Different Species Requirements within a Heterogeneous Spring Complex Affects Patch Occupancy of Threatened Snails in Australian Desert Springs


Abstract
:1. Introduction
2. Methods
2.1. Site Description
2.2. Study Species
2.3. Environmental Variables
2.4. Snail Survey Method
3. Data Analysis
4. Results
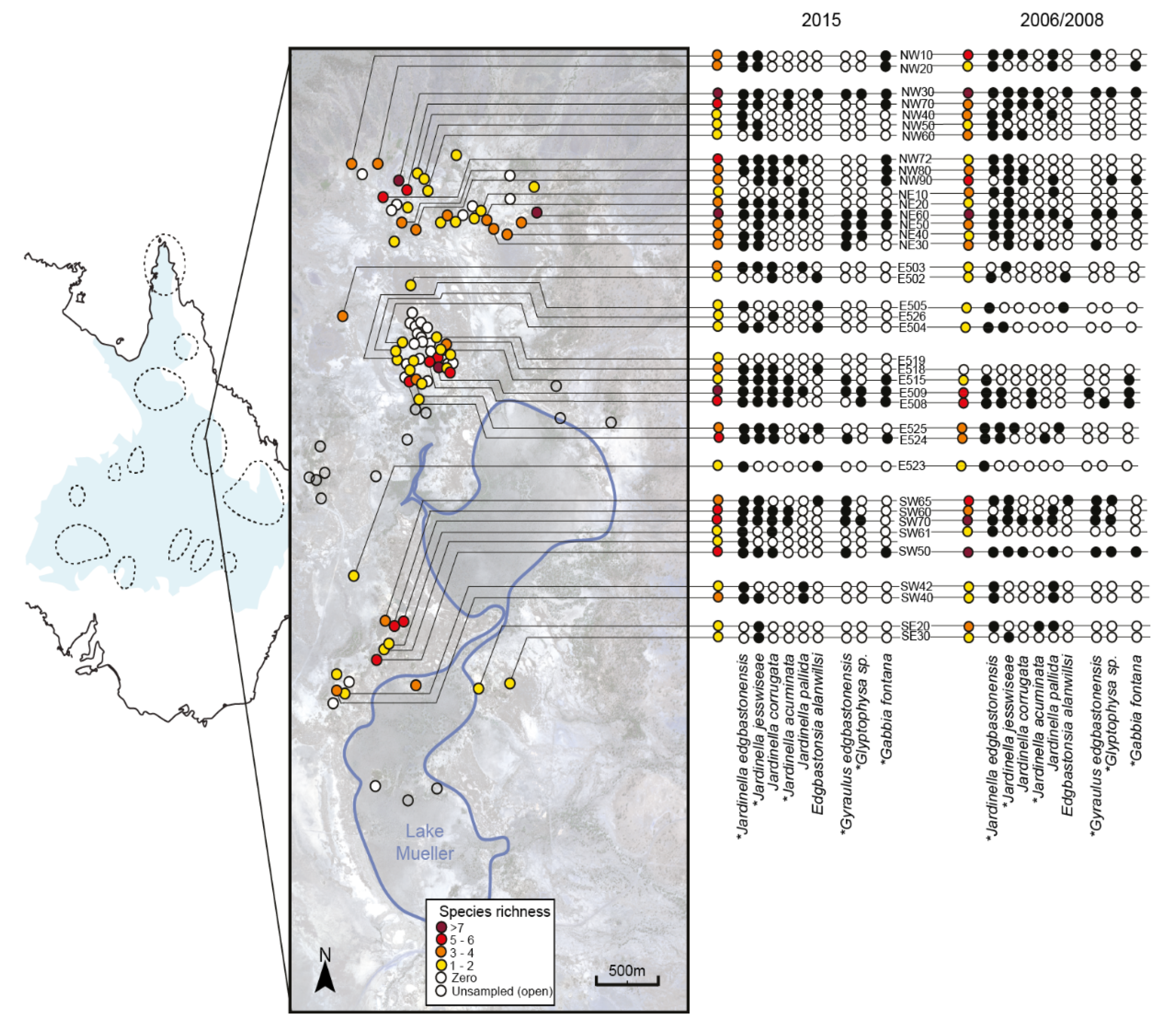
4.1. Patterns of Occupancy and Changes since 2006–2008
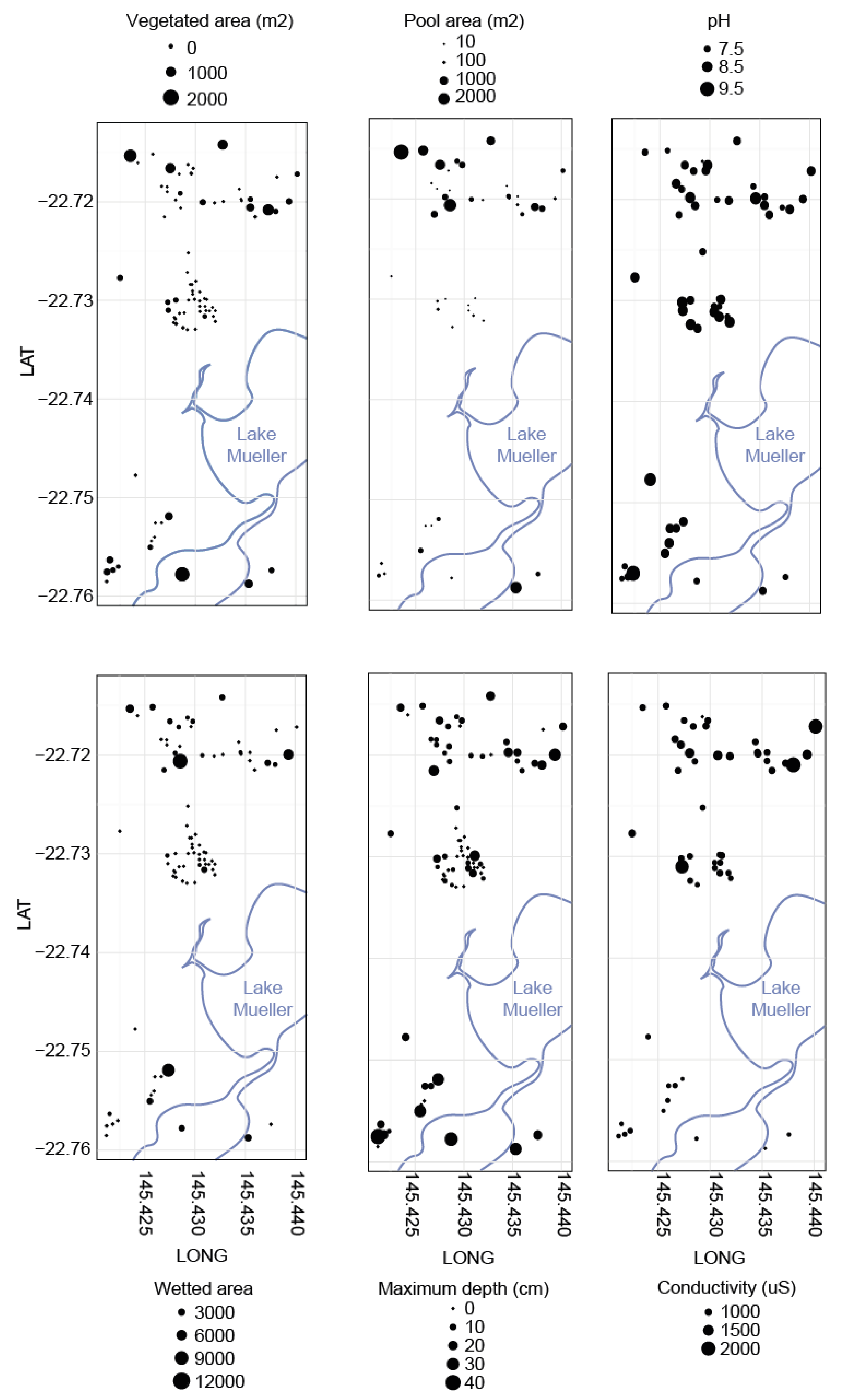
4.2. Patterns of Environmental Heterogeneity
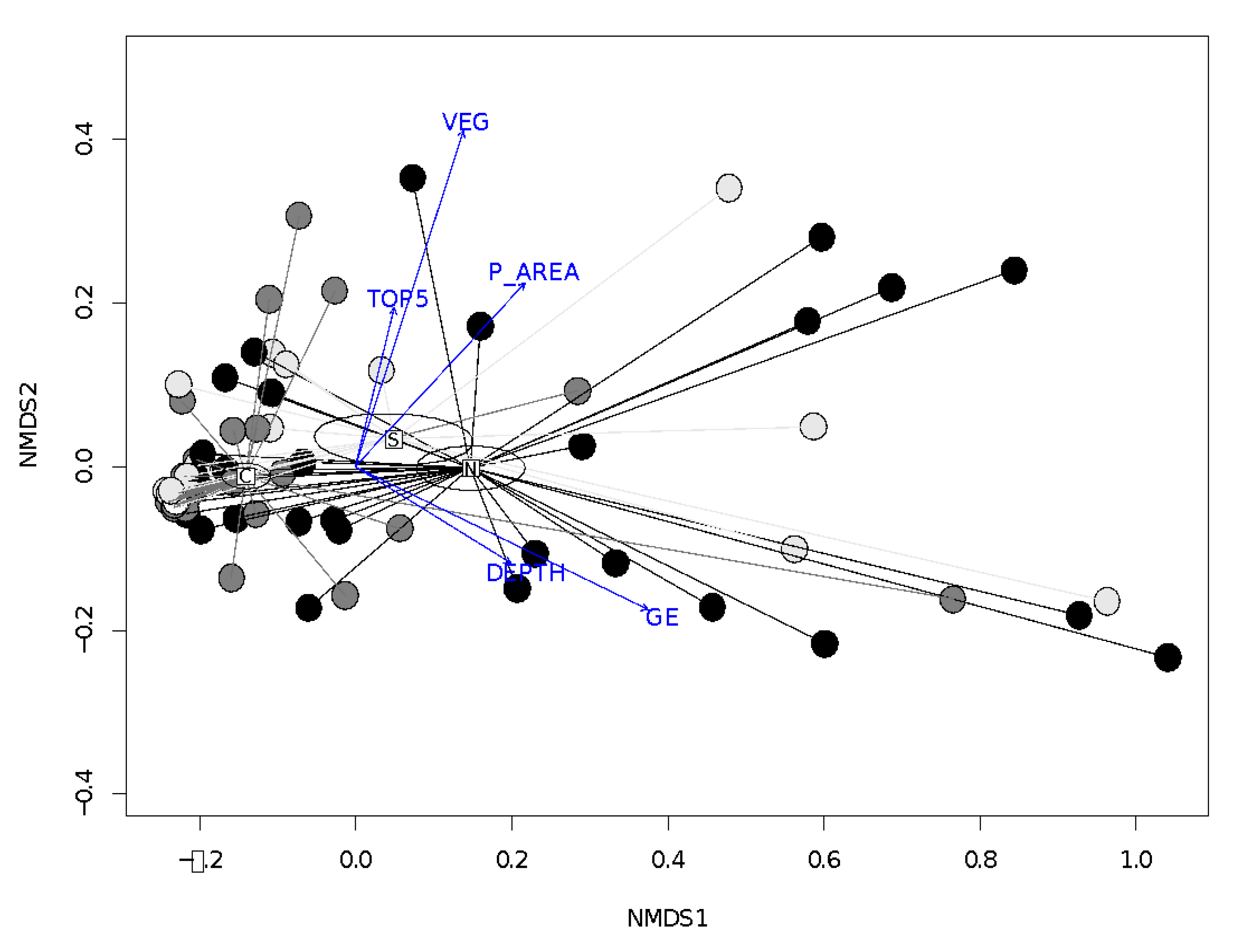
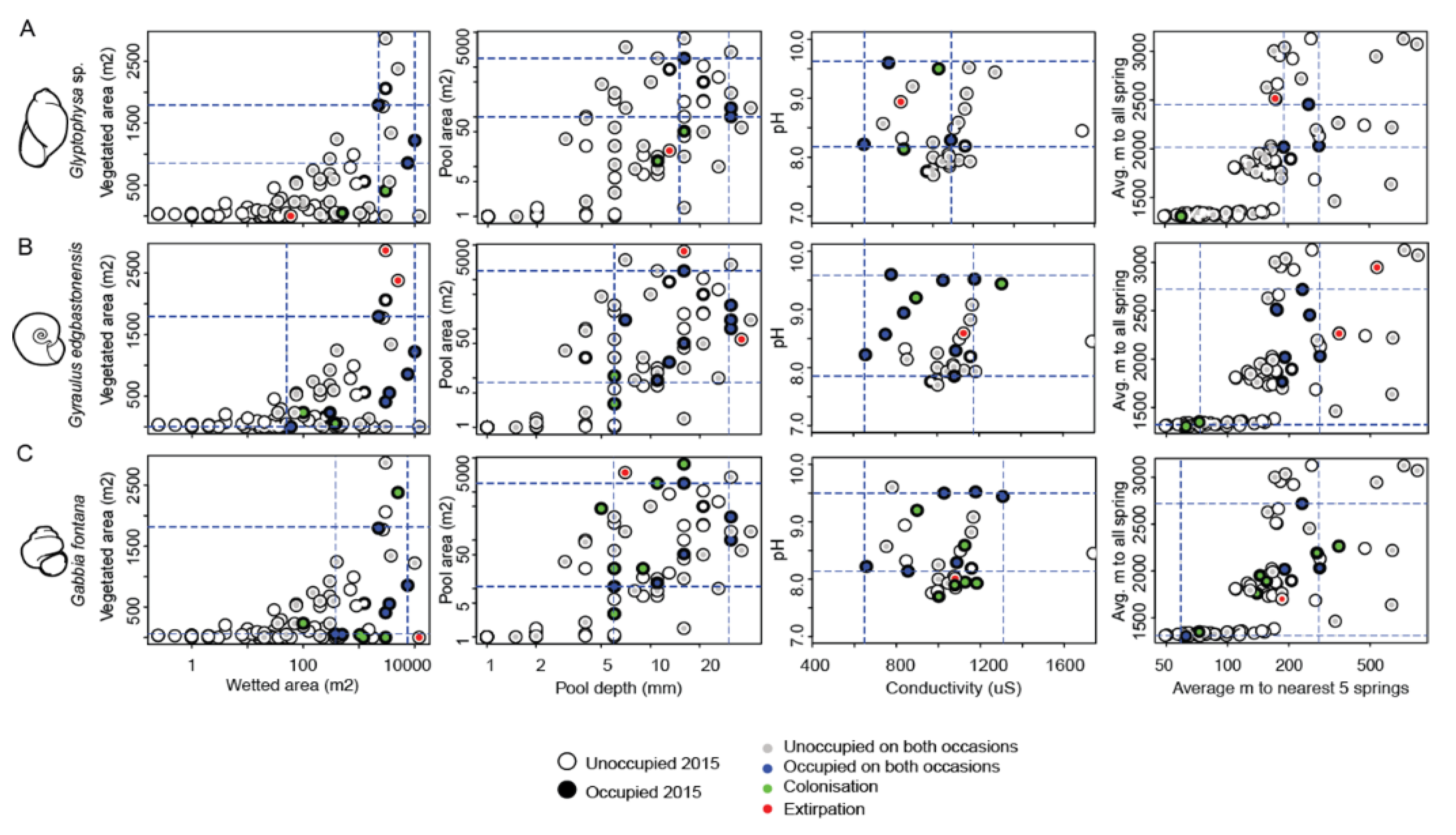
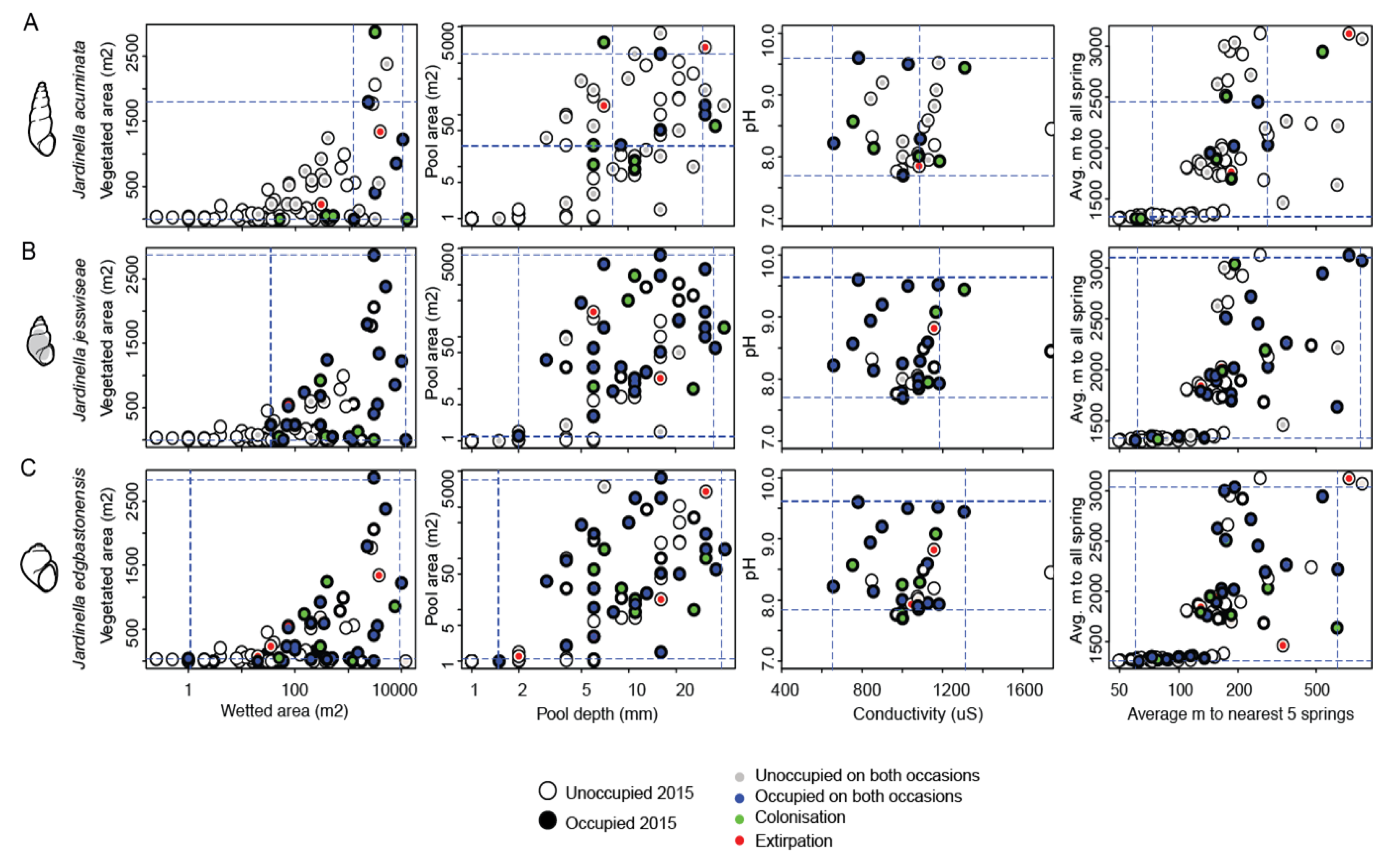
4.3. Relationship between Occupancy and Spring Characteristics
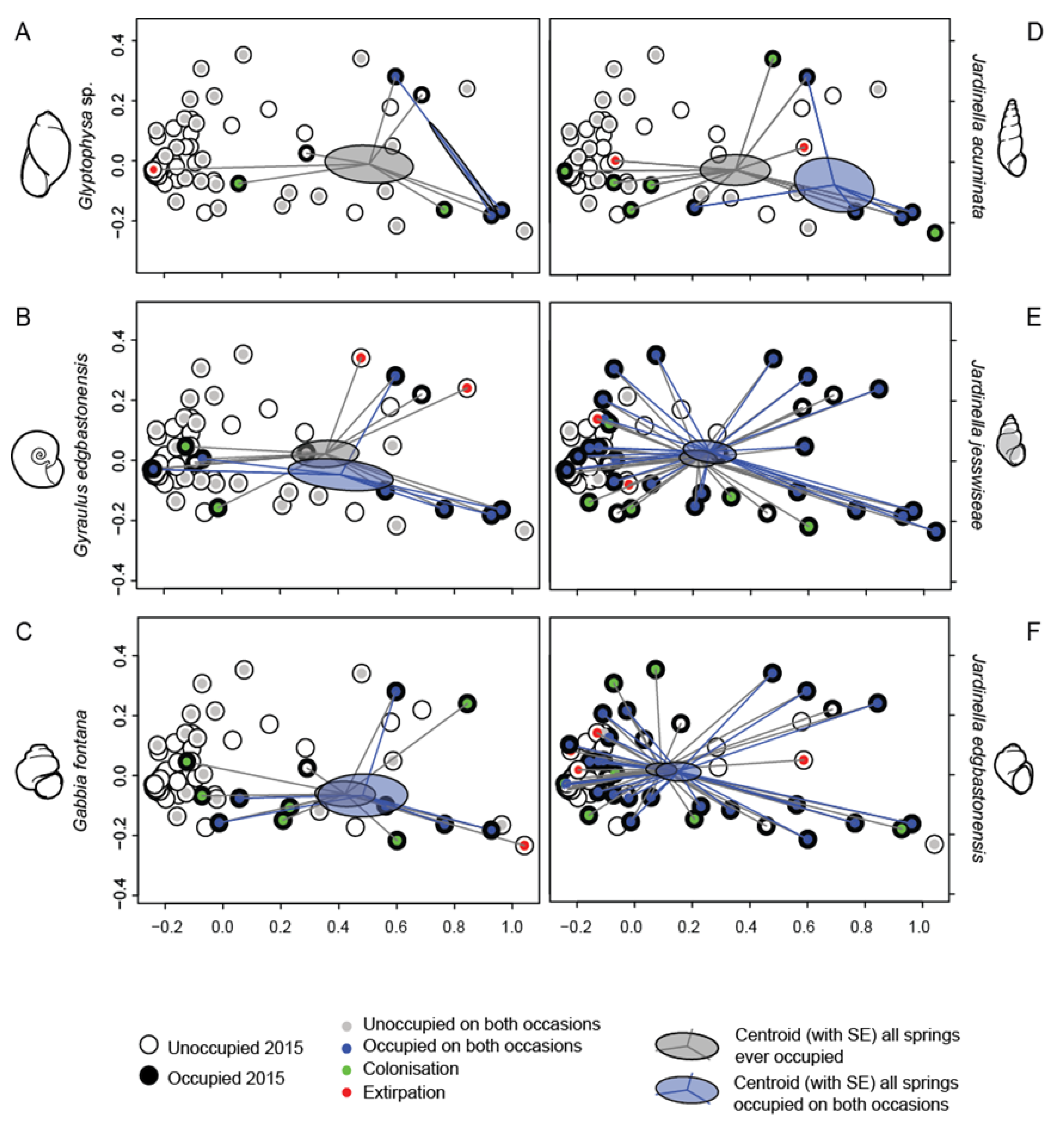
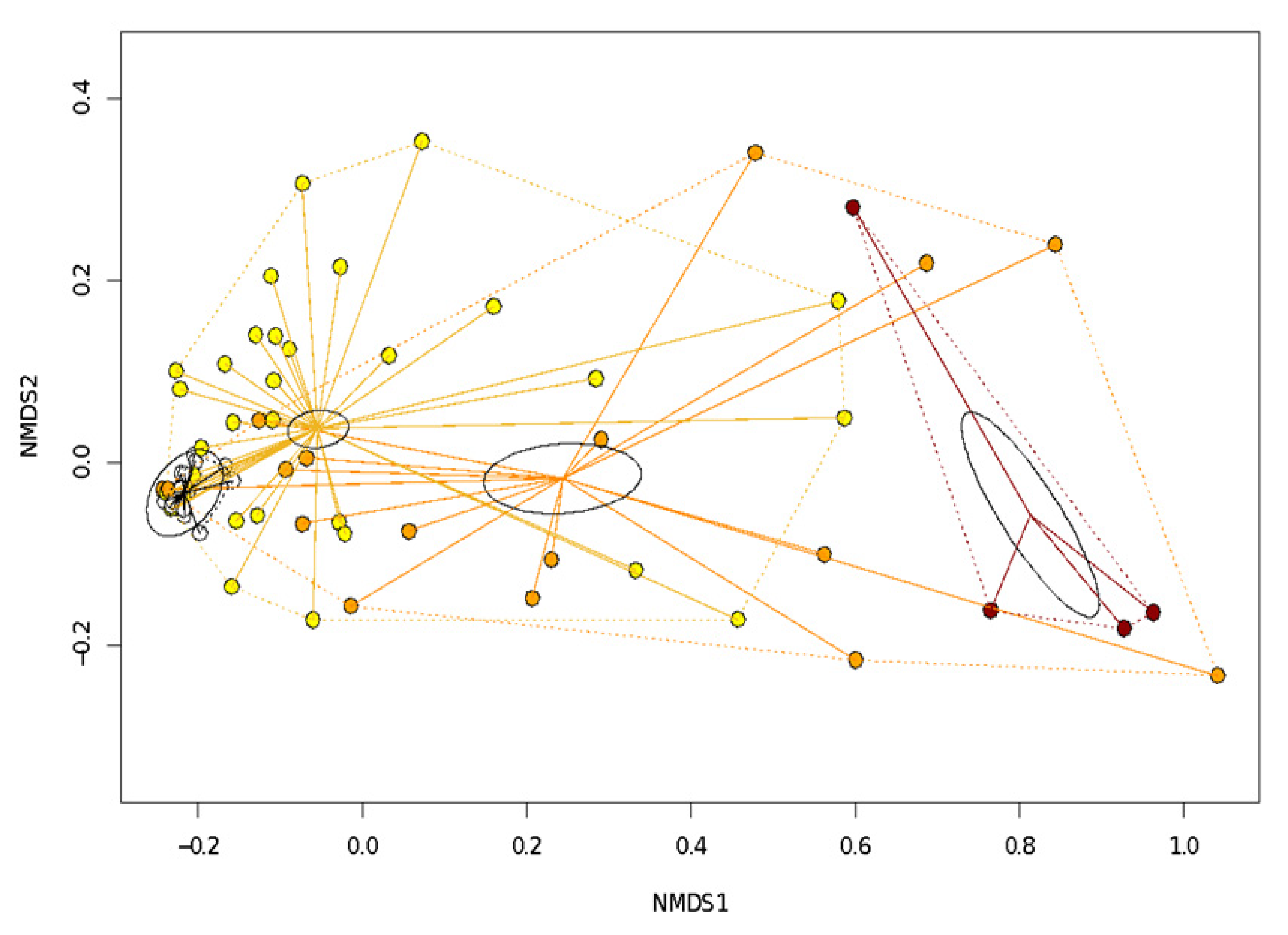
4.4. Relationship between Diversity and Spring Characteristics
5. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cantonati, M.; Füreder, L.; Gerecke, R.; Juttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Elsaied, A.; El-Ghamry, A.; Khafagi, O.-M.A.; Powell, O.; Bedair, R. Floristic diversity and vegetation analysis of Siwa Oasis: An ancient agro-ecosystem in Egypt’s Western Desert. Ann. Agric. Sci. 2015, 60, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Fairfax, R.; Fensham, R. In the Footsteps of J. Alfred Griffiths: A Cataclysmic History of Great Artesian Basin Springs in Queensland, Australia. Aust. Geogr. Stud. 2002, 40, 210–230. [Google Scholar] [CrossRef]
- Famiglietti, J.S. The global groundwater crisis. Nat. Clim. Chang. 2014, 4, 945–948. [Google Scholar] [CrossRef]
- Powell, O.; Fensham, R.J. The history and fate of the Nubian Sandstone Aquifer springs in the oasis depressions of the Western Desert, Egypt. Hydrogeol. J. 2015, 24, 395–406. [Google Scholar] [CrossRef]
- Unmack, P.; Minckley, W.L. The Demise of Desert Springs. In Aridland Springs in North America: Ecology and Conservation; Stevens, L.E., Meretsky, V.J., Eds.; The University of Arizona Press: Tucson, AZ, USA, 2008. [Google Scholar]
- Ponder, W.F.; Walker, K.F. From Mound Springs to Mighty Rivers: The conservation Status of Freshwater Molluscs in Australia. Aquat. Ecosyst. Health Manag. 2003, 6, 19–28. [Google Scholar] [CrossRef]
- Box, J.; Duguid, A.; Read, R.; Kimber, R.; Knapton, A.; Davis, J.; Bowland, A. Central Australian waterbodies: The importance of permanence in a desert landscape. J. Arid Environ. 2008, 72, 1395–1413. [Google Scholar] [CrossRef]
- Davis, J.A.; Kerezsy, A.; Nicol, S. Springs: Conserving perennial water is critical in arid landscapes. Biol. Conserv. 2017, 211, 30–35. [Google Scholar] [CrossRef]
- Shepard, W.D. Desert springs—Both rare and endangered. Aquat. Conserv. Mar. Freshw. Ecosyst. 1993, 3, 351–359. [Google Scholar] [CrossRef]
