In Vitro Antidiabetic and Antioxidant Effects of Different Extracts of Catharanthus roseus and Its Indole Alkaloid, Vindoline


Abstract
:1. Introduction
2. Results
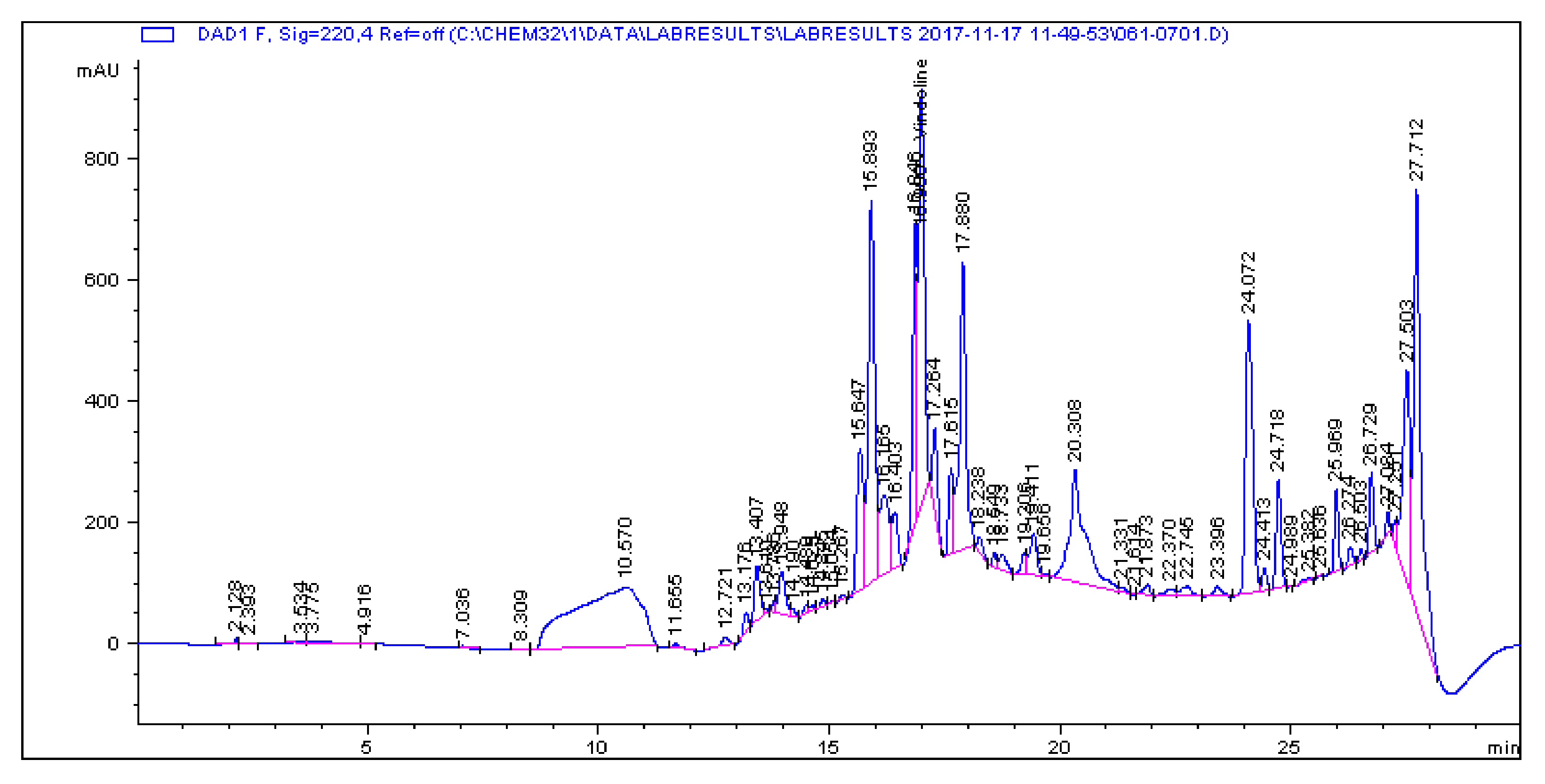
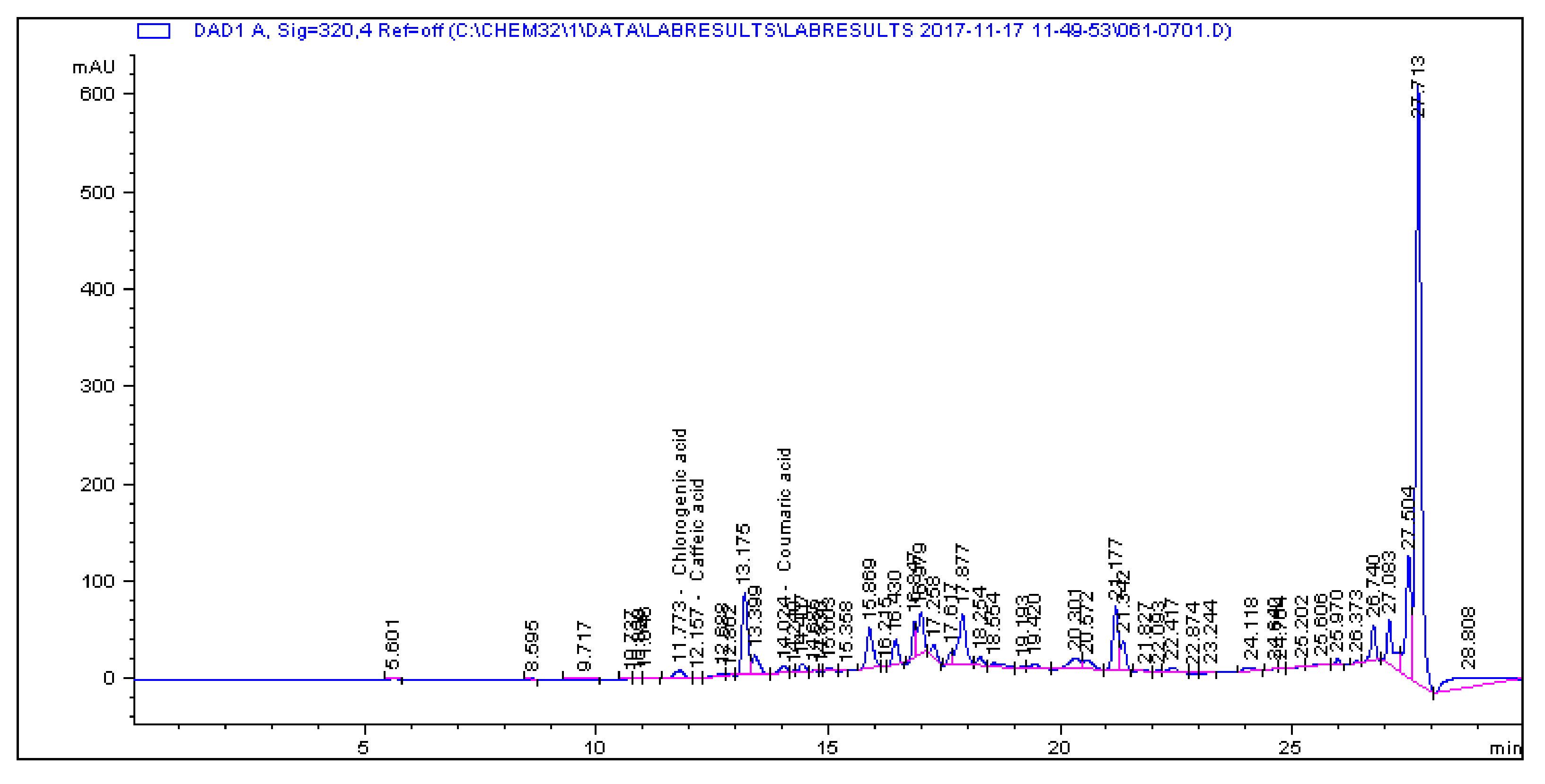
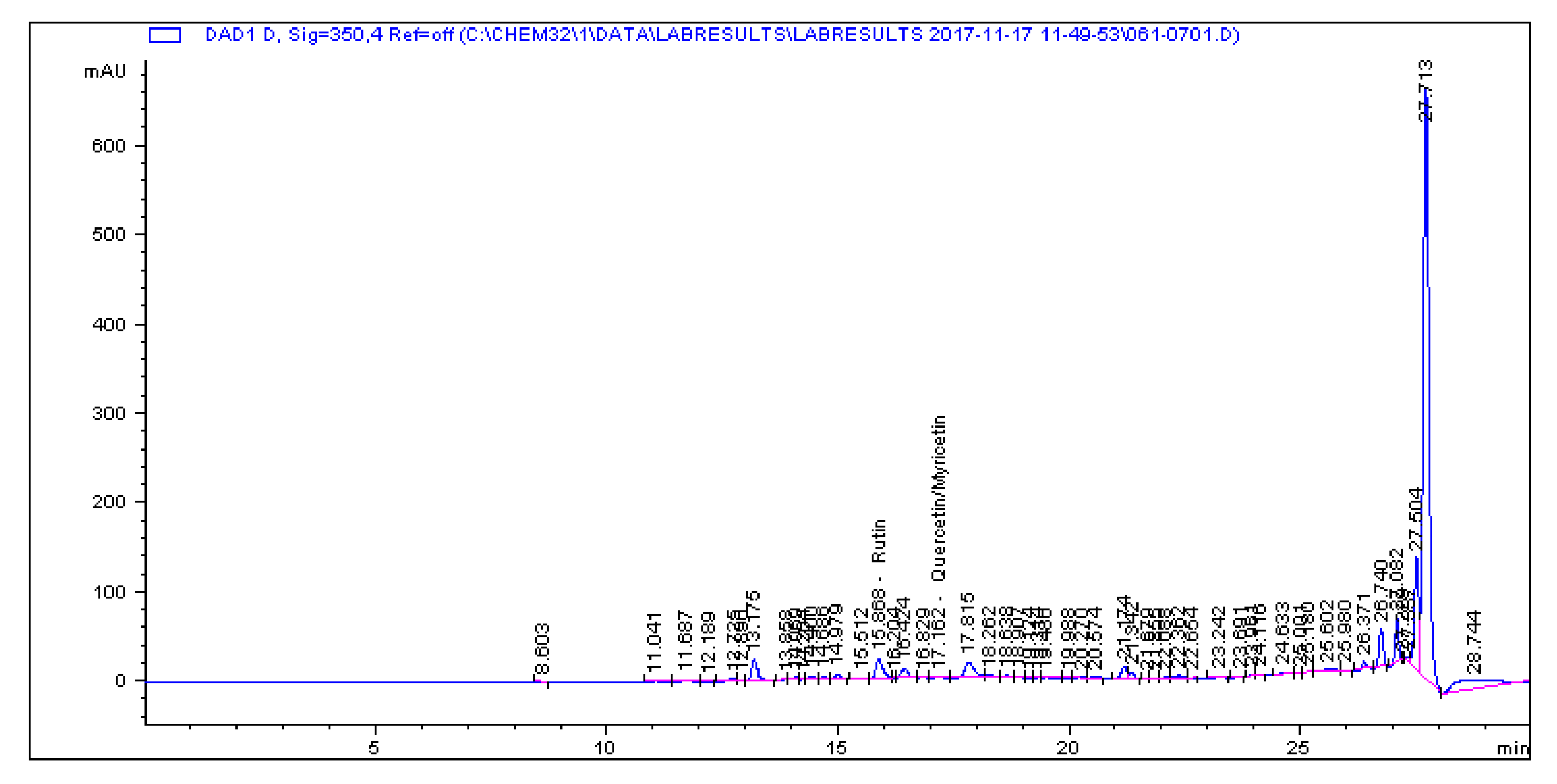
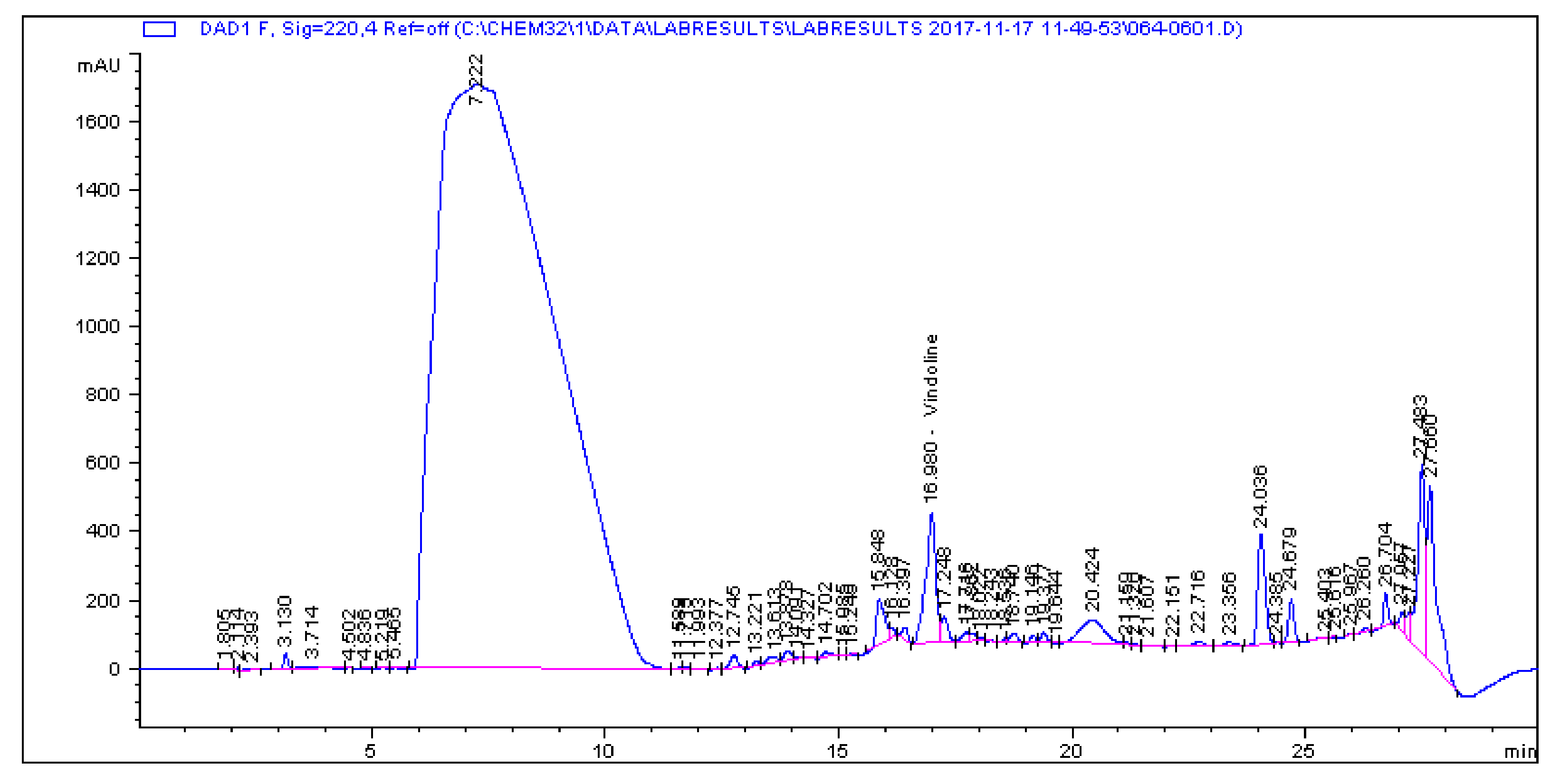
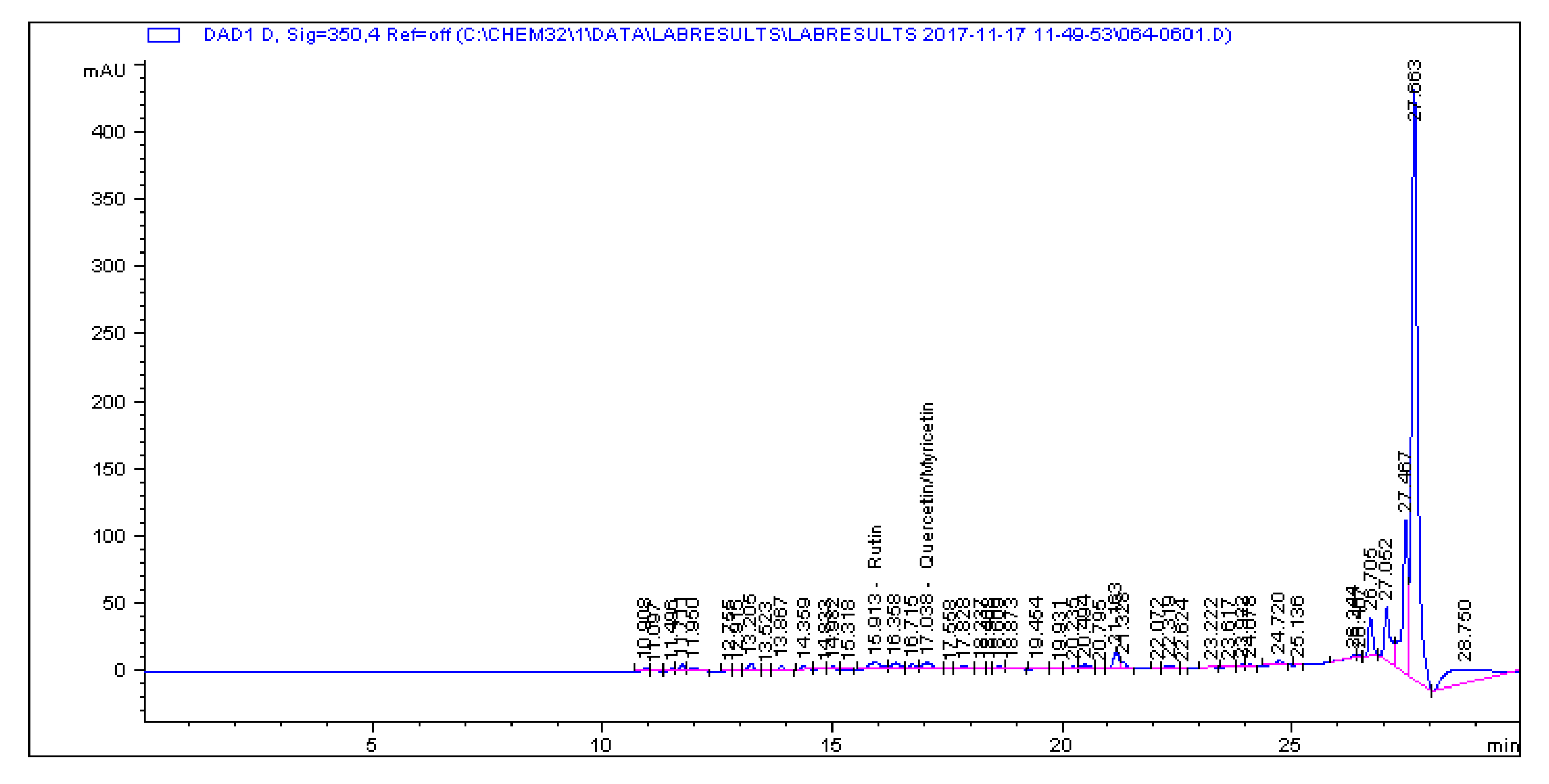
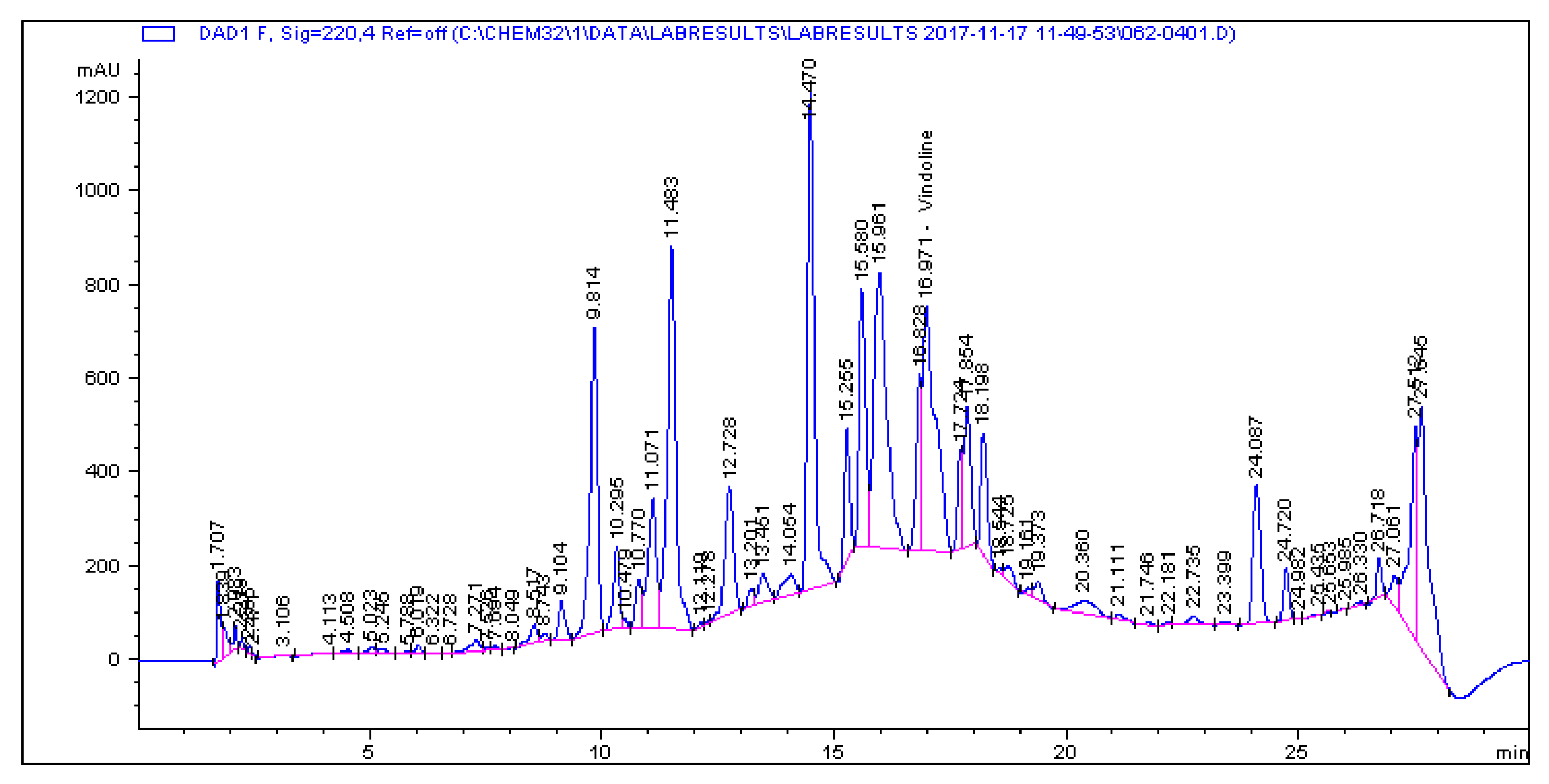
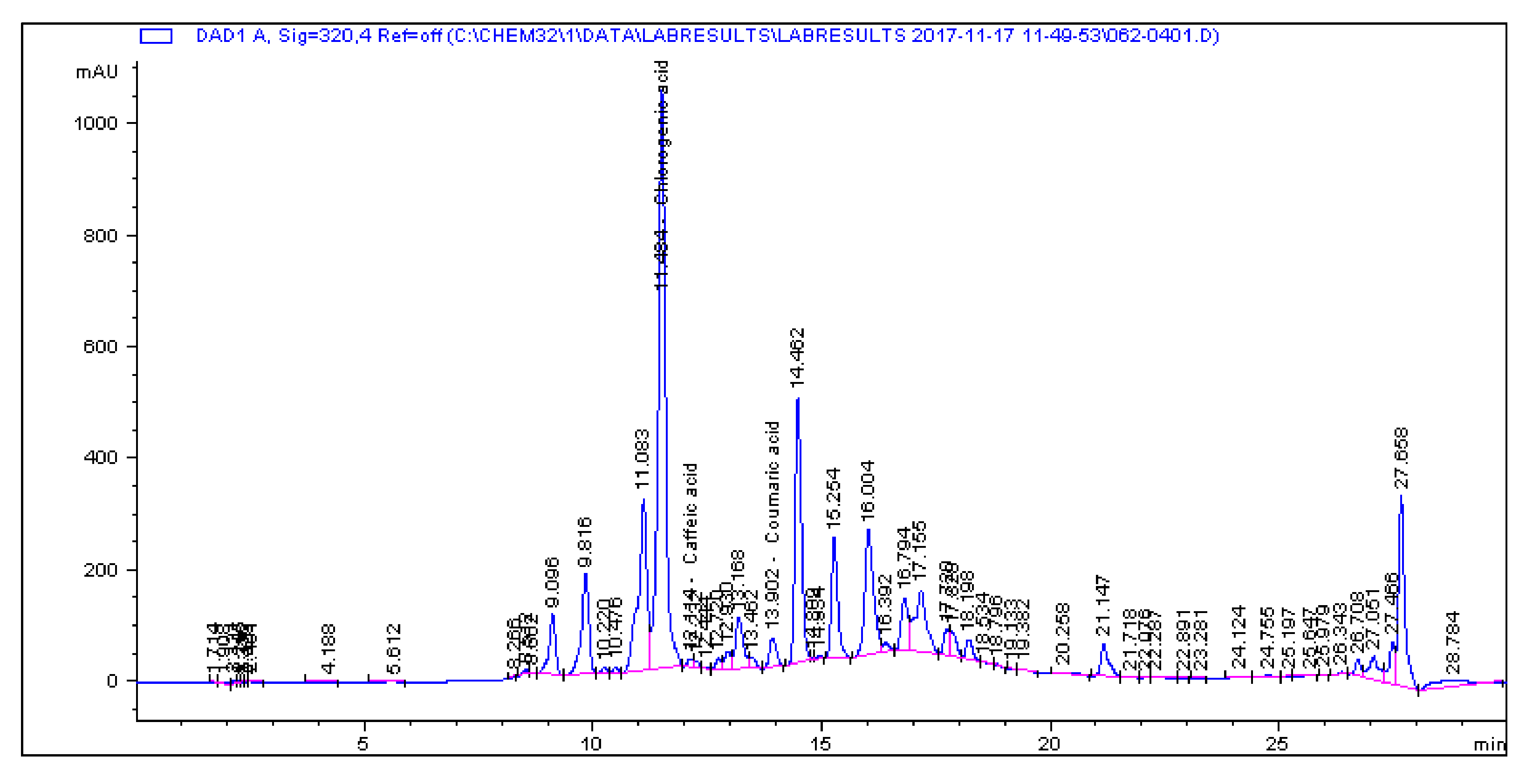
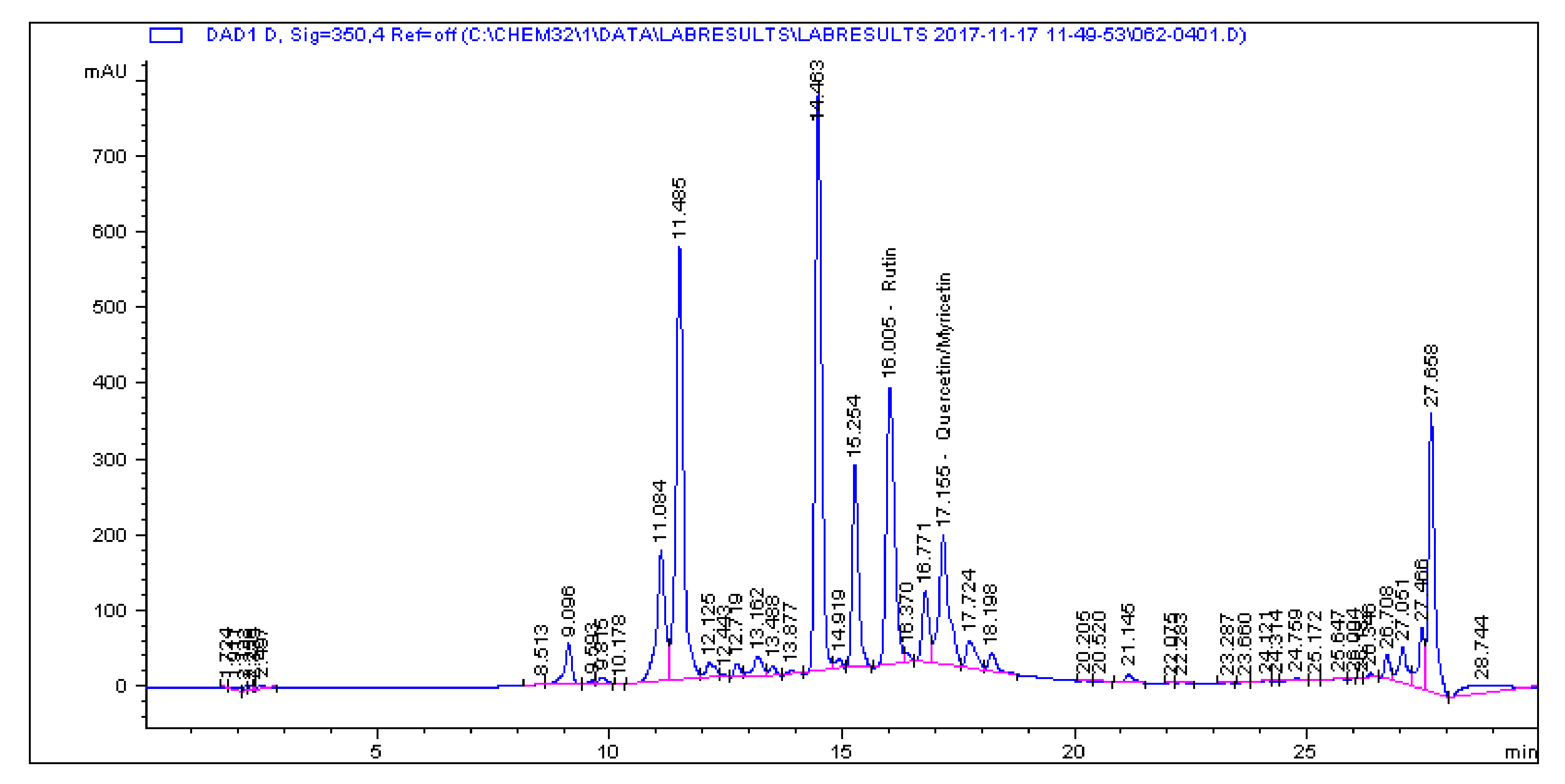
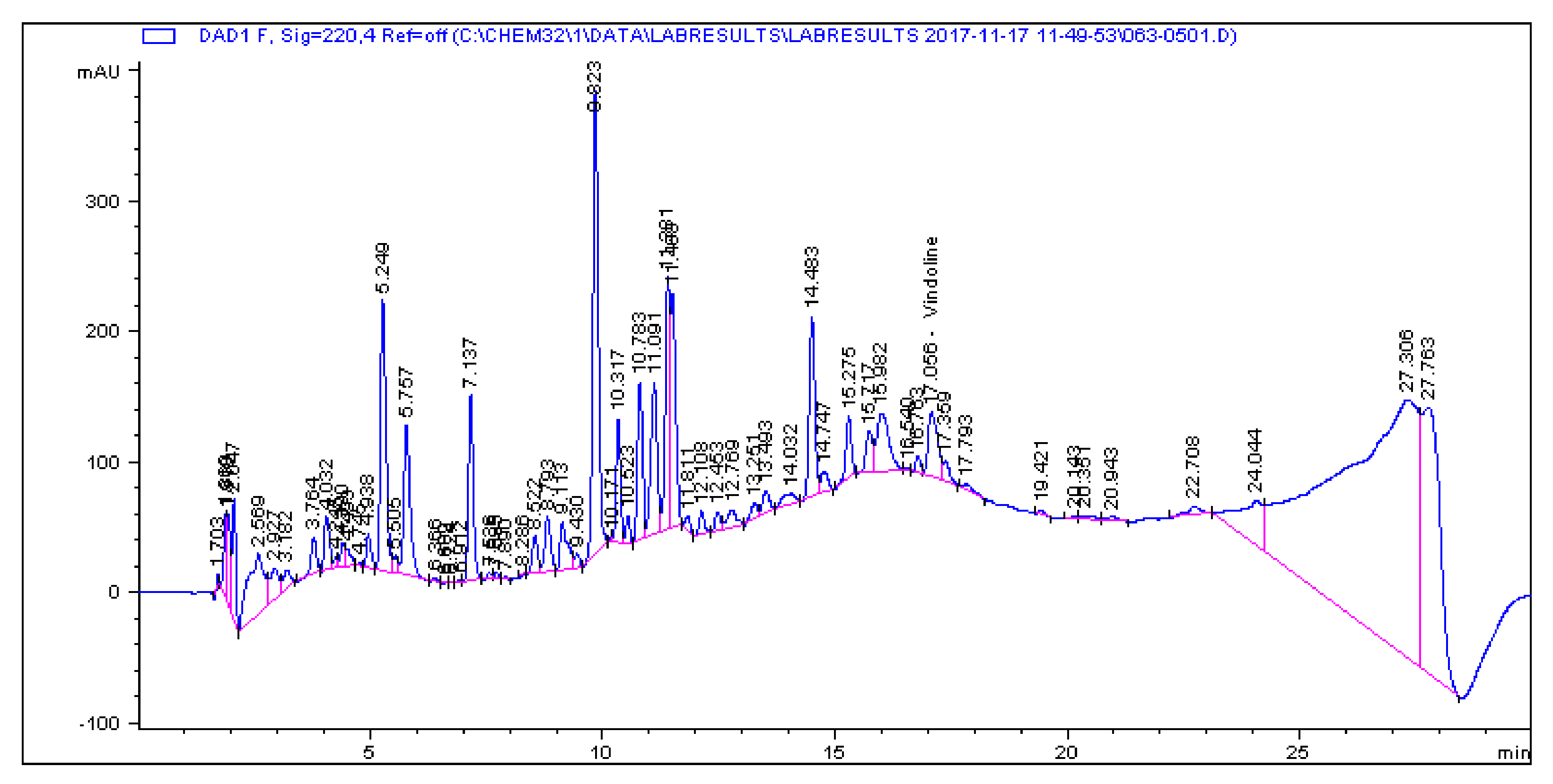
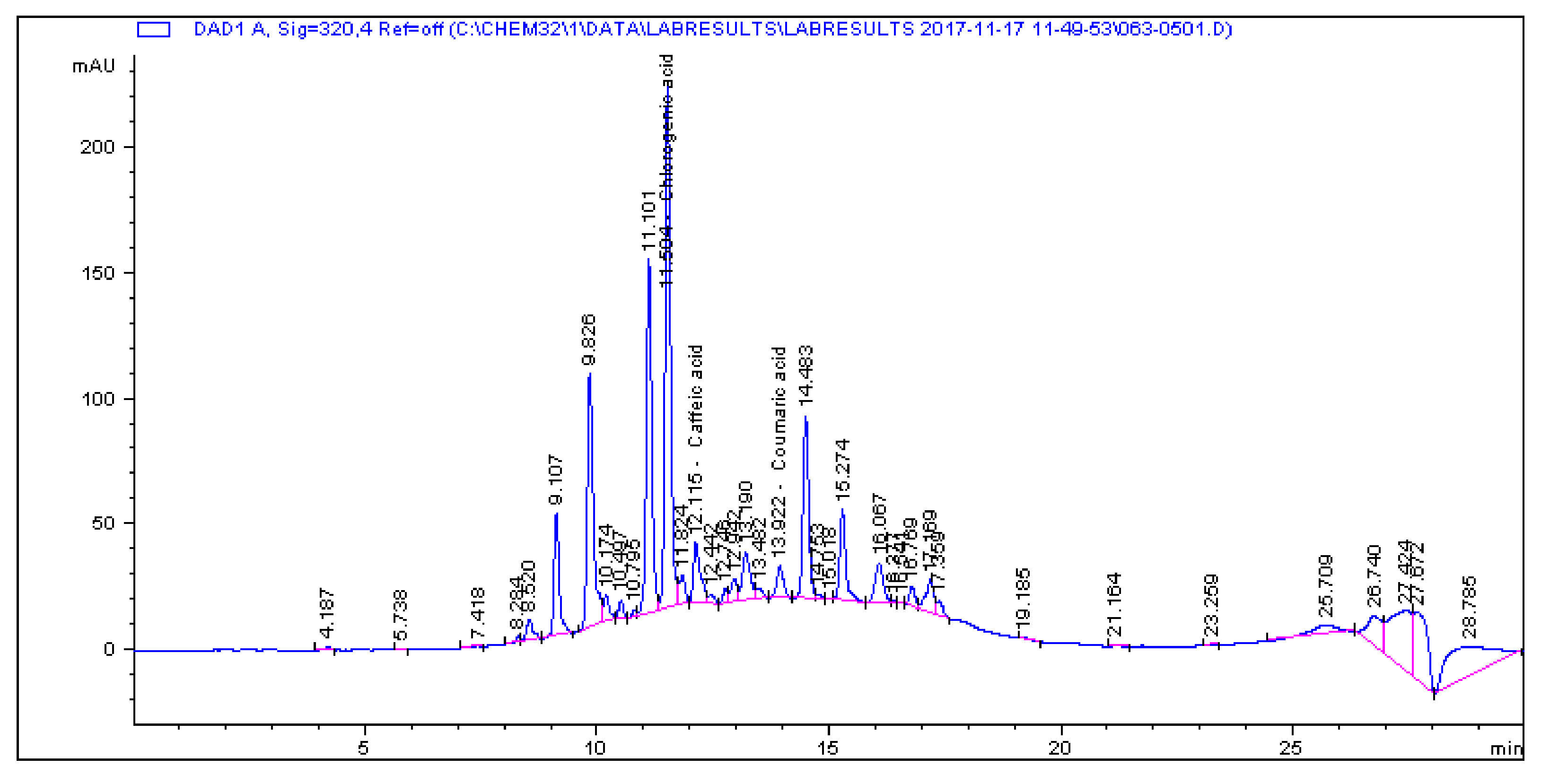
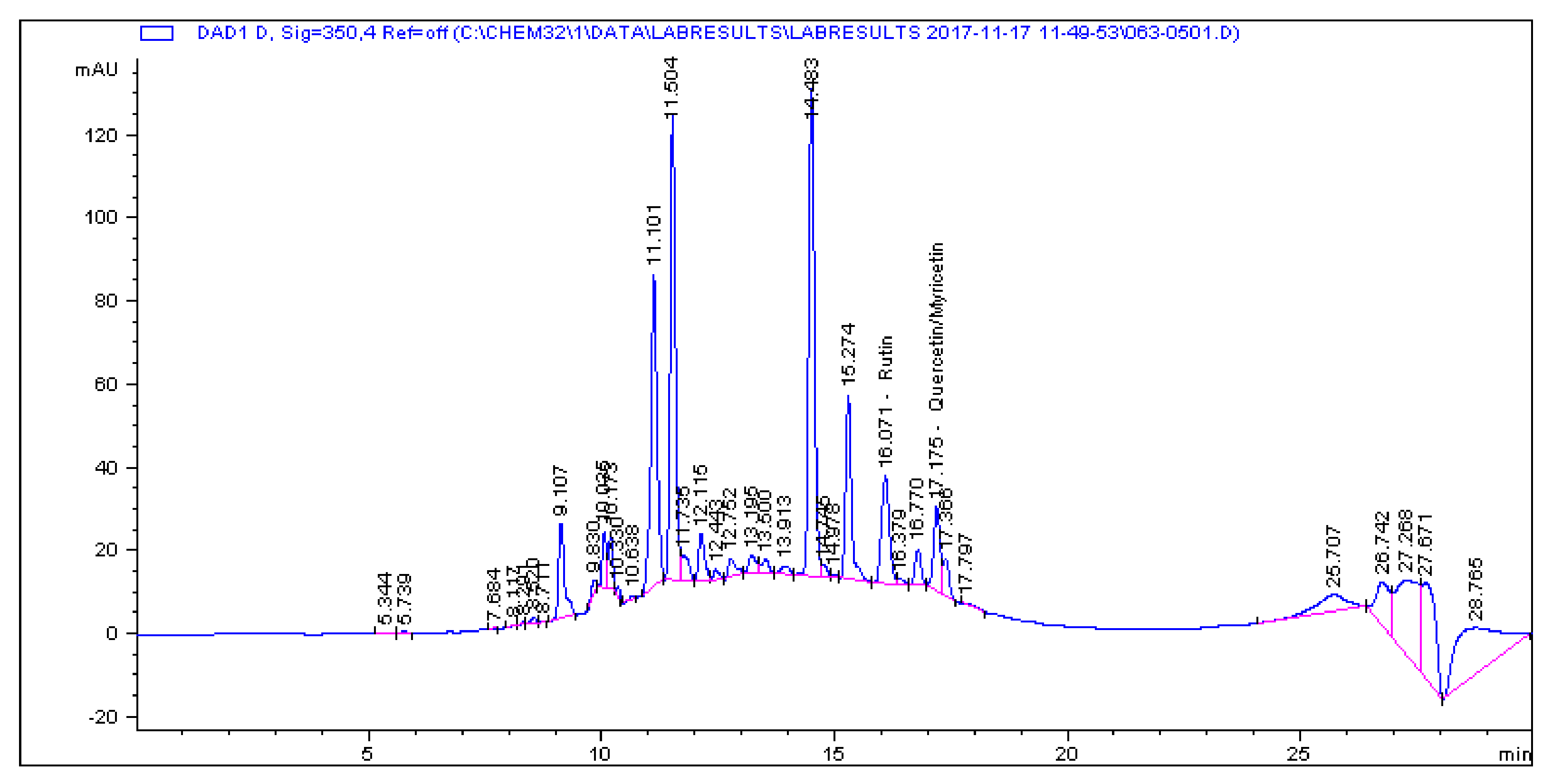
2.1. Determination and Quantification of Phenolic Compounds and Vindoline in C. roseus Extracts
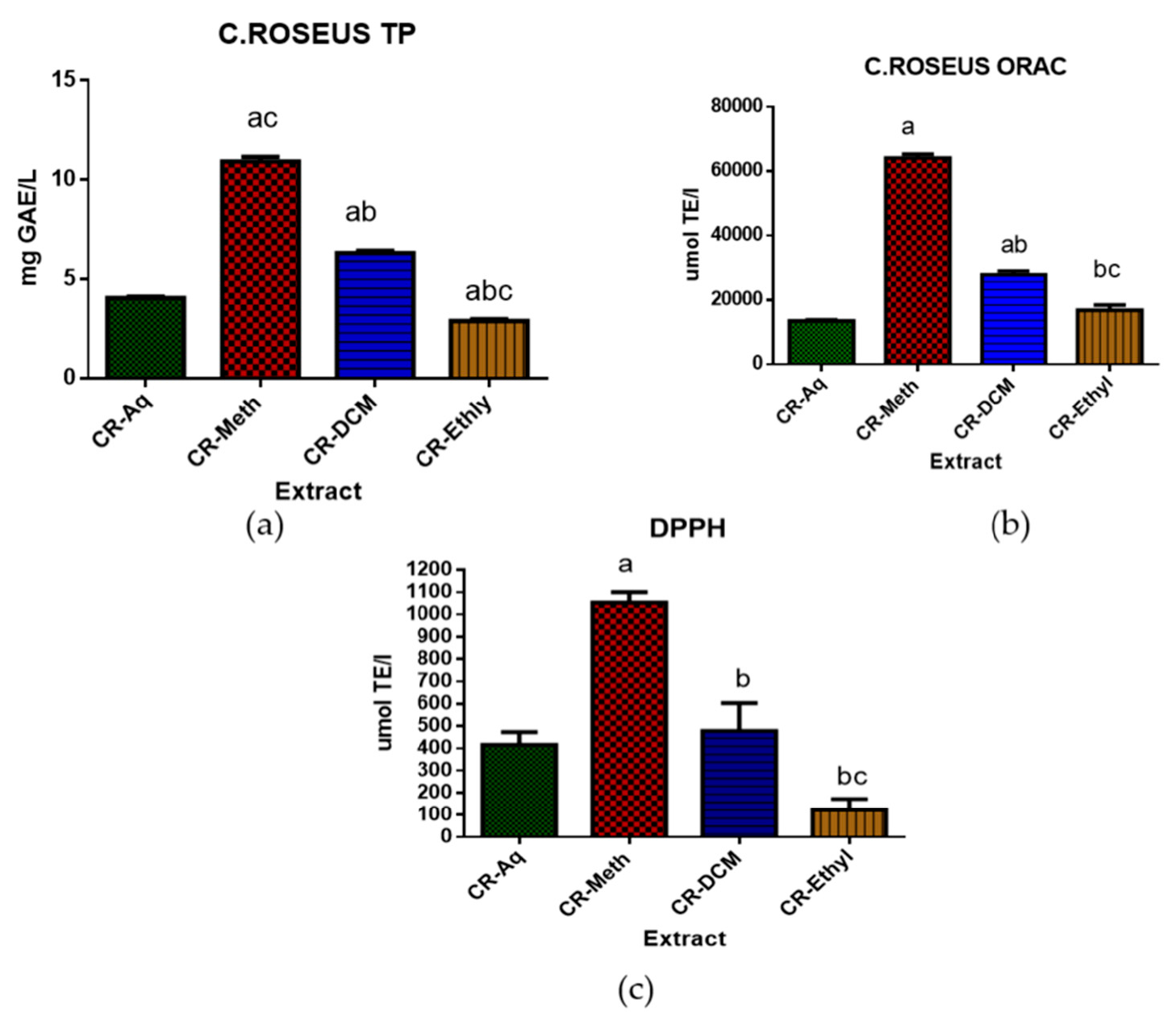
2.2. Total Polyphenolic and Antioxidant Assessment of C. roseus Extracts
2.3. Determination of Vindoline’s Antioxidant Capacity
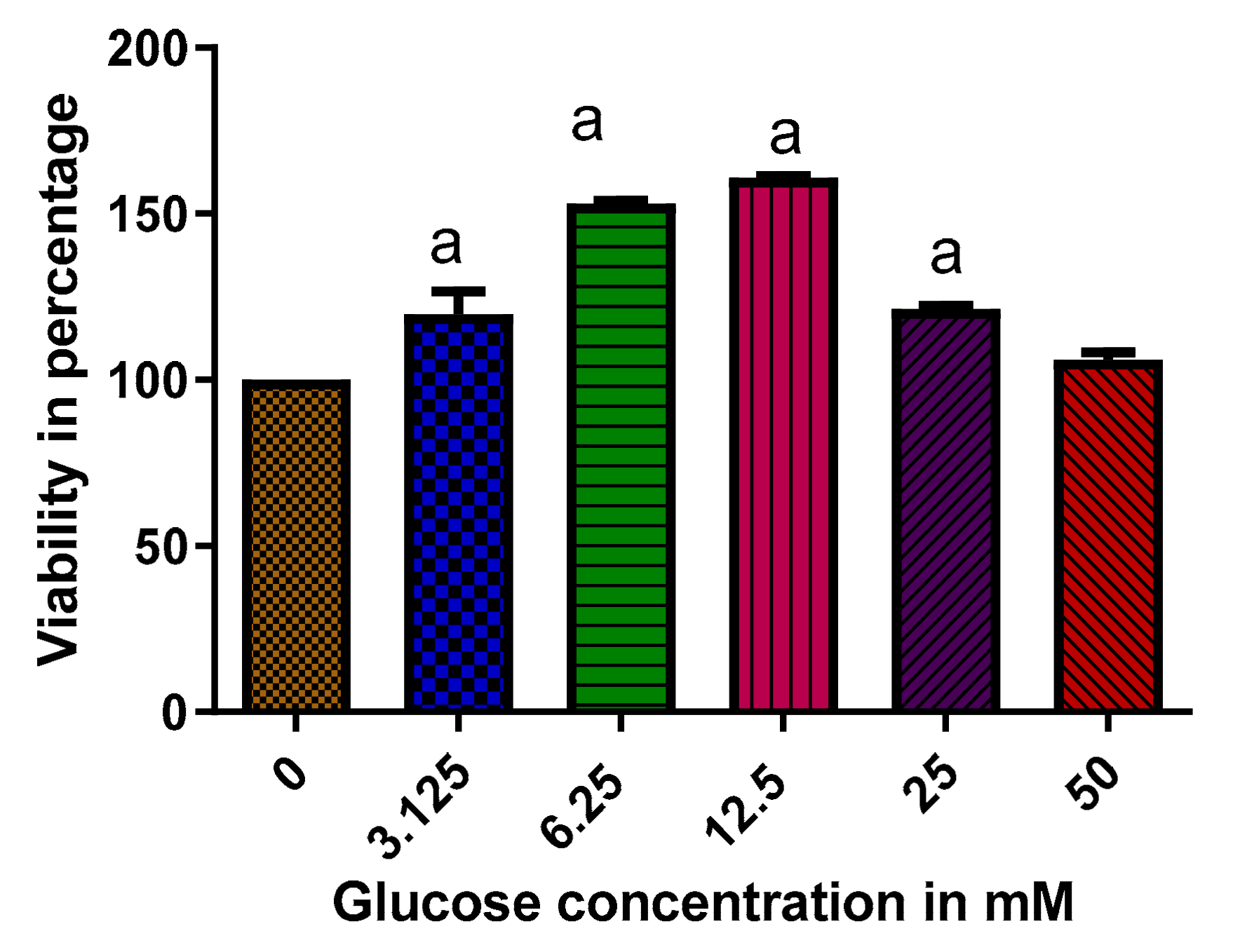
2.4. The Effect of High Glucose Concentration on the Viability of RIN-5F Cells
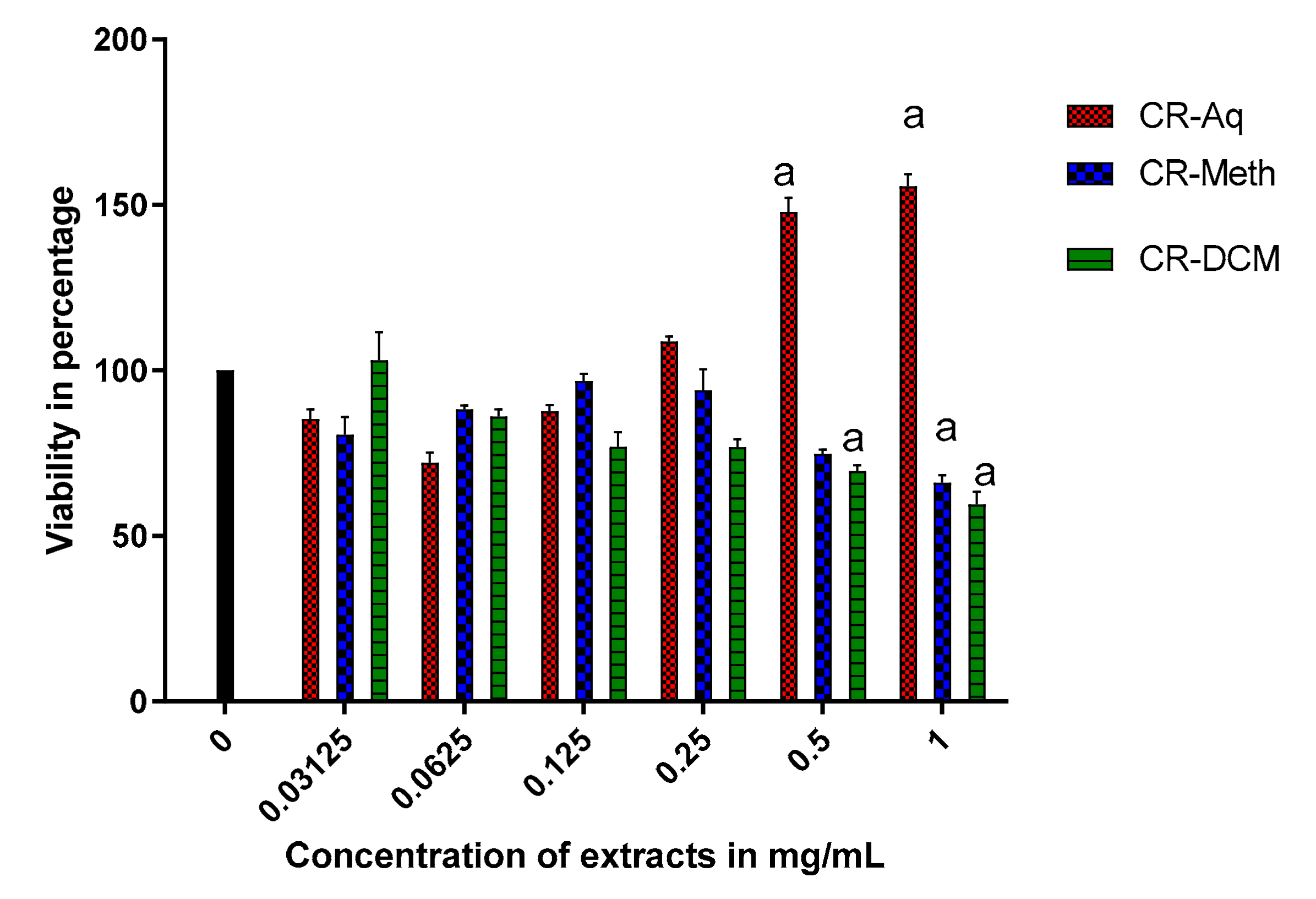
2.5. Effect of Different Extracts of C. roseus on the Cell Viability of RIN-5F Cells
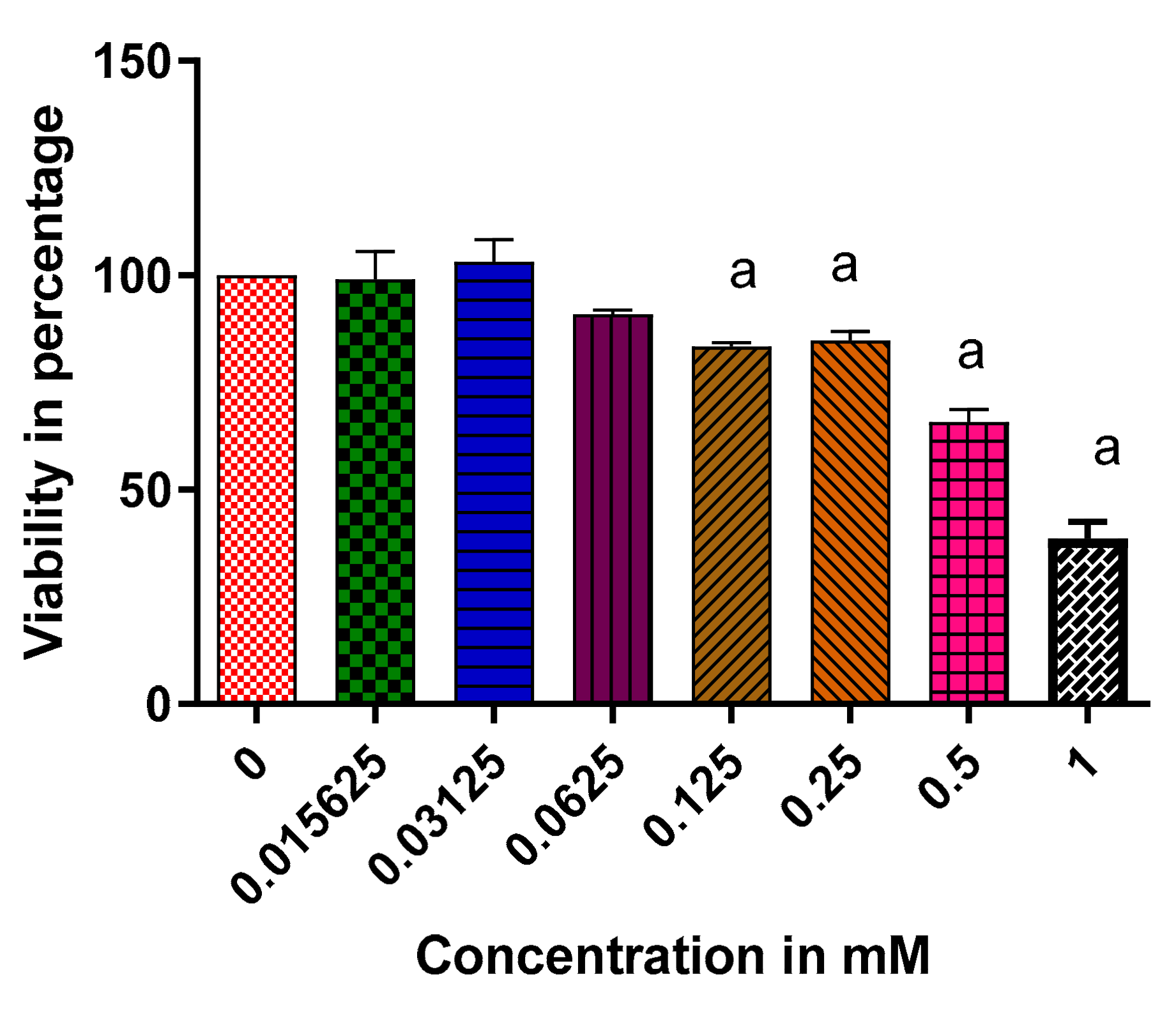
2.6. The Effect of Vindoline on the Cell Viability of RIN-5F Cells
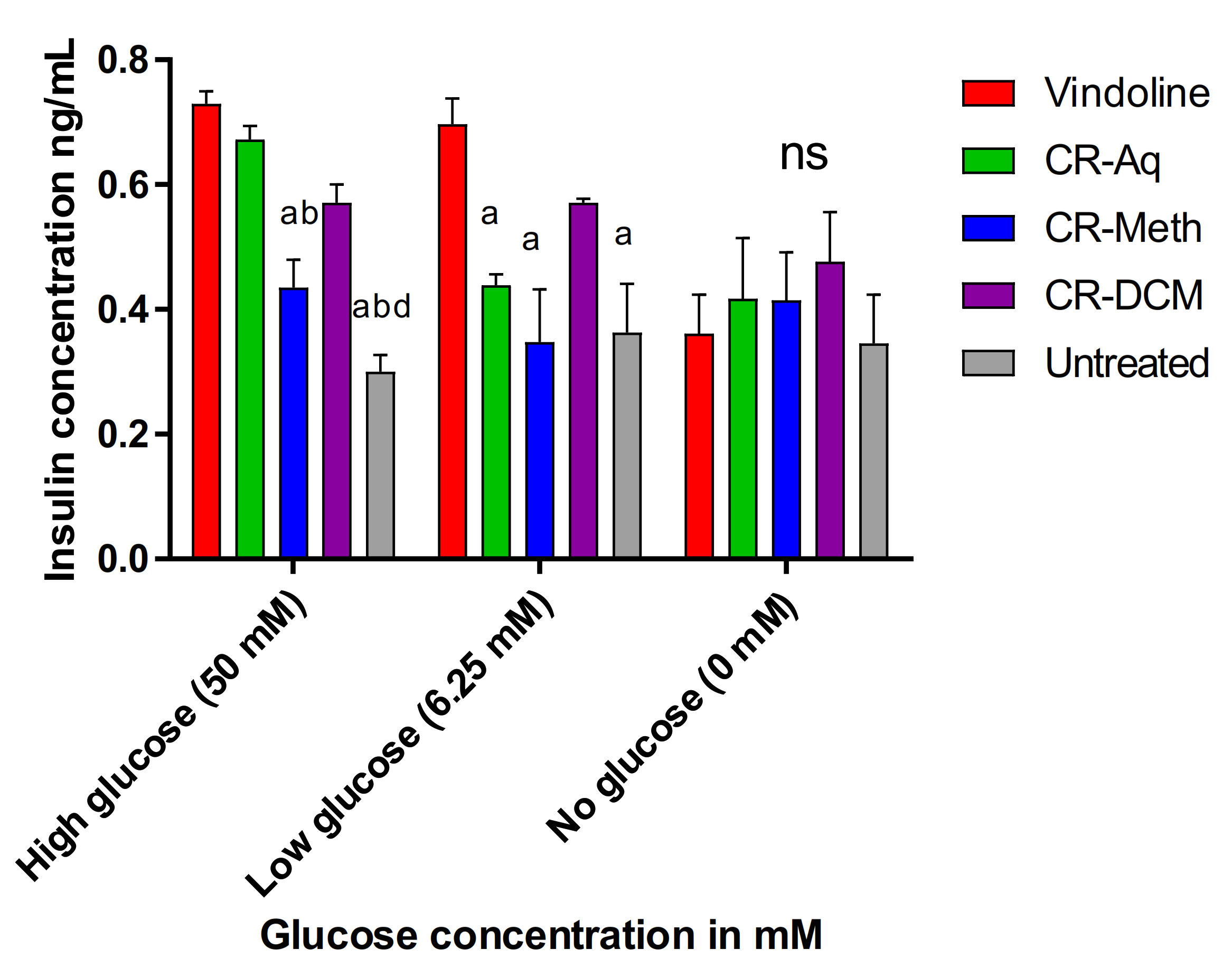
2.7. Effect of Vindoline and the Extracts of C. roseus on Insulin Secretion
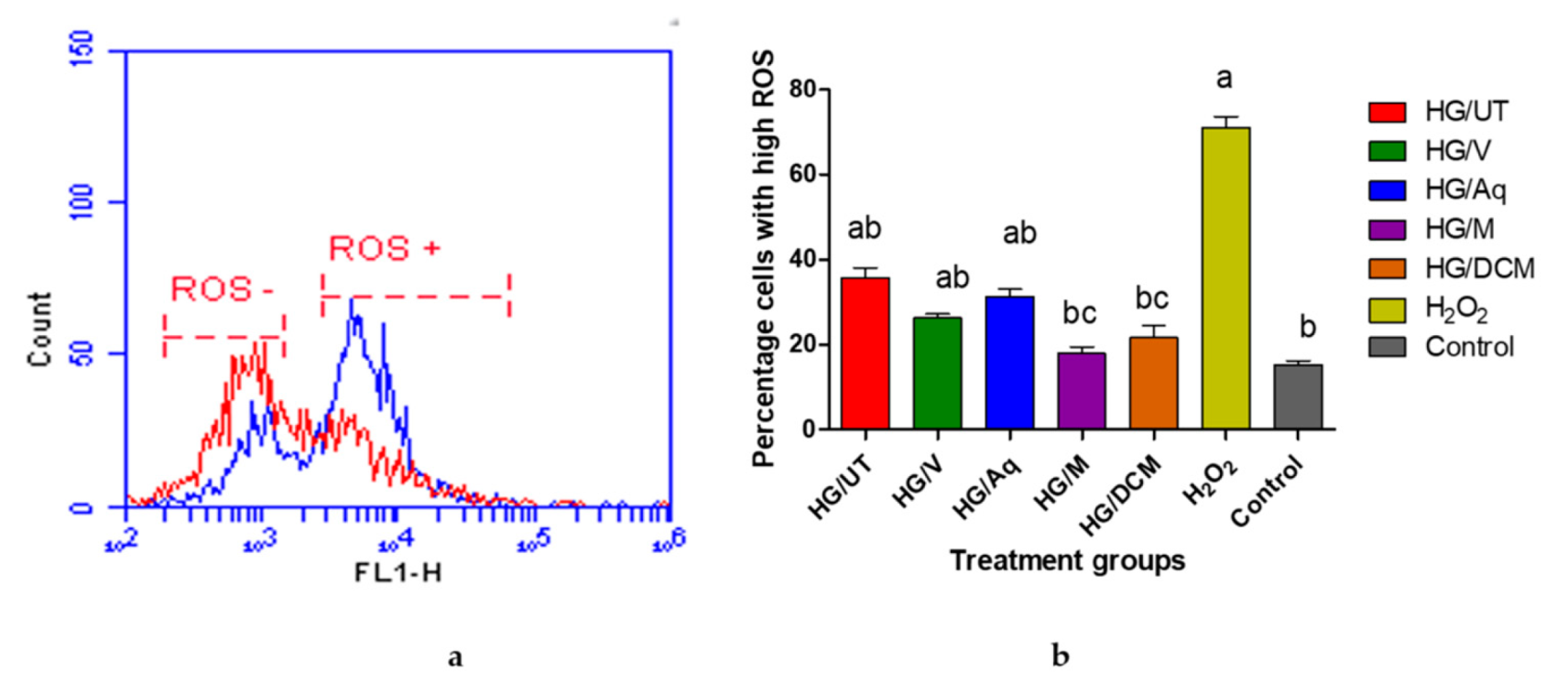
2.8. Analysis of the Generation of Intracellular Reactive Oxygen Species in RIN-5F Cells
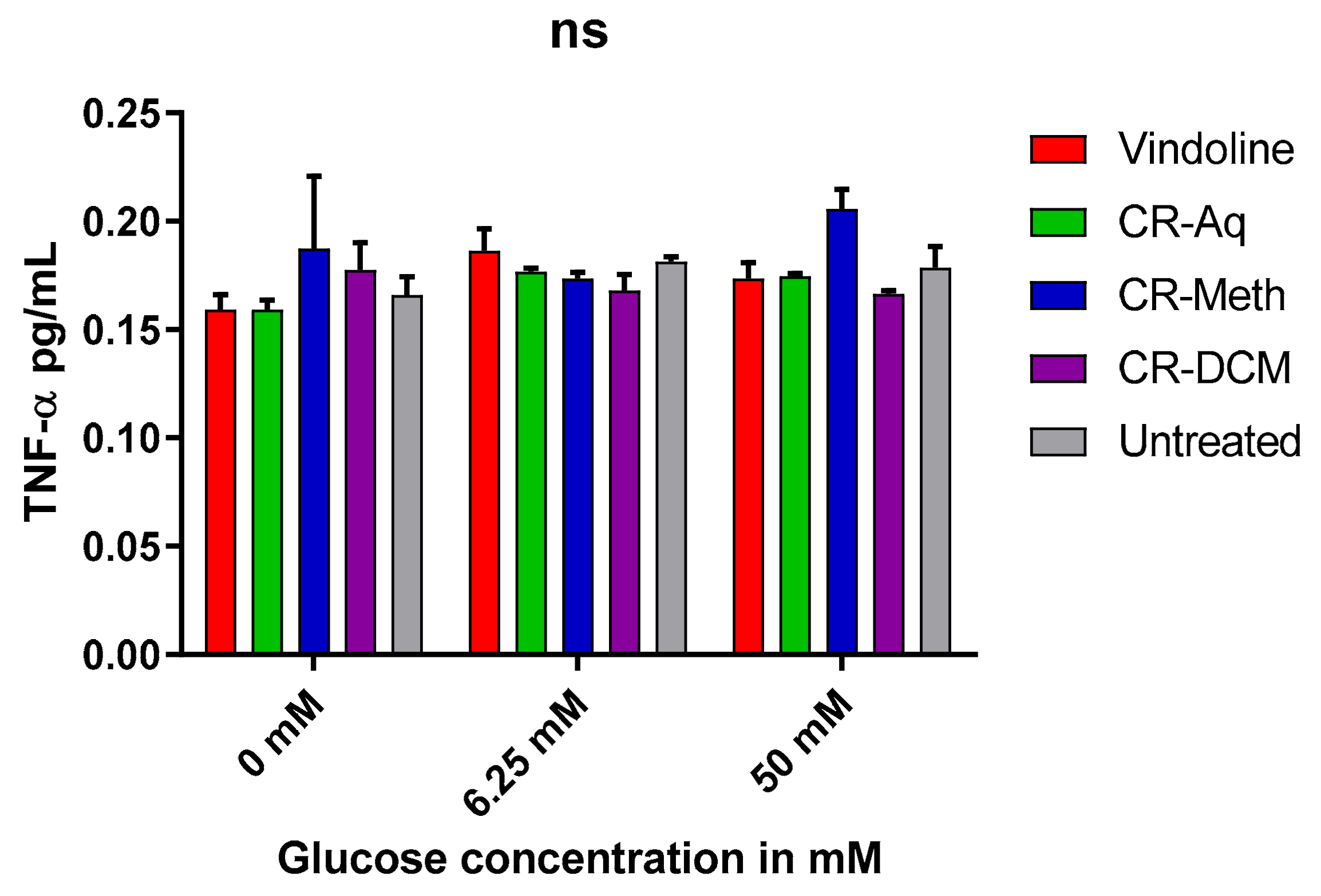
2.9. Effect of Vindoline and the Extracts on the Levels of TNF-α Levels
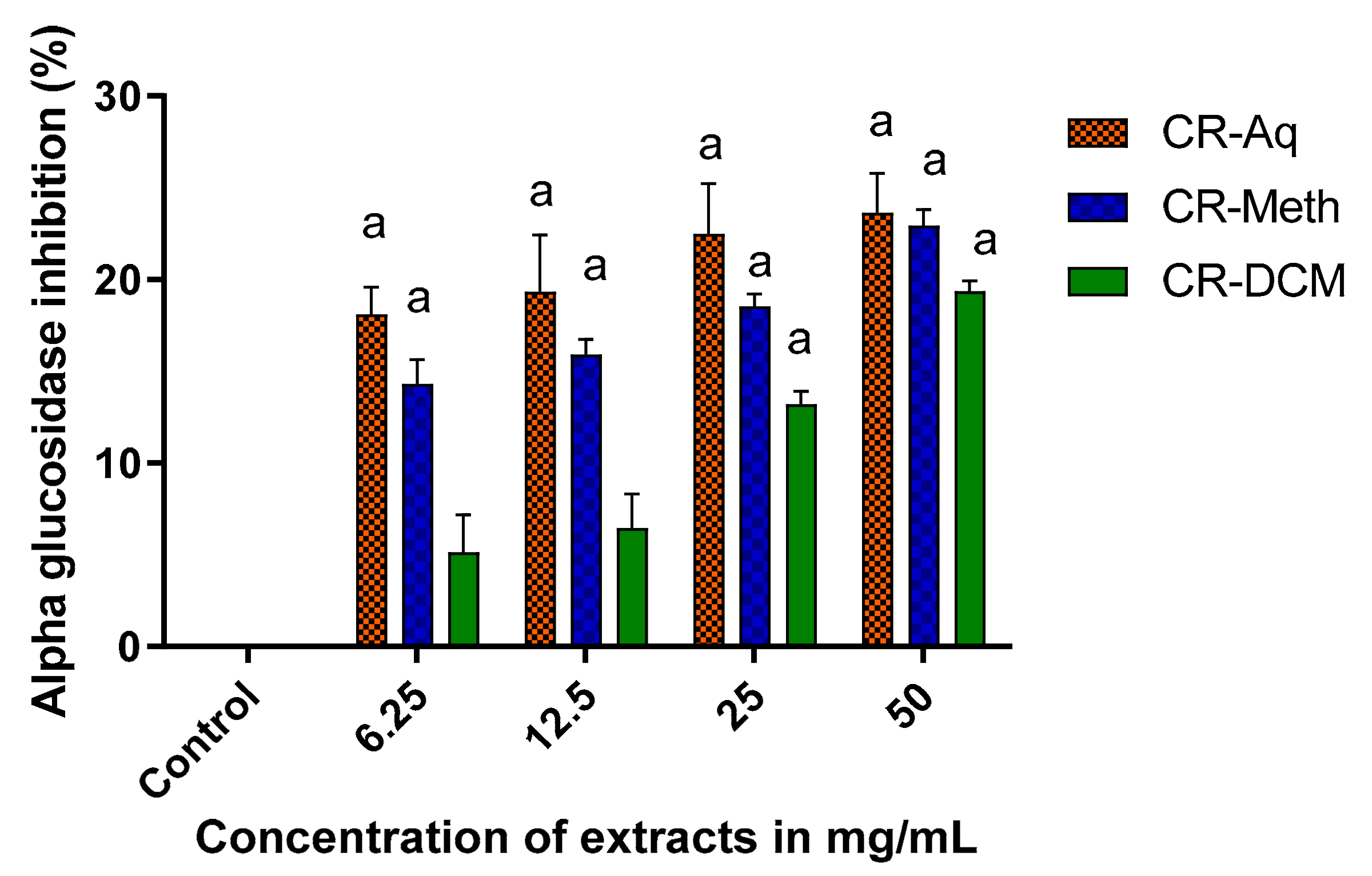
2.10. Alpha Glucosidase Inhibitory Activity
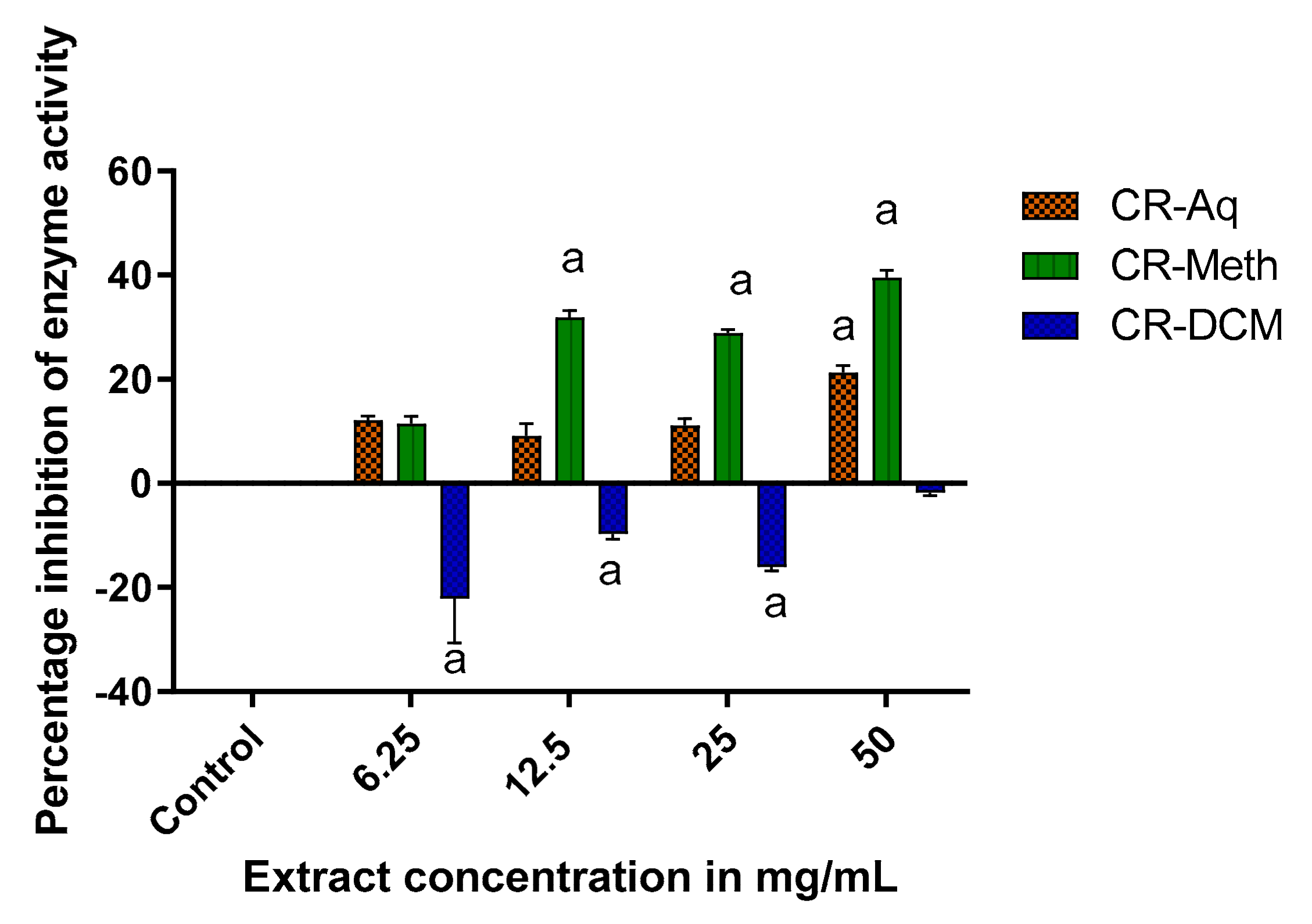
2.11. Alpha Amylase Inhibitory Effects of C. roseus Extracts
