Inheritance and Expressivity of Neoplasm Trait in Crosses between the Domestic Pea (Pisum sativum subsp. sativum) and Tall Wild Pea (Pisum sativum subsp. elatius)

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Screening for Pea Weevil
2.3. Plant Sowing and Growing
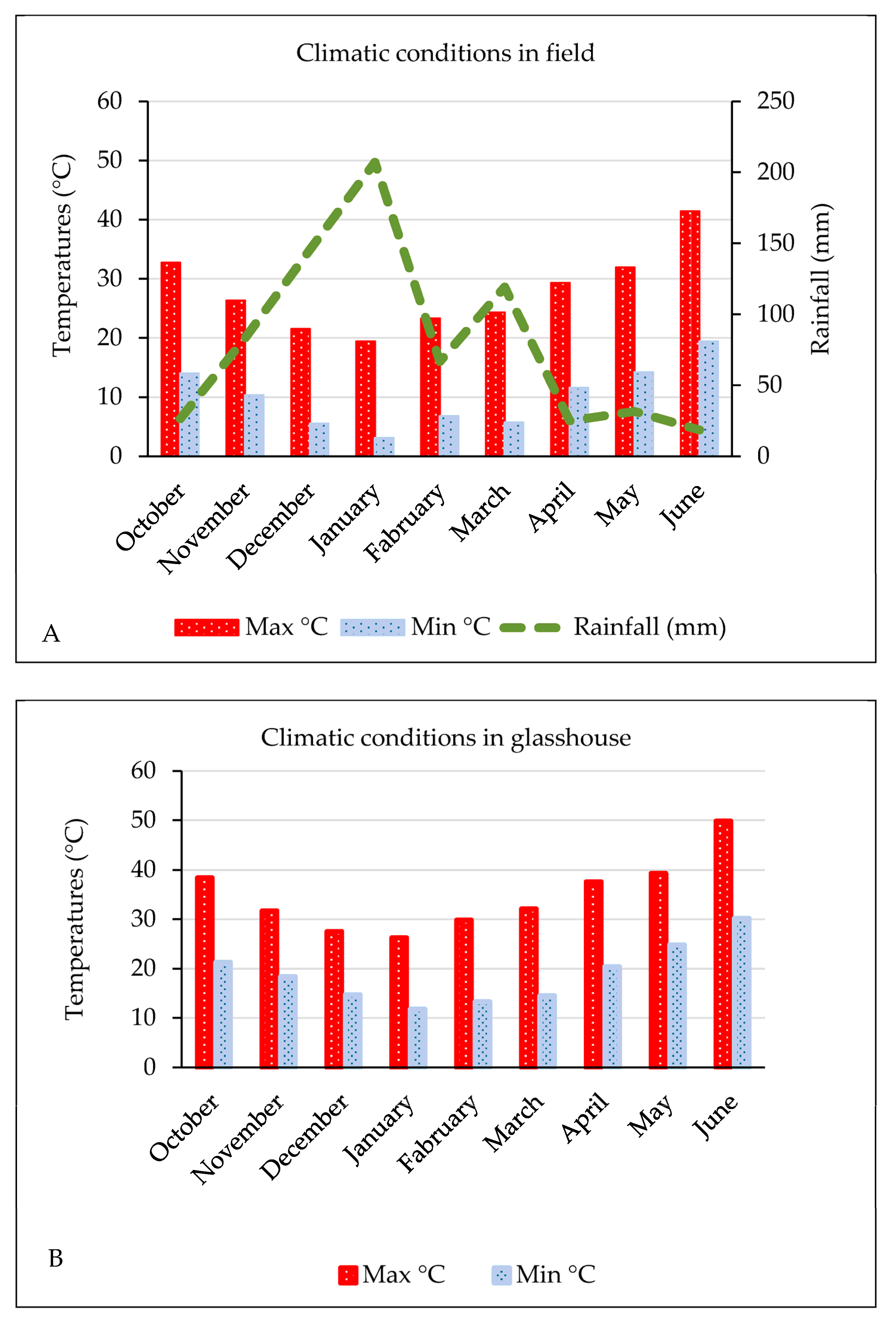
2.4. Climatic Conditions in Field and Glasshouse
2.5. Evaluation of Neoplasm
2.6. Expressivity of Neoplasm Trait
2.7. Plant Crosses and Progeny
2.8. Data Analysis
3. Results
3.1. Relationships between Neoplasm and Pea Weevil
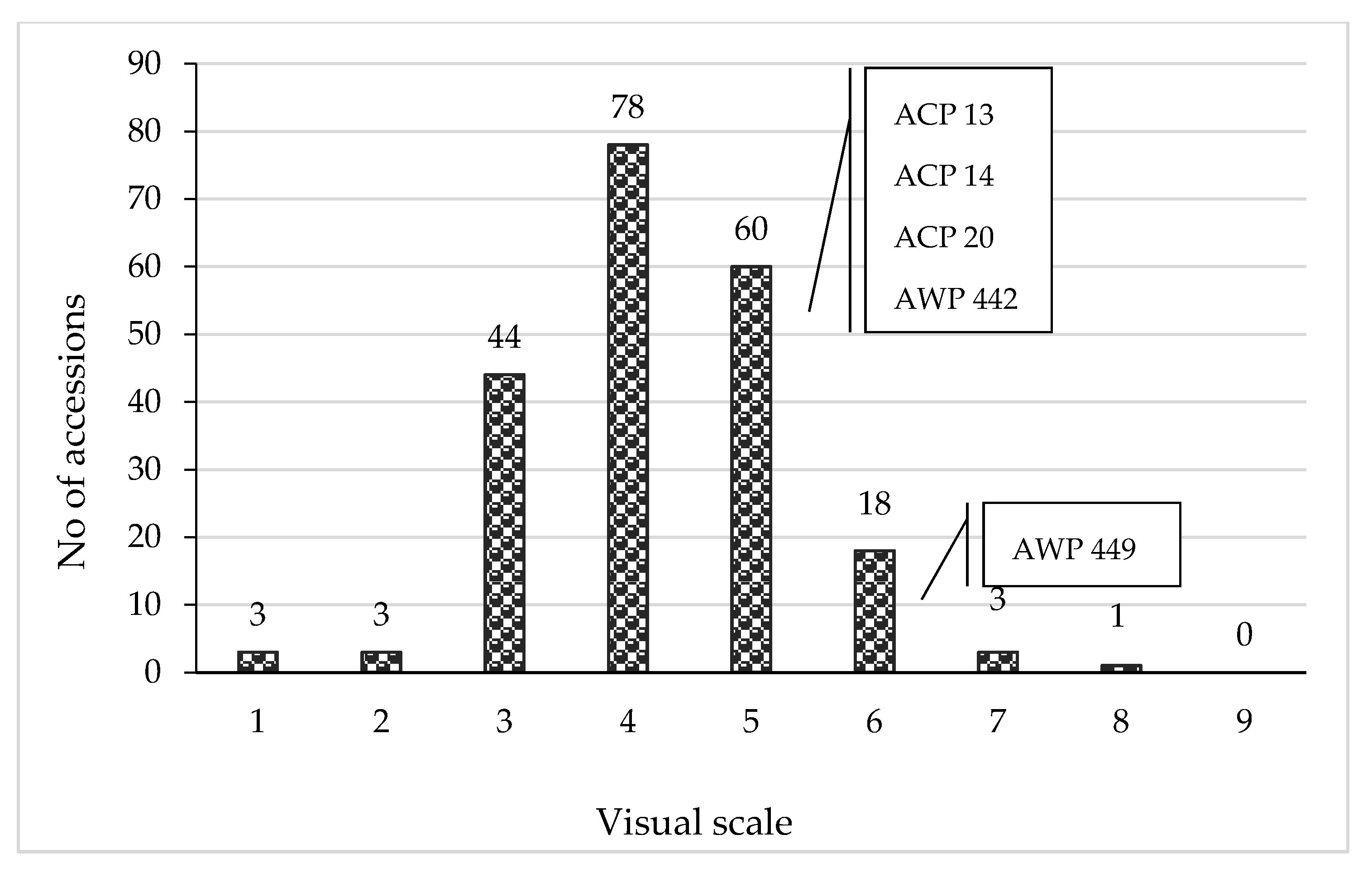
3.2. Expressivity of Neoplasm
3.3. Inheritance of Neoplasm in P. sativum subsp. elatius
3.4. Relationships between Neoplasm and Climatic Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Davis, P.H. Flora Turkey East Agean Islands 3; University of Edinburgh: Edinburgh, UK, 1970; pp. 370–372. [Google Scholar]
- Maxted, N.; Ambrose, N. Peas (Pisum L.) . In Plant Genetic Resources of Legumes in the Mediterranean; Maxted, N., Bennett, S.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Chapter 10; pp. 181–190. [Google Scholar]