- Ponder, W.F.; Deckker, P. Mound Springs of the Great Artesian Basin. In Limnology in Australia; Springer: Berlin, Germany, 1986. [Google Scholar]
- Brown, K.M.; Lang, B.; Perez, K.E. The conservation ecology of North American pleurocerid and hydrobiid gastropods. J. N. Am. Benthol. Soc. 2008, 27, 484–495. [Google Scholar] [CrossRef]
- Fensham, R.J.; Silcock, J.L.; Kerezsy, A.; Ponder, W. Four desert waters: Setting arid zone wetland conservation priorities through understanding patterns of endemism. Boil. Conserv. 2011, 144, 2459–2467. [Google Scholar] [CrossRef]
- Myers, M.J.; Resh, V.H. Spring-formed wetlands of the arid west—Islands of aquatic invertebrate biodiversity. In Invertebrates in Freshwater Wetlands of North America; John Wiley & Sons: Hoboken, NJ, USA, 1999; pp. 811–828. [Google Scholar]
- Powell, O.; Silcock, J.; Fensham, R.J. Oases to Oblivion: The Rapid Demise of Springs in the South-Eastern Great Artesian Basin, Australia. Ground Water 2013, 53, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Richey, A.S.; Thomas, B.F.; Lo, M.-H.; Reager, J.T.; Famiglietti, J.S.; Voss, K.; Swenson, S.; Rodell, M. Quantifying renewable groundwater stress with GRACE. Water Resour. Res. 2015, 51, 5217–5238. [Google Scholar] [CrossRef]
- Fensham, R.J.; Ponder, W.F.; Fairfax, R. Recovery Plan for the Community of Native Species Dependant on Natural Discharge of Groundwater from the Great Artesian Basin; Queensland Department of Environment and Resource Management: Brisbane, Australia, 2010. [Google Scholar]
- Ponder, W.F.; Vial, M.; Jefferys, E. The Aquatic Macroinvertebrates in the Springs on Edgbaston Station; Report for Bush Heritage Australia; The Queensland Museum: Brisbane, Australia, 2010. [Google Scholar]
- Worthington-Wilmer, J.W.; Elkin, C.M.; Wilcox, C.; Murray, L.; Niejalke, D.; Possingham, H. The influence of multiple dispersal mechanisms and landscape structure on population clustering and connectivity in fragmented artesian spring snail populations. Mol. Ecol. 2008, 17, 3733–3751. [Google Scholar] [CrossRef]
- Rossini, R.; Fensham, R.J.; Walter, G.H. Spatiotemporal variance of environmental conditions in Australian artesian springs affects the distribution and abundance of six endemic snail species. Aquat. Ecol. 2017, 51, 511–529. [Google Scholar] [CrossRef]
- Incagnone, G.; Marrone, F.; Barone, R.; Robba, L.; Naselli-Flores, L. How do freshwater organisms cross the “dry ocean”? A review on passive dispersal and colonization processes with a special focus on temporary ponds. Hydrobiologia 2014, 750, 103–123. [Google Scholar] [CrossRef]
- Worthington-Wilmer, J.W.; Wilcox, C. Fine scale patterns of migration and gene flow in the endangered mound spring snail, Fonscochlea accepta (Mollusca: Hydrobiidae) in arid Australia. Conserv. Genet. 2006, 8, 617–628. [Google Scholar] [CrossRef]