2.12. Alpha Glucosidase and Alpha Amylase Inhibitory Activity of Vindoline
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. High-Performance Liquid Chromatography (HPLC)
4.4. Measurement
4.5. Determination of the Oxygen Radical Absorbance Capacity (ORAC)
4.6. DPPH Assay
4.7. FRAP Assay
4.8. In Vitro Cell Line Studies
4.9. Cell Viability Assay
4.10. Reactive Oxygen Species Assay (ROSA)
4.11. Insulin Secretion Assay
4.12. Determination of Inflammation
4.13. Alpha Amylase Inhibitory Assay
4.14. Alpha Glucosidase Inhibitory Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A












References
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M.; et al. Prevention of Type 2 Diabetes Mellitus by Changes in Lifestyle among Subjects with Impaired Glucose Tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef]
- Pheiffer, C.; Pillay-van Wyk, V.; Joubert, J.D.; Levitt, N.; Nglazi, M.D.; Bradshaw, D. The prevalence of type 2 diabetes in South Africa: A systematic review protocol. BMJ Open 2018, 8, e021029. [Google Scholar] [CrossRef] [Green Version]
- Olokoba, A.B.; Obateru, O.A.; Olokoba, L.B. Type 2 diabetes mellitus: A review of current trends. Oman Med. J. 2012, 27, 269–273. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Francini, F.; Schinella, G.R. Natural Products for the Treatment of Type 2 Diabetes Mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar]
- Yao, X.G.; Chen, F.; Li, P.; Quan, L.; Chen, J.; Yu, L.; Ding, H.; Li, C.; Chen, L.; Gao, Z.; et al. Natural product vindoline stimulates insulin secretion and efficiently ameliorates glucose homeostasis in diabetic murine models. J. Ethnopharmacol. 2013, 150, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Zargaran, A.; Rafieian-Kopaei, M.; Saki, K. Ethnobotanical study of medicinal plants used in the management of diabetes mellitus in the Urmia, Northwest Iran. Asian Pac. J. Trop. Med. 2014, 7, S348–S354. [Google Scholar] [CrossRef] [Green Version]
