
Abstract
A multidisciplinary study of the Devonian–Carboniferous boundary interval in pelagic successions of the Holy Cross Mountains and Sudetes and the ramp successions in the Western Pomerania region (Poland) is presented herein. The analysis applies the results of new palaeontological and biostratigraphic studies based mainly on conodonts, ammonoids and palynomorphs, biostratigraphic results interpreted earlier by different authors that have been re-examined, and geochemical and mineralogical characteristics, as well as magnetic susceptibility measurements across the Devonian–Carboniferous boundary interval. The study is focused on the interval from the Famennian ultimus conodont Zone to the Tournaisian duplicata conodont Zone, and from the Famennian lepidophyta–explanatus (LE) miospore Zone to the Tournaisian verrucosus–incohatus (VI) miospore Zone, respectively. The paper highlights sections, which are the most representative for the Devonian–Carboniferous boundary in each region, illustrates and summarises current knowledge on the uppermost Famennian to lowermost Tournaisian in these regions, gives data and correlation of the important stratigraphic markers for each region, and briefly correlates them outside the region. The sedimentary successions and specific phenomena, together with microscale environmental perturbations, recognised close to the Devonian/Carboniferous boundary in Poland, display a pattern similar to that observed in many areas in Europe during the Hangenberg Event.
Similar content being viewed by others

Introduction
Previous and recent studies across the Devonian–Carboniferous boundary (DCB) interval have demonstrated several problems with its determination, resulting from the quality of the collected material and the species concept of individual researchers for the definition of this boundary (e.g. Kaiser et al. 2009; Becker et al. 2016; Corradini et al. 2017; Spalletta et al. 2017; Kaiser et al. 2019). It seems, therefore, that a single boundary marker selected in the future should be accompanied by alternatives such as ranges of other fossils or chemostratigraphic indices.
The goal of this study is to provide a detailed documentation of important stratigraphic markers across the DCB interval for different regions of Poland, as a contribution to the ongoing international revision of the DCB.
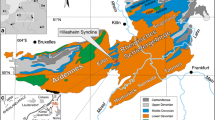
The several-hundred-kilometres-wide shelf area was located in the Devonian in the Polish part of a pericratonic basin, which stretched from Western Europe to the Ukraine along the periphery of Laurussia (Old Red Sandstone Continent). The pelagic successions are situated in the southern part of the Polish Basin, whereas the neritic successions are located in its northern part (Fig. 1). The most complete pelagic successions occur in the active Kowala Quarry near Kielce in the Holy Cross Mountains and in the old Wapnica Quarry near Dzikowiec village in the Sudetes, and are studied more thoroughly herein. Supplementary and comparative data were derived from neritic sections, where the uppermost Famennian ramp sequence was documented in the Western Pomerania area in the Gorzysław-9, Rzeczenica-1 and Chmielno-1 borehole sections. The analysis presented herein, including the uppermost Famennian and lowermost Tournaisian strata, together with the Hangenberg Black Shale (HBS) horizon, applies the results of new palaeontological and biostratigraphic studies based mainly on conodonts, ammonoids and palynomorphs, as well as biostratigraphic results interpreted earlier by different authors that have been re-examined wherever it was possible (for details, see “Analytical sections”). The present paper illustrates and summarises current knowledge on the uppermost Famennian to lowermost Tournaisian in these regions, gives detailed data and correlation of the important stratigraphic markers for each region and briefly correlates them outside the region; moreover, it highlights sections which are the most representative for the DCB in each region.

Position of the studied localities within the palaeogeographic and structural framework of the Devonian of Poland (modified after Racki et al. 2002); in asterisks: A—Kowala (Holy Cross Mountains), B—Dzikowiec (Sudetes), C—Gorzysław-9, Chmielno-1 and Rzeczenica-1 borehole sections (Western Pomerania area)
Material and methods
Thirty-five new samples were processed for conodonts in the section of the Kowala Quarry (Holy Cross Mountains); most of the samples were conodont productive and almost all of them contained adequate material for biostratigraphic analysis. From the Famennian clymeniid limestone and Tournaisian Gattendorfia limestones in the main section of the Wapnica Quarry near Dzikowiec (Sudety Mts), new samples for conodonts were collected in 2014 by TW-M. Fifteen limestone samples were processed, out of which 8 yielded conodont fauna. The presence of the uppermost Famennian–lowermost Tournaisian succession in the Western Pomerania area was recognised earlier (Matyja 1993; Matyja and Stempień-Sałek 1994) in two borehole sections, Gorzysław-9 and Rzeczenica-1, and a stratigraphic gap close to the DCB was suggested. Unfortunately, these two sections were regularly sampled at 1-m intervals only. High-resolution biostratigraphic study in the Chmielno-1 sections, where ninety samples at ~ 20-cm intervals were collected (Matyja et al. 2015), allows for the recognition of the first complete sequence of the uppermost Famennian and lowermost Tournaisian recorded in the Pomerania area. Therefore, the DCB interval in the Gorzysław-9 section was re-sampled in detail for conodonts and miospores, and 76 samples at ~ 20-cm intervals were collected from the depth range of 3187.0–3203.0 m. Unfortunately, there was no possibility to re-sample the rock material from the DCB interval in the Rzeczenica-1 section, as it was not available anymore; therefore, the previously collected biostratigraphic data from the depth range of 2920.7–2937.0 m was re-examined.
Standard procedures were employed to all conodont samples (e.g. Stone 1987; Barnes et al. 1987; Merrill et al. 1987), using acetic and monochloroacetic acids to dissolve samples and extract conodonts.
The critical part of the DCB interval in the Kowala section was also intensively tested for palynology in the past; unfortunately, not all samples were positive (e.g. Filipiak 2004, 2005; Filipiak and Racki 2010; Marynowski and Filipiak 2007; Marynowski et al. 2012). New palynologic investigations in the Kowala section revealed the presence of the lowermost Tournaisian miospore zones. Palynomorphs were analysed in the Wapnica Quarry section for the first time. Unfortunately, most of the 15 samples were barren and only three of them, obtained from the approx. 20-cm-thick black shales interval ( = HBS), possess an organic content. Two positive Carboniferous samples were additionally taken from two different walls of the quarry.
Standard laboratory methods were used to process the samples, including the HF–HCl–HF acid sequence (see Wood et al. 1996). Finally, four standard microscope slides were prepared for each residue. Cellosize was used as the organic dispersing agent and Petropoxy 154 as the mounting agent.
The new palaeontological material (conodonts, ammonoids) is housed at the Polish Geological Institute - National Research Institute in Warsaw, and slides and residues (palynomorphs) are housed at the Faculty of Natural Sciences, University of Silesia, in Katowice.
In recent years, the Kowala section was also intensively examined for multiple geochemical proxies that reflected environmental changes close to the DCB interval (e.g. Marynowski and Filipiak 2007; Trela and Malec 2007; Marynowski et al. 2012, 2017; Racki et al. 2018a, 2018b). Recently, the uppermost Famennian–lowermost Tournaisian interval in the Kowala section was analysed also by the present authors using inorganic geochemistry and mineralogy. A set of 11 samples was taken from the outcrop to analyse the bulk rock mineralogy, clay mineralogy (in the < 0.002-mm fraction), major element geochemistry and total organic carbon content (TOC). The samples were studied by XRD, XRF and coulometric methods at the laboratories of the Polish Geological Institute–National Research Institute. Some weathering indices were also calculated. Four samples (6a, 23a, 23b and 23) represent black shale layers, and one sample (29) corresponds to the grey claystone at the top of the Devonian. Other samples were collected from marly limestone strata. Clay minerals (and bulk rock mineralogy) were analysed for the first time in the uppermost Famennian–lowermost Tournaisian interval. Previously, only two samples were studied from the HBS horizon in the Kowala section by Marynowski et al. (2012). Carbon stable isotope (δ13Ccarb) was analysed in the Kowala Quarry (20 samples), in the Wapnica Quarry (16 samples) and in the Gorzysław-9 borehole section (12 samples). Comparative data from the Chmielno-1 section (Matyja et al. 2015) was also taken into consideration.
Rock magnetic susceptibility (MS) for the DCB interval was sampled and measured in the Kowala Quarry section (141 samples), in the Wapnica Quarry section near Dzikowiec (16 samples) and in the Gorzysław-9 borehole section (97 samples). Comparative data (93 samples) from the Chmielno-1 borehole section (Matyja et al. 2015) was also taken into consideration. Magnetic susceptibility (MS) was measured on Kappabridge KLY-2 (prod. Geofyzika, Brno) at a frequency of 920 Hz and a sensitivity of 4 × 10−8 SI. The obtained results were normalised to sample weight and expressed in cubic metres per kilogram. Rock magnetic measurements were performed in the Palaeomagnetic Laboratory of the Polish Geological Institute–National Research Institute.
Analytical sections
The presented high-resolution biostratigraphic analysis, encompassing the uppermost Famennian and lowermost Tournaisian strata together with the Hangenberg Black Shale (HBS) horizon, is based on a complete continuous pelagic sequence in the large active Kowala Quarry near Kielce in the Holy Cross Mountains and on the almost complete succession in the old non-active Wapnica Quarry near Dzikowiec village in the Sudety Mts, as well as on the relatively shallow ramp sequence in the Western Pomerania area (Gorzysław-9, Rzeczenica-1 and Chmielno-1 borehole sections) (Fig. 1). The presented interpretations use the results of recent palaeontological and biostratigraphic studies; when possible, previous biostratigraphic data was re-examined.
Kowala near Kielce—an overview of previous results
The area close to the village Kowala, located in the southern limb of the Gałęzice–Kowala syncline (Fig. 2a), is selected here as the principal reference site for the DCB studies.

a Simplified geological map of the Holy Cross Mountains with the position of the studied DCB section in the Kowala Quarry; b schematic plan of the Kowala Quarry (Google Maps: http://maps.google.com; October 18, 2018) with approximate location of the Kowala historical trench and the Kowala-1 borehole section (asterisk); c, d field images (photo B.A. Matyja) of the complete pelagic DCB succession (north-eastern level of the Kowala Quarry; compare Fig. 2b); lithological units after Malec (1995, 2014): A—nodular cephalopod limestone, B—shale (including the Hangenberg Black Shale horizon in the lower part of the unit), C—marly limestone, D—marl with nodular limestone intercalations; abbreviations: KBS—Kowala Black Shale, HBS—Hangenberg Black Shale
Firstly, Szulczewski (1971) followed by Berkowski (2002) sub-divided the upper Famennian part, exposed in the formerly Wola Quarry as well as in the adjacent railroad cut, into informal lithological sets H-2 to L (see also Racki et al. 2002). Independently, based on data from the historical trench exposed in the NE part of the Kowala Quarry (Fig. 2b), Malec (1995) sub-divided the DCB sequence into informal lithological units A to D, which are easily recognisable in the field (Fig. 2c, d) and have a correlation potential with the Rhenisch succession (comp. Fig. 3). [The upper part of set L sensu Berkowski 2002 is the equivalent of the lower part of set A sensu Malec 1995]. Later, De Vleeschouwer et al. (2013) proposed a new set M overlying set L, composed of a thick grey-greenish clayey/tuffite succession with several interbedded limestone layers [equivalent of the upper part of set B and set C sensu Malec 1995, 2014], and set N, which comprises a greenish clayey/tuffite interval with several thin black shale intercalations and some nodular carbonate horizons [equivalent of the lower part of set D sensu Malec].

Correlation of conodont, ammonoid and miospore zonations in relation to Rhenisch standard succession (after Becker et al. 2016, simplified)
Unit A sensu Malec (1995, 2014) consists mostly of alternating, cyclic, reddish-green weathering, grey nodular cephalopod limestones, and grey calcareous claystones, and is ca. 10 m thick (Fig. 2c, d and Fig. 4). The nodular beds vary between 3 and 10 cm in thickness and the calcareous claystones are between 1 and 6 cm thick. The Kowala black bituminous shale horizon (KBS), ca. 25 cm thick, dated as the LV or LL miospore Zone was recognised 3 m above the base of unit A (for details, see Marynowski and Filipiak 2007; comp. also Fig. 2c, d and Fig. 4). Just above the KBS horizon, nodular limestones display bioturbation (Fig. 4). Nodular limestones of unit A are mostly mud-wackestones with predominant cephalopods, but many corals, represented by rugosans, tabulates and heterocoralls, were described by Różkowska (1969), Berkowski (2002), Zapalski and Berkowski (2012), Berkowski et al. (2016) and Zapalski et al. (2016). Crinoids were described by Głuchowski (2002), and brachiopods by Halamski and Baliński (2009). Among the 23 taxa analysed by Halamski and Baliński (2009) from the middle and upper part of the Famennian (set J and K; see Szulczewski 1971; Berkowski 2002) and from the upper to uppermost Famennian (set L), only four are common between the two analysed faunas. Halamski and Baliński (2009) emphasised that the species common between the faunas of set J–K and L belong to the Rhynchonellida and Orthida. The latest Famennian brachiopod fauna from set L consists mainly of the species Schellwienella pauli Gallwitz, Aulacella interlineata (Sowerby), Rozmaria equitans (Schmidt) and Sphenospira julii (Dehée). Unit A is also characterised by the presence of small gastropods, abundant conodonts, benthic and planktonic ostracodes (Olempska 1997), and a relatively large amount of agglutinated foraminifers (Woroncowa-Marcinowska 2017). Some remarks about the macrofauna from the Wocklumeria limestone were also provided by Rakociński (2011).

The DCB interval in the Kowala Quarry (Holy Cross Mountains)—lithological log with position of conodont samples, biostratigraphy and conodont distribution
The uppermost part of unit A (ca. 1 m thick), lying directly below the HBS, consists of green and yellow thin-bedded marly limestones intercalated with shales and volcanic ash layers (Marynowski and Filipiak 2007; Marynowski et al. 2012).
Above unit A occurs a ca. 2.4-m-thick horizon (unit B sensu Malec 1995, 2014) of black bituminous shales with very thin layers of grey marly shales (ca. 1.2 m thick), overlain by light-grey marly shales, which interrupt the carbonate succession (Figs. 2c, d and Fig. 4). This black bituminous shale horizon, which contains compounds characteristic of photic zone euxinia, occurrence of wildfires, as well as volcanogenic material, was recognised as the equivalent of the global Hangenberg Black Shale (HBS) horizon by Marynowski and Filipiak (2007) and Marynowski et al. (2012). The most abundant fossil in the HBS is the bivalve Guerichia; cephalopods as well as entomozoids are common in some samples. The occurrence of volcanic ash layers within the HBS succession, as well as just below and above this horizon, was observed by Marynowski and Filipiak (2007) and Marynowski et al. (2012). Myrow et al. (2013, fig. 1 and 2) presented geochronological data for three volcanic ash beds, situated directly below and above the HBS horizon, that provide constraints on the age and short duration of the Hangenberg Event, between 358.97 ± 0.11 and 358.89 ± 0.20 Ma.
The overlying deposits, ca. 1.3 m thick, are composed of ca. 30 cm yellow–grey massive marly micritic limestones in the lower part of the horizon, reddish laminated limestones in its middle part and yellow–grey micritic limestones in the upper part (unit C sensu Malec 1995, 2014, Figs. 2c, d and Fig. 4). At the top of this package occur isolated limestone nodules in a layer within an approximately 26-cm-thick, yellow–grey marly claystone bed.
Above the limestones occurs a thick package (ca. 18 m thick) of green–grey marls with thin beds of grey micritic, sometimes nodular limestones (Radlin beds sensu Żakowa and Pawłowska 1961; unit D sensu Malec 1995, 2014).
Czarnocki (1928, 1933, 1989) recognised and described this part of the uppermost Famennian in the Kowala trench I and III (presently not existing), located near the active Kowala Quarry. He noted the presence of a black shale unit (0.81 m thick) with Imitoceras, underlain by a marly, often nodular limestone and shale unit (about 6.5 m thick), referred to by Czarnocki as the “Wocklumeria beds” (= “Stufe” in the German type area) and sub-divided into the Lower and Upper Wocklumeria beds. Czarnocki collected rich and diverse ammonoid fauna from the Wocklumeria beds. Unfortunately, he indicated the position of each ammonoid species in the Lower and Upper Wocklumeria beds only.
The first more detailed biostratigraphic data comes from the Kowala-1 borehole section, located close to the present-day Kowala Quarry (Fig. 2b). The uppermost part of the Famennian was investigated in a number of aspects: biostratigraphic (Żakowa et al. 1985; Żakowa and Radlicz (1990); Nehring-Lefeld 1990), palynostratigraphic (Turnau 1985, 1990) and lithological (Romanek and Rup 1990). Turnau (1990) recognised the LL miospore Zone and LN Zone within the black bituminous shale horizon (HBS), with index taxa Retispora lepidophyta, Vallatisporites verrucosus and Verrucosisporites nitidus, and the VI Zone above the HBS, with Vallatisporites vallatus and Vallatisporites pusillites. Conodonts identified just a few metres below the HBS, in the uppermost part of the nodular limestone unit, indicated as Bispathodus costatus, Bispathodus stabilis, Palmatolepis gracilis gracilis, Palmatolepis gracilis sigmoidalis and Pseudopolygnathis marburgensis trigonicus, were assigned by Nehring-Lefeld (1990) to the Upper expansa–Lower praesulcata ‘standard’ conodont zones of Ziegler and Sandberg (1984).
The pelagic Kowala sequence was also investigated in a trench, dug by Malec, located in Kowala village, close to the northern wall of Kowala Quarry (Fig. 2b). As previously, samples from the trench were investigated for biostratigraphy and lithology (Malec 1995; Dzik 1997; Olempska 1997), as well as palynology (Filipiak 2004, 2005). Based on the occurrence of conodonts, ammonoids and bivalves, the bottom part of the exposed section (units A and B sensu Malec 1995) was included in the expansa ‘standard’ conodont Zone and to the lower part of the praesulcata Zone. The overlying deposits, above the cephalopod limestone with Wocklumeria, were assigned to the sulcata Zone and succeeding conodont zones belonging to the Tournaisian. Accordingly, the DCB was indicated within unit D sensu Malec (2014) (Radlin beds). Unfortunately, palynologic remains in the critical DCB interval were partly weathered, although the LE and LN miospore zones were tentatively recognised just below the DCB (for details, see Filipiak 2004). Strong condensation of small acritarchs, just above the DCB, was noted (see Filipiak 2005).
The most abundant fossil in the HBS horizon is the bivalve Guerichia, with up to 7000 specimens determined as Guerichia venusta (Münster) being present in a square metre (Marynowski et al. 2012). Bivalves of the former “Posidonia venusta” group occur widespread within pelagic limestones and Culm facies in the uppermost Devonian to the lowermost Mississippian strata. Earlier attempts for a biostratigraphic zonation in Kazakhstan by Sadykov (1962) and later in Poland by Żakowa (1983) were successfully adapted to comparable facies in Europe (Amler 2004). Sadykov (1962) showed that the evolution of this group in Kazakhstan displays a development from short, compact forms [such as Guerichia nalivkini (Sadykov, 1962) and G. simorini (Sadykov, 1962)] in the early Famennian to elongate–triangular forms [such as Guerichia venusta (Münster, 1840)] in the late Famennian, and finally to elongate–transverse taxa [such as Guerichia venustiformis (Sadykov, 1962) = ?G. ratingensis (Paeckelmann, 1913)], known from the Wocklum limestone and the Culm facies. The oval Guerichia mariannae (Tschernyschev, 1941) is characteristic of the lower Mississippian or upper Tournaisian.
A diverse Famennian assemblage of agglutinated foraminifers from the Kowala trench was first documented by Olempska (1983) and Malec (in Żakowa et al. 1985). Most of the recognised species belong to the genera Hyperammina, Thurammina and Tolypammina. The stratigraphic position of this fauna corresponds to the upper part of the entomozoid hemisphaerica–dichotoma Zone and to the costatus ‘standard’ conodont Zone (Olempska 1983). A comparably rich collection of agglutinated foraminifers, studied by Woroncowa-Marcinowska (2017), comes from the nodular cephalopod limestone (= unit A sensu Malec 1995, 2014) in the Kowala Quarry. The foraminiferal assemblage (Woroncowa-Marcinowska 2017, fig. 3, pl. 1, 2) includes forms with free tests such as Hyperammina, Reophax, Psammosphaera, Pseudoastrorhiza, Thurammina, Paratikhinella and Septatournayella, as well as sessile forms, such as Tolypammina and Moravammina. Paratikhinella cannula, Tolypammina rotula and Hyperammina stabilis occur nearly in all samples. Agglutinated foraminifers do not appear to provide a basis for a detailed biostratigraphic scheme. Most of the species are long-ranging taxa, known from the entire Famennian and Tournaisian. However, the stratigraphic ranges of some species, e.g. Hyperammina aperta and Thurammina tubulata fixa, are limited to the uppermost part of the Famennian, to the costatus ‘standard’ conodont Zone (Olempska 1983, p. 395). Moravammina? constricta occurs also only in the uppermost part of the Famennian, in the Dasberg- and Wocklum-Stufe (Eickhoff 1970). Taking into account that some sedimentary environments are well characterised by typical foraminiferal associations, Gutschick and Sandberg (1983), Balthasar and Amler (2003) and Herbig (2006) distinguished three distinctive deeper-water associations of agglutinated foraminifers at the end of the Famennian and during the Tournaisian. The distribution of agglutinated foraminifera taxa in the Kowala section was examined at generic level to compare with the proposed models. The red nodular cephalopod limestone (Fig. 4, interval between samples 8–18) contains the mixed saccaminid and Hyperammina biofacies, in which the more “shallow” saccaminid elements constitute about 60% of the total foraminiferal assemblage, and the “deeper” Hyperammina contributes to about 30%. In the overlying green–grey limestone situated below the HBS horizon (compare Fig. 4, interval between samples 19–22), elements of the Hyperammina biofacies increase to about 50%, and a subtle biofacies shift to a deeper one is observed.
Kowala near Kielce—new results
Conodont biostratigraphy
As a contribution to the ongoing international revision of the DCB, new rock material for the presented biostratigraphic analysis was collected from the north-eastern wall of the Kowala Quarry (Fig. 2b) in 2010 and 2013. Due to the north-eastward extension of the exploitation front, a very well-preserved part of the Upper Famennian to lower Tournaisian succession has been exposed in 2018, and the upper part of the section (units B, C and lower part of the unit D sensu Malec 2014) was re-sampled (Fig. 2c, d). Our new conodont record in the Kowala Quarry section, based on 35 new samples, as well as conodonts from the collection of Woroncowa-Marcinowska (2017), is presented herein (Figs. 4, 5, 6, and 7; Table 1). Some conodont data of Malec (1995, 2014), the accuracy of which is not doubtful, were also incorporated in the biostratigraphic analysis (comp. Fig. 4, data of Malec (2014) indicated as asterisks).