- Ponder, W.F.; Colgan, D.J. What makes a narrow-range taxon? Insights from Australian freshwater snails. Invertebr. Syst. 2002, 16, 571–582. [Google Scholar] [CrossRef]
- Bilton, D.T.; Freeland, J.R.; Okamura, B. Dispersal in Freshwater Invertebrates. Annu. Rev. Ecol. Syst. 2001, 32, 159–181. [Google Scholar] [CrossRef] [Green Version]
- Morán-Ordóñez, A.; Pavlova, A.; Pinder, A.M.; Sim, L.; Sunnucks, P.; Thompson, R.M.; Davis, J. Aquatic communities in arid landscapes: Local conditions, dispersal traits and landscape configuration determine local biodiversity. Divers. Distrib. 2015, 21, 1230–1241. [Google Scholar] [CrossRef]
- Ponder, W.F.; Hershler, R.; Jenkins, B. An endemic radiation of Hydrobiid snails from Artesian Springs in Northern South Australia—Their taxonomy, physiology, distribution and anatomy. Malacologia 1989, 31, 1–140. [Google Scholar]
- Rossini, R.; Tibbetts, H.L.; Fensham, R.J.; Walter, G.H. Can environmental tolerances explain convergent patterns of distribution in endemic spring snails from opposite sides of the Australian arid zone? Aquat. Ecol. 2017, 51, 605–624. [Google Scholar] [CrossRef]
- Murphy, N.P.; Guzik, M.T.; Wilmer, J.W. The influence of landscape on population structure of four invertebrates in groundwater springs. Freshw. Biol. 2010, 55, 2499–2509. [Google Scholar] [CrossRef]
- Worthington-Wilmer, J.W.; Murray, L.; Elkin, C.M.; Wilcox, C.; Niejalke, D.; Possingham, H. Catastrophic Floods May Pave the Way for Increased Genetic Diversity in Endemic Artesian Spring Snail Populations. PLoS ONE 2011, 6, e28645. [Google Scholar] [CrossRef] [PubMed]
- Cantonati, M.; Rott, E.; Spitale, D.; Angeli, N.; Komárek, J. Are benthic algae related to spring types? Freshw. Sci. 2012, 31, 481–498. [Google Scholar] [CrossRef]
- Nicol, S.; Haynes, T.B.; Fensham, R.J.; Kerezsy, A. Quantifying the impact of Gambusia holbrooki on the extinction risk of the critically endangered red-finned blue-eye. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.L.; Guzman, L.M.; De Meester, L.; Horváth, Z.; Ptacnik, R.; Vanschoenwinkel, B.; Viana, D.; Chase, J. A process-based metacommunity framework linking local and regional scale community ecology. Ecol. Lett. 2020, 23, 1314–1329. [Google Scholar] [CrossRef]
- Hanski, I. Coexistence of Competitors in Patchy Environment with and without Predation. Oikos 1981, 37, 306. [Google Scholar] [CrossRef]
- Hanski, I. Coexistence of Competitors in Patchy Environment. Ecology 1983, 64, 493–500. [Google Scholar] [CrossRef]
- Hanski, I. A Practical Model of Metapopulation Dynamics. J. Anim. Ecol. 1994, 63, 151. [Google Scholar] [CrossRef] [Green Version]
- Hanski, I. Predictive and Practical Metapopulation Models: The Incidence Function Approach. In Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1997; Volume 30, pp. 21–45. [Google Scholar]