- Kendall, D.M.; Harmel, A.P. The Metabolic Syndrome, Type 2 Diabetes, and Cardiovascular Disease: Understanding the Role of Insulin Resistance. Am. J. Manag. Care 2002, 8, S635–S653. [Google Scholar] [PubMed]
- Alarcon-Aguilar, F.J.; Roman-Ramos, R.; Flores-Saenz, J.L.; Aguirre-Garcia, F. Investigation on the hypoglycaemic effects of extracts of four Mexican medicinal plants in normal and alloxan-diabetic mice. Phyther. Res. 2002, 16, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Fennell, C.W.; Lindsey, K.L.; McGaw, L.J.; Sparg, S.G.; Stafford, G.I.; Elgorashi, E.E.; Grace, O.M.; van Staden, J. Assessing African medicinal plants for efficacy and safety: Pharmacological screening and toxicology. J. Ethnopharmacol. 2004, 94, 205–217. [Google Scholar] [CrossRef]
- Bhogireddy, N.; Naga, A.; Ramesh, B.; Pradeep, M. Anti-inflammatory and anti-diabetic activities with their other ethnomedicinal properties of the plants. J. Med. Plants Stud. 2013, 1, 87–96. [Google Scholar]
- Salihu Shinkafi, T.; Bello, L.; Wara Hassan, S.; Ali, S. An ethnobotanical survey of antidiabetic plants used by Hausa–Fulani tribes in Sokoto, Northwest Nigeria. J. Ethnopharmacol. 2015, 172, 91–99. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. C. R. Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef]
- Jesmin, S.; Khan, A.R.; Jahan, W.A.; Yusuf, M.A.; Begum, W.; Begum, M.; Ara, S. Comparative Study of the Lipid Lowering Effect of Leaf Extract of Catharanthus roseus & Atorvastatin. J. Natl. Inst. Neurosci. Bangladesh 2017, 1, 53. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, N.R.; Verpoorte, R. Phenolic compounds in Catharanthus roseus. Phytochem. Rev. 2007, 6, 243–258. [Google Scholar] [CrossRef] [Green Version]