- Ladizinsky, G.; Abbo, S. The Pisum Genus. In the Search for Wild Relatives of Cool Season Legumes; Springer: Cham, Switzerland, 2015; pp. 55–69. [Google Scholar]
- Smykal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.H.; Flavell, A.J.; Ford, R.; Hybl, M.I.; Macas, J.; Neumann, P.; et al. Pea (Pisum sativum L.) in the genomic era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Smykal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef] [Green Version]
- Trneny, O.; Brus, J.; Hradilova, I.; Rathore, A.; Das, R.R.; Kopecky, P.; Coyne, C.J.; Reeves, P.; Richards, C.; Smykal, P. Molecular evidence for two domestication events in the pea crop. Genes 2018, 9, 535. [Google Scholar] [CrossRef] [Green Version]
- Smykal, P.; Trneny, O.; Brus, J.; Hanacek, P.; Rathore, A.; Roma, R.D.; Pechanec, V.; Duchoslav, M.; Bhattacharyya, D.; Bariotakis, M.; et al. Genetic structure of wild pea (Pisum sativum subsp. elatius) populations in the northern part of the Fertile Crescent reflects moderate cross-pollination and strong effect of geographic but not environmental distance. PLoS ONE 2018, 13, e0194056. [Google Scholar]
- Yang, C.; Fan, Z.; Chai, Q. Agronomic and economic benefits of pea/maize intercropping systems in relation to N fertilizer and maize density. Agronomy 2018, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Shiwakoti, S.; Zheljazkov, V.D.; Gollany, H.T.; Xing, B.; Kleber, M. Micronutrient Concentrations in Soil and Wheat Decline by Long-Term Tillage and Winter Wheat–Pea Rotation. Agronomy 2019, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Andersen, B.J.; Samarappuli, D.P.; Wick, A.; Berti, M.T. Faba bean and pea can provide late-fall forage grazing without affecting maize yield the following season. Agronomy 2020, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Fan, Z.; Coulter, J.A.; Yin, W.; Hu, F.; Yu, A.; Fan, H.; Chai, Q. High Maize Density Alleviates the Inhibitory Effect of Soil Nitrogen on Intercropped Pea. Agronomy 2020, 10, 248. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Food and Agriculture Organization Statistical Databases. Available online: http://www.fao.org/faostat/en/ (accessed on 17 July 2020).