a Conodonts of the ultimus Zone (sensu Kaiser et al. (2009) and Becker et al. (2016), Kowala Quarry (Holy Cross Mountains). a, Palmatolepis gracilis gonioclymeniae Müller, 1956, sample 3, oral view. b, Palmatolepis gracilis gracilis Branson and Mehl, 1934a, sample 18, oral view. c, Palmatolepis gracilis sigmoidalis Ziegler, 1962a, sample 8, oral view. d, Protognathodus sp. (aff. meischneri Ziegler, 1969), a very untypical form with a narrow strongly bended cup and an extended posterior tip, sample 11, oral view. e, Bispathodus aculeatus aculeatus (Branson and Mehl, 1934a), sample 18, oral view. f, Bispathodus costatus (Branson, 1934), sample 18, oral view. g, Bispathodus ultimus (Bischoff, 1957), sample 18, oral view. h and i, Pseudopolygnathus micropunctatus Bischoff and Ziegler, 1956, sample 8, aboral and views. j, Branmehla suprema (Ziegler, 1962a), sample 18, oral view. k, Bispathodus spinulicostatus (Branson, 1934), sample 5, oral view. l, Branmehla inornata (Branson and Mehl, 1934a), sample 18, lateral view. m, Mehlina strigosa (Branson and Mehl, 1934a), sample 8, lateral view. n, Neopolygnathus cf. vogesi (Ziegler, 1962a), sample 12, oral view. o and r, Neopoplygnathus communis communis (Branson and Mehl, 1934b); o, sample 12; p–r, sample 8, oral views. s and t, Bizignathus kayseri (Bischoff and Ziegler, 1956), sample 8, oral and aboral views. b Conodonts of the ultimus Zone (sensu Kaiser et al. (2009) and Becker et al. (2016), Kowala Quarry (Holy Cross Mountains): a, Pseudopolygnathus marburgensis trigonicus Ziegler, 1962a, sample 16, oral view. b–d, Pseudopolygnathus primus Branson and Mehl, 1934, sample 8, oral views. e, Pseudopolygnathus ostrovkensis Dzik, 1997, sample 8, oral view. Conodonts intermediate between polygnathids, pseudopolygnathids and siphonodellids: f and g, cf. N. Gen. 2 sensu Becker et al. (2013), sample 12, oral and aboral views. h, cf. N. Gen. 2 sensu Becker et al. (2013), sample 13, aboral view. i and j, “Pseudopolygnathus” graulichi Bouckaert and Groessens, 1976, sample 16, oral and aboral views. k and l, ?Polygnathus sp., from the oral view looks like Polygnathus and has a Polygnathus-type pit but is also characterised by a curvature of the platform similar to the genus Siphonodella, sample 19, oral and aboral views

Conodonts of the ?praesulcata Zone (sensu Kaiser et al. 2009 and Becker et al. 2016), Kowala Quarry (Holy Cross Mountains): a Palmatolepis gracilis sigmoidalis Ziegler, 1962a, sample 20, oral view. b Palmatolepis gracilis gracilis Branson and Mehl, 1934a, sample 22, oral view. c Palmatolepis gracilis expansa Sandberg and Ziegler, 1979, sample 20, oral view. d Bisphatodus aculeatus aculeatus (Branson and Mehl, 1934a), sample 20, oral view. e Bispathodus costatus (Branson, 1934), sample 20, oral view. f Bispathodus stabilis vulgaris Dzik, 2006, sample 22, lateral view. g and h Branmehla inornata (Branson and Mehl, 1934a), sample 19, lateral views. i and j Branmehla suprema (Ziegler, 1962a); i sample 22; j sample 30, oral views. k Branmehla disparilis (Branson and Mehl, 1934a), sample 22, oral view. l cf. Siphonodella (Eosiphonodella) sp., anteriorly broken specimen, sample 22, oral view

Conodonts of the ?kockeli (a–c, g–i) and sulcata/kuehni zones (d–f) (sensu Kaiser et al. 2009 and Becker et al. 2016), Kowala Quarry (Holy Cross Mountains): a–c ?Protognathodus sp.? similar to Bispathodus aculeatus, sample 30, oral and lateral views. d and e Siphonodella sulcata (Huddle, 1934) M5 after Kaiser and Corradini (2011), sample 35, aboral and oral views. f Neopolygnathus communis communis (Branson and Mehl, 1934b) > Neopolygnathus vogesi (Ziegler, 1962a), sample 35, oral view. g and h Polygnathus purus purus Voges, 1959, sample 30, oral views. i Polygnathus purus subplanus Voges, 1959, sample 35, oral view
The uppermost Famennian to lower Tournaisian Standard Conodont Zonation of Ziegler and Sandberg (1984, 1990) and Sandberg et al. (1978) has been extensively updated in the last three decades. Alternative zonal definitions and zonal schemes were proposed by some authors (for details, see discussion in Ji 1985; Corradini 2008; Kaiser et al. 2009; Kaiser and Corradini 2011; Hartenfels 2011; Hartenfels and Becker 2016; Becker et al. 2016; Corradini et al. 2017; Spaletta et al. 2017). Among the proposed new zonal schemes, the zonation of Kaiser et al. (2009) and Becker et al. (2016) is readily applied in the presented pelagic sections in Poland. A very useful review of the stratigraphic ranges of common and widespread Famennian conodont species and subspecies is given by Corradini et al. (2017) and Spaletta et al. (2017).
The lower part of unit A (samples 1–2) yielded Bispathodus bispathodus, Polygnathus inornatus, Polygnathus znepolensis and Palmatolepis rugosa ampla, as well as different representatives of the Palmatolepis gracilis group but without Palmatolepis gracilis gonioclymeniae. This part of unit A probably belongs to the costatus Subzone of the aculeatus Zone (Fig. 4).
The first occurrence of Palmatolepis gracilis gonioclymeniae in sample 3 characterises the lower boundary of the ultimus Zone. This part of the Kowala section, located between samples 3–19, contains abundant and widespread uppermost Famennian taxa (e.g. the Palmatolepis gracilis group, Pseudopolygnathus marburgensis trigonicus, several branmehlids, and Bispathodus costatus-ultimus group), as well as strange representatives of different conodont taxa (comp. Fig. 4). The entry of these forms took place within the ultimus Zone. First Protognathodus sp. (? aff. meischneri) enters in sample 11 (Fig. 4). This is a very untypical form, strongly bent, with a narrow cup and an extended posterior tip (Fig. 5a(d)), similar to the specimens noted by Kaiser et al. (2019, pp. 12-13) from Trolp (Palaeozoic of Graz). Very specific are specimens intermediate between the polygnathids, pseudopolygnathids and siphonodelloids (comp. Tragelehn 2010). Specimens with a flat pseudokeel, which continues to the posterior tip (samples 12 and 13; Fig. 5b(f, g, h)), may belong to N. Gen. 2 sensu Becker et al. (2013). Some forms with a relatively wide cavity resemble Pseudopolygnathus (?“Pseudopolygnathus” graulichi—sample 16, Fig. 5b(i, j)). The form illustrated as “k” and “l” in Fig. 5b (sample 19) in oral view looks like Polygnathus and have a Polygnathus-type pit in aboral view, but is characterised by a curvature of the platform similar to the genus Siphonodella. The upper boundary of the ultimus Zone and the base of the praesulcata Zone is marked by the first appearance of Siphonodella (Eosiphonodella) praesulcata in the top of unit A (data of Malec 2014—his beds 142 and 148; see fig. 6A). However, to be sure, the lower views of these specimens are important for a final evaluation; therefore, there are still some biostratigraphic uncertainties concerning the determinations of Malec. The first Siphonodella (Eosiphonodella) in our collection (unfortunately anteriorly broken specimen) is noted in the sample 22 (Fig. 4 and Fig. 6l). Conodont faunas from samples 21–22 (Figs. 4 and 6, Table 1) include the undoubted last occurrences of typical pre-Hangenberg taxa, such as Bispathodus bispathodus, Palmatolepis gracilis gracilis, Palmatolepis gracilis expansa, Bispathodus costatus, Palmatolepis gracilis gonioclymeniae, Bispathodus ultimus, Branmehla suprema, Pseudopolygnathus marburgensis trigonicus and Branmehla disparilis. The last occurrence of this conodont fauna at the base of the HBS horizon could define the base of the costatus-kockeli Interregnum (c-kI) sensu Kaiser et al. (2009), as the following conodont samples (23a-29) from unit B (= HBS together with the overlying grey shale horizon) contain no conodonts. The absence of conodonts could be due to lithofacies, as discussed by Kaiser et al. (2019).
Within unit C of Malec (2014), beginning with sample 30, the re-occurrence of long-ranging taxa (survivors such as Neopolygnathus vogesi, Bispathodus stabilis and Neopolygnathus communis communis) is observed together with the onset of some new taxa, e.g. ?Protognathodus sp. and Polygnathus purus purus in very small numbers. Characteristic is the absence of Protognathodus kockeli in our assemblage; Dzik (1997), fig. 2, table 3, p. 60), however, noted the first representatives of Protognathodus kockeli in his samples Ko-51 and Ko-24, located between 2.2 and 2.4 m above the top of the Wocklumeria limestone. It is not easy to indicate the estimated position of this species found by Dzik in relation to our section, but its position within unit C is very likely. It is possible that lithological unit C belongs to the kockeli conodont Zone.
The first Siphonodella sulcata, represented by morphotype 5 sensu Kaiser and Corradini (2011), was noted by Malec (2014, fig. 6E) from bed 184, located about 4 m above the base of unit D (comp. Fig. 4). We have found this species (Fig. 7d, e) together with Polygnathus purus subplanus in our bed 35 (Fig. 7i), at 3 m above the base of unit D, below the data of Malec. Both these species can be used for the approximation of the DCB in the Holy Cross Mountains and have a potential to correlate the Polish sections with other regions where siphonodellids or even only protognathodus fauna (with Protognathodus kuehni) exist.
Ammonoid biostratigraphy—re-examination of the ammonoid fauna collected by Czarnocki (1989)
Woroncowa-Marcinowska attempted to find an estimated position of the ammonoid fauna collected by Czarnocki [collection number MUZ PIG 284.II] in relation to our Kowala Quarry section (see Fig. 4, interval between samples 7–29 = Wocklumeria beds sensu Czarnocki 1989), comparing individual ammonoid specimens with the matrix attached with some characteristic details of lithological units observed in our section. Some important taxa are illustrated in Fig. 8). Apart from some exceptions, however, it was very difficult to determine the precise location of ammonoid fauna collected by Czarnocki (comp. Table 2).

Some typical prionoceratid and wocklumeriid ammonoids from the uppermost Famennian of the Kowala Quarry. a–e Effenbergia lens Korn, 1992: a–c MUZ PIG 284.II.385b, × 4; d MUZ PIG 284.II.385c, × 4,5; e MUZ PIG 284.II.385a, × 4; f–h Effenbergia minutula Korn, 1992, MUZ PIG 284.II.385d, × 3.5. i–k Kenseyoceras nucleus (Schmidt, 1924), MUZ PIG 284.II.727b, × 3.5. l and m Kenseyoceras rostratum Selwood, 1960, MUZ PIG 284.II.727a, × 2.8. n and o Balvia globularis (Schmidt, 1924), MUZ PIG 284.II.727c, × 2.7. p–r Epiwocklumeria applanata (Wedekind, 1918), × 1: p MUZ PIG 284.II.728c; q, r MUZ PIG 284.II.728e
In general, red limestones and shales dominate in the Lower Wocklumeria beds sensu Czarnocki (our section = interval between samples 7–16; see Fig. 4), and the unit was sub-divided by Czarnocki into three lithological sub-units. Set 1 (about 1.3 m thick) was characterised by red-grey variegated nodular limestone (our section = interval between samples 7–11; see Fig. 4), set 2 (about 1.6 m thick) was characterised by red nodular limestone (our section = interval between samples 12–14; see Fig. 4), and set 3 (1.3 m thick) was characterised by red–grey–green variegated limestone (our section = interval between samples 15–16).
The Lower Wocklumeria beds of Czarnocki (1989) are characterised by rich and diverse clymeniids. Among 29 species, local clymeniid fauna constitutes a significant component of the total ammonoid assemblage (comp. Table 2). The majority of the listed species is well known from the uppermost Famennian or Wocklumian (Schindewolf 1937). The list includes forms such as Muessenbiaergia sublaevis and M. bisulcata that can be correlated with UD VI A and with UD VI B, characterised here by “Glatziella glaukopis”, Kosmoclymenia tenuissima, Muessenbiaergia kowalensis and Trochoclymenia wysogorskii (Frech) (compare Becker et al. 2016). Representatives of the genus Glatziella, as well as the species Effenbergia lens (Korn) (see also Woroncowa-Marcinowska 2011), indicate the Effenbergia lens Zone (UD VI-B). Three specimens in the collection are registered under a single number 777 and named as “Glatziella glaukopis”. According to the opinion of TW-M, two of them (illustrated in Czarnocki 1989, pl. III, fig. 8 and probably coming from set 2) belong to the species Glatziella helenae (Renz), and only one (Czarnocki 1989, illustrated in pl. III, figs. 1, 2, 6) represents Glatziella glaukopis. In the opinion of Becker and Mapes (2010), the genera described by Czarnocki as Dimeroclymenia and Liroclymenia belong to the genus Rhiphaeclymenia. They supported the opinion of Dzik (2006) that Liroclymenia fundifera is rather closely related to Rhiphaeclymenia canaliculata Bogoslovsky. The latter species is known from the Kalloclymenia–Wocklumeria genozone in the southern Urals (Bogoslovsky 1981). Dzik (2006) suggested also that the three species of Dimeroclymenia created by Czarnocki as new species (pristina, semicostata and subacuta) are conspecific, and considered them as representing Biloclymenia pristina (Czarnocki) (red limestone of the Dasbergina trigonica Zone sensu Dzik 2006; this paper = above sample 11; see Fig. 4; possibly UD VI-B of Becker et al. 2016). Czarnocki (1989) created six new species within the genus Kielcensia. He indicated three levels, where K. bohdanowiczi, K. pisilla, K. inaequilobata and K. angustilobata were found (sets 1–3). It seems correct to agree with the opinion of Dzik (2006) that K. bohdanowiczi appeared probably in his late P. jugosus Zone (perhaps UD VI-B of Becker et al. 2016). One representative of Wocklumeria sphaeroides (Richter) from the Lower Wocklumeria beds (Czarnocki 1989, table 3) was mentioned incorrectly (see Czarnocki 1989, table 2). The presence of Kamptoclymenia aff. endogona (Schindewolf) in the Lower Wocklumeria beds of Czarnocki (1989) characterises UD VI-CI (compare Becker et al. 2016, fig. 4).
The ammonoid faunas from the Lower Wocklumeria beds sensu Czarnocki (1989) (our section = interval between samples 7–16) can be correlated with the genozones Linguaclymenia (UD VI-A), Effenbergia (VI-B) and lower Parawocklumeria (VI-C1) of Becker et al. (2016).
Green limestones and shales prevail in the Upper Wocklumeria beds sensu Czarnocki (our section = interval between samples 17–22; see Fig. 4); they were sub-divided into two lithological sets. The lower part of the Upper Wocklumeria beds (set 1) is composed of green–grey nodular limestone (about 1.3 m thick) and grey irregular nodular limestone and shale (1.6 m thick) (our section = interval between samples 17–20; see Fig. 4).
The upper part of the Upper Wocklumeria beds (set 2 sensu Czarnocki) is composed of dark grey, thick bedded nodular limestone (0.6 m thick) (our section = interval between samples 21–22, comp. Fig. 4).
In this part of the section, Czarnocki (1989), table 3) distinguished 15 species of clymeniids (Table 2). The first three species were found within set 1 (our section = sample 17; see Fig. 4). Parawocklumeria paradoxa appears here for the first time and may indicate the P. paradoxa Zone (UD VI-C2 of Becker et al. 2016). The appearance of Wocklumeria sphaeroides (Richter) and other representatives of the genus Wocklumeria in grey irregular nodular limestone (set 1 of Czarnocki; our section = interval between samples 18–20, see Fig. 4) indicates the Wocklumeria genozone (UD VI-D1 of Becker et al. 2016). The appearance of Epiwocklumeria applanata Wedekind together with the mentioned assemblage (set 2 of Czarnocki; our section = interval between samples 21–22; see Fig. 4) indicates the upper part of the Wocklumeria genozone (UD VI-D2 of Becker et al. 2016). Moreover, together with Epiwocklumeria applanata, Woroncowa-Marcinowska (2011) identified also in Czarnocki’s collection (specimen 284.II.727) the species Sporadoceras terminus Dzik, Kenseyoceras nucleus (Schmidt), K. rostratum Selwood and Balvia globularis (Schmidt).
Ammonoid fauna from the HBS interval was studied by Czarnocki (1989), Dzik (1997, 2006), Malec (1995, 2014) and Marynowski et al. (2012). All specimens are flattened and occur as imprints. Czarnocki (ibid.) indicated “Imitoceras” and Cymaclymenia. Dzik (1997) found Acutimitoceras prorsum (Schmidt) at the level of 1.2 m above the top of the Wocklumeria limestone, in the tuffite bed above the HBS horizon, as well as 1.5 m above the HBS horizon (unit B sensu Malec 1995, 2014). Marynowski et al. (2012, fig. 5) illustrated some cymaclymeniids from the HBS horizon, wrongly identified as “Platyclymenia sp.” Malec (1995, 2014) mentioned abundant Acutimitoceras in a 22-cm-thick bed of claystones (his bed 159).
Numerous new, poorly preserved (as imprints) ammonoids, represented probably by Postclymenia evoluta Schmidt and Acutimitoceras sp. (comp. Fig. 9a–e), as well as bivalve Guerichia and other fauna and plants (Fig. 9g) were found by TW-M between samples 22–24 on a bedding surface of the HBS interval. The well-preserved suture (Fig. 9f), similar to Postclymenia evoluta, was also found on the bedding surface (specimen 284.II.349 in Czarnocki’s collection). Guerichia protobecheri djailmensis (Sadykov, 1962), G. venustiformis (Sadykov, 1962) and G. aff. mariannae (Tchernyshev, 1941) were also recognised in the Kowala Quarry section (Fig. 10) in the HBS horizon. Sadykov (1962) considered all these species as characteristic of the Guerichia venustiformis (= G. ratingensis?) Zone.