- Ponarulselvam, S.; Panneerselvam, C.; Murugan, K.; Aarthi, N.; Kalimuthu, K.; Thangamani, S. Synthesis of silver nanoparticles using leaves of Catharanthus roseus Linn. G. Don and their antiplasmodial activities. Asian Pac. J. Trop. Biomed. 2012, 2, 574–580. [Google Scholar] [CrossRef] [Green Version]
- Almagro, L.; Fernández-Pérez, F.; Pedreño, M.A. Indole alkaloids from Catharanthus roseus: Bioproduction and their effect on human health. Molecules 2015, 20, 2973–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, K.; Tiloke, C.; Naidoo, P.; Chuturgoon, A.A. Phytonanotherapy for management of diabetes using green synthesis nanoparticles. J. Photochem. Photobiol. B Biol. 2017, 173, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O.; Meyer, S.; Aboua, Y.G.; Goboza, M. Hypoxis hemerocallidea Significantly Reduced Hyperglycaemia and Hyperglycaemic-Induced Oxidative Stress in the Liver and Kidney Tissues of Streptozotocin-Induced Diabetic Male Wistar Rats. Evid. Based Complement. Altern. Med. 2016, 2016, 8934362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quideau, S.; Deffieux, D.; Douat-casassus, C.; Pouysøgu, L. Natural Products Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. Engl. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Testa, R.; Bonfigli, A.R.; Genovese, S.; De Nigris, V.; Testa, R.; Bonfigli, R.; Genovese, S.; De Nigris, V.; Ceriello, A. The Possible Role of Flavonoids in the Prevention of Diabetic Complications. Nutrients 2016, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Chellan, N.; Muller, C.J.F.; De Beer, D.; Joubert, E.; Page, B.J.; Louw, J. An in vitro assessment of the effect of Athrixia phylicoides DC. aqueous extract on glucose metabolism. Phytomedicine 2012, 19, 730–736. [Google Scholar] [CrossRef]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic Acid on regulating glucose and lipids metabolism: A review. Evid. Based. Complement. Alternat. Med. 2013, 2013, 801457. [Google Scholar] [CrossRef] [PubMed]