- Duke, J.A. Hand Book of Legumes of World Economic Importance; Plenum Press: New York, NY, USA, 1981; pp. 99–265. [Google Scholar]
- Nuttall, V.W.; Lyall, L.H. Inheritance of neoplastic pod in the pea. J. Hered. 1964, 55, 184–186. [Google Scholar] [CrossRef]
- Dodds, K.S.; Matthews, P. Neoplastic pod in the pea. J. Hered. 1966, 57, 83–85. [Google Scholar] [CrossRef]
- Berdnikov, V.A.; Trusov, Y.A.; Bogdanova, V.S.; Kosterin, O.E.; Rozov, S.M.; Nedelkina, S.V.; Nikulina, Y.N. The neoplastic pod gene (Np) may be a factor for resistance to the pest Bruchus pisorum L. Pisum Genet. 1992, 24, 37–39. [Google Scholar]
- Doss, R.P.; Oliver, J.E.; Proebsting, W.M.; Potter, S.W.; Kuy, S.; Clement, S.L.; Willianmson, R.T.; Carney, J.R.; DeVilbiss, E.D. Bruchins: Insect-derived plant regulators that stimulate neoplasm formation. Proc. Natl. Acad. Sci. USA 2000, 97, 6218–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teshome, A.; Bryngelsson, T.; Mendesil, E.; Marttila, S.; Geleta, M. Enhancing neoplasm expression in field pea (Pisum sativum) via intercropping and its significance to pea weevil (Bruchus pisorum) management. Front. Plant Sci. 2016, 7, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, J.; Fleming, E.N. The structure and development of a genetic tumour of the pea. Protoplasma 1973, 76, 315–325. [Google Scholar] [CrossRef]
- Esen, A.; Sari, H.; Erler, F.; Adak, A.; Sari, D.; Eker, T.; Canci, H.; Ikten, C.; Kahraman, A.; Toker, C. Screening and selection of accessions in the genus Pisum L. for resistance to pulse beetle (Callosobruchus chinensis L.). Euphytica 2019, 4, 82. [Google Scholar] [CrossRef]
- Teshome, A.; Mendesil, E.; Geleta, M.; Andargie, D.; Anderson, P.; Rämert, B.; Seyoum, E.; Hillbur, Y.; Dagne, K.; Bryngelsson, T. Screening the primary gene pool of field pea (Pisum sativum L. subsp. sativum) in Ethiopia for resistance against pea weevil (Bruchus pisorum L.). Genet. Resour. Crop Evol. 2015, 62, 525–538. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Carrillo-Perdomo, E.; Flores, F.; Rubiales, D. Identification and multi-environment validation of resistance to pea weevil (Bruchus pisorum) in Pisum germplasm. J. Pest Sci. 2018, 91, 505–514. [Google Scholar] [CrossRef]
- Koseoglu, K.; Adak, A.; Sari, D.; Sari, H.; Ceylan, F.O.; Toker, C. Transgressive segregations for yield criteria in reciprocal interspecific crosses between Cicer arietinum L. and C. reticulatum Ladiz. Euphytica 2017, 213, 116. [Google Scholar] [CrossRef]
- Kivrak, K.G.; Eker, T.; Sari, H.; Sari, D.; Akan, K.; Aydinoglu, B.; Catal, M.; Toker, C. Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.). Agronomy 2020, 10, 901. [Google Scholar] [CrossRef]
- Yasar, M.; Ceylan, F.O.; Ikten, C.; Toker, C. Comparison of expressivity and penetrance of the double podding trait and yield components based on reciprocal crosses of kabuli and desi chickpeas (Cicer arietinum L.). Euphytica 2014, 3, 331–339. [Google Scholar] [CrossRef]
- Steel, R.G.; Torrie, J.H. Principles and Procedures of Statistics, a Biometrical Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Hardie, D.C.; Baker, G.J.; Marshall, D.R. Field screening of Pisum accessions to evaluate their susceptibility to the pea weevil (Coleoptera: Bruchidae). Euphytica 1995, 84, 155–161. [Google Scholar] [CrossRef]