Poorly preserved ammonoids and plant imprint from the Hangenberg Black Shale (HBS) horizon, Kowala Quarry. a Acutimitoceras sp., × 1. b–e Imprints of shells, similar to Postclymenia evoluta Schmidt, 1924 or c, d Cymaclymenia sulcata Czarnocki 1989 (= C.?striata in Nikolaeva and Bogoslovsky 2005), b, d × 1,5; c, e × 1. f Imprints of shell with suture typical for Postclymenia evoluta, ×1,5, MUZ PIG 284.II.349. g Plant imprint, × 1

Macrofauna from the HBS horizon, Kowala Quarry. a Brachiopods (rhynchoneliids). b–i Bivalve Guerichia (fragments of valves with impressions): b, g G. djailmensis (Sadykov, 1962), × 1.5. c, h G. venustiformis (Sadykov, 1962), × 2.5. e G. cf. venustiformis (Sadykov, 1962), × 2.5. d Accumulation of moulds of G. aff. mariannae (Tchernyshev, 1941) valves, × 2.5. f, i G. aff. mariannae (Tchernyshev, 1941), i representative of G. aff. mariannae (Tchernyshev, 1941), twice enlarged from d
The ammonoid faunas from the Upper Wocklumeria beds sensu Czarnocki (1989) (our section = interval between samples 22–29; see Fig. 4) can be correlated with UD VI-E (HBS) and UD VI-F of Becker et al. (2016).
Microflora from the DCB interval—overview of previous results and some new results
The critical part of the DCB interval in the Kowala section was intensively tested for palynology; unfortunately, not all samples were positive (e.g. Filipiak 2004, 2005; Marynowski and Filipiak 2007; Filipiak and Racki 2010). Unfortunately, palynologic remains in the critical DCB interval were partly weathered, although the LE and LN standard western European miospore zones (Streel et al. 1987) were tentatively recognised just below the DCB (for details, see Filipiak 2004). The best-preserved microflora generally came from the HBS or directly sub-/superjacent layers. New palynological results, unpublished up till now, revealed also the presence of the VI miospore Zone (base of the Tournaisian) just a few metres above the HBS, within the Radlin beds (set D sensu Malec 2014) and the next HD miospore Zone within the Zaręby beds (set E sensu Malec 2014). In the Kowala Quarry, the Zaręby beds are noted about 14 m above the HBS horizon, and can be treated as an equivalent of the globally known Tournaisian Lower Alum Shale horizon.
Microflora obtained from the HBS horizon indicates the presence of the Retispora lepidophyta–Verrucosisporites nitidus (LN) miospore Zone (Fig. 11). Both index species are represented by single specimens and constitute < 1% of the assemblages. The rare occurrence of these important taxa was noticed previously in other sections, from the LN Zone as well (Filipiak 2004). Other, much more frequent, miospores noticed in this palynological assemblage are typical of the LN miospore Zone and are represented by Apiculiretusispora verrucosa, Bascaudaspora submarginata, Cymbosporites minutus, Diducites versabilis, Grandispora echinata, G. lupata, Indotriradites explanatus, Kraeuselisporites mitratus, Pustulatisporites dolbii, Retusotriletes incohatus, Tumulispora malevkensis, T. rarituberculata, Umbonatisporites rarisetosus, Vallatisporites verrucosus, V. vallatus and V. pusillites. The most common miospores are Vallatisporites spp. (43.5%), Retusotriletes incohatus (15.5%) and Apiculiretusispora verrucosa (10.6%; Marynowski and Filipiak 2007). Peculiar is the presence of Vallatisporites vallatus in this level. In Western Europe, this species appears in the next, Tournaisian VI miospore Zone (Clayton et al. 1977; Clayton and Turnau 1990). In Belarus, however, this taxon is also present in the PM local Zone, which is the stratigraphic equivalent of the standard LN level (see Avkhimovitch 1993; Avkhimovitch et al. 1993). Therefore, it may indicate that, similarly as in Eastern Europe, this species appears in the Holy Cross Mountains diachronically earlier compared to Western Europe.

The DCB interval in the Kowala Quarry section with the main floristic events and possible biocrises. A!—concentration of small acritarch species (“Micrhystridium” complex, Veryhachium and Unellium); T!—concentration of abnormal miospores and undivided miospore tetrads; L.RL!—last recorded occurrence of Retispora lepidophyta. *palynology based on Turnau (1985, 1990), Filipiak (2004), Marynowski and Filipiak (2007) and Filipiak and Racki (2010). HBS—Hangenberg Black Shales, KBS—Kowala Black Shales
The sample taken from the overlying HBS, thin (10 cm) claystone layer, just below the distinctive tuffite layer, had a very interesting content (Marynowski and Filipiak 2007; Filipiak and Racki 2010). It yields a palynologically distinctive assemblage of abnormal miospores and undivided tetrads. The quantity of aberrant spores and pollen exceeded 3% of the entire palynological assemblage. It was interpreted as a signal of environmental stress (for details, see Filipiak and Racki 2010, Fig. 11). According to Foster and Afonin (2005), percentages above 3% of abnormal pollen are used as a proxy to monitor recent air pollution. Therefore, a comparable abundance of mutated palynoflora can be treated as a signal of fertilisation failure in stressed ancient habitats. Tetrad and abnormal pollen enrichment in the Kowala succession is accompanied by volcanic ash levels. Additionally, large amounts of charcoal debris and polycyclic aromatic biomarkers were noticed in this section, which is indicative of forest wildfires (for details, see Marynowski and Filipiak 2007). Accumulation of abnormal miospores together with undivided tetrads could reflect the mutagenic effect of regional(?) environmental acidification and/or other stresses due to e.g. volcanic eruptions (Fig. 11).
Abnormal miospores have been reported many times from strata close to the DCB in different areas, e.g. Combaz and Streel (1971); Brévilliers, northern France), Chibrikova et al. (1978, localities in the Russian Platform), Higgs and Streel (1994); Sauerland, Germany), Filipiak and Racki (2010), Holy Cross Mountains, Poland) and Prestianni et al. (2016), Chanxhe sections, Belgium). Kedo (1957) and later Streel (1966) noted that Retispora lepidophyta, a very important taxon for Upper Devonian palynostratigraphy, varied in size (see also Maziane et al. 2002) and, furthermore, frequently in morphology by the appearance of abnormal forms. Based on this morphological variability, Kedo (1957, 1963) introduced two forms: R. lepidophyta var. minor and R. lepidophyta var. tener. The former one in its extremity is characterised by a presence of just a few crests and/or verrucae instead of well-developed muri of the reticulum observed in the minor form. Streel (1966) noticed similar abnormal forms in Retispora and referred to them as R. lepidophyta type a. Recently, Prestianni et al. (2016) recognised and described morphological changes (mutations) in a broader miospore assemblage from the uppermost Famennian of the Belgium area. Based on the biostratigraphic importance of R. lepidophyta var. tener, they proposed to name the occurrence of a large population of abnormal miospores just before the DCB as the tener Event. According to them, the tener Event is possibly the continental equivalent of the Hangenberg Biocrisis (for further details, see Prestianni et al. 2016).
The next palynological data in the Kowala section is noted a few metres above the HBS, from the Radlin beds (unit D sensu Malec 2014), developed as marls and bright-coloured shale. The collected samples are Tournaisian in age and represent the VI (Vallatisporites verrucosus-Retusotriletes incohatus) miospore Zone (Fig. 11). The assemblage is low diversity and does not possess typical upper Famennian taxa, such as Retispora lepidophyta or Diducites spp. The palynological assemblage contains Aurorasopra spp., Convolutispora spp., Discernisporites micromanifestus, Lophozonotriletes excisus, Plicatispora scolecophora, Retusotriletes incohatus, Tumulispora spp. and Vallatisporites spp.
In the historical Kowala trench, approx. 2 m above the HBS, concentrations of small acritarch taxa were noticed (see Filipiak 2005, Fig. 11). Among them predominate taxa from the Micrhystridium “complex” together with Unellium and Veryhachium (Fig. 11). A similar composition of phytoplankton taxa in the lower Tournaisian samples was observed also in Wales by McNestry (1988) and in the Stockum section in the Rhenish Slate Mountains (Streel 1999). This event could have a correlation potential for the base of the lower Tournaisian. Current investigations in the Kowala Quarry did not confirm this observation, probably due to strong weathering of microflora, especially in samples from marls and limestones.
Wapnica Quarry near Dzikowiec (Western Sudetes)—an overview of previous results
The abandoned Wapnica Quarry, located near Dzikowiec village on the western slope of Wapnica Hill in the north-western margin of the Bardo Mts (Fig. 12a), is well known from the partly exposed, Upper Devonian to lower Carboniferous succession of unmetamorphosed rocks, representing one of very few fragments of the sedimentary cover deposited directly on a Lower Devonian oceanic crust (e.g. Dopieralska et al. 2006). The basal part of this cover is developed as large blocks of gabro embedded in the carbonate matrix, covered by carbonates, included in the Main Limestone and “Clymenia beds” by Guerich (1902), and dated as ?Frasnian to lower Mississippian (for details, see Berkowski 2002).

a Simplified geological map of the Sudety Mountains (after Muszer and Haydukiewicz 2011, simplified) with the position of the studied DCB section in the Wapnica Quarry near Dzikowiec; b schematic plan of the Wapnica Quarry; asterisks—location of the studied sections along the eastern quarry walls; c field image (photo, T. Woroncowa-Marcinowska) of the pelagic DCB succession (NE part of the Wapnica Quarry near Dzikowiec); lithological units: Clymenia Limestone and Gattendorfia Limestone with distinctive black shale intercalation (HBS) and sharply overlying Viséan grey sandstone; d HBS horizon = black shale and nodular limestone horizon
The DCB interval belongs to the upper part of the informal lithostratigraphic unit, named the Wapnica “formation” by Wajsprych (1995). This part of the Wapnica “formation” consists of the so-called clymeniid limestone and the overlying Gattendorfia limestone. The Famennian clymeniid limestone, about 2–3 m thick, is characterised by pink and grey nodular biomicrite and is separated from the Gattendorfia limestone by a thin layer of dark-grey shales. In the opinion of Haydukiewicz (1990) and Muszer and Haydukiewicz (2011), this thin (1–5 cm-thick) black shale horizon corresponds to the condensed uppermost Famennian and the lowermost Tournaisian, and is treated as being the equivalent of the Hangenberg shales horizon. Also Mistiaen and Weyer (1999) suggested that the Wocklumeria sphaeroides subzone is missing at least and this dark shale horizon is an equivalent of HBS, however without any biostratigraphic evidence. The Wapnica “formation” is overlain by the fragmentarily preserved Gołogłowy “formation” comprising black shales as well (lower Carboniferous; for details, see Wajsprych 1995). The Gołogłowy “formation” is discordantly overlain by sandstones of the Nowa Wieś “formation”, which probably belongs to the lowermost part of the Viséan (Głuszek and Tomaś 1993; Muszer and Haydukiewicz 2011).
The Famennian to Tournaisian interval in the Wapnica Quarry has been subject to intense and palaeontological studies. Cephalopods were studied by Schindewolf (1937), Lewowicki (1959), Weyer (1965), Korn (1993), Korn et al. (2005) and Dzik (1997, 2006). Unique deep-water corals were analysed by Berkowski (2001, 2002) and Berkowski and Bełka (2008).
At first, Schindewolf (1937) assigned the clymeniid limestone to the lower part of the Wocklumeria Stufe. Lewowicki (1959), after Schindewolf (1937), ascertained the lack of the upper part of the Wocklumeria Zone. The re-examination of two sections within the quarry by Korn et al. (2005) confirmed Schindewolf’s estimation. A section located in the mid-northern part of the quarry yielded representatives of the Famennian Muessenbergia sublaevis, Muessenbergia parundulata and Effenbergia lens zones, and a section at the northern end of the quarry, described earlier by Lewowicki (1959), in the topmost bed of the clymeniid limestone yielded Kamptoclymenia endogona Schindewolf, a species that characterises the Kamptoclymenia endogona Zone (Korn et al. 2005, fig. 2; Becker et al. 2016, fig. 4). The lack of characteristic genera of the uppermost zone of the Wocklumeria Stage, Lewowicki (1959) connected with the existence of hiatus. On the other hand, Effenbergia and Balvia were collected by Dzik (2006) at the southern end of the Wapnica Quarry among loose blocks, close to the vertical rock wall. Dark-grey limestone with numerous bivalves (sample Dz-75), also occurring in looser blocks, yield the uppermost Famennian Protognathodus-fauna, otherwise unknown from the wall of the quarry (Dzik 2006, fig. 116 M–N). The uppermost part of the carbonate succession in the Wapnica Quarry is represented by the Gattendorfia limestone (about 1.5 m thick) that belongs to the Tournaisian Gattendorfia crassa Zone (Weyer 1965).
Agglutinated foraminifers from the clymeniid limestone were studied by Woroncowa-Marcinowska (2017). This group, rather facies-sensitive, does not appear to provide a basis for detailed biostratigraphy. The foraminiferal assemblage (comp. Woroncowa-Marcinowska 2017, fig. 6 and plates 3 and 4) includes forms with free tests such as Hyperammina, Reophax, Paratikhinella, Lagenammina and Septatournayella, as well as sessile forms, Tolypammina and Moravammina. The association resembles the Hyperammina biofacies of Herbig (2006), dominated by species of Hyperammina; representatives of Psammosphaera and Psedoastrorhiza, as well as Thurammina, are absent. The Hyperammina biofacies is located within the deeper parts of the outer shelf environment (compare Balthasar and Amler 2003).
Conodonts in the Wapnica Quarry were studied by Freyer (1968), Chorowska (1979), Chorowska and Radlicz (1987), Haydukiewicz (1990), Dzik (1997, 2006) and Belka (in Dopieralska et al. 2006). According to Belka and Haydukiewicz (in Streel et al. 2004) and Belka (in Dopieralska et al. 2006), the clymeniid limestone contains e.g. Bispathodus ultimus, Palmatolepis gracilis gonioclymeniae and Pseudopolygnathus marburgensis trigonicus, conodonts representative of the Upper expansa and Lower praesulcata zones, and the first Siphonodella (Eosiphonodella) praesulcata occurs about 1 m above the base of the clymeniid limestone. Unfortunately, Siphonodella praesulcata is not illustrated; therefore, this information should be verified in the future, since this species has been used by different authors in a variable sense (see discussion in Kaiser and Corradini 2011).
Wapnica quarry near Dzikowiec – new results
Conodont biostratigraphy
As a contribution to the ongoing international revision of the DCB, 15 new samples were collected near the northern corner of the Wapnica Quarry (Fig. 12b–d) for conodonts and palynomorph analysis to make the previous biostratigraphic results more precise in relation to new recommendation of the DCB Working Group. Most of the samples were taken from the main section (see Fig. 12b, c), from the clymeniid limestone and Gattendorfia limestone. Most of the Famennian samples yielded a diverse and relatively numerous conodont fauna, very similar in its taxonomic composition to conodont faunas known from the Kowala section (comp. Fig. 4). Beds 1 to 7 contain (Figs. 13, 14, and 15a–m) Pseudopolygnathus marburgenis trigonicus and different bispathodids, represented by Branmehla inornata, Branmehla suprema, Mehlina strigose, Bispathodus costatus, Bispathodus ultimus, Bispathodus bispathodus, Bispathodus aculeatus aculeatus and Bispathodus aculeatus anteposicornis, as well as palmatolepids with Palmatolepis gracilis gonioclymenia, Palmatolepis gracilis gracilis, Palmatolepis gracilis sigmoidalis and Palmatolepis gracilis expansa. Polygnathus inornatus, Neopolygnathus communis communis and Neopolygnathus vogesi are long-ranging taxa. Only Pseudopolygnathus marburgensis trigonicus, Palmatolepis gracilis gonioclymenia and Bispathodus ultimus are relatively short-ranging taxa, which characterise the Upper expansa and Lower praesulcata zones in ‘standard’ zonations of Ziegler and Sandberg (1984) or the Bispathodus ultimus Zone sensu Kaiser et al. (2009) and Becker et al. (2016). There is no conodont fauna in four samples (Dz8–Dz11) from the black shale and limestone interval (HBS).

The DCB interval, Wapnica Quarry near Dzikowiec (Sudetes)—lithological log with position of conodont samples, biostratigraphy and conodont distribution

Conodonts of the ultimus–praesulcata zones, Wapnica Quarry near Dzikowiec (Sudetes). a Palmatolepis gracilis gracilis Branson and Mehl, 1934a, sample 1, oral view. b Pseudopolygnathus marburgensis trigonicus Ziegler, 1962a (juvenile form), sample 1, oral view. c Bispathodus costatus (Branson, 1934) morphotype 2, sample 1, oral view. d Branmehla inornata (Branson and Mehl, 1934a), sample 1, lateral view. e Mehlina strigosa (Branson and Mehl, 1934a), sample 1, lateral view. f Palmatolepis gracilis expansa Sandberg and Ziegler, 1979, sample 2, oral view. g Palmatolepis gracilis sigmoidalis Ziegler, 1962a, sample 2, oral view. h Pseudopolygnathus marburgensis trigonicus Ziegler, 1962a (juvenile form), sample 2, oral view. i Branmehla suprema (Ziegler, 1962a), sample 2, oral view. j Branmehla inornata (Branson and Mehl, 1934a), sample 2, lateral view. k Palmatolepis gracilis sigmoidalis Ziegler, 1962a, sample 6, oral view. l Neopolygnathus vogesi (Ziegler, 1962a), sample 6, oral view. m Pseudopolygnathus marburgensis trigonicus Ziegler, 1962a, sample 6, oral view. n and o Bispathodus ultimus ultimus (Bischoff, 1957), sample 6, oral views

Conodonts of the ultimus–praesulcata zones (a–m) and ?bransoni and/or ?duplicata zones (n–t), the Wapnica Quarry near Dzikowiec. a–c Palmatolepis gracilis sigmoidalis Ziegler, 1962a, sample 7, oral view. d Palmatolepis gracilis expansa Sandberg and Ziegler, 1979, sample 7, oral view. e Palmatolepis gracilis gonioclymeniae Müller, 1956, sample 7, oral view. f Neopolygnathus vogesi (Ziegler, 1962a), sample 7, oral view. g Pseudopolygnathus marburgensis trigonicus Ziegler, 1962a, sample 7, oral view. h, m Bispathodus ultimus ultimus (Bischoff, 1957), sample 7, oral views. i Branmehla suprema (Ziegler, 1962a), sample 7, oral view. j, l Bispathodus costatus (Branson, 1934) morphotype 1, sample 7, oral views. k Bispathodus aculeatus anteposicornis (Scott, 1961), sample 7, oral view. n Siphonodella belkai Dzik, 1997, sample 14, oral view. o, p Siphonodella. duplicata (Branson and Mehl, 1934), sample 14, oral views. r, s Polygnathus purus purus (Voges, 1959), sample 14, oral views. t Pseudopolygnathus? primus Branson and Mehl, 1934, sample 14, oral view
There is only a sparse conodont fauna in four samples (Dz12–Dz15) from the thick-bedded nodular Gattendorfia limestone above the HBS horizon. Only sample Dz14 yielded Siphonodella duplicata, Siphonodella belkai (see revision in Kaiser et al. 2017) and Pseudopolygnathus primus (Figs. 13 and 15n–t); therefore, some Tournaisian conodont data of Chorowska (1979) and Dzik (1997) were incorporated in the biostratigraphic analysis (see Fig. 13). Chorowska (1979) noted the presence of Polygnathus purus purus, Siphonodella sulcata, Protognathodus kockeli and specimens described at that time as Siphonodella duplicata in a position close to our sample Dz12 (see Fig. 13). Unfortunately, these important conodont taxa were not illustrated by Chorowska (1979); therefore, this information cannot be verified. These conodont taxa indicate a stratigraphic position not earlier than the duplicata Zone. However, taking into account the revision of Siphonodella duplicata (early morphotype M1 of S. duplicata = Siphonodella bransoni Ji, late morphotype M2 of Siphonodella duplicata = classical Siphonodella duplicata), as well as the revision of the Tournaisian conodont scheme, some biostratigraphic uncertainties concerning Chorowska’s determinations still remain. Branmehla inornata documented by Chorowska higher up in the section, in a position close to our sample Dz15 (see Fig. 16), supports rather a position within the Tournaisian bransoni Zone for this part of the section (samples Dz12 to Dz15). A very numerous collection of conodonts was recovered by Dzik (1997, comp. table 1) from the Wapnica Quarry. Unfortunately, with some exceptions, it was very difficult to indicate even an estimated position of the conodont fauna presented by Dzik in relation to our section.