- Hanski, I. Metapopulation theory, its use and misuse. Basic Appl. Ecol. 2004, 5, 225–229. [Google Scholar] [CrossRef]
- Hanski, I.; Gilpin, M. Metapopulation dynamics: Brief history and conceptual domain. Biol. J. Linn. Soc. 1991, 42, 3–16. [Google Scholar] [CrossRef]
- Hanski, I.; Thomas, C.D. Metapopulation dynamics and conservation: A spatially explicit model applied to butterflies. Biol. Conserv. 1994, 68, 167–180. [Google Scholar] [CrossRef]
- Dennis, R.L.; Eales, H.T. Patch occupancy in Coenonympha tullia (Muller, 1764) (Lepidoptera: Satyrinae): Habitat quality matters as much as patch size and isolation. J. Insect Conserv. 1997, 1, 167–176. [Google Scholar] [CrossRef]
- Fleishman, E.; Murphy, D.D.; Sada, D.W. Effects of Environmental Heterogeneity and Disturbance on the Native and Non-native Flora of Desert Springs. Biol. Invasions 2006, 8, 1091–1101. [Google Scholar] [CrossRef]
- Fleishman, E.; Ray, C.; Sjogren-Gulve, P.; Boggs, C.L.; Murphy, D.D. Assessing the Roles of Patch Quality, Area, and Isolation in Predicting Metapopulation Dynamics. Conserv. Biol. 2002, 16, 706–716. [Google Scholar] [CrossRef] [Green Version]
- Mortelliti, A.; Amori, G.; Boitani, L. The role of habitat quality in fragmented landscapes: A conceptual overview and prospectus for future research. Oecologia 2010, 163, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Mortelliti, A.; Sozio, G.; Driscoll, D.A.; Bani, L.; Boitani, L.; Lindenmayer, D. Population and individual-scale responses to patch size, isolation and quality in the hazel dormouse. Ecosphere 2014, 5, art107. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, T.; Tanaka, K.; Hamasaki, K.; Nakatani, Y.; Iwasaki, N.; Sprague, D.S.; Bjørnstad, O.N. Evaluating the relative importance of patch quality and connectivity in a damselfly metapopulation from a one-season survey. Oikos 2009, 118, 67–76. [Google Scholar] [CrossRef]
- Walter, G.H.; Hengeveld, R. Autecology: Organisms, Interactions and Environmental Dynamics; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Cantonati, M.; Angeli, N.; Bertuzzi, E.; Spitale, D.; Lange-Bertalot, H. Diatoms in springs of the Alps: Spring types, environmental determinants, and substratum. Freshw. Sci. 2012, 31, 499–524. [Google Scholar] [CrossRef]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Glazier, D. The fauna of North American temperate cold springs: Patterns and hypotheses. Freshw. Biol. 1991, 26, 527–542. [Google Scholar] [CrossRef]
- Rosati, M.; Cantonati, M.; Primicerio, R.; Rossetti, G. Biogeography and relevant ecological drivers in spring habitats: A review on ostracods of the Western Palearctic. Int. Rev. Hydrobiol. 2014, 99, 409–424. [Google Scholar] [CrossRef]