- Clement, S.L.; Hardie, D.C.; Elberson, L.R. Variation among accessions of Pisum fulvum for resistance to pea weevil. Crop Sci. 2002, 42, 2167–2173. [Google Scholar] [CrossRef] [Green Version]
- Aryamanesh, N.; Byrne, O.; Hardie, D.C.; Khan, T.; Siddique, K.H.M.; Yan, G. Large-scale density-based screening for pea weevil resistance in advanced backcross lines derived from cultivated field pea (Pisum sativum) and Pisum fulvum. Crop Pasture Sci. 2012, 63, 612–618. [Google Scholar] [CrossRef]
- Mendesil, E.; Ramert, B.; Marttila, S.; Hillbur, Y.; Anderson, P. Oviposition preference of pea weevil, Bruchus pisorum L. among host and non-host plants and its implication for pest management. Front. Plant Sci. 2016, 6, 1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoad, B.; Matthews, P. Neoplasms of the pea pod. Chromosom. Today 1969, 2, 126–131. [Google Scholar]
- Doss, R.P. Treatment of pea pods with Bruchin B results in up-regulation of a gene similar to MtN19. Plant Physiol. Biochem. 2005, 43, 225–231. [Google Scholar] [CrossRef]
- Doss, R.P.; Proebsting, W.M.; Potter, S.W.; Clement, S.L. Response of Np mutant of pea (Pisum sativum L.) to pea weevil (Bruchus pisorum L.) oviposition and extracts. J. Chem. Ecol. 1995, 21, 1–10. [Google Scholar] [CrossRef]
- Cooper, L.D.; Doss, R.P.; Price, R.; Peterson, K.; Oliver, J.E. Application of Bruchin B to pea pods results in the up-regulation of CYP93C18, a putative isoflavone synthase gene, and an increase in the level of pisatin, an isoflavone phytoalexin. J. Exp. Bot. 2005, 56, 1229–1237. [Google Scholar] [CrossRef]
- Ketter, A.P. Cellular Events Conditioned by the Np Gene of Pisum sativum L. in Response to Reduced UV Light, Weevil Oviposition, and Bruchins. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2007. [Google Scholar]
- Reddy, G.V.; Sharma, A.; Gadi, R.L. Biology, ecology, and management of the pea weevil (Coleoptera: Chrysomelidae). Ann. Entomol. Soc. Am. 2018, 111, 161–171. [Google Scholar] [CrossRef]
- Aznar-Fernandez, T.; Cimmino, A.; Masi, M.; Rubiales, D.; Evidente, A. Antifeedant activity of long-chain alcohols, and fungal and plant metabolites against pea aphid (Acyrthosiphon pisum) as potential biocontrol strategy. Nat. Prod. Res. 2019, 33, 2471–2479. [Google Scholar] [CrossRef]
- WOS. Web of Science. Available online: http://apps.webofknowledge.com/ (accessed on 18 August 2020).
- Weeden, N.F.; Ellis, T.H.N.; Timmerman-Vaughan, G.M.; Swiecicki, W.K.; Rozov, S.M.; Berdnikov, V.A. A consensus linkage map for Pisum sativum. Pisum Genet. 1998, 30, 1–4. [Google Scholar]
- Prioul, S.; Frankewitz, A.; Deniot, G.; Morin, G.; Baranger, A. Mapping of quantitative trait loci for partial resistance to Mycosphaerella pinodes in pea (Pisum sativum L.), at the seedling and adult plant stages. Theor. Appl. Genet. 2004, 108, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Saxena, M.C. Breeding for Stress Tolerance in Cool-Season Food Legumes; Wiley: Chichester, UK, 1993; p. 52. [Google Scholar]
