Deformed Wing Virus in Two Widespread Invasive Ants: Geographical Distribution, Prevalence, and Phylogeny

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Isolation and RT-PCR Detection of DWV
2.3. Phylogenetic Analysis
3. Results
3.1. DWV Prevalence in LCA and YCA
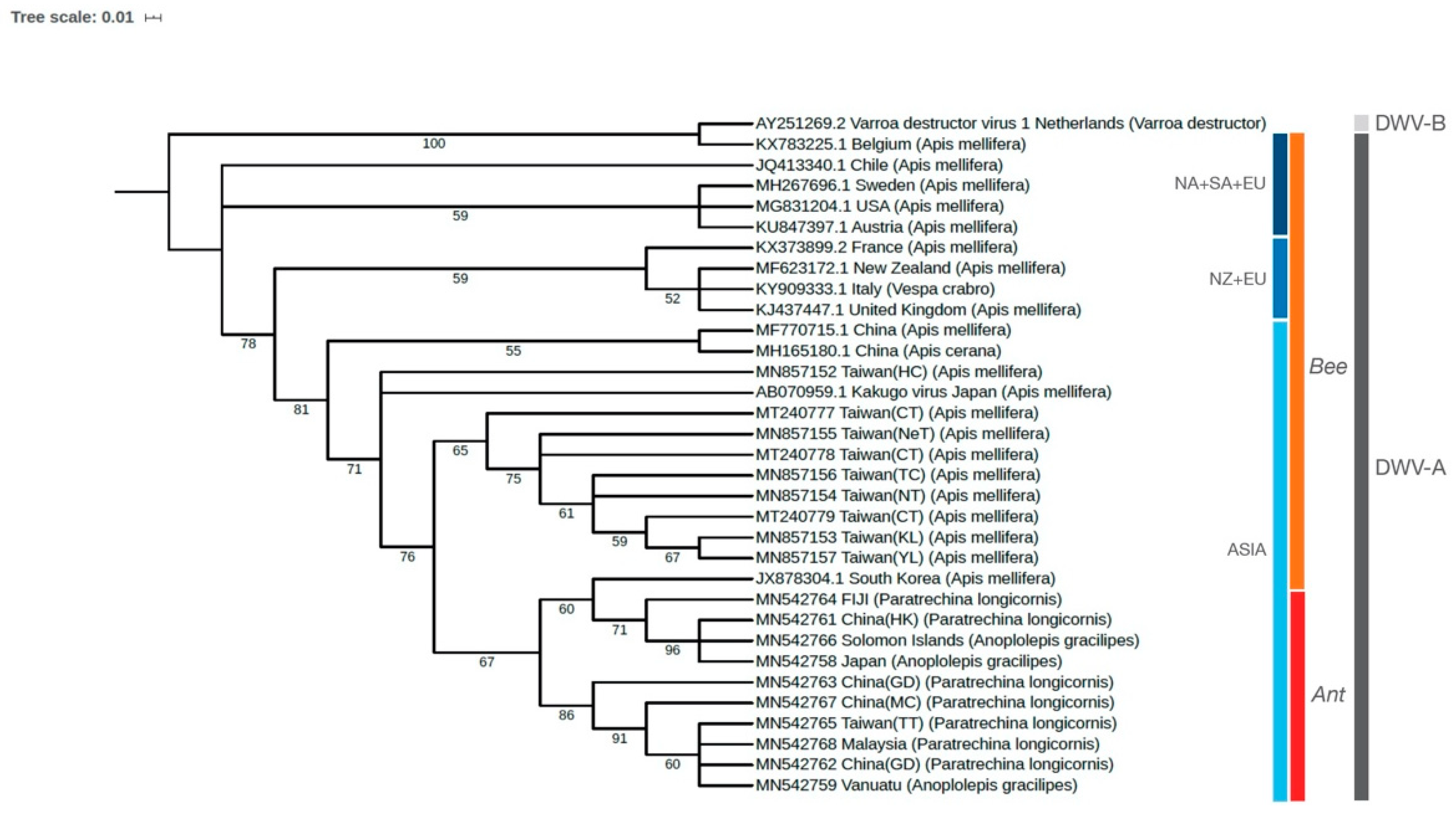
3.2. Phylogenetic Relationships of DWV in Ants and Honey Bees
4. Discussion
4.1. DWV Prevalence in Globally Invasive Ants
4.2. Origin of DWV in Invasive Ants
4.3. The role of Invasive Ants in DWV Transmission
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Vilcinskas, A. Pathogens as biological weapons of invasive species. PLoS Pathog. 2015, 11, e1004714. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.M.; Warnock, N.D.; Tosh, D.G.; McInnes, C.; Everest, D.; Montgommery, W.I.; Scantlebury, M.; Marks, N.; Dick, J.T.; Reid, N. Squirrelpox virus: Assessing prevalence, transmission and environmental degradation. PLoS ONE 2014, 9, e89521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurnell, J.; Rushton, S.P.; Lurz, P.W.W.; Sainsbury, A.W.; Nettleton, P.; Shirley, M.D.F.; Bruemmer, C.; Geddes, N. Squirrel poxvirus: Landscape scale strategies for managing disease threat. Biol. Conserv. 2006, 131, 287–295. [Google Scholar] [CrossRef]
- Alger, S.A.; Burnham, P.A.; Boncristiani, H.F.; Brody, A.K. RNA virus spillover from managed honeybees (Apis mellifera) to wild bumblebees (Bombus spp.). PLoS ONE 2019, 14, e0217822. [Google Scholar] [CrossRef] [Green Version]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proc. R. Soc. 2015, 282, 20142124. [Google Scholar] [CrossRef] [Green Version]
- Sébastien, A.; Lester, P.J.; Hall, R.J.; Wang, J.; Moore, N.E.; Gruber, M.A.M. Invasive ants carry novel viruses in their new range and form reservoirs for a honeybee pathogen. Biol. Lett. 2015, 11, 20150610. [Google Scholar] [CrossRef]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; Vanengelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA viruses in hymenopteran pollinators: Evidence of inter-Taxa virus transmission via pollen and potential impact on non-Apis hymenopteran species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef]
- Fürst, M.; McMahon, D.P.; Osborne, J.; Paxton, R.; Brown, M. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef]
