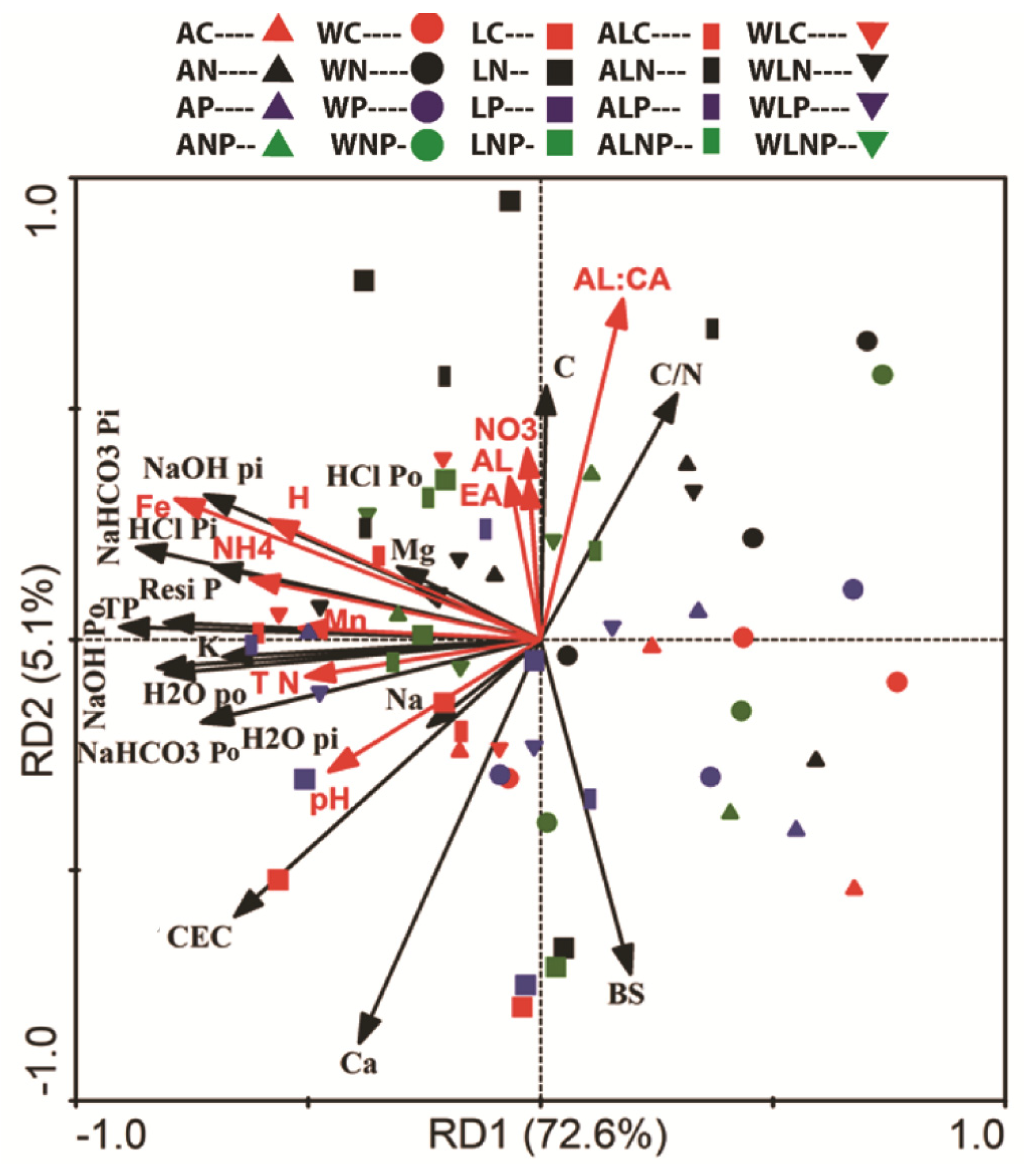
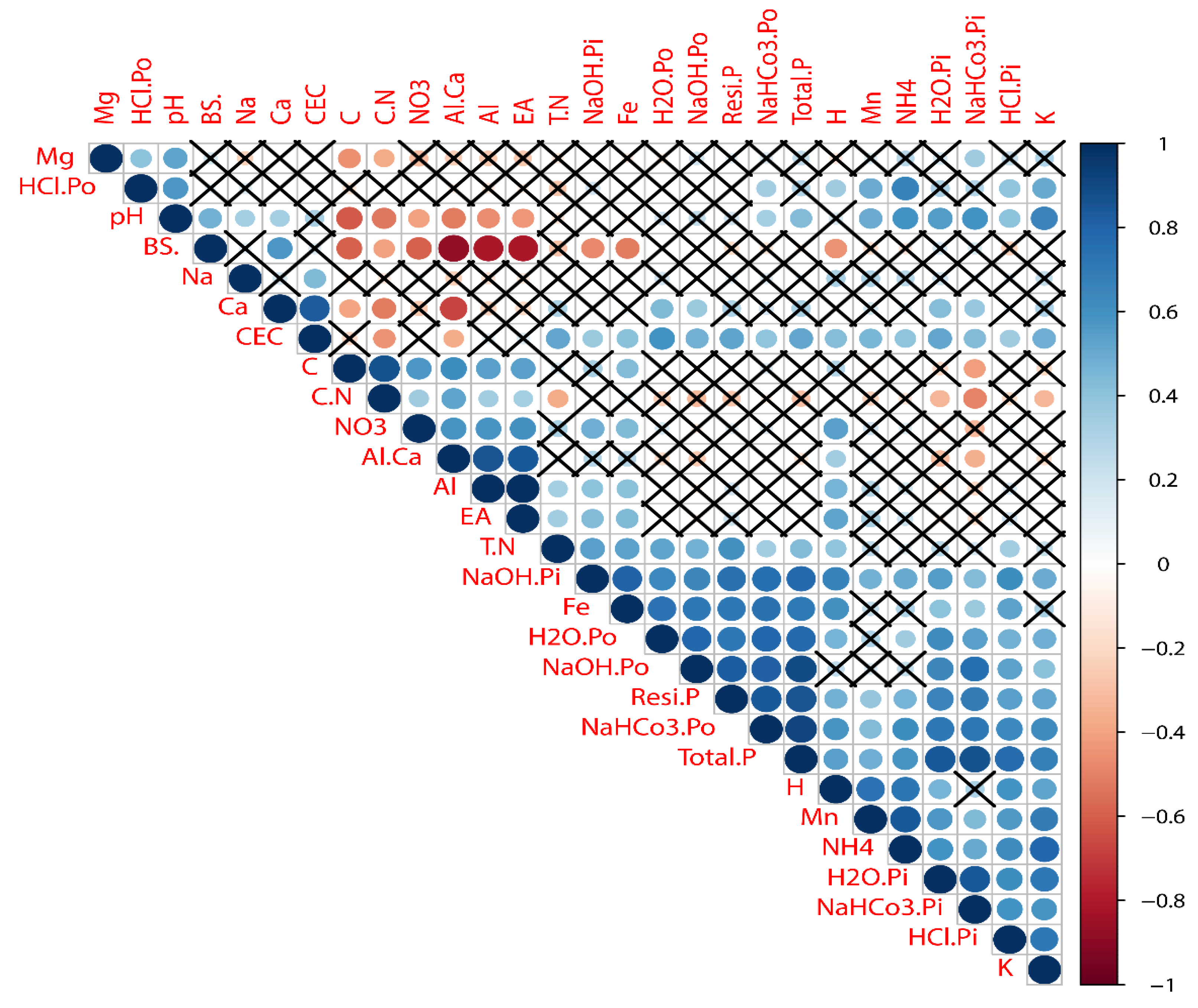
3.3. Correlation between Soil P Fractions and Chemical Properties
4. Discussion
4.1. Effects of Additional N on the Soil Chemical Properties
4.2. Effects of Additional P on the Soil Chemical Properties
4.3. Effects of Fertilizations on P Fractions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galloway, J.; Townsend, A.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.; Martinelli, L.; Seitzinger, S.; Sutton, M. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science (New York, N.Y.) 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penuelas, J.; Janssens, I.; Ciais, P.; Sardans, J. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Chang. Biol. 2020, 26. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Yu, G.; He, N.; Zhan, X.; Fang, H.; Sheng, W.; Zuo, Y.; Zhang, D.; Wang, Q.-F. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity. Sci. Rep. 2014, 4, 3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced Nitrogen Deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.P.; Wang, Y.S.; Tang, G.Q.; Wu, D. Wet and dry deposition of atmospheric nitrogen at ten sites in Northern China. Atmos. Chem. Phys. 2012, 12, 6515–6535. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.; Tang, A.; Zhang, Y.; Luo, X.; Shen, J.; Wang, W.; Li, K.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef]
- Suding, K.; Collins, S.; Gough, L.; Clark, C.; Cleland, E.; Gross, K.; Milchunas, D.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. PNAS 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Niu, S. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10. [Google Scholar] [CrossRef]
- Lucas, R.W.; Klaminder, J.; Futter, M.; Bishop, K.; Gustaf, E.; Laudon, H.; Högberg, P. A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams. For. Ecol. Manag. 2011, 262, 95–104. [Google Scholar] [CrossRef]
- Lebauer, D.; Treseder, K. Nitrogen Limitation of Net Primary Productivity in Terrestrial Ecosystems is Globally Distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.; Howarth, R. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Małek, S. Sustainability of Picea abies of Istebna provenance in Dupniański stream catchment as dependent on stand age class. Dendrobiology 2009, 61, 95–104. [Google Scholar]
- Yang, K.; Zhu, J.; Gu, J.; Yu, L.; Wang, Z. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Ann. For. Sci. 2015, 72, 435–442. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Gallo, M.; Lauber, C.; Waldrop, M.; Zak, D. Extracellular Enzyme Activities and Soil Organic Matter Dynamics for Northern Hardwood Forests receiving Simulated Nitrogen Deposition. Biogeochemistry 2005, 75. [Google Scholar] [CrossRef] [Green Version]
