Fitness Cost of Imazamox Resistance in Wild Poinsettia (Euphorbia heterophylla L.)

 , and
, and 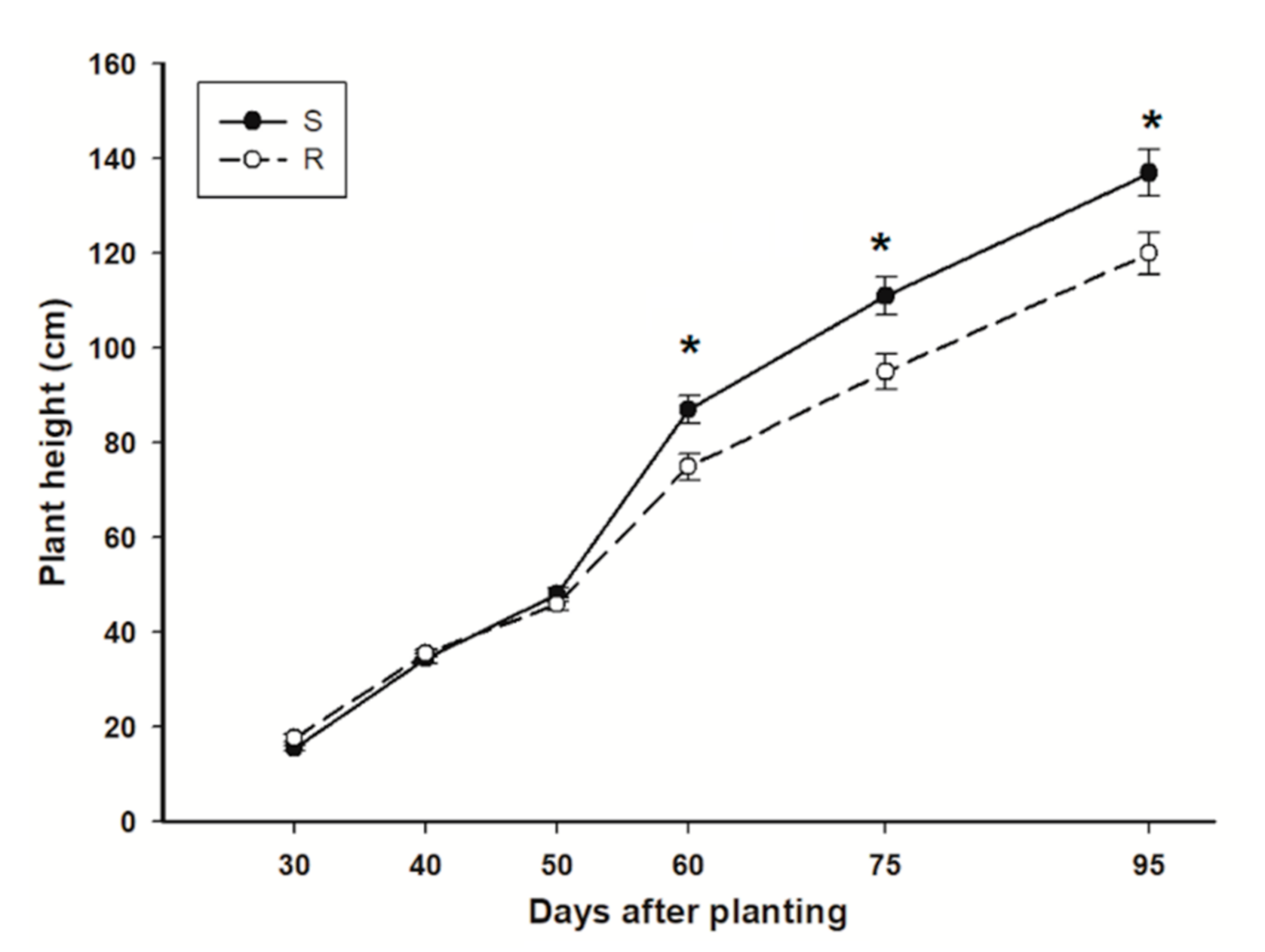{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growth Conditions and Measurement of Fitness Traits
2.3. Statistical Analysis
3. Results
3.1. Plant Height
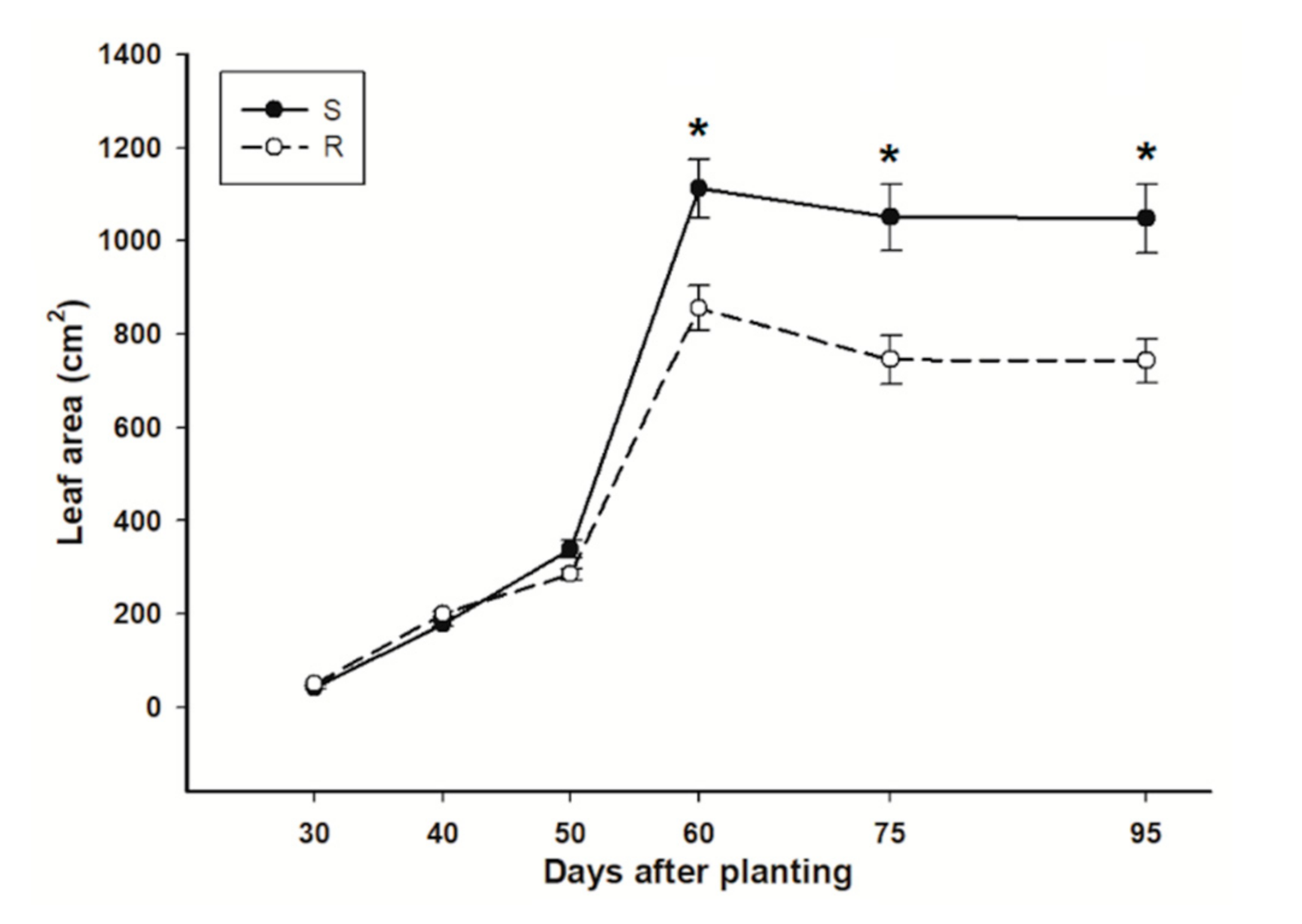
3.2. Leaf Area
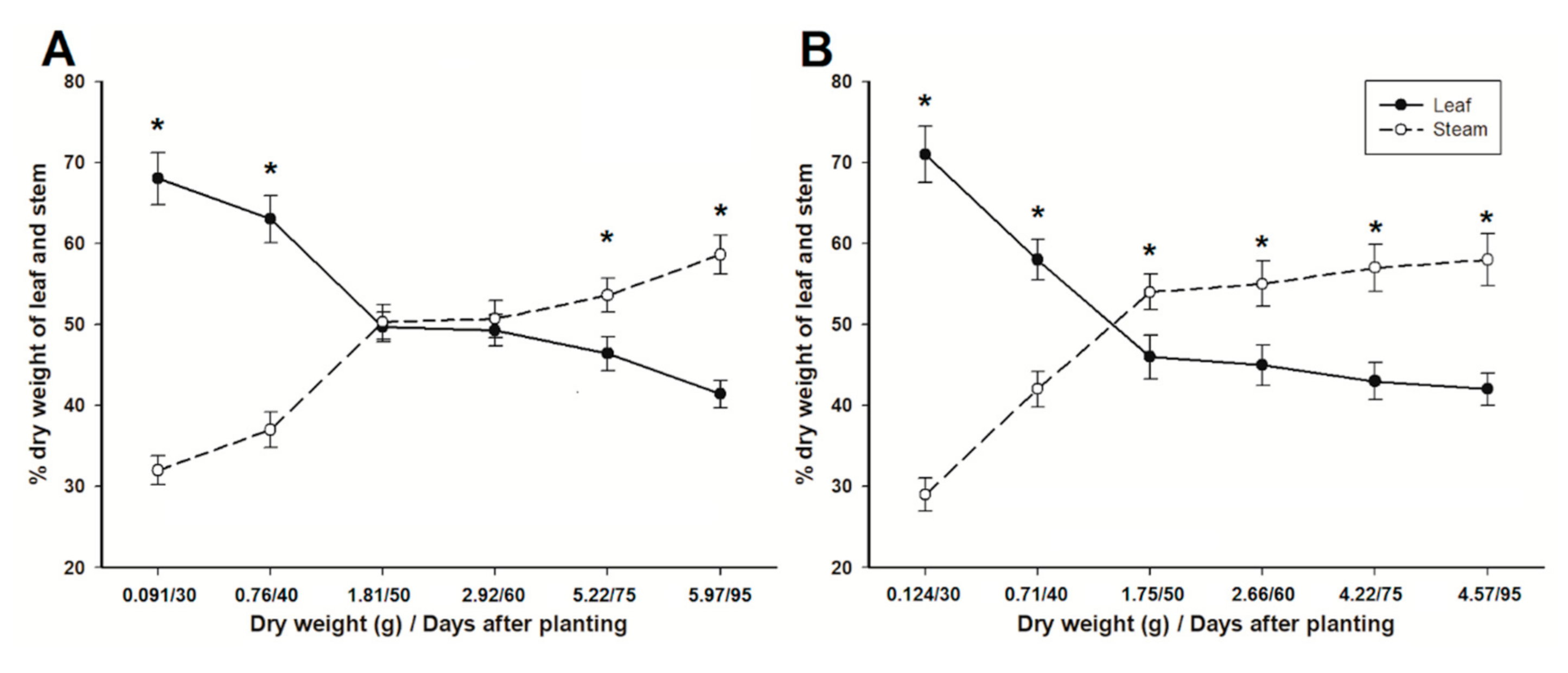
3.3. Dry Weights
3.4. Share of Weight and Seed Production
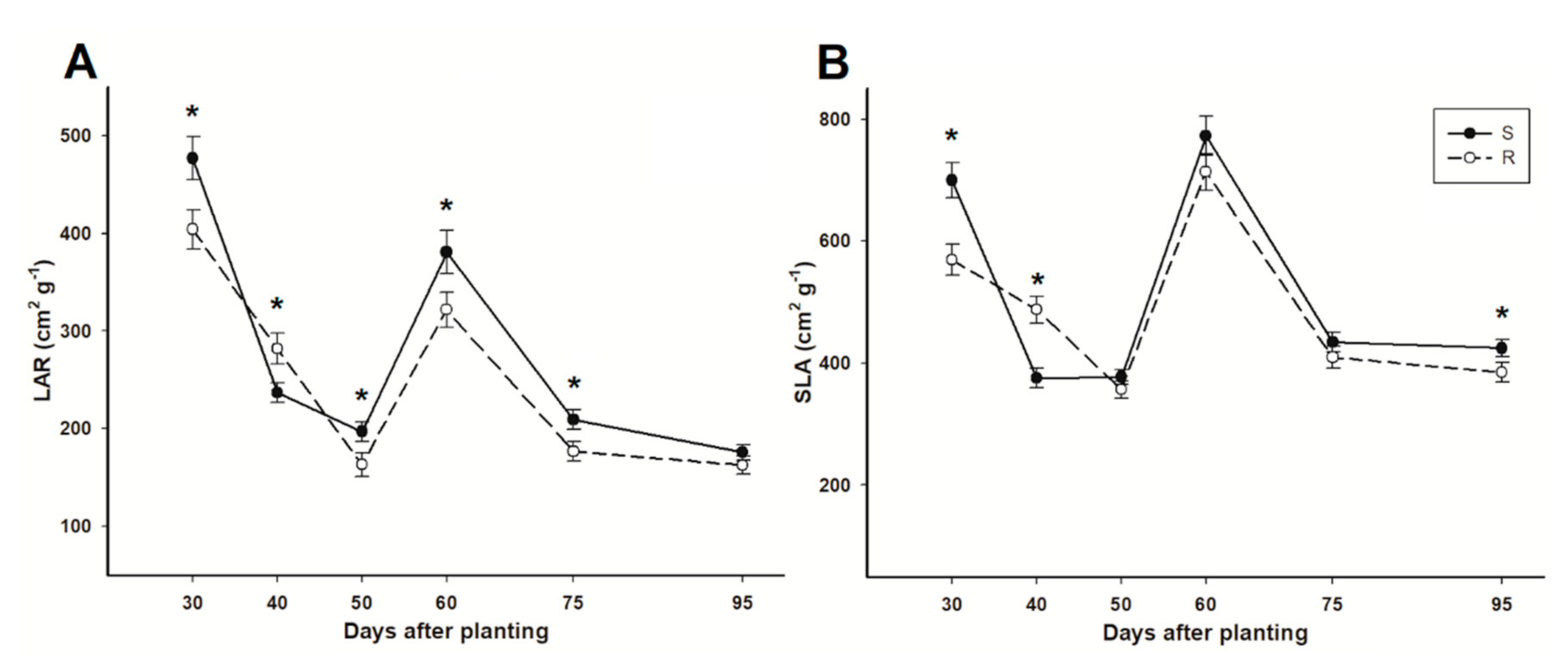
3.5. Leaf Area Ratio and Specific Leaf Area
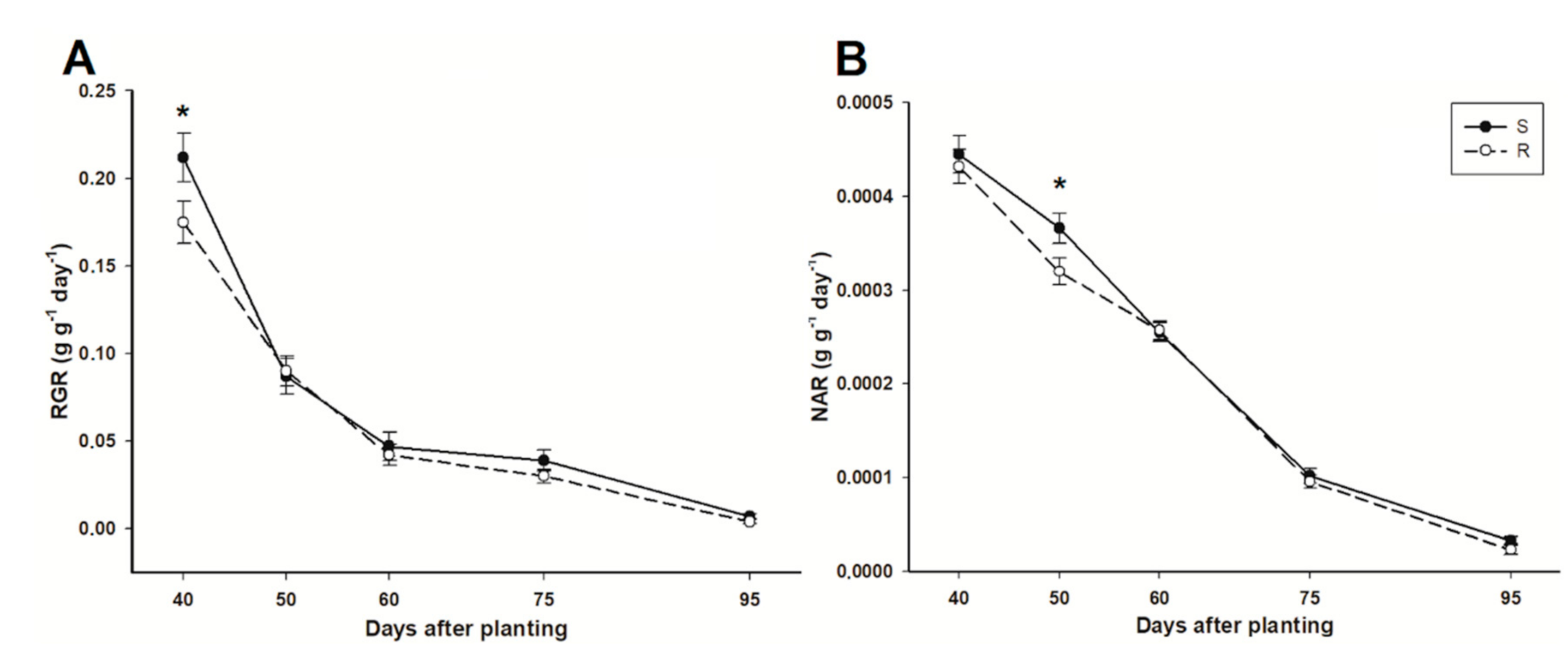
3.6. Relative Growth Rate and Net Assimilation Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chauhan, B.S. Grand challenges in weed management. Front. Agron. 2020, 1, 3. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: http://weedscience.org/ (accessed on 30 August 2020).
- Vila-Aiub, M.M. Fitness of herbicide-resistant weeds: Current knowledge and implications for management. Plants 2019, 8, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousens, R.D.; Fournier-Level, A. Herbicide resistance costs: What are we actually measuring and why? Pest. Manag. Sci. 2018, 74, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Vila-Aiub, M.M.; Yu, Q.; Powles, S.B. Do plants pay a fitness cost to be resistant to glyphosate? New Phytol. 2019, 223, 532–547. [Google Scholar] [CrossRef] [Green Version]