- Gisder, S.; Genersch, E. Viruses of commercialized insect pollinators. J. Invertebr. Pathol. 2017, 147, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Bailes, E.J.; Deutsch, K.R.; Bagi, J.; Rondissone, L.; Brown, M.J.F.; Lewis, O.T. First detection of bee viruses in hoverfly (syrphid) pollinators. Biol. Lett. 2018, 14, 20180001. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, L.L.; Blinder, M.; Grincavitch, C.; Jelinek, A.; Mann, E.K.; Merva, L.A.; Metz, L.E.; Zhao, A.Y.; Irwin, R.E.; McArt, S.H.; et al. Bee pathogen transmission dynamics: Deposition, persistence and acquisition on flowers. Proc. R. Soc. 2019, 286, 20190603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitt, A.L.; Singh, R.; Cox-Foster, D.L.; Rajotte, E.; Hoover, K.; Ostiguy, N.; Holmes, E.C. Cross-species transmission of honey bee viruses in associated arthropods. Virus Res. 2013, 176, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Yañez, O.; Piot, N.; Dalmon, A.; de Miranda, J.R.; Chantawannakul, P.; Panziera, D.; Amiri, E.; Smagghe, G.; Schroeder, D.; Chejanovsky, N. Bee viruses: Routes of infection in Hymenoptera. Front. Microbiol. 2020, 11, 943. [Google Scholar] [CrossRef] [PubMed]
- Gruber, M.A.M.; Cooling, M.; Baty, J.W.; Buckley, K.; Friedlander, A.; Quinn, O.; Russell, J.F.E.J.; Sébastien, A.; Lester, P.J. Single-stranded RNA viruses infecting the invasive Argentine ant, Linepithema humile. Sci. Rep. 2017, 7, 3304. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.N.; Shepherd, T.F.; Rangel, J. The detection of honey bee (Apis mellifera)-associated viruses in ants. Sci. Rep. 2020, 10, 2923. [Google Scholar] [CrossRef] [Green Version]
- Dobelmann, J.; Felden, A.; Lester, P.J. Genetic strain diversity of multi-host RNA viruses that infect a wide range of pollinators and associates is shaped by geographic origins. Viruses 2020, 12, 358. [Google Scholar] [CrossRef] [Green Version]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. Parasites in bloom: Flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. R. Soc. 2015, 282, 20151371. [Google Scholar] [CrossRef] [Green Version]
- Simoes, M.R.; Giannotti, E.; Tofolo, V.C.; Pizano, M.A.; Firmino, E.L.B.; Antonialli-Junior, W.F.; Andrade, L.H.C.; Lima, S.M. Morphological and chemical characterization of the invasive ants in hives of Apis mellifera scutellata Lepeletier (Hymenoptera: Apidae). Neotrop. Entomol. 2015, 45, 72–79. [Google Scholar] [CrossRef]
- Kojima, Y.; Toki, T.; Morimoto, T.; Yoshiyama, M.; Kimura, K.; Kadowaki, T. Infestation of Japanese native honey bees by tracheal mite and virus from non-native European honey bees in Japan. Microb. Ecol. 2011, 62, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, T.; Kojima, Y.; Yoshiyama, M.; Kimura, K.; Yang, B.; Kadowaki, T. Molecular identification of chronic bee paralysis virus infection in Apis mellifera colonies in Japan. Viruses 2012, 4, 1093–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.H.; Lo, C.F.; Nai, Y.S.; Wang, C.Y.; Chen, Y.R.; Huang, W.F.; Chien, T.Y.; Wu, C.Y. Honey bee colony collapse disorder. Formosan Entomol. 2009, 29, 119–138. [Google Scholar]