- Nave, L.E.; Vance, E.; Swanston, C.; Curtis, P.S. Impacts of elevated N inputs on north temperate forest soil C storage, C/N, and net N-mineralization. Geoderma 2009, 153. [Google Scholar] [CrossRef]
- Cusack, D.; Silver, W.; Torn, M.; Burton, S.; Firestone, M. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Futa, B.; Mocek-Płóciniak, A. The influence of uncontrolled grass burning on biochemical qualities of soil. J. Agric. Eng. 2016, 61, 98–100. [Google Scholar]
- de Groot, C.; Marcelis, L.F.M.; Boogaard, R.; Lambers, H. Growth and dry-mass partitioning in tomato as affected by phosphorus nutrition and light. Plant Cell Environ. 2001, 24, 1309–1317. [Google Scholar] [CrossRef]
- Aber, J.; Nadelhoffer, K.; Steudler, P.; Melillo, J. Nitrogen Saturation in Northern Forest Ecosystems. Bioscience 1989, 39, 378–386. [Google Scholar] [CrossRef]
- Binkley, D.; Burnham, H.; Allen, H. Waterquality Impacts of Forest Fertilization with Nitrogen and Phosphorus. For. Ecol. Manag. 1999, 121, 191–213. [Google Scholar] [CrossRef]
- Hou, E.; Luo, Y.; Kuang, Y.; Chen, C.; Lu, X.; Jiang, L.; Luo, X.; Wen, D. Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nat. Commun. 2020, 11, 637. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.; Porder, S.; Houlton, B.; Chadwick, O. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, K. Soil acidification and adaptations of plants and microorganisms in Bornean tropical forests. Ecol. Res. 2014, 29. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Xiankai, L.; Zhou, K.; Chen, H.; Zhu, X.; Mori, T.; Mo, J. Effects of long-term nitrogen and phosphorus additions on soil acidification in an N-rich tropical forest. Geoderma 2017, 285. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Fu, S. Effects of nitrogen deposition and increased precipitation on soil phosphorus dynamics in a temperate forest. Geoderma 2020, 380, 114650. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Wen, D.; Yu, K. Soil potential labile but not occluded phosphorus forms increase with forest succession. Biol. Fertil. Soils 2016, 52, 41–51. [Google Scholar] [CrossRef]
- Helfenstein, J.; Jegminat, J.; McLaren, T.I.; Frossard, E. Soil solution phosphorus turnover: Derivation, interpretation, and insights from a global compilation of isotope exchange kinetic studies. Biogeosciences 2018, 15, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Bünemann, E.K. Assessment of gross and net mineralization rates of soil organic phosphorus—A review. Soil Biol. Biochem. 2015, 89, 82–98. [Google Scholar] [CrossRef]