- Rasool, N.; Rizwan, K.; Zubair, M.; Naveed, K.U.R.; Imran, I.; Ahmed, V.U. Antioxidant potential of different extracts and fractions of Catharanthus roseus shoots. Int. J. Phytomed. 2011, 3, 108–114. [Google Scholar]
- Kabesh, K.; Senthilkumar, P.; Ragunathan, R.; Kumar, R.R. Phytochemical Analysis of Catharanthus roseus Plant Extract and its Antimicrobial Activity. Int. J. Pure Appl. Biosci. 2015, 3, 162–172. [Google Scholar]
- Islam, M.A.; Akhtar, M.A.; Khan, M.R.I.; Hossain, M.S.; Alam, M.K.; Wahed, M.I.I.; Rahman, B.M.; Anisuzzaman, A.S.M.; Shaheen, S.M.; Ahmed, M. Antidiabetic and Hypolipidemic Effects of Different Fractions of Catharanthus Roseus (Linn.) on Normal and Streptozotocin-induced Diabetic Rats. J. Sci. Res. 2009, 1, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Wang, S.Y. Antioxidant activity and phenolic compounds in selected herbs. J. Agric. Food Chem. 2001, 49, 5165–5170. [Google Scholar] [CrossRef]
- Kaur, S.; Mondal, P. Study of Total Phenolic and Flavonoid Content, Antioxidant Activity and Antimicrobial Properties of Medicinal Plants. J. Microbiol. Exp. 2014, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Padiya, R.; Adela, R.; Putcha, U.K.; Reddy, G.S.; Reddy, B.R.; Kumar, K.P.; Chakravarty, S.; Banerjee, S.K. Garlic and resveratrol attenuate diabetic complications, loss of β-cells, pancreatic and hepatic oxidative stress in streptozotocin-induced diabetic rats. Front. Pharmacol. 2016, 7, 360. [Google Scholar] [CrossRef] [Green Version]