- Spitale, D.; Leira, M.; Angeli, N.; Cantonati, M. Environmental classification of springs of the Italian Alps and its consistency across multiple taxonomic groups. Freshw. Sci. 2012, 31, 563–574. [Google Scholar] [CrossRef]
- Horsák, M.; Rádková, V.; Syrovátka, V.; Bojková, J.; Křoupalová, V.; Schenkova, J.; Zajacová, J. Drivers of aquatic macroinvertebrate richness in spring fens in relation to habitat specialization and dispersal mode. J. Biogeogr. 2015, 42, 2112–2121. [Google Scholar] [CrossRef]
- Shand, P.; Gotch, T.; Love, A.; Raven, M.; Priestley, S.; Grocke, S. Extreme environments in the critical zone: Linking acidification hazard of acid sulfate soils in mound spring discharge zones to groundwater evolution and mantle degassing. Sci. Total Environ. 2016, 568, 1238–1252. [Google Scholar] [CrossRef]
- Kodric-Brown, A.; Brown, J.H. Highly Structured Fish Communities in Australian Desert Springs. Ecology 1993, 74, 1847–1855. [Google Scholar] [CrossRef] [Green Version]
- White, D.C.; Lewis, M. A new approach to monitoring spatial distribution and dynamics of wetlands and associated flows of Australian Great Artesian Basin springs using QuickBird satellite imagery. J. Hydrol. 2011, 408, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Fensham, R.J.; Fairfax, R.; Pocknee, D.; Kelley, J. Vegetation patterns in permanent spring wetlands in arid Australia. Aust. J. Bot. 2004, 52, 719. [Google Scholar] [CrossRef] [Green Version]
- Zullini, A.; Gatti, F.; Ambrosini, R. Microhabitat preferences in springs, as shown by a survey of nematode communities of Trentino (south-eastern Alps, Italy). J. Limnol. 2011, 70, 93–105. [Google Scholar] [CrossRef]
- Klockmann, M.; Scharre, M.; Haase, M.; Fischer, K. Does narrow niche space in a ‘cold-stenothermic’ spring snail indicate high vulnerability to environmental change? Hydrobiology 2015, 765, 71–83. [Google Scholar] [CrossRef]
- Springer, A.E.; Stevens, L.E. Spheres of discharge of springs. Hydrogeol. J. 2008, 17, 83–93. [Google Scholar] [CrossRef]
- Department of Science Information Technology and Innovation Australia (DSITIA). Lake Eyre Basin Springs Assessment Project: Groundwater Dependent Ecosystem Mapping Report; Department of Science Information Technology and Innovation Australia: Brisbane, Australia, 2015. [Google Scholar]
- Ponder, W.F.; Hallan, A.; Shea, M.; Clark, S.A. Australian Freshwater Molluscs; Australian Museum: Brisbane, Australia, 2016; Available online: http://keys.lucidcentral.org/keys/v3/freshwater_molluscs/ (accessed on 20 October 2020).
- Ponder, W.F. Monograph of the Australian Bithyniidae (Caenogastropoda: Rissooidea). Zootaxa 2003, 230, 1. [Google Scholar] [CrossRef]
- Ponder, W.F.; Wilke, T.; Zhang, W.H.; Golding, R.E.; Fukuda, H.; Mason, R.A.B. Edgbastonia alanwillsi n. gen & n. sp (Tateinae: Hydrobiidae s.l.: Rissooidea: Caenogastropoda); a snail from an artesian spring group in western Queensland, Australia, convergent with some Asian Amnicolidae. Molluscan Res. 2008, 28, 89–106. [Google Scholar]