- Helfenstein, J.; Pistocchi, C.; Oberson, A.; Tamburini, F.; Goll, D.S.; Frossard, E. Estimates of mean residence times of phosphorus in commonly considered inorganic soil phosphorus pools. Biogeosciences 2020, 17, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Helfenstein, J.; Tamburini, F.; von Sperber, C.; Massey, M.S.; Pistocchi, C.; Chadwick, O.A.; Vitousek, P.M.; Kretzschmar, R.; Frossard, E. Combining spectroscopic and isotopic techniques gives a dynamic view of phosphorus cycling in soil. Nat. Commun. 2018, 9, 3226. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Chapin Iii, F.S.; Pons, T. Plant Physiological Ecology: Second Edition; Springer Science & Business Media: New York, NY, USA, 2008; pp. 1–604. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.; Shaver, G.; Smith, S. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Rosling, A.; Midgley, M.; Cheeke, T.; Urbina, H.; Fransson, P.; Phillips, R. Phosphorus cycling in deciduous forest soil differs between stands dominated by ecto- and arbuscular mycorrhizal trees. New Phytol. 2015, 209. [Google Scholar] [CrossRef] [Green Version]
- Oberson, A.; Joner, E.J. Microbial turnover of phosphorus in soil. In Organic Phosphorus in the Environment; CABI Publishing: Wallingford, UK, 2005; pp. 133–164. [Google Scholar] [CrossRef]
- Rhoades, C. Single-tree influences on soil properties in agroforestry: Lessons from natural forest and savanna ecosystems. Agrofor. Syst. 1997, 35, 71–94. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why Do Tree Species Affect Soils? The Warp and Woof of Tree–Soil Interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Bardgett, R.; Bowman, W.; Kaufmann, R.; Schmidt, S.; Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef]
- West, J.B.; Hobbie, S.E.; Reich, P.B. Effects of plant species diversity, atmospheric [CO2], and N addition on gross rates of inorganic N release from soil organic matter. Glob. Chang. Biol. 2006, 12, 1400–1408. [Google Scholar] [CrossRef]
- Walker, T.N.; Ward, S.E.; Ostle, N.J.; Bardgett, R.D. Contrasting growth responses of dominant peatland plants to warming and vegetation composition. Oecologia 2015, 178, 141–151. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, G.-h.; Fu, B.-J.; Zheng, X. Relationship between plant species diversity and soil microbial functional diversity along a longitudinal gradient in temperate grasslands of Hulunbeir, Inner Mongolia, China. Ecol. Res. 2008, 23, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Bragazza, L.; Bardgett, R.; Mitchell, E.; Buttler, A. Linking soil microbial communities to vascular plant abundance along a climate gradient. New Phytol. 2014, 205. [Google Scholar] [CrossRef]