- Lu, M.C.; Wu, H.H.; Lo, Y.C. Distribution and prevention strategy of honeybee (Apis mellifera L.) viruses in Taiwan (In Chinese). In Proceedings of the Symposium on the Agriculture Technology, Taipei, Taiwan, 3 March 2014; pp. 50–61. [Google Scholar]
- Forsgren, E.; Wei, S.; Guiding, D.; Zhiguang, L.; Tran, T.V.; Tang, P.T.; Truong, T.A.; Dinh, T.Q.; Fries, I. Preliminary observations on possible pathogen spillover from Apis mellifera to Apis cerana. Apidologie 2015, 46, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Bi, J.; Wang, L.; Zhou, D.; Ma, X.T.; Li, W.G.; Zhao, W.Z.; Yin, G.F.; Liu, J.P.; He, S.Y. Prevalence of four common bee RNA viruses in eastern bee populations in Yunnan Province, China. J. Veterinar. Sci. Technol. 2016, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Berényi, O.; Bakonyi, T.; Derakhshifar, I.; Köglberger, H.; Topolska, G.; Ritter, W.; Pechhacker, H.; Nowotny, N. Phylogenetic analysis of deformed wing virus genotypes from diverse geographic origins indicates recent global distribution of the virus. Appl. Environ. Microbiol. 2007, 73, 3605–3611. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.L. Surveillance of parasites and diseases of honeybees in Papua New Guinea and Indonesia. CSIRO Rep. 2008, 1, 1–41. [Google Scholar]
- Sanpa, S.; Chantawannakul, P. Survey of six bee viruses using RT-PCR in northern Thailand. J. Invertebr. Pathol. 2009, 100, 116–119. [Google Scholar] [CrossRef]
- Mookhploy, W.; Kimura, K.; Disayathanoowat, T.; Yoshiyama, M.; Hondo, K.; Chantawannakul, P. Capsid gene divergence of Black queen cell virus isolates in Thailand and Japan honey bee species. J. Econ. Entomol. 2015, 108, 1460–1464. [Google Scholar] [CrossRef]
- Chanpanitkitchote, P.; Chen, Y.; Evans, J.D.; Li, W.; Li, J.; Hamilton, M.; Chantawannakul, P. Acute bee paralysis virus occurs in the Asian honey bee Apis cerana and parasitic mite Tropilaelaps mercedesae. J. Invertebr. Pathol. 2018, 151, 131–136. [Google Scholar] [CrossRef]
- Carr, A.J. Asian Honeybee: Possible Environmental Impacts. Prepared for the Department of Sustainability, Environment, Water, Population and Communities. Available online: https://www.environment.gov.au/system/files/resources/16422f90-1acf-4223-a347-740f48401ced/files/asian-bees.pdf (accessed on 13 August 2011).
- Lloyd, D.; Somerville, D.; Roberts, J. Novel Approaches for Increasing Participation in the Honeybee Industries of the Pacific. Available online: https://aciar.gov.au/sites/default/files/project-page-docs/final_report_ls-2017-100_0.pdf (accessed on 18 October 2019).