- Ponder, W.F.; Clark, G.A. A Radiation of Hydrobiid Snails in Threatened Artesian Springs in Western Queensland. Rec. Aust. Mus. 1990, 42, 301–363. [Google Scholar] [CrossRef] [Green Version]
- Rossini, R.A.; Fensham, R.J.; Walter, G.H. Determining optimal sampling strategies for monitoring threatened endemic macro-invertebrates in Australia’s artesian springs. Mar. Freshw. Res. 2015, 67, 653–665. [Google Scholar] [CrossRef]
- Cunningham, R.B.; Lindenmayer, D.B. Modelling count data of rare species: Some statistical issues. Ecology 2005, 86, 1135–1142. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R.; Springer: Berlin, Germany, 2011. [Google Scholar]
- Legendre, P.; Legendre, L.F.J. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Suggests, M.A.S.S. The vegan package. In Community Ecology Package; 2007; Volume 10, pp. 631–637. [Google Scholar]
- Von Fumetti, S.; Blattner, L. Faunistic assemblages of natural springs in different areas in the Swiss National Park: A small-scale comparison. Hydrobiology 2016, 793, 175–184. [Google Scholar] [CrossRef]
- Pritchard, G. Insects in thermal springs. Memoirs Èntomol. Soc. Can. 1991, 123, 89–106. [Google Scholar] [CrossRef]
- Wiegert, R.G.; Mitchell, R. Ecology of yellowstone thermal effluent systems: Intersects of blue-green algae, grazing flies (Paracoenia, Ephydridae) and water mites (Partnuniella, Hydrachnellae). Hydrobiology 1973, 41, 251–271. [Google Scholar] [CrossRef]
- Fairfax, R.; Fensham, R.J.; Wager, R.; Brooks, S.; Webb, A.; Unmack, P.J. Recovery of the red-finned blue-eye: An endangered fish from springs of the Great Artesian Basin. Wildl. Res. 2007, 34, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Kodric-Brown, A.; Brown, J.H. Native fishes, exotic mammals, and the conservation of desert springs. Front. Ecol. Environ. 2007, 5, 549–553. [Google Scholar] [CrossRef]
- Kodric-Brown, A.; Wilcox, C.; Bragg, J.G.; Brown, J.H. Dynamics of fish in Australian desert springs: Role of large-mammal disturbance. Divers. Distrib. 2007, 13, 789–798. [Google Scholar] [CrossRef]
- Kovac, K.-J.; Mackay, D.A. An experimental study of the impacts of cattle on spider communities of artesian springs in South Australia. J. Insect Conserv. 2007, 13, 57–65. [Google Scholar] [CrossRef]
- Sada, D.W.; Fleishman, E.; Murphy, D.D. Associations among spring-dependent aquatic assemblages and environmental and land use gradients in a Mojave Desert mountain range. Divers. Distrib. 2005, 11, 91–99. [Google Scholar] [CrossRef]
- Schooley, R.L.; Branch, L.C. Enhancing the area-isolation paradigm: Habitat heterogeneity and metapopulation dynamics of a rare wetland mammal. Ecol. Appl. 2009, 19, 1708–1722. [Google Scholar] [CrossRef]