- Fahey, T.J.; Yavitt, J.B.; Sherman, R.E.; Maerz, J.C.; Groffman, P.M.; Fisk, M.C.; Bohlen, P.J. Earthworm effects on the incorporation of litter C and N into soil organic matter in a sugar maple forest. Ecol. Appl. 2013, 23, 1185–1201. [Google Scholar] [CrossRef] [Green Version]
- Payn, T.; Carnus, J.-M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Frédéric, A.; Mollicone, D.; Stibig, H.-J.; Aksenov, D.; Laestadius, L.; Li, Z.; Popatov, P.; Yaroshenko, A. Areas of rapid forest-cover change in boreal Eurasia. For. Ecol. Manag. 2006, 237, 322–334. [Google Scholar] [CrossRef]
- Zhang, P.; Shao, G.; Zhao, G.; Le Master, D.C.; Parker, G.R.; Dunning, J.B.; Li, Q. China’s Forest Policy for the 21st Century. Science 2000, 288, 2135. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, P.; Kong, C.-H. Effect of larch (Larix gmelinii Rupr.) root exudates on Manchurian walnut (Juglans mandshurica Maxim.) growth and soil juglone in a mixed-species plantation. Plant Soil 2010, 329, 249–258. [Google Scholar] [CrossRef]
- Shi, F.; Chen, X.; Chen, N. Study on the artificial mixed forest of Juglans mandshurica and Larix olgensis. J. Northeast For. Univ. 1991, 19, 32–43. [Google Scholar]
- Razaq, M.; Salahuddin; Shen, H.-l.; Sher, H.; Zhang, P. Influence of biochar and nitrogen on fine root morphology, physiology, and chemistry of Acer mono. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Guo, D.; Li, H.; Mitchell, R.; Han, W.; Hendricks, J.; Fahey, T.; Hendrick, R. Fine root heterogeneity by branch order: Exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods. New Phytol. 2008, 177, 443–456. [Google Scholar] [CrossRef]
- Gundersen, P.; Rasmussen, L. Nitrification in Forest Soils: Effects from Nitrogen Deposition on Soil Acidification and Aluminum Release. Rev. Environ. Contam. Toxicol. 1990, 113, 1–45. [Google Scholar] [CrossRef]
- Bowden, R. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. FEMS Microbiol. Lett. 2004. [Google Scholar] [CrossRef]
- Mei, l.; Gu, J.; Zhang, Z.; Wang, Z. Responses of fine root mass, length, production and turnover to soil nitrogen fertilization in Larix gmelinii and Fraxinus mandshurica forests in Northeastern China. J. For. Res. 2010, 15, 194–201. [Google Scholar] [CrossRef]
- Hedley, M.; Stewart, J.; Chauhan, B. Changes in Inorganic and Organic Soil Phosphorus Fractions Induced by Cultivation Practices and by Laboratory Incubations1. Soil Sci. Soc. Am. J. 1982, 46. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of available P by sequential extraction. Soil Sampl. Methods Anal. 1993, 7, 75–86. [Google Scholar]
- Robertson, G.P.; Sollins, P.; Ellis, B.G.; Lajtha, K. Exchangeable ions, pH, and cation exchange capacity. Stand. Soil Methods Long Term Ecol. Res. 1999, 28, 462. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; Team, R.C.: Vienna, Austria, 2013. [Google Scholar]