- Roberts, J.M.K.; Anderson, D.L.; Durr, P.A. Absence of deformed wing virus and Varroa destructor in Australia provides unique perspectives on honeybee viral landscapes and colony losses. Sci. Rep. 2017, 7, 6925. [Google Scholar] [CrossRef]
- Roper, T.; Gonzalez, M. Pacific Horticultural and Agricultural Market Access Program (PHAMA) Technical Report 49: Disease Survey of Honey Bees in Fiji (FIJI15). Available online: https://phama.com.au/wp-content/uploads/2016/06/TR-49-FIJI15-Disease-Survey-of-Honey-Bees-in-Fiji-v1.0-FINAL.pdf (accessed on 27 August 2013).
- Taylor, B.; Roper, T. Pacific Horticultural and Agricultural Market Access Program (PHAMA) Technical Report 34: Disease Survey of Honey Bees in Vanuatu (VAN10). Available online: https://phamaplus.com.au/wp-content/uploads/2016/07/TR-34-VAN10-Disease-Survey-of-Honey-Bees-FINAL-160413.pdf (accessed on 15 April 2013).
- Chen, Y.; Zhao, Y.; Hammond, J.; Hsu, H.T.; Evans, J.; Feldlaufer, M. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. J. Invertebr. Pathol. 2004, 87, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsik, C.G.; Tayal, A.; Diesh, C.M.; Unni, D.R.; Emery, M.L.; Nguyen, H.N.; Hagen, D.E. Hymenoptera Genome Database: Integrating genome annotations in HymenopteraMine. Nucleic Acids Res. 2016, 44, 793–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.W.; Chiu, M.C.; Lee, C.C.; Lee, C.Y.; Yang, C.C.S. The association between virus prevalence and intercolonial aggression levels in the yellow crazy ant, Anoplolepis gracilipes (Jerdon). Insects 2019, 10, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A new and scalable tool for the selection of DNA and protein evolutionary models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Fonio, E.; Heyman, Y.; Boczkowski, L.; Gelblum, A.; Kosowski, A.; Korman, A.; Feinerman, O. A locally-blazed ant trail achieves efficient collective navigation despite limited information. eLife 2016, 5, e20185. [Google Scholar] [CrossRef] [Green Version]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef] [Green Version]
- Lester, P.J.; Buick, K.H.; Baty, J.W.; Felden, A.; Haywood, J. Different bacterial and viral pathogens trigger distinct immune responses in a globally invasive ant. Sci Rep. 2019, 9, 5780. [Google Scholar] [CrossRef] [Green Version]
- Schläppi, D.; Chejanovsky, N.; Yañez, O.; Neumann, P. Foodborne transmission and clinical symptoms of honey bee viruses in ants Lasius spp. Viruses 2020, 12, 321. [Google Scholar] [CrossRef] [Green Version]
- Cooling, M.; Gruber, M.A.M.; Hoffmann, B.D.; Sébastien, A.; Lester, P.J. A metatranscriptomic survey of the invasive yellow crazy ant, Anoplolepis gracilipes, identifies several potential viral and bacterial pathogens and mutualists. Insectes Soc. 2017, 64, 197–207. [Google Scholar] [CrossRef]
- Wetterer, J.K. Worldwide distribution and potential spread of the long-legged ant, Anoplolepis gracilipes (Hymenoptera: Formicidae). Sociobiology 2005, 45, 77–97. [Google Scholar]
- Ito, F.; Asfiya, W.; Kojima, J.I. Discovery of independent-founding solitary queens in the yellow crazy ant Anoplolepis gracilipes in East Java, Indonesia (Hymenoptera: Formicidae). Entomol. Sci. 2016, 19, 312–314. [Google Scholar] [CrossRef]
- Yang, C.C.S.; Yu, Y.C.; Valles, S.M.; Oi, D.H.; Chen, Y.C.; Shoemaker, D.; Wu, W.J.; Shih, C.J. Loss of microbial (pathogen) infections associated with recent invasions of the red imported fire ant Solenopsis invicta. Biol. Invasions 2010, 12, 3307–3318. [Google Scholar] [CrossRef]
- Felden, A.; Paris, C.; Chapple, D.G.; Suarez, A.V.; Tsutsui, N.D.; Lester, P.J.; Gruber, M.A.M. Native and introduced Argentine ant populations are characterised by distinct transcriptomic signatures associated with behaviour and immunity. NeoBiota 2019, 49, 105–126. [Google Scholar] [CrossRef]
- Wale, N.; Sim, D.G.; Jones, M.J.; Salathe, R.; Day, T.; Read, A.F. Resource limitation prevents the emergence of drug resistance by intensifying within-host competition. Proc. Natl. Acad. Sci. USA 2017, 114, 13774–13779. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.C.; Lin, C.Y.; Hsu, H.W.; Yang, C.C.S. Complete genome sequences of two novel dicistroviruses detected in the yellow crazy ant, Anoplolepis gracilipes. Arch. Virol. 2020, 165, 2715–2719. [Google Scholar] [CrossRef]
- Roberts, M.K.; Simbiken, N.; Dale, C.; Armstrong, J.; Anderson, D.L. Tolerance of honey bees to Varroa mite in the absence of deformed wing virus. Viruses 2020, 12, 575. [Google Scholar] [CrossRef]
- Martin, S.J.; Brettell, L.E. Deformed wing virus in honeybees and other insects. Annu. Rev. Virol. 2019, 6, 12.1–12.21. [Google Scholar] [CrossRef] [PubMed]
- Bradbear, N. Bees and Their Role in Forest Livelihoods: A Guide to the Services Provided by Bees and the Sustainable Harvesting, Processing and Marketing of Their Products. Available online: http://www.fao.org/3/a-i0842e.pdf (accessed on 15 November 2009).