Distribution of microflora in the Wapnica Quarry section near Dzikowiec
Microflora from the DCB interval
Palynomorphs were analysed from the Wapnica Quarry section for the first time. Unfortunately, a large number of the samples were barren (see Fig. 16); only three of them possessed an organic content. Positive Devonian samples were obtained from the approx. 20-cm-thick interval of black shale with carbonate nodules (samples Dz9–Dz11; see Fig. 16). Two positive Carboniferous samples (20 and 30A) were additionally taken from two different walls of the quarry (asterisks in Fig. 12b).
Important miospores and phytoplankton taxa are presented in Figs. 17, 18, and 19. The palynomorph assemblage from the uppermost Famennian is high diversity with prevalent miospores; amorphic organic matter (AOM) was noticed as well. The microflora is moderately preserved and slightly overheated. Conodonts show a colour alteration index (CAI) value of 2.5, indicating that the rocks have not been exposed to temperatures higher than about 100 °C (Dopieralska et al. 2006).

The uppermost Famennian phytoplankton assemblage from the Wapnica Quarry near Dzikowiec. a Micrhystridium stellatum Deflandre, 1945; sample 9. b Micrhystridium stellatum Deflandre, 1945; sample 10. c Dictyotidium sp., sample 10. d Dictyotidium sp., sample 9. e Cymatiosphaera sp., sample 11. f Cymatiosphaera chelina Wicander and Loeblich, 1977; sample 10. g Gorgonisphaeridium winslowiae Staplin, Jonsonius and Pocock, 1965; sample 11. h Lophosphaeridium segregum Playford in Playford and Dring, 1981; sample 9. i Leisphaeridia sp.; sample 9

Abnormal ornamentation of the Famennian Retispora lepidophyta and lowermost Tournaisian miospore assemblage from the Wapnica Quarry near Dzikowiec. a–e Retispora lepidophyta (Kedo) Playford, 1976; morphological variation of reticulum, a and d sample 9; b, c and e sample 10. f Plicatispora scolecophora (Neves and Ioannides) Higgs et al., 1988; sample 30A. g unidentified; sample 30A. h Tumulispora malevkensis Sullivan, 1968; sample 30A. i–k Densosporites sp., sample 30A. l Murospora sp.; sample 20. m Discernisporites micromanifestus (Hacquebard) Sabry and Neves, 1971; sample 20. n Densosporites sp., sample 30A. o unidentified; sample 20. p Convolutispora major (Kedo) Turnau, 1978; sample 20

The uppermost Famennian miospore assemblage from the Wapnica Quarry near Dzikowiec. a Auroraspora macra Sullivan, 1968; sample 10. b Tumulispora malevkensis (Kedo) Turnau, 1978; sample 9. c Retusotriletes incohatus Sullivan, 1964; sample 10. d Auroraspora macra Sullivan, 1968; sample 10. e Vallatisporites vallatus Hacquebard, 1957; sample 9. f Retispora lepidophyta (Kedo) Playford, 1976 var. minor; sample 9. g Abnormal miospore similar to Vallatisporites sensu Filipiak and Racki (2010), sample 10. h Verrusosisporites(?) sp.; sample 9. i Raistrickia sp.; sample 9. j Neoraistrickia sp.; sample 9. k Bascaudaspora submarginata (Playford) Higgs et al., 1988; sample 9. l Vallatisporites verrucosus Hacquebard, 1957; sample 11. m Grandispora famenensis (Naumova) Streel, 1974 var. famenensis; sample 9. n Grandispora gracilis (Kedo) Streel in Becker et al., 1974; sample 9. o Grandispora femenensis (Naumova) Streel, 1974 var. famenensis; sample 10. p Vallatisporites pusillites (Kedo) Dolby and Neves, 1970; sample 9. q Diducites poljessicus (Kedo) Van Veen, 1981; sample 10. r Cordylosporites sp.; sample 9. s Grandispora micronulata (Kedo) Avkhimovitch in Higgs et al., 2000; sample 9. t Knoxisporites literatus (Waltz) Playford, 1963; sample 10. u Tumulispora(?) sp.; sample 11. v Vallatisporites pusillites (Kedo) Dolby and Neves, 1970; sample 9. w Spinozonotriletes sp., sample 11
Based on the presence of Retispora lepidophyta var. minor, together with Vallatisporites verrucosus, the LN miospore Zone was tentatively recognised (Fig. 16). The second index taxon Verrucosisporites nitidus is absent in all samples studied. The presence of Vallatisporites verrucosus was taken into account in determining the precise palynological zone (Fig. 16). The first appearance of V. verrucosus is considered to coincide with that of Verrucosisporites nitidus (see Higgs et al. 1988). However, when not using Vallatisporites verrucosus as an important LN marker, rather the LE (R. lepidophyta–Indotriradites explanatus) level should be distinguished here (see Prestianni et al. 2016). Generally, Retusotriletes and Apiculiretusispora predominate in the assemblage. More frequently appear Auroraspora asparella, A. macra, Bascaudaspora submarginata, Cyrtospora cristifera, Grandispora spp., Tumulispora spp. and Vallatisporites spp. Less frequent are Cymbosporites acutus, Grandispora famenensis, Indotriradites explanatus, Knoxisporites literatus, K. triradiatus, Plicatispora scolecophora, Raistrickia spp. and Spinozonotriletes uncatus. Vallatisporites vallatus (Fig. 19e) was noticed in samples 9 and 10 from the upper Famennian as well.
The palynostratigraphic analysis confirmed that the thin complex of black shale with carbonate nodules in the Wapnica section (see Fig. 16) should be treated as a HBS equivalent.
The second miospore assemblage was identified in two samples from the Gattendorfia limestone (Fig. 16; Dz30A and Dz20). The samples were taken from two different walls and lithologically correlated with the main section (Fig. 16). Significant is the lack of the taxa important for the uppermost Famennian, such as Retispora lepidophyta, Diducites spp. and Rugospora flexuosa among other miospores. This fact may tentatively point to the VI (Vallatisporites vallatus–Retusotriletes incohatus) miospore Zone. Only the latter index species was recognised, whereas no Vallatisporites spp. were noticed. The microfloristic assemblage is less diverse and consists of Convolutispora major, Corbulispora cancellata, Densosporites sp., Cymbosporites acutus, Cyrtospora cristifera, Plicatispora scolecophora, Pustulatisporites dolbii, Retusotriletes incohatus, Secarisporites sp. and Tumulispora spp. Lack of index taxa of the HD miospore Zone (Kraeuselisporites hibernicus and Umbonatisporites distinctus), the next Tournaisian miospore assemblage, also supports attributing this assemblage to the VI level. However, it should be stressed that miospores are poorly preserved and not very common; therefore, especially this identification should be verified in the future.
The Devonian phytoplankton assemblage from samples collected from the HBS comprises typical Upper Devonian acritarcha and prasinophyta (Fig. 17). Among them common were Cymatiosphara, Dictyotidium, Gorgonisphaeridium, Leiosphaeridia, Lophosphaeridium and Micrhystridium. Few remains of higher plants were noticed as well. Carboniferous samples, dated at the VI level, possess restricted in number taxa of Leiosphaeridia and Maranhites and large amounts of land plant debris (Fig. 16).
The frequency of Retispora lepidophyta in samples from the HBS (Dz9–Dz11) is very rare (< 1%). Moreover, R. lepidophyta is frequently represented by “abnormal” forms sensu Prestianni et al. (2016); see also Streel 1966) from the uppermost Famennian of Belgium. A characteristic feature is the interrupted reticulum with a different degree of muri disappearance (see Fig. 18). Reduction of ornamentation in more extreme forms is reduced to a few crests and verrucae. Moreover, some other abnormal (or mutated?) miospores have been observed in the analysed assemblage (Fig. 19g). This phenomenon corresponds well to the tener Event, noticed in the DCB sections in the eastern Dinant Synclinorium (Belgium; see Prestianni et al. 2016), as well as in the Holy Cross Mountains (Filipiak and Racki 2010; this paper) and in the Western Pomerania area (Matyja et al. 2015, fig. 7a).
Significant is the lack of Verrucosisporites nitidus in the HBS samples, analogous to Belgian profiles. Among others, Prestianni et al. (2016) consider ecological factors (e.g. sorting during transport) as being responsible for the lack of ‘heavier’ miospores (e.g. Tumulispora, Lophotriletes and Verrucosisporites). Meanwhile, this phenomenon is not fully observed in the Sudetes and the Holy Cross Mountains. Among the miospores observed, ‘heavier’ forms were noticed as well (e.g. Tumulispora, Corbulispora and Raistrickia). Additionally, the presence of mutant forms (abnormal microflora) points rather to some kind of biocrisis on continents.
The nature of this phenomenon is still unclear. In sections from the HCM, numerous tuffite layers occur close to the DCB interval in the Kowala section (Holy Cross Mountains). This helps to connect volcanic activity (Myrow et al. 2014; Racki et al. 2018a, 2018b; Percival et al. 2017) as the potential reason for flora mutation and occurrence of unseparated tetrads (for details, see Filipiak and Racki 2010) at least on a local scale. Last data on significant mercury enrichment corresponding to the Hangenberg Crisis in Vietnam suggests that the mercury is most likely sourced from distal volcanic emissions (Paschall et al. 2019). It is possible that large-scale volcanic activity acted as a trigger mechanism also for the Hangenberg Crisis.
According to Prestianni et al. (2016), on a global scale, the impoverished palynological assemblage from the DCB interval reflects rather climatic changes. A cooler and drier climate (see Streel and Marshall 2006) can be responsible for the rarefaction of coastal swamps and disappearance of Cyclostigma–Archaeopteris forests.
Western Pomerania—an overview of previous biostratigraphic results
The Western Pomerania region (NW Poland), buried under thick Permian and Mesozoic/Cenozoic sequences of the Polish Basin, during its Devonian and Carboniferous history was situated within the Trans European Suture Zone (TESZ) and located along the margin of the stable East European Craton to the north and northeast, and the Variscan-influenced areas to the south (Dadlez 2000; see Fig. 20a). The major palaeogeographic elements that affected the Devonian and early Carboniferous sedimentary evolution of the Pomeranian Basin were land areas representing uplifted parts of the East European Craton (Fennoscandian High extending to the north, outside Poland, and the Masurian–Belarus High, situated to the east). The sedimentary evolution and lithofacies pattern of the Pomeranian Basin during its Devonian and Mississippian history followed these main structural outlines (Matyja 2006, 2008, 2009).

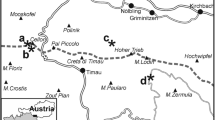
a Simplified sub-Pennsylvanian geological map of north-western Poland (Western Pomerania area) with location of studied borehole sections (modified after Matyja 2009, simplified). TESZ—Trans-European Suture Zone; b lithofacies pattern for the uppermost Famennian–lowermost Tournaisian in north-western Poland (modified after Matyja 2009); c some microfacies types across the DCB interval as examples of successions where the Hangenberg Black Shale horizon is not developed: Gorzyslaw-9 section—palaeoberesellid algae-crinoid packstone/grainstone, sample G 56, depth 3194.2 m, middle part of the LN miospore Zone; Chmielno-1 section—grey marl with relatively high amounts of silt and organic matter, sample Ch 52, depth 4010.6 m, uppermost part of the LN miospore Zone; compare Matyja et al. (2015); and Rzeczenica-1 section—crinoid wackestone/packstone, depth 2925.0 m, uppermost part of the LN miospore Zone
The uppermost part of the Famennian and lowermost part of the Tournaisian succession, widespread throughout the whole Pomerania area, belong to the Sąpolno Calcareous Shale Formation, which is a succession of carbonate and clayey deposits, and reflects an open-marine, middle/outer ramp phase of sedimentation (Matyja 1993, 2009; comp. Fig. 20b). Deposits studied in detail, in which a complete/or almost complete succession of the uppermost Famennian to lowermost Tournaisian has been recognised (Matyja and Stempień-Sałek 1994; Matyja et al. 2015), show a generally monotonous lithological pattern, and the characteristic HBS horizon is not developed here (Fig. 20c; see also Matyja et al. 2015). The uppermost Famennian part of the formation consists of dark marls and marly limestones, with crinoids, small palaeoberesellid green algae, calcareous and encrusting foraminifers, benthic ostracodes, rare brachiopods, laminar stromatoporoids, conodonts and miospores in the shallower part of the ramp (e.g. Gorzysław-9 section), and dark marls and claystones, with brachiopods, bivalves, gastropods, very rare ammonoids, trilobites and solitary corals, as well as conodonts and miospores in the relatively deeper part of the ramp (e.g. Rzeczenica-1 section). Brachiopods and bivalves are quite abundant in some parts of the succession (e.g. Korejwo 1975; Matyja 1976; Kłapciński and Muszer 1995a, 1995b, 1995c; Muszer 1998), but require a modern systematic study to assess their biostratigraphic potential. Cephalopods, corals and trilobites are too rare within this ramp facies to be of more than occasional application (e.g. Chwediuk 2003; Korejwo 1975).
There are no palmatolepids and Protognathodus fauna in the upper part of the Famennian in the Pomerania area. The last palmatolepids definitively retreat from the Pomeranian shelf at the end of the ‘standard’ marginifera or early in the trachytera Chron (‘standard’ conodont zonation of Ziegler and Sandberg (1984), owing to palaeoecological reasons (compare Matyja 1993). Therefore, the Famennian Upper expansa and Lower praesulcata ‘standard’ conodont zones of Ziegler and Sandberg (1984) (= ultimus Zone of Kaiser et al. 2009 and Becker et al. 2016) were recognisable owing to the presence of numerous species of Bispathodus (Matyja 1993). It seems that only the combined evidence from palynomorphs, conodonts and other microfossils provides more precise age determinations for this relatively shallow-water part of the DCB succession in relation to the pelagic succession (comp. Fig. 21), which was hitherto unfavourable with regard to individual fossil groups.

The uppermost Famennian–lowermost Tournaisian succession in Western Pomerania has been firstly recognised in the Gorzysław-9 and Rzeczenica-1 borehole sections (Matyja 1993; Matyja and Stempień-Sałek 1994). The uppermost dated part of the Famennian sequence in the Rzeczenica-1 section yielded conodont fauna indicative of the Upper expansa and/or Lower praesulcata zones (= ultimus Zone in new zonation of Kaiser et al. 2009 and Becker et al. 2016) due to the presence of numerous species of Bispathodus and Branmehla, i.e. Bispathodus costatus, Bispathodus aculeatus aculeatus, Bispathodus ultimus and Branmehla suprema (Fig. 22). In the Gorzysław-9 section, where conodonts are less abundant and poorly preserved, the uppermost part of the Famennian is characterised by Polygnathus delicatus, Polygnathus inornatus and Polygnathus streeli, as well as by Bispathodus jugosus and Bispathodus spinulicostatus (Fig. 23a, b). The presence of advanced siphonodellids, such as Siphonodella duplicata morphotype 1 (= Siphonodella bransoni), as well as Siphonodella quadruplicata (Matyja 1993; Matyja and Stempień-Sałek 1994; Matyja et al. 2000), suggests that the lowermost dated part of the Tournaisian succession in the Rzeczenica-1 borehole section (Matyja 1993, table 7) may be correlated with the upper part of the sandbergi Zone (sensu Sandberg et al. 1978; Clausen et al. 1989). Accompanying forms include single representatives of Polygnathus spicatus and Siphonodella belkai. Other conodont fauna consists almost entirely of long-ranging taxa (Matyja et al. 2000, fig. 3): Bispathodus spinulicostatus, Neopolygnathus communis morphotype 1, Polygnathus purus purus, Clydagnathus plumulus, Elictognathus bialatus and Elictognathus lacerates, Bispathodus stabilis morphotype 1 (= Bispathodus stabilis vulgaris Dzik) and Polygnathus inornatus.