- Benke, M.; Brändle, M.; Albrecht, C.; Wilke, T. Patterns of freshwater biodiversity in Europe: Lessons from the spring snail genus Bythinella. J. Biogeogr. 2011, 38, 2021–2032. [Google Scholar] [CrossRef]
- Hershler, R.; Liu, H.-P.; Howard, J. Springsnails: A New Conservation Focus in Western North America. Bioscience 2014, 64, 693–700. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Lawler, J.J.; Poff, N.L. Challenges and Opportunities in Implementing Managed Relocation for Conservation of Freshwater Species. Conserv. Biol. 2010, 25, 40–47. [Google Scholar] [CrossRef]
- Sgro, C.M.; Lowe, A.J.; Hoffmann, A.A. Building evolutionary resilience for conserving biodiversity under climate change. Evol. Appl. 2010, 4, 326–337. [Google Scholar] [CrossRef]
- Peck, S. Evaluating the Effectiveness of Fencing to Manage Feral Animal Impacts on High Conservation Value Artesian Spring Wetland Communities of Currawinya National Park. Royal Soc. Queensland 2020, in press. [Google Scholar]
- Fairfax, R.J.; Fensham, R.J. Great Artesian Basin Springs in Southern Queensland 1911–2000. In Memoirs of the Queensland Museum; Queensland Museum: Brisbane, Australia, 2003; Volume 49, pp. 285–293. [Google Scholar]
- Kerezsy, A.; Fensham, R.J. Conservation of the endangered red-finned blue-eye, Scaturiginichthys vermeilipinnis, and control of alien eastern gambusia, Gambusia holbrooki, in a spring wetland complex. Mar. Freshw. Res. 2013, 64, 851. [Google Scholar] [CrossRef]
- Kerezsy, A. Fish Survey of the Edgbaston Spring Complex with an Emphasis on the Distribution of the Endangered Edgbaston Goby, Chlamydogobius squamigenus; Clontarf, Queensland, Australia, 2014. [Google Scholar]
- Faulks, L.K.; Kerezsy, A.; Unmack, P.J.; Johnson, J.B.; Hughes, J.M. Going, going, gone? Loss of genetic diversity in two critically endangered Australian freshwater fishes, Scaturiginichthys vermeilipinnisand Chlamydogobius squamigenus, from Great Artesian Basin springs at Edgbaston, Queensland, Australia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 27, 39–50. [Google Scholar] [CrossRef]
- Silcock, J.L.; Macdermott, H.; Laffineur, B.; Fensham, R.J. Obscure oases: Natural, cultural and historical geography of western Queensland’s Tertiary sandstone springs. Geogr. Res. 2016, 54, 187–202. [Google Scholar] [CrossRef]
- Cantonati, M.; Segadelli, S.; Ogata, K.; Tran, H.; Sanders, D.; Gerecke, R.; Rott, E.; Filippini, M.; Gargini, A.; Celico, F.; et al. A global review on ambient Limestone-Precipitating Springs (LPS): Hydrogeological setting, ecology, and conservation. Sci. Total Environ. 2016, 568, 624–637. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Snail Species | 2006–2008 | 2015 | Same | Extirpation | Colonisation |
|---|---|---|---|---|---|