- Shi, L.; Zhang, H.; Liu, T.; Zhang, W.; Shao, Y.; Ha, D.; Li, Y.; Zhang, C.; Cai, X.-A.; Rao, X.; et al. Consistent effects of canopy vs. understory nitrogen addition on the soil exchangeable cations and microbial community in two contrasting forests. Sci. Total Environ. 2016, 553, 349–357. [Google Scholar] [CrossRef]
- Carreira, J.; García-Ruiz, R.; Lietor, J.; Harrison, A. Biochemistry. Changes in soil phosphatase activity and P transformation rates induced by application of N-and S-containing acid-mist to a forest canopy. Soil Biol. Biochem. 2000, 32, 1857–1865. [Google Scholar] [CrossRef]
- Bowman, W.; Cleveland, C.; Halada, L.; Hreško, J.; Baron, J. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Lu, X.; Mo, J.; Gundersern, P.; Zhu, W.-X.; Zhou, G.-Y.; Li, D.; Zhang, X. Effect of Simulated N Deposition on Soil Exchangeable Cations in Three Forest Types of Subtropical China. Pedosphere 2009, 19, 189–198. [Google Scholar] [CrossRef]
- Bolan, N.; Hedley, M.; White, R. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Raubuch, M.; Beese, F.; Bolger, T.; McCarthy, F.; Anderson, J.M.; Splatt, P.; Willison, T.; Coûteaux, M.M.; Ineson, P.; Berg, M.P.; et al. Acidifying processes and acid-base reactions in forest soils reciprocally transplanted along a European transect with increasing pollution. Biogeochemistry 1998, 41, 71–88. [Google Scholar] [CrossRef]
- Indian People Organizing for Change (IPOC). Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; ISBN 110705799X. [Google Scholar]
- Yan, E.-R.; Wang, X.; Huang, J.-J.; Li, G.-Y.; Zhou, W. Decline of soil nitrogen mineralization and nitrification during forest conversion of evergreen broad-leaved forest to plantations in the subtropical area of Eastern China. Biogeochemistry 2008, 89, 239–251. [Google Scholar] [CrossRef]
- Tafazoli, M.; Hojjati, S.; Jalilvand or Djalilvand, H.; Lamersdorf, N. Simulated Nitrogen Deposition Reduces the Concentration of Soil Base Cations in Acer velutinum Bioss. Plantation, North of Iran. J. Soil Sci. Plant Nutr. 2019, 19, 440–449. [Google Scholar] [CrossRef]
- Falkengren-Grerup, U.; Brunet, J.; Diekmann, M. Nitrogen mineralisation in deciduous forest soils in south Sweden in gradients of soil acidity and deposition. Environ. Pollut. 1998, 102, 415–420. [Google Scholar] [CrossRef]
- Eberwein, J.; Shen, W.; Jenerette, G. Michaelis-Menten kinetics of soil respiration feedbacks to nitrogen deposition and climate change in subtropical forests. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Katou, H. A pH-dependence implicit formulation of cation-and anion-exchange capacities of variable-charge soils. Soil Sci. Soc. Am. J. 2002, 66, 1218–1224. [Google Scholar] [CrossRef]