- Ali, S.M.; Sharma, B.; Ambrose, M.J. Current status and future strategy in breeding pea to improve resistance to biotic and abiotic stresses. In Expanding the Production and Use of Cool Season Food Legumes; Springer: Dordrecht, The Netherlands, 1994; pp. 540–558. [Google Scholar]
- Toker, C.; Uzun, B.; Ceylan, F.O.; Ikten, C. Chickpea. In Alien Gene Transfer in Crop Plants; Springer: New York, NY, USA, 2014; Volume 2, pp. 121–151. [Google Scholar]
- Clement, S.L.; McPhee, K.E.; Elberson, L.R.; Evans, M.A. Pea weevil, Bruchus pisorum L. (Coleoptera: Bruchidae), resistance in Pisum sativum × Pisum fulvum interspecific crosses. Plant Breed. 2009, 128, 478–485. [Google Scholar] [CrossRef]
- Byrne, O.M.; Hardie, D.C.; Khan, T.; Yan, G. Genetic analysis of pod and seed resistance to pea weevil in a Pisum sativum x P. fulvum interspecific cross. Aust. J. Agric. Res. 2008, 59, 854–862. [Google Scholar] [CrossRef]
- Aryamanesh, N.; Zeng, Y.; Byrne, O.; Hardie, D.C.; Al-Subhi, A.M.; Khan, T.; Siddique, K.H.M.; Yan, G. Identification of genome regions controlling cotyle don, pod wall/seed coat and pod wall resistance to pea weevil through QTL mapping. Theor. Appl. Genet. 2014, 127, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Fondevilla, S.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of a new gene for resistance to powdery mildew in Pisum fulvum, a wild relative of pea. Breed. Sci. 2007, 57, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Barilli, E.; Sillero, J.C.; Moral, A.; Rubiales, D. Characterization of resistance response of pea (Pisum spp.) against rust (Uromyces pisi). Plant Breed. 2009, 128, 665–670. [Google Scholar] [CrossRef]
- Barilli, E.; Satovic, Z.; Rubiales, D.; Torres, A.M. Mapping of quantitative trait loci controlling partial resistance against rust incited by Uromyces pisi (Pers.) Wint. in a Pisum fulvum L. intraspecific cross. Euphytica 2010, 175, 151–159. [Google Scholar] [CrossRef] [Green Version]
- McPhee, K.E.; Tullu, A.; Kraft, J.M.; Muehlbauer, F.J. Resistance to Fusarium wilt race 2 in the Pisum core collection. J. Am. Soc. Hortic. Sci. 1999, 124, 28–31. [Google Scholar] [CrossRef] [Green Version]
- Hance, S.T.; Grey, W.; Weeden, N.F. Identification of tolerance to Fusarium solani in Pisum sativum ssp. elatius. Pisum Genet. 2004, 36, 9–13. [Google Scholar]
- Wroth, J.M. Possible role of wild genotypes of Pisum spp. to enchance ascochyta blight resistance in pea. Aust. J. Exp. Agric. 1998, 38, 469–479. [Google Scholar] [CrossRef]
- Fondevilla, S.; Ávila, C.M.; Cubero, J.I.; Rubiales, D. Response to Mycosphaerella pinodes in a germplasm collection of Pisum spp. Plant Breed. 2005, 124, 313–315. [Google Scholar] [CrossRef]
- Carrillo, E.; Rubiales, D.; Pérez-de-Luque, A.; Fondevilla, S. Characterization of mechanisms of resistance against Didymella pinodes in Pisum spp. Eur. J. Plant Pathol. 2013, 135, 761–769. [Google Scholar] [CrossRef]
- Naim-Feil, E.; Toren, M.; Aubert, G.; Rubinstein, M.; Rosen, A.; Eshed, R.; Sherman, A.; Ophir, R.; Saranga, Y.; Abbo, S. Drought response and genetic diversity in Pisum fulvum, a wild relative of domesticated pea. Crop Sci. 2017, 57, 1145–1159. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies | Accessions | Landrace/Wild | Np in Field | Np in Glasshouse (Np/np) | Flower Color | 100-Seed Weight (g) | Tolerance to Cold | Resistance to Seed Beetle | Resistance to Pea Weevil |