Conodont, entomozoid and miospore distribution in the Rzeczenica-1 borehole section (Western Pomerania area); solid bars indicate certain ranges of species; empty bars indicate uncertain ranges of species, probably redeposited from older strata

a Distribution of conodonts, most important calcareous foraminifers (after Matyja and Tomaś, unpublished data) and miospores (after Stempień-Salek 2002 and Matyja and Stempień-Salek, unpublished data) in the uppermost Famennian–lowermost Tournaisian part of the Gorzyslaw-9 succession (Western Pomerania area); solid bars indicate certain species ranges; empty bars indicate uncertain species ranges, probably redeposited from older strata; b Conodonts of the ?ultimus –?praesulcata zones, Gorzyslaw-9 borehole section (Western Pomerania area): a, Polygnathus delicatulus Ulrich and Bassler, 1926, depth 3202.8 m, oral view. b Polygnathus inornatus Branson and Mehl, 1934b, depth 3198.7 m, oral view. c, siphonodelloid, depth 3198.7 m, oral view. d, Bispathodus spinulicostatus (Branson, 1934), depth 3202.8 m, oral view. e, Bispathodus ?jugosus (Branson and Mehl, 1934a), depth 3198.7 m, oral view. f, Polygnathus streeli Dreesen, Dusar and Groessens, 1976, depth 3202.8 m, oral view
A stratigraphic gap close to the DCB was suggested in these sections by conodont and miospore data, as the equivalents of the Middle and Upper praesulcata, sulcata and duplicata conodont zones (= costatus-kockeli Interregnum kockeli, sulcata-kuehni, bransoni and duplicata zones in the new zonation of Kaiser et al. (2009) and Becker et al. (2016)), as well as the western European lepidophyta–explanatus (LE), lepidophyta–nitidus (LN) and verrucosus-incohatus (VI) miospore zones were considered missing (comp. Turnau 1978, 1979; Matyja and Turnau 1989; Matyja and Stempień-Sałek 1994). The only physical manifestation of sedimentological disturbances within this interval was the occurrence of pyrite and organic matter. No surfaces with peculiar microrelief, which might evidence corrosion, have been found. Moreover, there was no evidence of any pre-sandbergi abrasion affecting the Famennian and Tournaisian deposits. It was suggested at that time that this stratigraphic gap resulted from some chemical (carbonate crisis) or hydrodynamic factors rather than from any tectonic uplift of the Pomeranian Basin floor (Matyja 1993). In some Pomeranian sections, however, the presence of the goniatite Pseudoarietites dorsoplanus dorsoplanus (Schmidt) (= Paprothites dorsoplanus (Schmidt)) was recognised (Korejwo 1993), which is the index taxon for the Paprothites dorsoplanus Zone (Korn 2006), correlated with the Upper duplicata conodont Zone (sensu Sandberg et al. 1978); this fact suggested that the gap could be smaller. Apart from the question on the range of the gap, it was clear, however, that the uppermost Famennian–lowermost Tournaisian deposits show reduced thicknesses in Pomerania, not exceeding several metres (compare Matyja et al. 2000, figures 18 and 19).
Later, the uppermost Famennian–lowermost Tournaisian interval, marked also by monotonous dark grey marls, marly claystones and claystones, was analysed in detail using biostratigraphy, sedimentology, magnetic susceptibility and geochemistry in the Chmielno-1 borehole section (Matyja et al. 2015). Almost a hundred samples were collected from the 20-m interval for conodont fauna; unfortunately, only one conodont sample was positive. Twenty-eight samples were palynologically productive and contained adequate palynological material for detailed biostratigraphic analysis. A complete sequence of the uppermost Famennian–lowermost Tournaisian, from the European standard lepidophyta–nitidus (LN) miospore Zone, through the vallatus–incohatus (VI) up to the local Pomeranian Convolutisopora major Zone (equivalent of the standard hibernicus–distinctus (HD) Zone), were recognised for the first time within the Pomeranian Basin (Matyja et al. 2015, fig. 5).
Western Pomerania—new results
Unfortunately, in the past, the Gorzysław-9 and Rzeczenica-1 sections were regularly sampled at 1-m intervals only. High-resolution biostratigraphic study in the Chmielno-1 sections, where ninety samples at ~ 20-cm intervals were collected (Matyja et al. 2015), allows for the recognition of the first complete sequence of the uppermost Famennian and lowermost Tournaisian recorded in the Pomerania area. The DCB interval in the Gorzysław-9 section was re-sampled for conodonts and miospores, and 76 new samples (G21–G97) at ~ 20-cm intervals were collected from the depth range of 3187.0–3203.0 m. Unfortunately, similarly as in the Chmielno-1 section (comp. Matyja et al. 2015), there were no positive conodont samples above the ultimus Zone. There was no possibility to re-sample the rock material from the DCB interval in the Rzeczenica-1 section, as it was not available any more. Therefore, the critical part of the DCB interval in both sections was re-examined for the palynostratigraphic analysis (Stempień-Sałek 2002; Matyja and Stempień Sałek unpublished data) to make the previous biostratigraphic results precise. It looks that the microflora characterises the Retispora lepidophyta–Verrucosisporites nitidus (LN) Zone between samples G73–G38, with both index species being represented. Other miospores noted within the LN Zone are represented by Diductites versabilis, Grandispora lupata, Tumulispora rarituberculata, Tumulispora malevkensis and others (Fig. 23a). The succeeding miospore assemblage belongs to the Vallatisporites verrucosus–Retusotriletes incohatus (VI) Zone. This assemblage is represented only by one index species, Retusotriletes incohatus, but does not possess other typical upper Famennian markers.
New results revealed the presence of LE, LN and VI standard western European miospore zones (comp. Figs. 22 and 23). Probably, there is no stratigraphic gap in these two sections, taking into account the miospore data; however, it looks that the lowermost Tournaisian deposits (VI miospore Zone) show extremely reduced thicknesses. Looking for the thicknesses of the LN miospore Zone in both sections, there is enough place above the unseparated ultimus–praesulcata conodont zones for the costatus–kockeli Interregnum and kockeli Zone (comp. Figs. 22 and 23). The absence of conodonts in this part of the DCB interval could be due to lithofacies (discussion in Kaiser et al. 2009).
Apart from palynomorphs and conodonts, entomozoacean ostracodes and calcareous foraminifers represent groups that are commonly used as biostratigraphic tools for the DCB interval. The great value of entomozoacean ostracodes with a fingerprint sculpture for European Devonian biostratigraphy remains undisputed (Gross-Uffenorde and Wang 1989; Gross-Uffenorde and Schindler 1990; Gross-Uffenorde et al. 2000). Entomozoaceans characterise pelagic environments; representatives of this group, however, sometimes invaded shelf facies. The Upper hemisphaerica–dichotoma Zone and lower hemisphaerica/latior Interregnum was defined in the Western Pomerania area (e.g. in the Rzeczenica-1 borehole section) by the presence of Richterina (Richterina) striatula (Żbikowska 1992), together with conodonts of the Upper expansa–Middle praesulcata zones, in terms of the new zonation—the ultimus–praesulcata zones (comp. Figs. 21 and 22). Famennian calcareous foraminifers occur sporadically in the Western Pomerania area, and the assemblages are generally of low diversity. Representatives of Quasiendothyra kobeitusana, Q. konensis, Q. regularis and Q. communis (Fig. 23a) have been found in the Gorzysław-9 borehole section (Matyja and Tomaś, unpubl. data). This assemblage, which is calibrated by the presence of conodont fauna, corresponds probably to the Quasiendothyra kobeitusana–Quasiendothyra konensis Zone (Kalvoda et al. 2015; comp. Figs. 21 and 23a).
Entomozoacean ostracodes, as well as calcareous foraminifers, are too rare in the Western Pomerania area to be of more than occasional use; however, representatives of these two groups confirm and emphasise previous biostratigraphic interpretations based on miospores or conodonts.
Geochemical and mineralogical characteristics of the DCB interval
Bulk rock and clay mineralogy, major element geochemistry and TOC content
The huge active Kowala Quarry is well established as a major reference site for new modern multidisciplinary studies (Marynowski and Filipiak 2007; Trela and Malec 2007; Marynowski et al. 2012; De Vleeschouwer et al. 2013; Myrow et al. 2014; Racki et al. 2018a, 2018b) of the Upper Devonian–Lower Carboniferous succession in the Holy Cross Mountains that reflect the dramatic environmental changes at the DCB. The intensely studied distinctive uppermost Famennian black bituminous horizons record several recurrent anoxic pulses that are known to be of global extent: annulata, Dasberg and Hangenberg (e.g. Marynowski and Filipiak 2007; Marynowski et al. 2010; Racka et al. 2010; Marynowski et al. 2012). Additionally, a new black shale horizon was recognised, located approximately 6.5 m below the HBS, referred to as the Kowala Black Shale (KBS; for details, see Marynowski and Filipiak 2007).
High values of U/Th, Ni/Co and V/(V + Ni) ratios, as well as high total organic carbon (TOC) contents and degree of pyritisation (DOP), point to anoxic/euxinic conditions particularly during the early deposition of the HBS (Marynowski et al. 2012). However, palaeoredox proxies suggest also the presence of transitional oxic episodes in the middle part of the HBS. Increased TOC contents, total sulphur (TS) and decreased CaCO3 concentrations clearly indicate excessive burial of organic matter (Marynowski et al. 2012; De Vleeschouwer et al. 2013). Moreover, the co-occurrence of charcoal and high concentrations of polycyclic aromatic hydrocarbons (PAHs) was observed at the Hangenberg Event horizon evidencing wildfires (Marynowski and Filipiak 2007). Furthermore, the δ13C curve for the uppermost Famennian–lowermost Tournaisian interval in the Kowala succession reveals also disturbances of the biogeochemical cycle (Trela and Malec 2007). This curve was recently supplemented by new data (see below).
Recently, the uppermost Famennian–lowermost Tournaisian interval in the Kowala section was also analysed by the present authors using inorganic geochemistry and mineralogy. A set of 11 samples was collected (Figs. 24 and 25) from units B and C (sensu Malec 2014) to analyse the bulk rock mineralogy, clay mineralogy (in < 0.002 mm fraction), major element geochemistry and TOC content.

Mineralogy of samples obtained from the Kowala Quarry section: a bulk rock data, b clay mineral data (< 0.002 mm fraction)

Carbon isotope record, magnetic susceptibility (MS) changes; solid line—smoothed data (5 samples window) and weight percentage of total organic carbon (TOC) and sulphur trioxide (SO3) across the DCB interval in the Kowala section (Holy Cross Mountains)
Major bulk-rock mineralogical changes in Kowala (Table 3, Fig. 24a) reflect the main lithological diversity. Shale (claystone) layers consist predominantly of phyllosilicates (35–45%) and detrital quartz (19–39%), and reveal a high detrital index (DI) in the range of 2.8–8.8. Calcite is observed here only in subsidiary amounts (8–22%). Moreover, feldspars are detected in minor quantities (0–6%). In the black shale, pyrite and dolomite appear in the range of 0–4% and 0–3%, respectively (Table 3, Fig. 24a). The marly limestones are mainly composed of calcite (with a proportion ranging from 63 to 86%), accompanied by subsidiary quantities of phyllosilicates (5–20%) and detrital quartz (5–12%) (Table 3, Fig. 24a). Other minerals are absent or they were detected in accessory amounts.
The results of bulk-rock mineralogy are confirmed by chemical data. The shale samples (6a, 23a, 23b, 23 and 29) are characterised by a high content of Al2O3, SiO2, TiO2 and K2O and a minor content of CaCO3 (Table 4). Reverse results were obtained for the limestone samples. Importantly, all black shale samples (6a, 23a, 23b, 23) contain very high amounts of organic carbon (TOC in the range of 10.1–16.6%). In other samples (also in grey shale), the TOC data does not exceed 0.2% (Table 3, Fig. 25). Interestingly, in some black shale samples (6a, 23a, 23b), much higher contents of sulphur (SO3) were also detected (Table 4, Fig. 25).
Typically, the clay fraction of the recently examined samples from Kowala is mostly dominated by illite and chlorite or vermiculite (Fig. 24b). The clay minerals are frequently accompanied by illite–smectite mixed layers and/or chlorite–vermiculite mixed layers. However, in sample 23a (from the lower part of the HBS), smectite and kaolinite predominate (Fig. 24b). Generally, rocks of the Kowala succession are thermally immature to early mature, with Rock-Eval Tmax values between 421 and 425 °C (Marynowski and Filipiak 2007); thus, the diagenetic overprint is insignificant here. A chlorite to vermiculite transformation occurs quite frequently in continental strata and usually documents moderate chemical weathering in acidic conditions (Thorez 1985; Velde and Meunier 2008). Successively, the weathering products may be eroded and supplied to the sedimentary basin. However, according to Marynowski et al. (2017), vermiculitisation of chlorite in at least the Tournaisian part of the Kowala succession is related to epigenetic (late Permian?) weathering. The presence of smectitic minerals in the Kowala section confirms a nearby volcanic activity at that time. Kaolinite typically dominates in mature soils that develop as a result of intense hydrolysis. The relatively high kaolinite content (up to 25% of the clay fraction) may evidence warm and humid climatic conditions in the adjacent continent at the beginning of the Hangenberg Event. In the case of Kowala, however, it may also be a result of fast volcanic ash alternation and kaolinisation of volcanic glass in an acidic environment. Nevertheless, CIW′ values (Cullers 2000) in the shale samples are very high (ranging from 96.3 to 98.4), suggesting a warm and humid climate and intense chemical weathering (Table 4). The highest CIW′ value is calculated for the “kaolinitic” sample 23a. CIW′ (named also CPA; see Buggle et al. 2011) is the Ca-free version of the classical chemical index of weathering—CIW (Harnois 1988). Increased chemical weathering is also confirmed by other indices (not illustrated herein).
The origin of major environmental changes across the DCB and the reasons of the Hangenberg Crisis are still debated. Elevated external nutrient input and fluxes of terrestrial runoff (Algeo and Scheckler 1998; Algeo et al. 2001; Rimmer et al. 2004; Carmichael et al. 2016), coastal and equatorial upwelling (Perkins et al. 2008; Caplan and Bustin 2001) or climate/salinity-driven upwelling that inverted oceanic stratification (Kaiser et al. 2015) were proposed as causes of tropical eutrophication and black shale formation during the Hangenberg Event. Presumably, the factors that triggered enhanced organic carbon burial and dysoxic–anoxic conditions were diverse and complex. Different times of the end-Devonian extinctions of different taxonomic groups may also suggest more than one lethal factor (Carmichael et al. 2016), although the main extinction episode occurred at the base of the shale interval (Kaiser et al. 2015). Nevertheless, most authors (e.g. Caplan and Bustin 1999; Rimmer 2004; Marynowski and Filipiak 2007; Kaiser et al. 2006, 2008; Marynowski et al. 2012) concur that the deposition of the HBS occurred in an anoxic or significantly oxygen-depleted environment. The results of former and recent geochemical and mineralogical research in the Kowala succession seem to confirm the current view that climatic and eustatic changes were the main cause of dramatic changes across the DCB (Kaiser et al. 2008 and references therein). More recent work indicates short-term climatic fluctuations in the DCB interval (Kaiser et al. 2011, 2015; Carmichael et al. 2016). De Vleeschouwer et al. (2013) established (using cyclostratigraphic methods) that the late Famennian major environmental changes and the Hangenberg Crisis were linked with climate shifts that may be predicted by orbital forcing. Moreover, widespread trap magmatism with injections of carbon dioxide and sulphur dioxide into the atmosphere was also suggested as a trigger of the Late Devonian climatic and environmental crisis (Marynowski et al. 2012 and references therein; Bond and Wignall 2014). Indeed, tuffites and other traces of volcanic activity were recognised in the Kowala succession (Marynowski and Filipiak 2007; Marynowski et al. 2012). A suggested greenhouse episode, which may have preceded the transgressive pulse and HBS deposition (Kaiser et al. 2006; De Vleeschouwer et al. 2013), could have initiated a carbonate crisis and profound perturbations in the carbon cycle. Warm and humid climate promoted strong chemical weathering of the continents, which is suggested in Kowala by the deficiency of limestones, domination of shale (with high values of weathering indices) and significant amounts of kaolinite. Successively, intense weathering of silicate rocks associated with enhanced burial of organic matter, and the development of the first leafy forests, prominently reduced the carbon dioxide level in the atmosphere. This sequestration of greenhouse gasses would have caused global cooling and resulted in the end-Devonian high-latitude glaciation and subsequent sea-level fall (Sandberg et al. 2002; Isaacson et al. 2008; Algeo et al. 2008; Kaiser et al. 2011, 2015).
The uppermost Famennian–lowermost Tournaisian interval was also analysed using organic, inorganic and stable isotope geochemistry in the Chmielno-1 reference section of the relatively shallow carbonate ramp environment within the Pomeranian Basin (Matyja et al. 2015). The DCB transition interval in the Chmielno-1 section is marked by monotonous dark grey marls, marly claystones and claystones; however, the HBS horizon in the latest Famennian (LN miospore Zone) is not developed here. Nevertheless, perturbations of the carbon cycle are reflected in positive δ13C excursions (see below). Geochemical features of this section indicate that the bottom water redox conditions were generally oxic during the sedimentation of the uppermost Devonian and lowermost Tournaisian deposits (Matyja et al. 2015). Total organic carbon (TOC) and total sulphur (TS) contents are much lower than in the Kowala section (cf. Marynowski et al. 2012). Moreover, the trace metal ratios (U/Th, V/Cr and most of Ni/Co data) and pyrite characteristics suggest domination of oxic conditions in the Pomeranian Basin. Nevertheless, short intermittent pulses of anoxia/euxinia have also been recorded by biomarker data across the DCB. Most probably, the recurrence of H2S-rich waters, where euxinia was present in the photic zone of the water column, was the reason for the almost complete absence of conodonts and other fauna at that time (Matyja et al. 2015). Moreover, elevated PAH concentrations were detected in several samples within the LN miospore Zone. They are very similar to those measured for the HBS horizon in the Kowala succession (cf. Marynowski et al. 2012). This evidences widespread wildfires that occurred in the hinterland area in the latest Devonian as a result of carbon dioxide sequestration and O2 level increase.
Carbon isotope record
Carbon isotope studies have been intensively used to disentangle perturbations in the global carbon cycle. Moreover, carbon isotope data (and other stable isotope signatures) fixed in sedimentary inorganic and organic matter are among the most powerful proxies used in chemostratigraphy (Weissert et al. 2008). Perturbations in the global carbon cycle during the DCB interval were revealed through the carbon isotopic record in numerous sections of the world (Xu et al. 1986; Schönlaub et al. 1992, 1994; Brand et al. 2004; Buggisch and Joachimski 2006; Kaiser et al. 2006, 2008; Trela and Malec 2007; Cramer et al. 2008; Day et al. 2011; Kumpan et al. 2014a, 2014b; Matyja et al. 2015; Qie et al. 2015).
We present herein carbon isotope data, collected from the uppermost Famennian to lowermost Tournaisian marine successions in Poland. The current paper contains the carbon isotope curves of two outcrop sections (Kowala, Wapnica) and two borehole sections (Chmielno-1, Gorzysław-9) (Figs. 25, 26, 27, and 28). All profiles analysed show (at least to some extent) the same well-developed excursion towards heavy isotopic values across the DCB. Generally, rocks of the investigated successions are thermally immature to early mature (Grotek et al. 1998; Narkiewicz et al. 1998; Dopieralska et al. 2006; Marynowski et al. 2007); thus, the diagenetic overprint is rather insignificant here.

Carbon isotope record and magnetic susceptibility (MS) changes across the DCB interval in the Wapnica Quarry section near Dzikowiec (Sudetes)

Carbon isotope record and magnetic susceptibility (MS) changes across the DCB interval in the Gorzysław-9 borehole section (Western Pomerania)