- Malhi, S.; Nyborg, M.; Harapiak, J. Effects of long-term N fertilizer-induced acidification and liming on micronutrients in soil and in bromegrass hay. Soil Till. Res. 1998, 48, 91–101. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.E.R.; Fang, Y.; Li, D.; Wang, H.U.I. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Chen, H.; Gurmesa, G.A.; Zhang, W.; Zhu, X.; Zheng, M.; Mao, Q.; Zhang, T.; Mo, J. Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing. Funct. Ecol. 2015, 30. [Google Scholar] [CrossRef]
- Homeier, J.; Hertel, D.; Camenzind, T.; Cumbicus, N.; Maraun, M.; Martinson, G.; Poma López, L.; Rillig, M.; Sandmann, D.; Scheu, S.; et al. Tropical Andean Forests Are Highly Susceptible to Nutrient Inputs—Rapid Effects of Experimental N and P Addition to an Ecuadorian Montane Forest. PLoS ONE 2012, 7, e47128. [Google Scholar] [CrossRef]
- Zhu, F.; Lu, X.; Liu, L.; Mo, J. Phosphate addition enhanced soil inorganic nutrients to a large extent in three tropical forests. Sci. Rep. 2015, 5, 7923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrich, B. Natural and anthropogenic components of soil acidification. Zeitschrift für Pflanzenernährung und Bodenkunde 1986, 149, 702–717. [Google Scholar] [CrossRef]
- Chen, H. Phosphatase activity and P fractions in soils of an 18-year-old Chinese fir (Cunninghamia lanceolata) plantation. For. Ecol. Manag. 2003, 178, 301–310. [Google Scholar] [CrossRef]
- Dossa, E.; Diedhiou, S.; Compton, J.; Assigbetse, K.; Dick, R. Spatial patterns of P fractions and chemical properties in soils of two native shrub communities in Senegal. Plant Soil 2009, 327, 185–198. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.; Zhang, Y.; Shen, J.; Han, X.-W.; Zhang, W.; Christie, P.; Goulding, K.; Vitousek, P.M.; Zhang, F.S. Significant Acidification in Major Chinese Croplands. Science (New York, N.Y.) 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.D.; Smethurst, P.J. Surface soil changes in base cation concentrations in fertilised hardwood and softwood plantations in Australia. For. Ecol. Manag. 2004, 191, 253–265. [Google Scholar] [CrossRef]
- Weand, M.; Arthur, M.; Lovett, G.; Sikora, F.; Weathers, K. The phosphorus status of northern hardwoods differs by species but is unaffected by nitrogen fertilization. Biogeochemistry 2010, 97, 159–181. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, X.Y.; Hu, Y.; Zeng, D.-H. Effects of nitrogen addition on nutrient allocation and nutrient resorption efficiency in Larix gmelinii. Sci. Silvae Sin. 2010, 46, 14–19. [Google Scholar]