- Hassanpour-Bourkheili, S.; Gherekhloo, J.; Kamkar, B.; Ramezanpour, S.S. Comparing fitness cost associated with haloxyfop-R methyl ester resistance in winter wild oat biotypes. Planta Daninha 2020, 38, e020213759. [Google Scholar] [CrossRef]
- Wang, T.; Picard, J.C.; Tian, X.; Darmency, H. A herbicide-resistant ACCase 1781 Setaria mutant shows higher fitness than wild type. Heredity 2010, 105, 394–400. [Google Scholar] [CrossRef]
- Keshtkar, E.; Abdolshahi, R.; Sasanfar, H.; Zand, E.; Beffa, R.; Dayan, F.E.; Kudsk, P. Assessing fitness costs from a herbicide-resistance management perspective: A review and insight. Weed Sci. 2019, 67, 137–148. [Google Scholar] [CrossRef]
- Du, L.; Qu, M.; Jiang, X.; Li, X.; Ju, Q.; Lu, X.; Wang, J. Fitness costs associated with acetyl-coenzyme A carboxylase mutations endowing herbicide resistance in American sloughgrass (Beckmannia syzigachne Steud.). Ecol. Evol. 2019, 9, 2220–2230. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.A.L.; Owen, M.D.K.; Leon, R.G. Influence of multiple herbicide resistance on growth in Amaranthus tuberculatus. Weed Res. 2019, 59, 235–244. [Google Scholar] [CrossRef]
- Henckes, J.R.; Cechin, J.; Schmitz, M.F.; Piasecki, C.; Vargas, L.; Agostinetto, D. Fitness cost and competitive ability of ryegrass susceptible and with multiple resistance to glyphosate, iodosulfuron-methyl, and pyroxsulam. Planta Daninha 2019, 37, e019197532. [Google Scholar] [CrossRef]