Correlation of the DCB interval between the pelagic successions in Holy Cross Mountains and Sudetes (represented by the Kowala and Wapnica sections) and the ramp successions in the Western Pomerania region (represented by Chmielno-1 and Gorzysław-9 borehole sections), based on biostratigraphy (conodonts, miospores), carbon isotope records (δ13Ccarb), magnetic susceptibility (MS) and total organic carbon (TOC)
Most comprehensive are new carbon isotope values measured in the Kowala section in limestones and inorganic matter, combined with earlier data (Trela and Malec 2007). A prominent carbon isotope excursion in the DCB was depicted with a relatively high temporal resolution (Fig. 25). The δ13C values from the Kowala section show a stepwise positive shift with a magnitude of approximately 5‰ (V-PDB). First, a short-lived negative excursion of δ13Ccarb ~ 3.5‰ is marked at the top of the praesulcata Zone, just before the black shale deposits. Next, an increase in carbon isotope values from about − 2‰ to about + 1‰ δ13C may be observed, leading to the top of the costatus–kockeli Interregnum and continuing with the highest values of up to + 3‰ into the kockeli Zone. Finally, there is an abrupt return to much lower values (negative excursion of over − 5‰) at the top of the kockeli Zone (= ‘standard’ Upper praesulcata Zone). The abrupt decrease of values across the DCB are well known from several other regions (see Kaiser et al. 2006, 2008) and could be used as a correlation marker. Interestingly, the major positive anomaly is recurrently interrupted by short-lived shifts to lower values. Especially, near the base of the kockeli Zone, a brief but pronounced negative excursion (with an amplitude of approximately − 4‰) is recorded.
In the Wapnica section, a record of the Hangenberg Crisis (costatus - kockeli Interregnum) is very condensed (with a thickness of only about 0.2 m). The positive carbon isotope excursion of 1‰ δ13Ccarb is observed at the DCB, although a gap must be considered, which has influence on the isotopic record (Fig. 26). The subsequent minor positive excursion (after a minor negative shift) is related with the Tournaisian ?bransoni or ?duplicata conodont Zone.
In the Chmielno-1 borehole section (Pomerania area), a positive δ13C excursion is well visible; however, the sampling resolution is not as high as in the Kowala succession (Fig. 28). Nevertheless, the Chmielno-1 section has yielded the first complete sequence of uppermost Famennian and lowermost Tournaisian miospore zones recorded in Pomerania, although the HBS horizon is not developed here (Matyja et al. 2015). The major positive carbon isotope excursion (CIE) starts in the uppermost Famennian over 4 m below the LN miospore Zone with a value of − 1.7‰ δ13Ccarb and continues until the lowermost part of the Ma1 miospore Zone, where the maximum value of + 3.5‰ δ13Ccarb was noted (Matyja et al. 2015). Thus, the magnitude of the positive anomaly reaches approximately 5‰ δ13Ccarb. Subsequently, a prominent negative excursion of over 3‰ δ13Ccarb is noticed, with values of ca. 0 ‰ δ13Ccarb.
In the Gorzysław-9 borehole section (located also in the Pomerania area), only the final part of the positive δ13C anomaly at the DCB is probably visible (Fig. 27). Just above, a negative CIE of approximately 2.5‰ can be observed.
There are no real geochemical indices of the DCB interval, except of the carbonate isotope record and beginning of carbonate sedimentation. Carbon-isotope chemostratigraphy is useful in continuous limestone successions, with only minor shale interruptions. Kaiser et al. (2015) summarised the carbon isotope records around the DCB, and some of the excursions by Myrow et al. (2014) or Qie et al. (2015) are not necessarily of Hangenberg Crisis age. Moreover, precise correlation of curves are difficult or even impossible, as peak values vary significantly between different regions (e.g. Kumpan et al. 2014b; Myrow et al. 2014; Qie et al. 2015; Kaiser et al. 2015). Major positive CIEs may start close to the bottom of the costatus–kockeli Interregnum or slightly higher. Additionally, this positive anomaly may represent the c–kI alone or the costatus–kockeli Interregnum and kockeli together or the kockeli Zone alone in several regions. Therefore, a possible revised position of the DCB could be established between two distinct positive isotope excursions, which represent these conodont zones (see Kaiser et al. 2015). In many cases, the two distinct excursions cannot be recorded or were not recorded due to facies change, low sampling resolution or probable gaps. This has to be considered as well. This option for a redefined DCB level (base of the kockeli Zone) seems to be preferred for some palaeobiologic, sedimentologic and palaeoclimatic reasons. A negative episodic peak, which separates the positive anomalies of the δ13C curve, is clearly noticeable in the Kowala section and some other sections in Europe. However, this negative shift of carbon isotope values is often ambiguous or invisible as in the Chmielno-1 borehole and the Wapnica Quarry section because of regional diversity, stratigraphic gaps or less dense sampling. Therefore, the CIE at the Famennian/Tournaisian transition should be applied with caution in precise stratigraphic comparisons. Kaiser et al. (2015) argued that chemostratigraphy could be used, but it should be confirmed by biostratigraphy in general to avoid a circular argumentation. The DCB interval is well characterised by two positive CIEs, which are well supported by biostratigraphy: the first one is the c–kI and the second one is the kockeli Zone.
Rock magnetism
Magnetic susceptibility measurements were performed to find characteristic general trends in selected sections from Poland and to attempt their mutual correlation and also on regional scale with other European sections.
The average MS value for the Kowala section is 50.83 (*10−9 m3/kg), changing from 7.41 to 144.11 (*10−9 m3/kg). Slightly higher values were noted for the Gorzysław-9 borehole, where the mean value is 59.1 (*10−9 m3/kg), from min. 10 to max. 128.57 (*10−9 m3/kg). The lowermost values are noted in the Wapnica section near Dzikowiec, where the average MS is 30.97 (*10−9 m3/kg) and varies from 4.86 to 97.5 (*10−9 m3/kg). Results from the Kowala Quarry and Gorzysław-9 borehole are close to the MS marine standard of Ellwood et al. (2011) (55 *10−9 m3/kg), calculated for limestones, marls and shales, whereas in Wapnica, the values are comparable more to the Belgian fore-reef settings by Da Silva et al. (2009) (33 *10−9 m3/kg). In the Chmielno-1 borehole, the noted magnetic susceptibility values were the highest, with an average of 78.11 (*10−9 m3/kg), changing from min. 22.73 to max. 121.31 (*10−9 m3/kg), being significantly higher than the MS marine standard of Ellwood et al. (2011).
The Kowala Quarry section (Fig. 25) was sampled in detail, especially in the ultimus and praesulcata conodont zones. Low and stable MS values are characteristic for grey and greenish nodular limestones of the aculeatus Zone and lowermost ultimus Zone, and only a small positive peak marks the KBS interval. MS begins to increase higher up, and a substantial, very high signal variability is noted in the red and dark red nodular limestones of the ultimus Zone. The highest MS values are noted here and also the mean MS curve gradually increases. Slightly lower and more stable MS data, beginning with a rapid MS decrease at the lithological boundary with grey and greenish irregular nodular limestones, are noted in the uppermost ultimus and the lower praesulcata zones. A clear, stable MS low covers the HBS interval and is preceded by an MS high at the middle part of the praesulcata Zone. Beginning from the middle part of the costatus–kockeli Interregnum (upper claystone part of complex B), the MS values increase with some fluctuations and MS low at the middle part of the kockeli Zone. The upper part of the kockeli Zone is characterised by a minor increase of the MS value.
The Wapnica section was sporadically sampled in a 2.5-m interval, despite the fact that some interesting MS trends can be seen (Fig. 26). The lower, mostly red and dark red part of the section (ultimus–praesulcata zones) represents slightly higher MS values in thick- and thin-bedded nodular limestone, with a prominent MS peak in its middle part. Black shale and nodular limestone of the HBS horizon in the costatus–kockeli Interregnum is characterised by the lowest magnetic susceptibility, followed by a slight MS increase in the Tournaisian thick-bedded nodular limestones of the ?bransoni and/or ?duplicata conodont Zone.
In the Gorzysław-9 borehole (Fig. 27), the lowest interval, comprising mainly marly limestones, belongs to the Ra local miospore Zone (= part of the LE Zone in the western European standard miospore zonation; comp. Fig. 21) and represents relatively stable and low MS values. Considerable fluctuations may be observed in the marly and shaly lower LN miospore Zone, followed by an MS low in the middle, more calcareous part, and again an increase in the marly upper part of the LN Zone. In the bottom of the claystone and marly shale of the uppermost LN Zone, the MS low covers the potential HBS zone, which due to intense condensation across the DCB interval was not recognised in the section. Rapid increase in the MS values in the middle part of the VI miospore Zone begins with higher values with stable fluctuations of magnetic susceptibility in the upper segment of the section, associated with shale and marly claystone–dominated sedimentation.
In the Chmielno-1 borehole, a significant MS low with some positive peaks is noted in the uppermost Famennian (upper part of the LN miospore Zone) interval, which is treated as the HBS equivalent. It is followed by a characteristic MS high in the LN and VI boundary interval. The details are discussed by Matyja et al. (2015).
Conclusions
Biostratigraphic correlation between pelagic sections (e.g. the Kowala section in the HCM) and more shallow-water Pomeranian sections still remains problematic; however, some levels with correlation potential exist:
-
The undoubted last occurrence of typical pre-Hangenberg conodont taxa at the base of the HBS horizon in both pelagic sections or its equivalents in the ramp succession could define the base of the costatus–kockeli Interregnum (c–kI) sensu Kaiser et al. (2009) and Becker et al. (2016).
-
Conodont samples from the HBS horizon or from its equivalents contain no conodonts.
-
Within the overlying marly limestone unit in the Kowala section (unit C), probably belonging to the kockeli conodont Zone, the re-occurrence of long-ranging taxa (survivors), such as Neopolygnathus vogesi, Bispathodus stabilis and Neopolygnathus communis communis, together with the onset of some new taxa, i.e. ?Protognathodus sp. and some polygnathids such as Polygnathus purus purus in very small numbers, is observed.
-
There is no Protognathodus kockeli close to the DCB interval in our sections, although data of Dzik (1997, 2006) must be considered.
-
Siphonodella sulcata M5 and Polygnathus purus subplanus characterise the beginning of the Tournaisian sulcata/kuehni conodont Zone in pelagic Kowala sections; the lowermost dated part (up to now) of the Tournaisian succession in the Western Pomerania area is probably the duplicata conodont Zone.
-
The best miospores generally came from the HBS horizon or its equivalents (lepidiphyta–nitidus (LN) miospore Zone) and directly superjacent layers (verrucosus–incohatus (VI) miospore Zone) in all investigated sections. Two events, the tener Event within the uppermost Famennian LN Zone and concentration of phytoplankton taxa in the lowermost Tournaisian VI Zone, have a correlation potential with different areas (this paper, section “Microflora from the DCB interval—new results”).
-
Apart from palynomorphs and conodonts, entomozoacean ostracodes and calcareous foraminifers represent groups that are commonly used as biostratigraphic tools for the DCB interval. Unfortunately, entomozoacean ostracodes, as well as calcareous foraminifers, are too rare in the Western Pomerania area to be of more than occasional application; however, representatives of the two groups confirm and emphasise previous biostratigraphic interpretations based on miospores and conodonts.
-
It seems that combined evidence of palynomorphs and conodonts, coupled with other microfossils, provides more precise age determinations for the relatively shallow-water part of the DCB succession.
Most of the geochemical indices (TOC and TS content, biomarkers, U/Th, Ni/Co and V/(V + Ni) ratios) define only the HBS horizon or its equivalent. Mineralogical and major element data in the Kowala coincide mainly with lithological changes, which are expressed in the base of the HBS horizon and at the top of the grey shale layer (HS equivalent—Kaiser et al. 2006, 2008, 2015). The former boundary coincides with the base of the costatus–kockeli Interregnum (Middle praesulcata Zone). The latter surface corresponds to the top of this zone. A contact between the grey shale and the reappearing marly limestones (top of the c–k Interregnum) seems to be the most appropriate option for the regional correlation. However, distinct HBS and HS horizons are frequently lacking. For example, Pomeranian sections show a generally monotonous lithological pattern, and the characteristic HBS horizon, with some correlation potential, is not developed here. However, important microscale environmental perturbations, including fluctuations in the water column euxinia, wildfire evidence, relative sea-level changes and perturbations of the carbon cycle reflected by positive carbon excursions, were recognised in one of the studied-in-detail sections (Chmielno-1) across the DCB interval (Matyja et al. 2015). These perturbations display a pattern similar to that observed in many areas in Europe during the Hangenberg Event.
Recently, some clay mineral analyses were also performed. The presence of smectite and kaolinite in the lower part of the HBS in the Kowala section may indicate volcanic activity and warm-humid climatic conditions, respectively. So far, however, any distinct change in the clay mineral composition was not observed close to the stratigraphic boundaries.
The perturbation of the global carbon cycle near the DCB is commonly evidenced by broad positive δ13C excursions due to increased global burial rates of organic matter in the ocean (Buggisch and Joachimski 2006; Kaiser et al. 2006, 2008, 2015; Kumpan et al. 2014a). The carbon isotope curves from Polish sections display a pattern generally similar to that known from many sections in Europe. In Kowala (Fig. 25), the deposition of black shale and the main positive CIE are preceded by a distinct negative excursion, which records a warming episode beginning in the latest Early praesulcata Chron and a global transgression (Kaiser et al. 2006; De Vleeschouwer et al. 2013). Precise examination of the Kowala curve shows the potential presence of two positive excursions (more gradual in the c–k Interregnum and relatively sharp in the kockeli Zone). This is in accordance with some former studies from the Carnic Alps, Rhenish Massif and Moravian Karst (Kaiser et al. 2006, 2008, 2015; Kumpan et al. 2014a). The formerly recognised positive anomaly encompasses the black shale (HBS) and grey shale (HS) horizons. It records high rates of Corg burial, intense weathering and drawdown of atmospheric CO2, which contributed to global cooling, a glacial pulse in Gondwana and a major glacial–eustatic drop in sea level. The latter positive excursion comprises the limestone horizon (unit C of Malec 2014; comp. Figs. 3, 4and 25) considered to be an equivalent of the Stockum Limestone. It is preceded by a prominent negative shift in δ13C values close to the major sequence boundary (base of the kockeli Zone) and corresponds to the re-warming and another transgression immediately after the glacial event. A stepped nature of the positive CIE is clearly visible in the Kowala section (Fig. 25) and less distinctly in the Chmielno-1 borehole section (Matyja et al. 2015; Fig. 28. The stepwise increase in δ13C values in the latest Famennian is also well documented in several other marine successions in Europe and may provide a support for orbitally and climatically controlled burial of organic carbon (De Vleeschouwer et al. 2013). Minor negative δ13C shifts that punctuate the Famennian–Tournaisian positive anomaly may reflect brief fluxes of isotopically light carbon into the ocean and atmosphere during the warming episodes.
An attempt to correlate the sections above described is possible when similar trends and characteristic peaks are found on the geophysical curves. Some similarities can be seen despite differences in sample resolution and variable stratigraphic record causing problems with direct correlation of sections. This is mainly caused by possible condensation across the key interval in the Gorzysław-9 borehole and Wapnica Quarry sections. The intensively variable magnetic susceptibility signal in the ultimus–praesulcata zones, observed in detail in the pelagic Kowala section, reflects specific conditions in the Famennian basin that are additionally highlighted by changes in rock colour and nodular/normal limestone lithology. Even in the Wapnica Quarry section, increased MS values can be traced in the ultimus–praesulcata zones, with a positive peak close to the boundary between nodular breccia and thick-bedded limestone. Some similarities in the MS variability signal can be traced also in the Gorzysław-9 borehole, in the interval above the ?ultimus-?praesulcata zones, beginning at the boundary of the LN miospore Zones, and ranging up to the middle part of the LN Zone. In the Chmielno-1 borehole, such strong MS variations are not visible (Matyja et al. 2015, fig. 11; Fig. 28); however, the lowermost part of the section of the LN Zone, representing stable high MS values in marls and marly limestones, might be a time equivalent that does not record the changes, possibly because they did not significantly influence that part of the basin. The HBS interval in the Kowala Quarry or its equivalents close to the DCB in other sections is quite similar and characterised by a slight MS high just below the HBS, where the MS values go down, antecedent to an increase which is also observed in all four sections but at a different intensity (Fig. 28). Rapid increase in the MS value in the VI miospore Zone in the Gorzysław-9 borehole may document an important change in the carbonate ramp environment. Perhaps, this interval may be correlated with the Chmielno-1 Event, taking place in the lowest Tournaisian, at the boundary between Ma0 and VI miospore zones, or in the lowest part of the Ma1 Zone (Matyja et al. 2015; comp. Fig. 28). It can be followed also in the pelagic Kowala Quarry section, starting from the upper part of the costatus–kockeli Interregnum, through the kockeli Zone. The chance for an unambiguous correlation is not obvious, since it is not clear exactly which MS peak in Kowala is to be correlated. A clear MS low is also noted in the middle part of the kockeli Zone. The trends mentioned above are not recognised in the Wapnica Quarry section due to condensation and gaps, and only a small-scale increase is observed in the ?bransoni and/or ?duplicata Zone.
The obtained magnetic susceptibility curves have also some correlation potential in other DCB sections. The Kowala Quarry record, described in detail in this paper, allows for its direct correlation with two sections for the Rhenish Slate Mountains (Oese and Oberrödinghausen), described by Kumpan et al. (2015). As noticed by them, a stable MS low in the uppermost Famennian of the Chmielno-1 section is correlatable with another Drewer section, located in the Northern Rhenish Massif.
References
Algeo, T. J., & Scheckler, S. E. (1998). Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events. Transactions of the Royal Society B: Biological Sciences, 353, 113–130.