|---|---|---|---|---|---|---|---|---|---|
| P. s. ssp. sativum | 1 ACP 13 | Landrace | No | np | Pink | 23.5 | Medium | Susceptible | Susceptible |
| P. s. ssp. sativum | ACP 14 | Landrace | No | np | Pink | 29.7 | Medium | Susceptible | Susceptible |
| P. s. ssp. sativum | ACP 20 | Landrace | No | np | White | 34.1 | Medium | Susceptible | Susceptible |
| P. s. ssp. sativum | ACP 773 | Landrace | No | np | White | 28.9 | * | * | * |
| P. s. ssp. elatius | 2 AWP 442 | Wild | No | Np | Lilac purple | 9.8 | Tolerant | Resistant | Susceptible |
| P. s. ssp. elatius | AWP 449 | Wild | No | Np | Lilac purple | 10.6 | Tolerant | Susceptible | Susceptible |
| P. s. ssp. elatius | AWP 451 | Wild | No | Np | Lilac purple | 11.2 | * | * | * |
| Score | Response to Pea Weevil | Damages on Seeds |
|---|---|---|
| 1 | Very Highly Resistant | Free from any seed damage after careful observation |
| 2 | Highly Resistant | Damage present in 1 to 10% of the seeds |
| 3 | Resistant | Damage present 11 to 20% of the seeds |
| 4 | Moderately Resistant | Damage present in 21 to 30% of the seeds |
| 5 | Less susceptible | Damage present in 31 to 40% of the seeds |
| 6 | Moderately susceptible | Damage present in 41 to 50% of the seeds |
| 7 | Susceptible | Damage present in 51 to 70% of the seeds |
| 8 | Highly susceptible | Damage present in 71 to 90% of the seeds |
| 9 | Very highly susceptible | Damage present in more than 91% of the seeds |
| Inter-Subspecific Crosses | Expected Ratio of Neoplasm | Observed Neoplasm Progeny | Expressivity (%) |
|---|---|---|---|
| ACP 13 (np) × AWP 442 (Np) | 3/4 (82.5) | 85 | 77.2 |
| ACP 773 (np) × AWP 451 (Np) | 3/4 (98.3) | 89 | 67.9 |
| Inter-Subspecific Crosses | Phenotype of F1s | F2 | ||||
|---|---|---|---|---|---|---|
| No of Plants | Observed | Expected | χ2 | p | ||
| ACP 13 × AWP 442 | Np | 85 | Np | 3:1 | 0.3 | 0.9-0.1 |
| 25 | np | |||||
| ACP 773 × AWP 451 | 89 | Np | 3:1 | 3.5 | 0.9-0.1 | |
| 42 | np | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sari, H.; Sari, D.; Eker, T.; Aydinoglu, B.; Canci, H.; Ikten, C.; Gokturk, R.S.; Zeybek, A.; Bakir, M.; Smykal, P.; et al. Inheritance and Expressivity of Neoplasm Trait in Crosses between the Domestic Pea (Pisum sativum subsp. sativum) and Tall Wild Pea (Pisum sativum subsp. elatius). Agronomy 2020, 10, 1869. https://doi.org/10.3390/agronomy10121869
Sari H, Sari D, Eker T, Aydinoglu B, Canci H, Ikten C, Gokturk RS, Zeybek A, Bakir M, Smykal P, et al. Inheritance and Expressivity of Neoplasm Trait in Crosses between the Domestic Pea (Pisum sativum subsp. sativum) and Tall Wild Pea (Pisum sativum subsp. elatius). Agronomy. 2020; 10(12):1869. https://doi.org/10.3390/agronomy10121869
Chicago/Turabian StyleSari, Hatice, Duygu Sari, Tuba Eker, Bilal Aydinoglu, Huseyin Canci, Cengiz Ikten, Ramazan S. Gokturk, Ahmet Zeybek, Melike Bakir, Petr Smykal, and et al. 2020. "Inheritance and Expressivity of Neoplasm Trait in Crosses between the Domestic Pea (Pisum sativum subsp. sativum) and Tall Wild Pea (Pisum sativum subsp. elatius)" Agronomy 10, no. 12: 1869. https://doi.org/10.3390/agronomy10121869