- Anthimidou, E.; Ntoanidou, S.; Madesis, P.; Eleftherohorinos, I. Mechanisms of Lolium rigidum multiple resistance to ALS-and ACCase-inhibiting herbicides and their impact on plant fitness. Pestic. Biochem. Physiol. 2019, 164, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Vila-Aiub, M.M.; Goh, S.S.; Gaines, T.A.; Han, H.; Busi, R.; Yu, Q.; Powles, S.B. No fitness cost of glyphosate resistance endowed by massive EPSPS gene amplification in Amaranthus palmeri. Planta 2014, 239, 793–801. [Google Scholar] [CrossRef]
- Travlos, I.S. Competition between ACCase-inhibitor resistant and susceptible sterile wild oat (Avena sterilis) biotypes. Weed Sci. 2013, 61, 26–31. [Google Scholar] [CrossRef]
- Wilson, A.K. Euphorbia heterophylla: A review of distribution, importance and control. Trop Pest. Manag. 1981, 27, 32–38. [Google Scholar] [CrossRef]
- Lati, R.N.; Goldwasser, Y.; Horesh, A.; Igbariya, K. Wild poinsettia biology and management–determining optimal control with herbicides and propane flaming. Crop. Prot. 2019, 115, 20–26. [Google Scholar] [CrossRef]
- Chachalis, D. Wild poinsettia (Euphorbia heterophylla): An emerging weed in cotton and processing tomato in Greece. Hell. Plant Prot. 2015, 8, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Agostinetto, D.; Ulguim, A.R.; Zandoná, R.R.; Silva, J.D.G.; Silva, B.M.; Várgas, L. Low level resistance of wild poinsettia biotypes to glyphosate and alternative chemical control. Biosci. J. 2019, 35, 1060–1070. [Google Scholar] [CrossRef]