- Alshatwi, A.A.; Subash-Babu, P. Aloe-emodin protects RIN-5F (Pancreatic β-cell) cell from glucotoxicity via regulation of pro-inflammatory cytokine and downregulation of bax and caspase 3. Biomol. Ther. 2016, 24, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.C.; Mustafa, M.R.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef] [Green Version]
- Weir, G.C.; Bonner-Weir, S. Five of stages of evolving β-cell dysfunction during progression to diabetes. Diabetes 2004, 53, S16–S21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taha, H.; Arya, A.; Paydar, M.; Looi, C.Y.; Wong, W.F.; Vasudeva Murthy, C.R.; Noordin, M.I.; Ali, H.M.; Mustafa, A.M.; Hadi, A.H.A. Upregulation of insulin secretion and downregulation of pro-inflammatory cytokines, oxidative stress and hyperglycemia in STZ-nicotinamide-induced type 2 diabetic rats by Pseuduvaria monticola bark extract. Food Chem. Toxicol. 2014, 66, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Wickramaratne, M.N.; Punchihewa, J.C.; Wickramaratne, D.B.M. In-vitro alpha amylase inhibitory activity of the leaf extracts of Adenanthera pavonina. BMC Complement. Altern. Med. 2016, 16, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, P.M.; de Sales, P.M.; Simeoni, L.A.; Silva, E.C.; Silveira, D.; de Magalhães, P.O. Inhibitory Activity of α -Amylase and α -Glucosidase by Plant Extracts from the Brazilian Cerrado. Planta Med 2012, 78, 393–399. [Google Scholar] [CrossRef]
- Bhutkar, M.A.; Bhise, S.B. In vitro assay of alpha amylase inhibitory activity of some indigenous plants. Int. J. Chem. Sci. 2012, 10, 457–462. [Google Scholar] [CrossRef]
- Jyothi, K.S.N.; Hemalatha, P.; Avanthi, A.; Challa, S. A comparative analysis on the alpha amylase inhibitory potential of six ornamental medicinal plants. J. Nat. Prod. Plant. Resour. 2013, 3, 1–6. [Google Scholar]