Algeo, T. J., Scheckler, S. E., & Maynard, J. B. (2001). Effects of the Middle to Late Devonian spread of vascular land plants on weathering regimes, marine biotas, and global climate. In P. G. Gensel & D. Edwards (Eds.) Plants invade the land - evolutionary and environmental perspectives (pp. 213–236). New York: Columbia University Press.
Algeo, T.J., Heckel, P.H., Maynard, J.B., Blakey, R., & Rowe, H. (2008) Modern and ancient epicratonic seas and the superestuarine circulation model of marine anoxia. In C. Holmden & B.R. Pratt (Eds.) Dynamics of Epeiric Seas: sedimentological, paleontological and geochemical perspectives (pp. 7–38). Geological Association of Canada Special Publication, 48.
Amler, M. R. W. (2004). Late Famennian bivalve, gastropod and belerophontid mollusks from the Refrath 1 Borehole (Bergisch Gladbach-Paffrath Syncline; Ardennes-Rhenish Massif, Germany). Courier Forschungsinstitut Senckenberg, 251, 151–173.
Avkhimovitch, V. I. (1993). Zonation and spore complexes of Devonian and Carboniferous boundary deposits of the Pripyat depression (Byelorussia). Annales de la Société Géologique de Belgique, 115, 425–452.
Avkhimovitch, V. I., Turnau, E., & Clayton, G. (1993). Correlation of Uppermost Devonian and Lower Carboniferous miospore zonation in Belorussia, Poland and Western Europe. Annales de la Société Géologique de Belgique, 115, 453–458.
Balthasar, U., & Amler, M. R. W. (2003). Silicified foraminifera from the Lower Carboniferous Genicera Formation (Cantabrian Mountains, Northern Spain) and their palaeoenvironmental interpretation. Courier Forschungsinstitut Senckenberg, 242, 1–19.
Barnes, C. R., Zhang, Y., Jeppson, L., Fredholm, D., Varker, W. J., Swift, A., Merrill, G. K., & Dorning, K. J. (1987). Recent developments in rock disintegration techniques for the extraction of conodonts. In R. L. Austin (Ed.) Conodonts: investigative techniques and applications. The British Micropalaeontological Society Series (pp. 35–53). Chichester: Ellis Horwood.
Becker, R.T. (2010). The chance of evolutionary success – the case of the oldest (Tournaisian) ceratitoid ammonoids (Prodromitidae, Tornoceratina). 8th International Symposium ‘Cephalopods – Present and Past’, 30 August – 3 September 2010, Dijon, abstracts volume, 25-26
Becker, R. T., & Mapes, R. H. (2010). Uppermost Devonian ammonoids from Oklahoma and their palaeobiogeographic significance. Acta Geologica Polonica, 60, 139–163.
Becker, R.T., Hartenfels, S., Aboussalam, Z.S., Tragelehn, H., Brice, D. & El Hassani, A. (2013). The Devonian-Carboniferous boundary at Lalla Mimouna (Northern Maider) – a progress report. In R.T. Becker, A. El Hassani, & A. Tahiri (Eds.) International field symposium “the Devonian and Lower Carboniferous of Northern Gondwana”, Morocco, 2013. Documents de l’Institut Scientifique Rabat, 27, 109–120.
Becker, R.T., Kaiser, S.I. & Aretz, M. (2016). Review of chrono-, litho- and chronostratigraphy across the global Hangenberg Crisis and Devonian-Carboniferous Boundary. In R.T. Becker, P. Koenigshof, & C.E. Brett (Eds.) Devonian climate, sea level and evolutionary events. Geological Society, London, Special Publications, 423, 355–386
Berkowski, B. (2001). Astogeny of amural colonial Rugosa from the Famennian of the Sudetes – a palaeoenvironmental study. Acta Geologica Polonica, 51, 109–120.
Berkowski, B. (2002). Famennian rugosa and heterocorallia from southern Poland. Paleontologica Polonica, 61, 1–87.
Berkowski, B., & Bełka, Z. (2008). Seasonal growth bands in famennian rugose coral Scruttonia kunthi and their environmental significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 265, 87–92.
Berkowski, B., Zapalski, M., & Wrzołek, T. (2016). New Famennian colonial coral (Rugosa) from the Holy Cross Mountains (Poland): an example of local evolution after Frasnian-Famennian extinction. The Science of Nature, 103(3–4), 33.
Bogoslovsky, B. I. (1981). Devonian ammonoids. III. Clymeniids (Suboder Gonioclymeniina). Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, 191, 1–122. [in Russian]
Bond, D.P., & Wignall, P.B. (2014). Large igneous provinces and mass extinctions: an update. In G. Keller, & A.C. Kerr (Eds.) Volcanism, impacts, and mass extinctions: causes and effects. Geological Society of America Special Paper 505, 29–55.
Brand, U., Legrand-Blain, M., & Streel, M. (2004). Biochemostratigraphy of the Devonian–Carboniferous boundary global stratotype section and point, Griotte Formation, La Serre, Montagne Noire, France. Palaeogeography, Palaeoclimatology, Palaeoecology, 205, 337–357.
Buggisch, W., & Joachimski, M. M. (2006). Carbon isotope stratigraphy of the Devonian and central and southern Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 68–88.
Buggle, B., Glaser, B., Hambach, U., Gerasimenko, N., & Markovic, S. (2011). An evaluation of geochemical weathering indices in loess-paleosol studies. Quaternary International, 240, 12–21.
Caplan, M. L., & Bustin, M. R. (1999). Devonian-Carboniferous Hangenberg mass extinction event, widespread organic-rich mudrock and anoxia: causes and consequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 187–207.
Caplan, M. L., & Bustin, M. R. (2001). Palaeoenvironmental and palaeoceanographic controls on black, laminated mudrock deposition: example from Devonian-Carboniferous boundary, Alberta, Canada. Sedimentary Geology, 145, 45–72.
Carmichael, S. K., Waters, J. K., Batchelor, C. J., Coleman, D. M., Suttner, T. J., Kido, E., Moore, L. M., & Chadimová, L. (2016). Climate instability and tipping points in the Late Devonian: detection of the Hangenberg Event in an open oceanic island arc in the Central Asian Orogenic Belt. Gondwana Research, 32, 213–231.
Chibrikova, E.V., Kedo, G.I. & Streel, M. (1978). Important spores for the delineation of the Devonian-Carboniferous Boundary through USSR and Western Europe. In Compte Rendu 8ème Congrès international de Stratigraphie et de Géologie du Carbonifère, Moscou 1975, 1, 198-203. [In Russian]
Chorowska, M. (1979). Nowe wyniki badań stratygraficznych dewonu okolic Kłodzka i problem granicy dewon/karbon. Materiały Konferencji Terenowej Nowa Ruda (pp. 142-153). Wrocław: 8-9 września 1979 r. [in Polish]
Chorowska, M. & Radlicz, K. (1987). Wapienie górnego dewonu i dolnego karbonu w kamieniołomie na górze Wapnica w Dzikowcu. In Baranowski et al. (Ed.) Przewodnik LVIII Zjazdu PTG Wałbrzych (Field-guide of the LVIII Annual Meeting of the Geological Society of Poland), 188–190. Kraków: 17–19 września 1987 r. [in Polish]
Chwediuk, E. (2003). Late Devonian and early Carboniferous Rugosa from Western Pomerania, northern Poland. Acta Geologica Polonica, 55(4), 392–443.
Clausen, C.-D., Leuteritz, K., Ziegler, W., & Korn, D. (1989). Ausgewählte Profile an der Devon/Carbon – Grenze im Sauerland (Rheinisches Schiefergebirge). Fortschritte in der Geologie von Rheinland und Westfalen, 35, 161–226.
Clayton, G., & Turnau, E. (1990). Correlation of the Tournaisian miospore zonation of Poland and the British Isles. Annales Societatis Geologorum Poloniae, 60, 45–58.
Clayton, G., Coquel, G., Doubinger, J., Gueinn, K. J., Loboziak, S., Owens, B., et al. (1977). Carboniferous miospores of western Europe: illustration and zonation. Mededelingen Rijks Geologische Dienst, 29, 1–71.
Combaz A. & Streel M. (1971). Microfossiles végétaux du Tournaisien inférieur dans le core-drill de Brévillers (Pas-de-Calais, France). In M. Streel & R. Wagner (Eds.) Compte-rendus de la 8e réunion de la Commission internationale de Microflore du Paléozoïque et de l’Assemblée générale de l’IUGS, Liège, 1970. Subcommission on Carboniferous Stratigraphy. Les Congrès et Colloques de l’Université de Liège, 55, 227–240.
Corradini, C. (2008). Revision of the Famennian-Tournaisian (Late Devonian-Early Carboniferous) conodont biostratigraphy of Sardinia, Italy. Revue de Micropaleontologie, 51, 123–132.
Corradini, C., Spalletta, C., Mossoni, A., Matyja, H., & Over, J. (2017). Conodonts across the Devonian/Carboniferous boundary: a review and implication for the redefinition of the boundary and a proposal for an updated conodont zonation. Geological Magazine, 154(4), 888–902.
Cramer, B. D., Saltzman, M. R., Day, J. E., & Witzke, B. J. (2008). Record of the Late Devonian Hangenberg global positive carbon-isotope excursion in an epeiric sea setting: carbonate production, organic-carbon burial and paleoceanography during the late Famennian. In H. Holmden, & B. R. Pratt (Eds.) Dynamics of Epeiric Seas: sedimentological, paleontological and geochemical perspectives. Geological Association of Canada, Special Paper, 48, 103–118.
Cullers, R. L. (2000). The geochemistry of shales, siltstones and sandstones of Pennsylvanian-Permian age, Colorado, USA: implications for provenance and metamorphic studies. Lithos, 51, 181–203.
Czarnocki, J. (1928). Przegląd stratygrafii famenu i karbonu dolnego (kulmu) w zachodniej i środkowej części Gór Świętokrzyskich. Posiedzenia Naukowe Państwowego Instytutu Geologicznego, 21, 55–59- [in Polish]
Czarnocki, J. (1933). Stratygrafia warstw granicznych między dewonem i karbonem w okol. Kowali. Posiedzenia Naukowe Państwowego Instytutu Geologicznego, 35, 31–34. [in Polish]
Czarnocki, J. (1989). Klimenie Gór Świętokrzyskich. Prace Państwowego Instytutu Geologicznego, CXXVII, 1–91. [in Polish]
Da Silva, A.-C., Potma, K., Weissenberger, J. A. W., Whalen, M. T., Humblet, M., Mabille, C., & Boulvain, F. (2009). Magnetic susceptibility evolution and sedimentary environments on carbonate platform sediments and atolls, comparison of the Frasnian from Belgium and Alberta, Canada. Sedimentary Geology, 214, 3–18.
Dadlez, R. (2000). Pomeranian Caledonides (NW Poland), fifty years of controversies: a review and a new concept. Geological Quarterly, 44, 221–236.
Day, J., Witzke, B., & Rowe, H. (2011). Development of an epeirical subtropical paleoclimate record from western Euramerica: late Frasnian–earliest Tournaisian stable carbon isotope record from the Yellow Spring–New Albany Groups of the northwestern Illinois Basin. In Geological Society of America Annual Meeting, Abstracts with Programs, 43, 151.
De Vleeschouwer, D., Rakociński, M., Racki, G., Bond, D. P. G., Sobień, K., & Claeys, P. (2013). The astronomical rhythm of Late-Devonian climate change (Kowala section, Holy Cross Mountains, Poland). Earth and Planetary Science Letters, 356, 25–37.
Dopieralska, J., Belka, Z., & Haack, U. (2006). Geochemical decoupling of water masses in the Variscan oceanic system during Late Devonian times. Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 108–119.
Dzik, J. (1997). Emergence and succession of Carboniferous conodont and ammonoid communities in the Polish part of the Variscan Sea. Acta Palaeontologica Polonica, 42, 57–170.
Dzik, J. (2006). The Famennian "Golden Age" of conodonts and ammonoids in the Polish part of the Variscan Sea. Palaeontologia Polonica, 63, 1–359.
Eickhoff, G. (1970). Foraminiferen aus dem Wocklumer Kalk am Borke-Wehr bei Balve (Oberdevon, Rheinisches Schifergebirge). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 135, 227–267.
Ellwood, B. B., Algeo, T. J., El Hassani, A., Tomkin, J. H., & Rowe, H. D. (2011). Defining the timing and duration of the Kačák interval within the Eifelian/Givetian boundary GSSP, Mech Irdane, Morocco, using geochemical and magnetic susceptibility patterns. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 74–84.
Filipiak, P. (2004). Miospore stratigraphy of Upper Famennian and Lower Carboniferous deposits of the Holy Cross Mountains (Central Poland). Review of Palaeobotany and Palynology, 128, 291–322.
Filipiak, P. (2005). Late Devonian and Early Carboniferous acritarchs and prasinophytes from the Holy Cross Mountains (Central Poland). Review of Palaeobotany and Palynology, 134, 1–26.
Filipiak, P., & Racki, G. (2010). Proliferation of abnormal palynoflora during the end-Devonian biotic crisis. Geological Quarterly, 54(1), 1–14.
Foster, C. B., & Afonin, S. A. (2005). Abnormal pollen grains: an outcome of deteriorating atmospheric conditions around the Permian–Triassic boundary. Journal of the Geological Society (London), 162, 653–659.
Freyer, G. (1968). Conodontenfunde aus dem Oberdevon und Unterkarbon von Dzikowiec Kłodzki (Ebersdorf) und Gołogłowy (Hollenau) in Dolny Śląsk (Niederschleisien). Geologie, Berlin, 17(1), 60–67.
Głuchowski, E. (2002). Crinoids from the Famennian of the Holy Cross Mountains, Poland. Acta Palaeontologica Polonica, 47, 319–328.
Głuszek, A., & Tomaś, A. (1993). Age of the Nowa Wieś Formation (Bardzkie Mts, Middle Sudetes, SW Poland). Annales Societatis Geologorum Poloniae, 62, 293–308.
Gross-Uffenorde, H. & Schindler, E. (1990). The effect of the global events on entomozoacean ostracoda. In R. Whatley & C. Maybury (Eds.) Ostracoda and global events. British Micropalaeontological Society Series London, 11-112.
Gross-Uffenorde, H., & Wang, S. (1989). The entomozoacean succession of China and Germany (Ostracoda, Devonian). Courier Forschungsinstitut Senckenberg, 110, 61–79.
Gross-Uffenorde, H., Lethiers, F., & Blumenstengel, H. (2000). Ostracodes and Devonian stratigraphy. Courier Forschungsinstitut Senckenberg, 220, 99–111.
Grotek, I., Matyja, H., & Skompski, S. (1998). Dojrzałość termiczna materii organicznej w osadach karbonu obszaru radomsko-lubelskiego i pomorskiego. Prace Państwowego Instytutu Geologicznego, 65, 245–254. [in Polish]
Guerich, G. (1902). Zur Diskussion ueber das Profil von Ebersdorf. Zeitschrift der Deutschen Geologischen Gesellschaft, 54, 57–65.
Gutchick, R.C. & Sandberg, Ch.A. (1983). Mississippian continental margin of conterminous United States. In D. J. Stanley & G. T. Moore (Eds.) The shelfbreak: Critical interface of continental margins. Special publication - Society of Economic Paleontologists and Mineralogists 33, 79–96.
Halamski, A. T., & Baliński, A. (2009). Latest Famennian brachiopods from Kowala, Holy Cross Mountains, Poland. Acta Palaeontologica Polonica, 54, 289–306.
Harnois, L. (1988). CIW index: a new chemical index of weathering. Sedimentary Geology, 55, 319–322.
Hartenfels, S. (2011). Die globalen Annulata-Events und die Dasberg-Krise (Famennium, Oberdevon) in Europa und Nord-Afrika: hochaufloesende Conodonten-Stratigraphie, Karbonat-Mikrofazies, Palaeooekologie und Palaeodiversitaet. Münstersche Forschungen zur Geologie und Paläontologie, 105, 17–527.
Hartenfels, S. & Becker R.T. (2016). The global Annulata events: review and new data from the Rheris Basin (northern Tafilalt) of SE Morocco. In R. T. Becker, P. Koenigshof & C. E. Brett (Eds.) Devonian climate, sea level and evolutionary events. Geological Society, London, Special Publications, 423, 291–354.
Haydukiewicz, J. (1990). Stratigraphy of Paleozoic rocks of the Góry Bardzkie and some remarks on their sedimentation (Poland). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 179(2/3), 275–284.
Herbig, H.-G. (2006). Sandschalig-agglutnierte Foraminiferen. In Deutsche Stratigraphische Kommission (Eds.) Stratigraphie von Deutschland VI, Unterkarbon (Mississippium). Schriftenreihe der Deutschen Gesselschaft für Geowissenschaften (SDGG), 41, 243–249.
Higgs, K., & Streel, M. (1994). Palynological age for the lower part of the Hangenberg Shales in Sauerland, Germany. Annales de la Société Géologique de Belgique, 116, 243–247.
Higgs, K., Clayton, G., & Keegan, B. J. (1988). Stratigraphic and systematic palynology of the Tournaisian rocks of Ireland. Geol. Surv. Irel. Spec. Pap., 7, 1–93.
Isaacson, P. E., Diaz-Martinez, E., Grader, G. W., Kalvoda, J., Babek, O., & Devuyst, F. X. (2008). Late Devonian-earliest Mississippian glaciation in Gondwanaland and its biogeographic consequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 268, 126–142.
Ji, Q. (1985). Study on the phylogeny, taxonomy, zonation and biofacies of Siphonodella (Conodonta). Institute of Geology, Bulletin, 11, 51–75.
Kaiser, S. I., & Corradini, C. (2011). The early siphonodellids Conodonta, Late Devonian-Early Carboniferous: overview and taxonomic state. Neues Jahrbuch für Geologie und Paläontologie, 261, 19–35.
Kaiser, S. I., Steuber, T., Becker, R. T., & Joachimski, M. M. (2006). Geochemical evidence for major environmental change at the Devonian-Carboniferous boundary in the Carnic Alps and the Rhenish Massif. Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 146–160.
Kaiser, S. I., Steuber, T., & Becker, R. T. (2008). Environmental change during the Late Famennian and Early Tournaisian (Late Devonian-Early Carboniferous): implications from stable isotopes and conodont biofacies in southern Europe. Geological Journal, 43, 241–260.
Kaiser, S.I., Becker, R.T., Spaletta, C. & Steuber T. (2009). High resolution conodont stratigraphy, biofacies, and extinctions around the Hangenberg Event in pelagic successions from Austria, Italy, and France. In D.J. Over (Ed.) Studies in Devonian stratigraphy: Proceedings of the 2007 International Meeting of the Subcommission on Devonian stratigraphy and IGCP 499 (pp. 99-143). Palaeontographica Americana, 2009.
Kaiser, S. I., Becker, R. T., Steuber, T., & Aboussalam, S. Z. (2011). Climate-controlled mass extinctions, facies, and sea-level changes around the Devonian-Carboniferous boundary in the eastern Anti-Atlas (SE Morocco). Palaeogeography, Palaeoclimatology, Palaeoecology, 310, 340–364.
Kaiser, S. I., Aretz, M., & Becker, R. T. (2015). The global Hangenberg Crisis (Devonian-Carboniferous transition): review of a first-order mass extinction. In R. T. Becker, P. Koenigshof, & C. E. Brett (Eds.) Devonian climate, sea level and evolutionary events (Vol. 50, pp. 423–438). Geological Society, London, Special Publications. https://doi.org/10.1002/gj.2523.
Kaiser, S. I., Kumpan, T., & Cigler, V. (2017). New unornamented siphonodellids (Conodonta) of the lower Tournaisian from the Rhenish Massif and Moravian Karst (Germany and Czech Republic). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 286(1), 1–33.
Kaiser, S. I., Kumpan, T., & Rasser, M. W. (2019). High-resolution conodont biostratigraphy in two key sections from the Carnic Alps (Grüne Schneid) and Graz Paleozoic (Trolp) – implications for the biozonation concept at the Devonian-Carboniferous boundary. Newsletters on Stratigraphy. https://doi.org/10.1127/nos/2019/0520XX.
Kalvoda, J., Kumpan, T., & Bábek, O. (2015). Upper Famennian and Lower Tournaisian sections of the Moravian Karst (Moravo-Silesian Zone, Czech Republic): a proposed key area for correlation of the conodont and foraminiferal zonations. Geological Journal, 50, 17–38.
Kedo, G. I. (1957). Spores from the supra salt Devonian deposits of the Pripyat Depression and their stratigraphic significance. Akademia Nauk Belrussk SSR, Trudy Instituta Geologicheskikh Nauk, Seria Stratigrafia i Paleontologia, 2, 3–43. [in Russian]
Kedo, G. I. (1963). Spores of the Tournaisian Stage of the Pripyat Depression and their stratigraphical significance. Rep Palaeontologiskii I Stratigrafiskii Byelorussian SSR, 4, 3–121. [in Russian]
Kłapciński, J., & Muszer, J. (1995a). Utwory podpermskie z wybranych otworów wiertniczych Pomorza Zachodniego (synklinorium szczecińskie, synklinorium i antyklinorium pomorskie). Prace Geologiczno-Mineralogiczne, 48, 15–25. [in Polish]
Kłapciński, J., & Muszer, J. (1995b). Stratygrafia podłoża permu w otworze Kłanino 3 na podstawie makroskamieniałości (Pomorze Zachodnie). Prace Geologiczno-Mineralogiczne, 50, 191–198. [in Polish]
Kłapciński, J., & Muszer, J. (1995c). Utwory górnego dewonu i dolnego karbonu w otworach Brzozówka 2, Myśligoszcz 1, Rzeczenica 2 i Wałdowo Królewskie 1 (Pomorze Wschodnie). Prace Geologiczno-Mineralogiczne, 50, 209–217. [in Polish]
Korejwo, K. (1975). Utwory najniższego dinantu z profilu Babilon 1 (Pomorze Zachodnie). Acta Geologica Polonica, 25(4), 451–504. [in Polish]
Korejwo, K. (1993). Biostratigraphy of the Dinantian in the Koszalin-Chojnice area (Western Pomerania). Acta Geologica Polonica, 103, 5–47.
Korn, D. (1993). The ammonoid faunal change nera the Devonian-Carboniferous boundary. Annales de la Société Géologique de Belgique, 115(2), 581–593.
Korn, D. (2006). Ammonoideen. In Deutsche Stratigraphische Komission (Eds.) Stratigraphie von Deutschland VI Unterkarbon (Mississippium). Schriftenreihe der Deutschen Gesselschaft für Geowissenschaften, (SDGG), 4, 147-170.