- Khan, K.S.; Joergensen, R.G. Relationships between P fractions and the microbial biomass in soils under different land use management. Geoderma 2012, 173–174, 274–281. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. Increasing drought decreases phoshorus availability in an evergreen Mediterranean forest. Plant Soil 2004, 267, 367–377. [Google Scholar] [CrossRef]
- Cross, A.; Schlesinger, W. A Literature Review and Evaluation of the Hedley Fractionation: Applications to the Biogeochemical Cycle of Soil Phosphorus in Natural Ecosystems. Geoderma 1995, 64, 197–214. [Google Scholar] [CrossRef]
- Liu, Q.; Loganathan, P.; Hedley, M.; Skinner, M. The mobilisation and fate of soil and rock phosphate in the rhizosphere of ectomycorrhizal Pinus radiata seedlings in an Allophanic soil. Plant Soil 2004, 264, 219–229. [Google Scholar] [CrossRef]
- Beck, M.; Sanchez, P. Soil Phosphorus Fraction Dynamics during 18 Years of Cultivation on a Typic Paleudult. Soil Sci. Soc. Am. J. 1994, 58, 1424–1431. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Species | Density (ha−1) | DBH (cm) | Height (m) |
|---|---|---|---|---|
| Mono-Culture | Walnut | 1495 | 57.3 | 12.0 |
| Mixed-Culture | Walnut | 650 | 67.1 | 12.5 |
| Larch | 735 | 44.8 | 13.4 | |
| Mono-Culture | Ash | 1535 | 63.5 | 13.1 |
| Mixed-Culture | Ash | 685 | 64.7 | 13.5 |
| Larch | 740 | 43.3 | 12.8 | |
| Mono-Culture | Larch | 1285 | 81.2 | 13.7 |
| Forest Type | Species | Soil pH | Total N (g kg−1) | Total P (mg kg−1) | SOC (g kg−1) | C:N Ratio |
|---|---|---|---|---|---|---|
| Monoculture | Walnut | 5.65 | 7.1 | 1530.0 | 91.4 | 12.9 |
| Mixed-Culture | Walnut*Larch | 5.58 | 6.2 | 1487.7 | 100.2 | 16.1 |
| Monoculture | Ash | 5.65 | 6.3 | 1479.6 | 91.7 | 14.5 |
| Mixed-Culture | Ash*Larch | 5.61 | 6.1 | 1465.9 | 100.7 | 16.5 |
| Monoculture | Larch | 5.57 | 6.2 | 1464.5 | 90.7 | 14.6 |
| Source of Variance | df | H+ | Al3+ | Fe3+ | EA | BS% | Al:Ca | Mg2+ | Ca2+ |
|---|---|---|---|---|---|---|---|---|---|
| Species (Spp.) | 4 | 0.546 | <0.001 | <0.001 | <0.001 | 0.004 | 0.228 | 0.973 | 0.183 |
| Fertilization (Fert) | 3 | 0.012 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.032 | 0.044 |
| Spp.*Fert | 12 | 1.000 | 0.720 | 0.853 | 0.770 | 0.955 | 0.931 | 1.000 | 0.974 |
| Source of Variance | df | NO3−-N | SOC | C:N ratio | pH | H2O Pi | NaHCO3 Pi | NaOH Pi | NaOH Po |
| Species (Spp.) | 4 | 0.394 | 0.024 | <0.001 | 0.725 | 0.028 | <0.001 | <0.001 | <0.001 |
| Fertilization (Fert) | 3 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 | 0.009 | 0.001 |
| Spp.*Fert | 12 | 0.742 | 0.997 | 0.989 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Species/ Fertilizer | Mg2+ (mmol kg−1) | Ca2+ (mmol kg−1) | NO3−-N (mg kg−1) | SOC (g kg−1) | C:N Ratio | Soil pH | |