- Singh, S.N.; Vats, P.; Suri, S.; Shyam, R.; Kumria, M.M.; Ranganathan, S.; Sridharan, K. Effect of an antidiabetic extract of Catharanthus roseus on enzymic activities in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2001, 76, 269–277. [Google Scholar] [CrossRef]
- Jayanthi, M.; Sowbala, N.; Rajalakshmi, G.; Kanagavalli, U.; Sivakumar, V. Study of anti hyperglycemic effect of Catharanthus roseus in alloxan induced diabetic rats. Int. J. Pharm. Pharm. Sci. 2010, 2, 114–116. [Google Scholar]
- Malathi, V.; Devi, S.S.; Revathi, K. Anti Diabetic Activity By The in vitro alpha amylase and alpha-glucosidase inhibitory activity of catharanthus roseus. Biascan Int. J. life Sci. 2010, 5, 655–659. [Google Scholar]
- Bramati, L.; Minoggio, M.; Gardana, C.; Simonetti, P.; Mauri, P.; Pietta, P. Quantitative Characterization of Flavonoid Compounds in Rooibos Tea (Aspalathus linearis) by LC−UV/DAD. J. Agric. Food Chem. 2002, 50, 5513–5519. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Berridge, M.V.; Tan, A.S. Trans-plasma membrane electron transport: A cellular assay for NADH- and NADPH-oxidase based on extracellular, superoxide-mediated reduction of the sulfonated tetrazolium salt WST-1. Protoplasma 1998, 205, 74–82. [Google Scholar] [CrossRef]
- Venditti, P.; Balestrieri, M.; Di Meo, S.; Leo, T. De Effect of thyroid state on lipid peroxidation, antioxidant defences, and susceptibility to oxidative stress in rat tissues. J. Endocrinol. 1997, 155, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, M.; Varshney, R.; Chopra, M.; Sekhri, T.; Adhikari, J.S.; Dwarakanath, B.S. Flow-cytometric analysis of reactive oxygen species in peripheral blood mononuclear cells of patients with thyroid dysfunction. Cytom. Part B Clin. Cytom. 2005, 70B, 20–23. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Chlorogenic Acid (µg/g) | Caffeic Acid (µg/g) | Quercetin (µg/g) | Coumaric Acid (µg/g) | Vindoline (µg/g) | Rutin (µg/g) |