- Vidal, R.A.; Winkler, L.M.; Hernandes, G.C.; Fleck, N.G.; Merotto, A., Jr.; Trezzi, M.M. A field survey of crop management practices and distribution of ALS resistant Euphorbia heterophylla in two states in southern Brazil. Planta Daninha 2004, 22, 403–410. [Google Scholar] [CrossRef]
- Trezzi, M.M.; Felippi, C.L.; Mattei, D.; Silva, H.L.; Nunes, A.L.; Debastiani, C.; Vidal, R.A.; Marques, A. Multiple resistance of acetolactate synthase and protoporphyrinogen oxidase inhibitors in Euphorbia heterophylla biotypes. J. Environ. Sci. Health B 2005, 40, 101–109. [Google Scholar] [CrossRef]
- Vidal, R.A.; Trezzi, M.M.; De Prado, R.; Ruiz-Santaella, J.P.; Vila-Aiub, M. Glyphosate resistant biotypes of wild poinsettia (Euphorbia heterophylla L.) and its risk analysis on glyphosate-tolerant soybeans. J. Food Agric. Environ. 2007, 5, 265–269. [Google Scholar]
- Rojano-Delgado, A.M.; Portugal, J.M.; Palma-Bautista, C.; Alcántara-de la Cruz, R.; Torra, J.; Alcántara, E.; De Prado, R. Target site as the main mechanism of resistance to imazamox in a Euphorbia heterophylla biotype. Sci. Rep. 2019, 9, 15423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menchari, Y.; Chauvel, B.; Darmency, H.; Délye, C. Fitness costs associated with three mutant acetyl-coenzyme A carboxylase alleles endowing herbicide resistance in black-grass Alopecurus myosuroides. J. Appl. Ecol. 2008, 45, 939–947. [Google Scholar] [CrossRef]