|---|---|---|---|---|---|---|---|
| Ash | C | 26.9 ± 1.6 a | 264.8 ± 9.7 a | 35.2 ± 0.9 a | 87.1 ± 5.3 a | 12.1 ± 0.8 a | 5.6 ± 0.2 a |
| N | 23.8 ± 1.9 b | 243.3 ± 8.4 b | 66.4 ± 1.4 b | 118.9 ± 4.0 b | 16.2 ± 0.4 b | 5.2 ± 0.1 b | |
| P | 26.0 ± 1.0 a | 254.8 ± 10.3 a | 34.1 ± 0.8 a | 89.4 ± 4.3 a | 12.9 ± 1.1 a | 5.4 ± 0.3 a | |
| NP | 25.6 ± 1.6 a | 252.2 ± 10.9 a | 38.4 ± 1.6 c | 104.7 ± 3.4 b | 14.6 ± 0.8 b | 5.4 ± 0.1 a | |
| Walnut | C | 27.3 ± 1.8 a | 247.1 ± 10.3 a | 35.0 ± 0.9 a | 84.0 ± 7.7 a | 13.4 ± 1.2 a | 5.6 ± 0.5 a |
| N | 24.1 ± 1.8 b | 230.9 ± 17.4 b | 66.2 ± 1.4 b | 115.8 ± 4.1 b | 17.1 ± 0.5 b | 5.2 ± 0.1 b | |
| P | 26.3 ± 1.5 a | 247.6 ± 16.6 a | 34.1 ± 0.8 a | 85.1 ± 3.8 a | 13.9 ± 0.8 a | 5.7 ± 0.3 a | |
| NP | 25.7 ± 1.6 a | 244.7 ± 21.4 a | 41.8 ± 6.2 c | 111.2 ± 7.7 b | 17.7 ± 1.5 b | 5.6 ± 0.1 a | |
| Larch | C | 26.5 ± 1.6 a | 292.5 ± 14.3 a | 35.2 ± 0.9 a | 87.9 ± 4.8 a | 11.1 ± 0.7 a | 5.5 ± 0.2 a |
| N | 23.4 ± 1.6 b | 242.0 ± 16.3 b | 75.6 ± 1.1 b | 119.4 ± 5.7 b | 14.5 ± 1.1 b | 5.1 ± 0.1 b | |
| P | 25.9 ± 1.0 a | 275.1 ± 15.4 a | 34.2 ± 0.8 a | 87.1 ± 5.1 a | 11.3 ± 0.7 a | 5.5 ± 0.2 a | |
| NP | 25.3 ± 1.0 a | 263.9 ± 18.3 a | 48.8 ± 7.1 c | 101.8 ± 3.0 b | 13.6 ± 0.3 b | 5.5 ± 0.2 a | |
| Ash + Larch | C | 26.7 ± 1.4 a | 266.6 ± 11.0 a | 45.6 ± 9.3 a | 95.9 ± 4.9 a | 13.5 ± 0.9 a | 5.6 ± 0.2 a |
| N | 23.4 ± 2.1 b | 230.5 ± 10.8 b | 64.9 ± 1.4 b | 125.9 ± 7.2 b | 17.3 ± 1.5 b | 5.1 ± 0.1 b | |
| P | 25.8 ± 1.4 a | 258.1 ± 11.2 a | 34.0 ± 0.9 a | 97.3 ± 4.9 a | 13.6 ± 0.2 a | 5.6 ± 0.3 a | |
| NP | 25.1 ± 1.7 a | 247.9 ± 9.8 a | 47.0 ± 5.7 c | 112.6 ± 9.8 b | 16.0 ± 1.6 b | 5.5 ± 0.2 a | |
| Walnut + Larch | C | 26.7 ± 0.9 a | 254.4 ± 10.1 a | 35.1 ± 1.0 a | 97.9 ± 4.8 a | 13.9 ± 0.9 a | 5.5 ± 0.4 a |
| N | 24.9 ± 2.4 b | 253.1 ± 18.9 b | 66.1 ± 1.3 b | 126.6 ± 7.9 b | 17.5 ± 0.8 b | 5.2 ± 0.1 b | |
| P | 25.9 ± 0.8 a | 271.7 ± 16.4 a | 34.1 ± 1.0 a | 98.1 ± 4.4 a | 14.9 ± 0.9 a | 5.6 ± 0.3 a | |
| NP | 25.3 ± 0.9 a | 247.9 ± 9.8 a | 45.1 ± 4.7 c | 113.0 ± 4.7 b | 16.0 ± 0.8 b | 5.6 ± 0.3 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarif, N.; Khan, A.; Wang, Q. Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China. Forests 2020, 11, 1274. https://doi.org/10.3390/f11121274
Zarif N, Khan A, Wang Q. Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China. Forests. 2020; 11(12):1274. https://doi.org/10.3390/f11121274
Chicago/Turabian StyleZarif, Nowsherwan, Attaullah Khan, and Qingcheng Wang. 2020. "Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China" Forests 11, no. 12: 1274. https://doi.org/10.3390/f11121274