|---|---|---|---|---|---|---|
| CR-Aq | 33.461 | 1.179 | 0.445 | 2.195 | 7.056 | 5.891 |
| CR-Meth | 225.19 | 0.614 | 1.945 | 28.822 | 15.397 | 85.916 |
| CR-Ethyl | 0.466 | 0.396 | 1.263 | 0.693 | 57.323 | 1.811 |
| CR-DCM | 2.308 | 0.017 | 0.253 | 0.197 | 57.891 | 4.506 |
| Concentration | Vindoline | Ascorbic Acid | |
|---|---|---|---|
| FRAP (µM) | 0.05 µM | 23,842 ± 339.3 ns | 24.514 ± 95.7 ns |
| ORAC (µmol TE/L) DPPH (%) | 0.05 µM 0.15 mg/mL | 56.0 ± 4.9 a 40.56 ± 9.28 ns | 40.86 ± 3.85 7.36 ± 11.31 ns |
| Test | Concentration | Vindoline Inhibition (%) | Acarbose Inhibition (%) |
|---|---|---|---|
| Alpha Glucosidase | 0.375 mM | 11.31 ± 2.351a | 83.35 ± 0.3 |
| Alpha Amylase | 0.375 mM | 18.57 ± 2.881a | 79.50 ± 1.081 |
Data Availability: Raw data used for this study is available from the authors upon reasonable request. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goboza, M.; Meyer, M.; Aboua, Y.G.; Oguntibeju, O.O. In Vitro Antidiabetic and Antioxidant Effects of Different Extracts of Catharanthus roseus and Its Indole Alkaloid, Vindoline . Molecules 2020, 25, 5546. https://doi.org/10.3390/molecules25235546
Goboza M, Meyer M, Aboua YG, Oguntibeju OO. In Vitro Antidiabetic and Antioxidant Effects of Different Extracts of Catharanthus roseus and Its Indole Alkaloid, Vindoline . Molecules. 2020; 25(23):5546. https://doi.org/10.3390/molecules25235546
Chicago/Turabian StyleGoboza, Mediline, Mervin Meyer, Yapo G. Aboua, and Oluwafemi O. Oguntibeju. 2020. "In Vitro Antidiabetic and Antioxidant Effects of Different Extracts of Catharanthus roseus and Its Indole Alkaloid, Vindoline " Molecules 25, no. 23: 5546. https://doi.org/10.3390/molecules25235546