Korn, D., Niedźwiecki, R., & Posieczek, J. B. (2005). Age, distribution, and phylogeny of the peculiar Late Devonian ammonoids Soliclymenia. Acta Geologica Polonica, 55(2), 99–109.
Kumpan, T., Bábek, O., Kalvoda, J., Frýda, J., & Grygar, T. M. (2014a). A high-resolution, multiproxy stratigraphic analysis of the Devonian–Carboniferous boundary sections in the Moravian Karst (Czech Republic) and a correlation with the Carnic Alps (Austria). Geological Magazine, 151(2), 201–215.
Kumpan, T., Bábek, O., Kalvoda, J., Grygar, T. M., & Frýda, J. (2014b). Sea-level and environmental changes around the Devonian-Carboniferous boundary in the Namur-Dinant Basin (S Belgium, NE France): a multiproxy stratigraphic analysis of carbonate ramp archives and its use in regional and interregional correlations. Sedimentary Geology, 311, 43–59.
Kumpan, T., Bábek, O., Kalvoda, J., Grygar, T. M., Frýda, J., Becker, R. T., & Hartenfels, S. (2015). Petrophysical and geochemical signature of the Hangenberg Events: an integrated stratigraphy of the Devonian–Carboniferous boundary interval in the Northern Rhenish Massif (Avalonia, Germany). Bulletin of Geosciences, 90(3), 667–694.
Lewowicki, S. (1959). Fauna of Clymenia Limestones from Dzikowiec near Kłodzko (Lower Silesia). Biuletyn Instytutu Geologicznego, 146, 73–119. [in Polish]
Malec, J. (1995). Devonian/Carboniferous boundary. In Guide to excursion A2, development of the Variscan Basin and epi-variscan cover at the margin of the East European Platform (Pomerania, Holy Cross Mts., Kraków Upland). Państwowy Instytut Geologiczny, 20–21.
Malec, J. (2014). The Devonian/Carboniferous boundary in the Holy Cross Mountains (Poland). Geological Quarterly, 58(2), 217–234.
Marynowski, L., & Filipiak, P. (2007). Water column euxinia and wildfire evidence during deposition of the Upper Famennian Hangenberg event horizon from the Holy Cross Mountains (Central Poland). Geological Magazine, 144, 569–595.
Marynowski, L., Filipiak, P., & Zatoń, M. (2010). Geochemical and palynological study of the Upper Famennian Dasberg event horizon from the Holy Cross Mountains (central Poland). Geological Magazine, 147, 527–550. https://doi.org/10.1017/S0016756809990835.
Marynowski, L., Zatoń, M., Rakociński, M., Filipiak, P., Kurkiewicz, S., & Pearce, T. (2012). Deciphering the upper Famennian Hangenberg black shale depositional environments based on multi-proxy record. Palaeogeography, Palaeoclimatology, Palaeoecology, 346–347, 66–86.
Marynowski, L., Pisarzowska, A., Derkowski, A., Rakociński, M., Szaniawski, R., Środoń, J., & Cohen, A. S. (2017). Influence of palaeoweathering on trace metal concentrations and environmental proxies in black shales. Palaeogeography, Palaeoclimatology, Palaeoecology, 472, 177–191.
Matyja, H. (1976). Biostratigraphy of the Devonian–Carboniferous passage beds from some selected profiles of NW Poland. Acta Geologica Polonica, 26(4), 489–539.
Matyja, H. (1993). Upper Devonian of Western Pomerania. Acta Geologica Polonica, 42(1/2), 27–94.
Matyja, H. (2006). Stratigraphy and facies development of the Devonian and Carboniferous deposits in the Pomeranian Basin and in the western part of the Baltic Basin and palaeogeography of the northern TESZ during late Palaeozoic time. Prace Państwowego Instytutu Geologicznego, 186, 79–122 [in Polish].
Matyja, H. (2008). Pomeranian basin (NW Poland) and its sedimentary evolution during Mississippian times. In M. Aretz., H-G. Herbig, I. D. Somerville (Eds.) Carboniferous platforms and basin Geological Journal, 43 (2–3), 123–150.
Matyja, H. (2009). Depositional history of the Devonian succession in the Pomeranian Basin, NW Poland. Geological Quarterly, 53, 63–92.
Matyja, H., & Stempień-Sałek, M. (1994). Devonian/Carboniferous boundary and the associated phenomena in Western Pomerania (NW Poland). Annales de la Société Géologique de Belgique, 116(2), 249–263.
Matyja, H., & Turnau, E. (1989). Conodonts and spores from the Devonian/Carboniferous boundary beds in Poland. XI Congrès International de Stratigraphie et de Géologie du Carbonifère Bejing, Compte Rendu, 3, 61–72. Bejing.
Matyja, H., Turnau, E., & Żbikowska, B. (2000). Lower Carboniferous (Mississippian) stratigraphy of northwestern Poland: conodont, miospore and ostracod zones compared. Annales Societatis Geologorum Poloniae, 70, 193–217.
Matyja, H., Sobień, K., Marynowski, L., Stempień-Sałek, M., & Małkowski, K. (2015). The expression of the Hangenberg Event (latest Devonian) in a relatively shallow-marine succession (Pomeranian Basin, Poland): the results of a multi-proxy investigation. Geological Magazine, 52(3), 400–428.
Maziane, N., Higgs, K., & Streel, M. (2002). Biometry and paleoenvironment of Retispora lepidophyta (Kedo) Playford 1976 and associated miospores in the latest Famennian nearshore marine facies, eastern Ardenne (Belgium). Review of Palaeobotany and Palynology, 118, 211–226.
McNestry, A. (1988). The palynostratigraphy of two uppermost Devonian–Lower Carboniferous borehole sections in South Wales. Review of Palaeobotany and Palynology, 56, 69–87.
Merrill, G. K., Swift, A., Ryley, C. C., Barnes, C. R., O'Brien, F. H. C., Varker, W. J., Stone, J., Saunders, R., Fredholm, D., & Jeppson, L. (1987). Recent developments in conodont concentration techniques. In R. L. Austin (Ed.) Conodonts: investigative techniques and applications. The British Micropalaeontological Society Series (pp. 54–76). Chichester: Ellis Horwood.
Mistiaen, B., & Weyer, D. (1999). Late Devonian stromtoporoid from the Sudetes Mountains (Poland), and endemicity of the Upper Famennian to Uppermost Famennian (= “Strunian”) stromatoporoid fauna in western Europe. Senckenbergiana lethaea, 79, 51–61.
Muszer, J. (1998). Makrofauna i stratygrafia osadów górnego dewonu i dolnego karbonu północnej części Pomorza Zachodniego. Prace Geologiczno-Mineralogiczne, 66, 1–153. [in Polish]
Muszer, J., & Haydukiewicz, J. (2011). The quarry in Dzikowiec as a geotouristic site and its importance for studies on the Upper Paleozoic in the Sudetes. In T. Słomka (Ed.) Geotourism a variety of aspects (pp. 243–257). Kraków: AGH University of Science and Technology. [in Polish]
Myrow, P. M., Ramezani, J., Hanson, A. E., Bowring, S. A., Racki, G., & Rakocinski, M. (2014). High precision U-Pb age and duration of the latest Devonian (Famennian) Hangenberg event, and its implications. Terra Nova, 26, 222–229.
Narkiewicz, K., Grotek, I., & Matyja, H. (1998). Dojrzałość termiczna materii organicznej w utworach górnodewońskich obszaru radomsko-lubelskiego i pomorskiego. Prace Państwowego Instytutu Geologicznego, 65, 235–244. [in Polish]
Nehring-Lefeld, M. (1990). Famennian biostratigraphy of the Kowala 1 borehole on the basis of conodots. Geological Quarterly, 34, 271–290. [in Polish]
Nikolaeva, S. V., & Bogoslovsky, B. I. (2005). Devonian ammonoids. IV. Clymeniids. Trudy Paleontologicheskogo Instituta RAN, 287, 1–220. [in Russian]
Olempska, E. (1983). The uppermost Devonian foraminifers of the Świetokrzyskie (Holy Cross) Mts., Poland. Acta Palaeontologica Polonica, 28(2–3), 393–416.
Olempska, E. (1997). Changes in the benthic ostracod assemblages across the Devonian-Carboniferous boundary in the Holy Cross Mountains, Poland. Acta Palaeontologica Polonica, 42, 291–332.
Paschall, O., Carmichael, S. K., Königshof, P., Waters, J. A., Ta, P. H., Komatsu, T., & Dombrowski, A. (2019). The Devonian-Carboniferous boundary in Vietnam: sustained ocean anoxia with a volcanic trigger for the Hangenberg Crisis? Global and Planetary Change, 175, 64–81. https://doi.org/10.1016/j.gloplacha.2019.01.021.
Percival, L. M. E., Ruhla, M., Hesselbo, S. P., Jenkyns, H. C., Mather, T. A., & Whiteside, J. H. (2017). Mercury evidence for pulsed volcanism during the end Triassic mass extinction. Proceedings of the National Academy of Sciences of the United States of America, 114(30), 7929–7934.
Perkins, R. B., Piper, D. Z., & Mason, C. E. (2008). Trace-element budgets in the Ohio/Sunbury shales of Kentucky: constraints on ocean circulation and primary productivity in the Devonian–Mississippian Appalachian Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 265, 14–29.
Prestianni, C., Sautois, M., & Denayer, J. (2016). Disrupted continental environments around the Devonian-Carboniferous Boundary: introduction of the tener event. Geologica Belgica, 19(1–2), 135–145.
Qie, W., Liu, J., Chen, J., Wang, X., Mii, H., Zhang, X., Huang, X., Yao, L., Algeo, T. J., & Luo, G. (2015). Local overprints on the global carbonate δ 13C signal in Devonian–Carboniferous boundary successions of South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 418, 290–303.
Racka, M., Marynowski, L., Filipiak, P., Sobstel, M., Pisarzowska, A., & Bond, D. P. J. (2010). Anoxic Annulata events in the Late Famennian of the Holy Cross Mountains (Southern Poland): geochemical and palaeontological record. Palaeogeography, Palaeoclimatology, Palaeoecology, 297, 549–575.
Racki, G., Racka, M., Matyja, H., & Devleeschouwer, X. (2002). The Frasnian/Famennian boundary interval in the South Polish-Moravian shelf basins: integrated event-stratigraphical approach. Palaeogeography, Palaeoclimatology, Palaeoecology, 181, 251–297.
Racki, G., Rakociński, M., & Marynowski, L. (2018a). Anomalous Upper Devonian mercury enrichments: comparison of inductively coupled plasma–mass spectrometry (ICPMS) and AAS analytical data. Geological. Quarterly, 62(3), 487–495. https://doi.org/10.7306/gq.1419.
Racki, G., Rakociński, M., Marynowski, L., & Wignall, P. B. (2018b). Mercury enrichment and the Frasnian-Famennian biotic crisis: a volcanic trigger proved? Geology, 46(6), 543–546.
Rakociński, M. (2011). Sclerobionts on upper Famennian cephalopods from the Holy Cross Mountains, Poland. Palaeobiodiversity and Palaeoenvironments, 91(1), 63–73.
Rimmer, S. M. (2004). Geochemical paleoredox indicators in Devonian-Mississippian black shales, Central Appalachian Basin (USA). Chemical Geology, 206, 373–391.
Rimmer, S. M., Thompson, J. A., Goodnight, S. A., & Robl, T. L. (2004). Multiple controls on the preservation of organic matter in Devonian-Mississippian marine black shales: geochemical and petrographic evidence. Palaeogeography, Palaeoclimatology, Palaeoecology, 215, 125–154.
Romanek, A., & Rup, M. (1990). Lithostratigraphic subdivision of the Devonian sequence in the Kowala 1 borehole. Geological Quarterly, 34, 221–242. [in Polish]
Różkowska, M. (1969). Famennian Tetracoralloid and Heterocoralloid fauna from the Holy Cross Mountains (Poland). Acta Palaeontologia Polonica, 14(1), 5–187.
Sadykov, A.M. (1962). Srednepaleozojskie dvustvorčatye molluski Atasu (Centralny Kazachstan). [Middle paleozoik Bivalvia from Atasu (Central Kazachstan). 1–114. Alma-Ata (Academija Nauk Kazachskoj SSR, Instytut Geologičeskich Nauk). [in Russian]
Sandberg, C. A., Ziegler, W., Leuteritz, K., & Brill, S. M. (1978). Phylogeny, speciation, and zonation of Siphonodella (Conodonta, Upper Devonian and Lower Carboniferous). Newsletters on Stratigraphy, 7, 102–120.
Sandberg, C.A., Morrow, J.R., & Ziegler, W. (2002) Late Devonian sea-level changes, catastrophic events, and mass extinctions. In C. Koeberl & K.G. MacLeod (Eds.) Catastrophic events and mass extinctions: impacts and beyond. Boulder, Colorado (pp. 473–487). Special Paper 356 Geological Society of America.
Schindewolf, O. H. (1937). Zur Stratigraphie und Palaeontologie der Wocklumer Schichten (Oberdevon). Abhandlungen der Preußischen Geologischen Landesanstalt, Neue Folge, 178, 1–132.
Schönlaub, H. P., Attrep, M., Boeckelmann, K., Dreesen, R., Feist, R., Fenninger, A., Hahn, G., Klein, P., Korn, D., Kratz, R., Magaritz, M., Orth, C. J., & Schramm, J.-M. (1992). The Devonian/Carboniferous boundary in the Carnic Alps (Austria)—a multidisciplinary approach. Jahrbuch der geologischen Bundesanstalt, 135, 57–98.
Schönlaub, H., Kreutzer, L., Joachimski, M.M., & Buggisch, W. ( 1994). Sedimentology and geochemistry of boundary sections from the Northern Calcareous (K/T) and Carnic Alps (O/S, S/D, F/F, D/C, P/T). In M.M. Joachimski (Ed.) Sedimentology and geochemistry of boundary sections from the northern calcareous (K/T) and Carnic Alps (O/S, S/D, F/F, D/C, P/T), Austria—A field guide. Erlanger Geologische Abhandlungen, 122, 77–103.
Spalletta, C., Perri, M. C., Over, D. J., & Corradini, C. (2017). Famennian (Upper Devonian) conodont zonation: revised global standard. Bulletin of Geosciences, 92(1), 31–57.
Stempień-Sałek, M. (2002). Miospore taxonomy and stratigraphy of Upper Devonian and lowermost Carboniferous in Western Pomerania (NW Poland). Annales Societatis Geologorum Poloniae, 72(2), 163–190.
Stone, J. (1987) Review of investigative techniques used in the study of conodonts. In R.L. Austin (Ed.) Conodonts: investigative techniques and applications. The British Micropalaeontological Society Series. Chichester: Ellis Horwood, 17-34.
Streel, M. (1966). Critères palynologiques pour une stratigraphie détaillée du Tn1a dans les bassins Ardenno-Rhénans. Annales de la Société Géologique de Belgique, 89, 65–96.
Streel, M. (1999). Quantitative palynology of Famennian events in the Ardenne-Rhine regions. In R. Feist, J.A. Talent & A. Daurer (Eds.) North Gondwana: Mid – Paleozoic Terranes, stratigraphy and biota. Abhandlungen der Geologischen Bundesanstalt, 54, 201–212.
Streel, M. & Marshall, J.E.A. (2006). Devonian-Carboniferous boundary global correlations and their paleogeographic implications for the assembly of Pangaea. In Th. E. Wong (Ed.) Proceedings of the XVth International Congress on Carboniferous and Permian Stratigraphy, Utrecht, the Netherlands, 10-16 August 2003. Royal Netherlands Academy of Arts and Sciences, 481-496.
Streel, M., Higgs, K., Loboziak, S., Riegel, W., & Steemans, P. (1987). Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions. Review of Palaeobotany and Palynology, 50, 211–229.
Streel, M., Bełka, Z., Dreesen, R., Durkina, A. V., Gross-Uffenorde, H., Hance, L., Hartkopf-Frőder, C., Haydukiewicz, J., Korn, D., Perri, M. C., Piecha, M., & Spaletta, C. (2004). Relation of the neritic microfaunas and continental microflora with the conodont and other pelagic faunas within the latest part of the Famennian. SDS Annual Meeting Rabat, 2004, 67–73.
Szulczewski, M. (1971). Upper Devonian conodonts, stratigraphy and facial development in the Holy Cross Mts. Acta Geologica Polonica, 21, 1–129.
Thorez, J. (1985). Argillogenesis and the hydrolysis index. Acta Mineralogica-Petrographica, 29, 313–338.
Tragelehn, H. (2010). Short note on the origin of the conodon genus Siphonodella in the Famennian. SDS Newsletter, 25, 41–43.
Trela, W., & Malec, J. (2007). Carbon isotope record in sediment of the Devonian-Carboniferous boundary in the southern Holy Cross Mountains. Przegląd Geologiczny, 55(5), 411–415. [in Polish]
Turnau, E. (1978). Spore zonation of uppermost Devonian and lower Carboniferous of Western Pomerania. Mededelingen Rijks Geologische Dienst, 30(1), 1–34.
Turnau, E. (1979). Correlations of Upper Devonian and Carboniferous deposits of Western Pomerania based on miospore study. Rocznik Polskiego Towarzystwa Geologicznego, 49(3/4), 231–269. [in Polish]
Turnau, E. (1985). Devonian-Carboniferous boundary in the borehole Kowala 1 (Southern Holy Cross Mts., Poland); spores. Bulletin of the Polish Academy of Sciences, Earth Sciences, 33, 2–5.
Turnau, E. (1990). Spore zones of Famennian and Tournaisian deposits from the Kowala 1 borehole (in Polish). Geological Quarterly, 34, 291–304. [in Polish]
Velde, B., & Meunier, A. (2008). The origin of clay minerals in soils and weathered rocks. Berlin Heidelberg: Springer-Verlag.
Wajsprych, B. (1995). The Bardo Mts rock complex: the Famennian–Lower Carboniferous preflysch (platform) – to flysch (foreland) basin succession, the Sudetes. Guide to Excursion B2 of XIII Inter. Congr. on Carboniferous–Permian 28.08-02.09. Kraków, Państwowy Instytut Geologiczny, Warszawa, pp. 23–42.
Weissert, H., Joachimski, M., & Sarnthein, M. (2008). Chemostratigraphy. Newsletters on Stratigraphy, 42(3), 145–179.
Weyer, D. (1965). Zur Ammonoideen-Fauna der Gattendorfia-Stufe von Dzikowiec (Ebersdorf) in Dolny Śląsk (Niederschlesien), Polen. Berichte der Geologischen Gessellschaft in der DDR, 10, 443–464.
Wood, G. D., Gabriel, A. M. & Lawson, J. C. (1996). Palynological techniques – processing and microscopy. In J. Jansonius & D. C. McGregor (Eds.) Palynology: principles and applications (pp., 29–50). 1, American Association of Stratigraphic Palynologists.
Woroncowa-Marcinowska, T. (2011). Late Famennian (Devonian) Balviinae (Ammonoidea) from the Holy Cross Mountains, Poland. Acta Geologica Polonica, 61, 35–45.
Woroncowa-Marcinowska, T. (2017). Latest Devonian silicified agglutinated foraminifera near the Hangenberg event horizon from Kowala (Holy Cross Mountains) and Wapnica quarries (Sudetes), Poland. In M. A. Kaminski, & L. Alegret (Eds.) Proceedings of the Ninth International Workshop on Agglutinated Foraminifera (pp. 251-261). Grzybowski Foundation Special Publication, 22, 251-261.
Xu, D.-Y., Yan, Z., Zhang, Q.-W., Shen, Z.-D., Sun, Y.-Y., & Ye, L.-F. (1986). Significance of a δ13C anomaly near the Devonian/Carboniferous boundary at the Muhua section, South China. Nature, 321, 854–855.
Żakowa, H. (1983). Małże rodzaju Guerichia Rzehak, 1910 z utworów famenu i turneju Jabłonnej, Góry Świętokrzyskie. Biuletyn Instytutu Geologicznego, 345, 177–223. [in Polish]
Żakowa, H., & Pawłowska, J. (1961). The Lower Carboniferous in the area between Radlin and Górno in the Kielce-Łagów synclinorium (Święty Krzyż Mountains). Biuletyn Instytutu Geologicznego, 167, 101–166. [in Polish]
Żakowa, H., & Radlicz, K. (1990). Macro- and microfauna and petrography of the Famennian deposits in the Kowala 1 borehole. Geological Quarterly, 34, 243–270. [in Polish]
Żakowa, H., Nehring-Lefeld, M., & Malec, J. (1985). Devonian-Carboniferous Boundary in the borehole Kowala 1 (Southern Holy Cross Mts., Poland); micro- and macrofauna. Bulletin of the Polish Academy of Sciences, Earth Sciences, 33, 87–95.
Zapalski, M. K., & Berkowski, B. (2012). The oldest species of ?Yavorskia (Tabulata) from the Upper Famennian of the Holy Cross Mountains (Poland). Acta Geologica Polonica, 62(2), 197–204.
Zapalski, M. K., Berkowski, B., & Wrzołek, T. (2016). Tabulate corals after the Frasnian/Famennian Crisis: a unique fauna from the Holy Cross Mountains, Poland. PLoS One, 11(3), e0149767. https://doi.org/10.1371/journal.pone.0149767.
Żbikowska, B. (1992). Entomozoidy (małżoraczki) górnego dewonu i najniższego karbonu Pomorza Zachodniego. Przegląd Geologiczny, 40(10), 612. [in Polish]
Ziegler, W. & Sandberg, Ch.A. (1984). Palmatolepis-based revision of upper part of standard Late Devonian conodont zonation. In D.L. Clark (Ed.) Conodont biofacies and provincialism (pp. 179–194). Geological Society of America Special Papers, 196.
Ziegler, W., & Sandberg, C. A. (1990). The Late Devonian standard conodont zonation. Courier Forschungsinstitut Senckenberg, 121, 1–115.
Acknowledgements
This work has been partly financed by the Polish Geological Institute-National Research Institute Project 61.2908.1304.00.0. The authors are deeply grateful to Dr. Sandra Kaiser and an anonymous reviewer, who carefully reviewed the manuscript and offered many suggestions. Special thanks go to Bronisław Andrzej Matyja, University of Warsaw, first of all for his valuable comments and suggested improvements to this paper, as well as help during the fieldwork. Anna Żylińska, University of Warsaw, took care of linguistic problems. Michał Janik is acknowledged for assistance with all figures, and Leszek Giro, both PGI – NRI, for assistance with all SEM photographs.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “Global review of the Devonian-Carboniferous Boundary”.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Matyja, H., Woroncowa-Marcinowska, T., Filipiak, P. et al. The Devonian/Carboniferous boundary interval in Poland: multidisciplinary studies in pelagic (Holy Cross Mountains and Sudetes) and ramp (Western Pomerania) successions. Palaeobio Palaeoenv 101, 421–472 (2021). https://doi.org/10.1007/s12549-020-00442-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-020-00442-3