- Hinton, J.; Schouten, C.; Austin, A.; Lloyd, D. An overview of rural development and small-scale beekeeping in Fiji. Bee World 2020, 97, 39–44. [Google Scholar] [CrossRef]
- Anderson, D.L. Pests and pathogens of the honeybee (Apis mellifera L.) in Fiji. J. Apicul. Res. 1990, 29, 53–59. [Google Scholar] [CrossRef]
- Manley, R.; Boots, M.; Wilfert, L. Emerging viral disease risk to pollinating insects: Ecological, evolutionary and anthropogenic factors. J. Appl. Ecol. 2015, 52, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Schläppi, D.; Lattrell, P.; Yañez, O.; Chejanovsky, N.; Neumann, P. Foodborne transmission of deformed wing virus to ants (Myrmica rubra). Insects 2019, 10, 394. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Region | DWV | ||
|---|---|---|---|
| LCA | YCA | Reference | |
| Prevalence (%) (No. of DWV—positive colony/total no. of the tested colony) [a] | (+/−) [b] | ||
| Regions with Abundant Collections c | |||
| Japan | 15.4 (2/13) | 3.4 (1/29) | + [21,22] |
| Taiwan | 4.6 (4/87) | − (0/68) | + [23,24] |
| Malaysia | 13.6 (3/22) | − (0/49) | N d |
| Regions with Few Collections c | |||
| China | 26.7 (4/15) | − (0/4) | + [25,26] |
| Nepal | − (0/4) | + [27] | |
| Indonesia | − (0/2) | − (0/5) | − [28] |
| Sri Lanka | − (0/1) | + [27] | |
| Thailand | − (0/9) | − (0/7) | + [29,30,31] |
| Singapore | − (0/5) | − (0/11) | N d |
| Solomon Island | 100 (1/1) | − [32,33] | |
| Australia | − (0/7) | − (0/2) | − [34] |
| Vanuatu | 50 (1/2) | − [35] | |
| Fiji | 33.3 (2/6) | − (0/1) | − [36] |
| Total | 8.82% (170) | 1.67% (180) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-Y.; Lee, C.-C.; Nai, Y.-S.; Hsu, H.-W.; Lee, C.-Y.; Tsuji, K.; Yang, C.-C.S. Deformed Wing Virus in Two Widespread Invasive Ants: Geographical Distribution, Prevalence, and Phylogeny. Viruses 2020, 12, 1309. https://doi.org/10.3390/v12111309
Lin C-Y, Lee C-C, Nai Y-S, Hsu H-W, Lee C-Y, Tsuji K, Yang C-CS. Deformed Wing Virus in Two Widespread Invasive Ants: Geographical Distribution, Prevalence, and Phylogeny. Viruses. 2020; 12(11):1309. https://doi.org/10.3390/v12111309
Chicago/Turabian StyleLin, Chun-Yi, Chih-Chi Lee, Yu-Shin Nai, Hung-Wei Hsu, Chow-Yang Lee, Kazuki Tsuji, and Chin-Cheng Scotty Yang. 2020. "Deformed Wing Virus in Two Widespread Invasive Ants: Geographical Distribution, Prevalence, and Phylogeny" Viruses 12, no. 11: 1309. https://doi.org/10.3390/v12111309