| Gabbia fontana | 7 | 12 | 6 | 1 | 6 |
| Gyraulus edgbastonensis | 10 | 10 | 8 | 2 | 2 |
| Glyptophysa sp. | 4 | 5 | 3 | 1 | 2 |
| Jardinella acuminata | 7 | 11 | 5 | 2 | 6 |
| Jardinella jesswiseae | 25 | 28 | 23 | 2 | 5 |
| Jardinella edgbastonensis | 28 | 31 | 24 | 4 | 7 |
| Spring Attribute | Number of Species | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3–4 | >4 | |||
| WETTED AREA (m2) | Avg | 16 | 382 | 446 | 1340 | 2426 | p ≤ 0.05 |
| (±SE) | (±4) | (±221) | (±172) | (±494) | (±929) | ||
| TOTAL POOL AREA (m2) | Avg | 1 | 151 | 122 | 573 | 184 | ns |
| (±SE) | (±0) | (±88) | (±61) | (±319) | (±122) | ||
| MAXIMUM POOL DEPTH (mm) | Avg | 1.4 | 7.1 | 8.8 | 13.4 | 13.8 | p ≤ 0.05 |
| (±SE) | (±0.8) | (±2.1) | (±2.3) | (±2.7) | (±3.0) | ||
| PH | Avg | 8.1 | 8.6 | 8.2 | 8.2 | 8.7 | ns |
| (±SE) | NA | NA | (±0.1) | (±0.1) | (±0.2) | ||
| COND. (uS) | Avg | 875 | 802 | 1022 | 857 | 775 | ns |
| (±SE) | NA | NA | (±173) | (±29) | (±59) | ||
| AVERAGE DISTANCE TO ALL SPRINGS (m) | Avg | 63 | 111 | 124 | 150 | 77 | ns |
| (±SE) | (±10) | (±32) | (±45) | (±49) | (±12) | ||
| AVERAGE DISTANCE TO CLOSEST 5 SPRINGS (m) | Avg | 190 | 212 | 203 | 227 | 152 | ns |
| (±SE) | (±53) | (±60) | (±46) | (±42) | (±22) | ||
| # OF SPRINGS WITHIN 300M | Avg | 14.3 | 9.9 | 7.5 | 7.7 | 12.1 | ns |
| (±SE) | (±2.6) | (±2.2) | (±2.0) | (±2.2) | (±2.8) | ||
| # SPRINGS CONNECTED BY WETLAND | Avg | 0.1 | 0.2 | 0.3 | 1 | 0.8 | p ≤ 0.05 |
| (±SE) | (±0.1) | (±0.1) | (±0.1) | (±0.4) | (±0.3) | ||
| Species | Number of Springs Occupied | Spring Size (m2) | Pool Size (m2) | Pool Depth (cm) | Total Wetland Area of OCCUPIED Springs |
|---|---|---|---|---|---|
| (ha) | |||||
| Glyptophysa sp. | 5 | >1000 | >10 | >10 | 2.8 |
| Gy. edgbastonensis | 10 | >100 | >5 | >5 | 3.2 |
| J. acuminata | 11 | >1000 | >5 | >5 | 4.0 |
| J. jesswiseae | 25 | >10 | >0 | >1 | 6.9 |
| J. edgbastonensis | 31 | >1 | ≥0 | 0 | 5.2 |
| Ga. fontana | 12 | >100 | >5 | >5 | 2.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. Rossini, R.; J. Fensham, R.; H. Walter, G. Different Species Requirements within a Heterogeneous Spring Complex Affects Patch Occupancy of Threatened Snails in Australian Desert Springs. Water 2020, 12, 2942. https://doi.org/10.3390/w12102942
A. Rossini R, J. Fensham R, H. Walter G. Different Species Requirements within a Heterogeneous Spring Complex Affects Patch Occupancy of Threatened Snails in Australian Desert Springs. Water. 2020; 12(10):2942. https://doi.org/10.3390/w12102942
Chicago/Turabian StyleA. Rossini, Renee, Roderick J. Fensham, and Gimme H. Walter. 2020. "Different Species Requirements within a Heterogeneous Spring Complex Affects Patch Occupancy of Threatened Snails in Australian Desert Springs" Water 12, no. 10: 2942. https://doi.org/10.3390/w12102942