- Schmitz, M.F.; Cechin, J.; Henckes, J.R.; Piasecki, C.; Agostinetto, D.; Vargas, L. Fitness cost and competitive ability to different ploidy levels in ryegrass genotypes. Planta Daninha 2019, 37, e019181921. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Ann. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef] [Green Version]
- Yanniccari, M.; Vila-Aiub, M.; Istilart, C.; Acciaresi, H.; Castro, A.M. Glyphosate resistance in perennial ryegrass (Lolium perenne L.) is associated with a fitness penalty. Weed Sci. 2016, 64, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, E.; Matzrafi, M.; Rubin, B.; Peleg, Z. Effects of environmental conditions on the fitness penalty in herbicide resistant Brachypodium hybridum. Front. Plant Sci. 2017, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Schaedler, C.E.; Burgos, N.R.; Noldin, J.A.; Alcober, E.A.; Salas, R.A.; Agostinetto, D. Competitive ability of ALS-inhibitor herbicide-resistant Fimbristylis miliacea. Weed Res. 2015, 55, 482–492. [Google Scholar] [CrossRef]
- Park, K.W.; Mallory-Smith, C.A. Multiple herbicide resistance in downy brome (Bromus tectorum) and its impact on fitness. Weed Sci. 2005, 53, 780–786. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Yu, Q.; Han, H.; Powles, S.B. Effect of herbicide resistance endowing Ile-1781-Leu and Asp-2078-Gly ACCase gene mutations on ACCase kinetics and growth traits in Lolium rigidum. J. Exp. Bot. 2015, 66, 4711–4718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, D.E. Components of resource competition in plant communities. In Perspectives in Plant Competition, 1st ed.; Grace, B.J., Tilman, D., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 27–49. [Google Scholar]
- Crooks, H.L.; Burton, M.G.; York, A.C.; Brownie, C. Vegetative growth and competitiveness of common cocklebur resistant and susceptible to acetolactate synthase-inhibiting herbicides. Cotton Sci. 2005, 9, 229–237. [Google Scholar]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Shergill, L.S.; Boutsalis, P.; Preston, C.; Gill, G.S. Fitness costs associated with 1781 and 2041 ACCase-mutant alleles conferring resistance to herbicides in Hordeum glaucum Steud. Crop. Prot. 2016, 87, 60–67. [Google Scholar] [CrossRef]
- Panozzo, S.; Scarabel, L.; Rosan, V.; Sattin, M. A new Ala-122-Asn amino acid change confers decreased fitness to ALS-resistant Echinochloa crus-galli. Front. Plant Sci. 2017, 8, 2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanizadeh, H.; Harrington, K.C. Fitness costs associated with multiple resistance to dicamba and atrazine in Chenopodium album. Planta 2019, 249, 787–797. [Google Scholar] [CrossRef]
- Abdollahipour, M.; Gherekhloo, J.; Bagherani, N.; Taghvaei, M.R. Effects relative fitness of susceptible and tribenuron methyl resistant biotypes of wild mustard (Sinapis arvensis L.). Scinzer J. Agric. Biol. Sci. 2016, 2, 15–23. [Google Scholar]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef]
- Lehnhoff, E.A.; Keith, B.K.; Dyer, W.E.; Peterson, R.K.; Menalled, F. Multiple herbicide resistance in wild oat and impacts on physiology, germinability, and seed production. Agron. J. 2013, 105, 854–862. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Growth and allocation. In Plant Physiological Ecology, 1st ed.; Lambers, H., Chapin, F.S., Pons, T.L., Eds.; Springer: New York, NY, USA, 1998; pp. 299–351. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassanpour-bourkheili, S.; Heravi, M.; Gherekhloo, J.; Alcántara-de la Cruz, R.; De Prado, R. Fitness Cost of Imazamox Resistance in Wild Poinsettia (Euphorbia heterophylla L.). Agronomy 2020, 10, 1859. https://doi.org/10.3390/agronomy10121859
Hassanpour-bourkheili S, Heravi M, Gherekhloo J, Alcántara-de la Cruz R, De Prado R. Fitness Cost of Imazamox Resistance in Wild Poinsettia (Euphorbia heterophylla L.). Agronomy. 2020; 10(12):1859. https://doi.org/10.3390/agronomy10121859
Chicago/Turabian StyleHassanpour-bourkheili, Saeid, Mahtab Heravi, Javid Gherekhloo, Ricardo Alcántara-de la Cruz, and Rafael De Prado. 2020. "Fitness Cost of Imazamox Resistance in Wild Poinsettia (Euphorbia heterophylla L.)" Agronomy 10, no. 12: 1859. https://doi.org/10.3390/agronomy10121859