Agro-Morphological, Microanatomical and Molecular Cytogenetic Characterization of the Medicinal Plant Chelidonium majus L.

 , , and
, , and
Abstract
:1. Introduction
2. Results
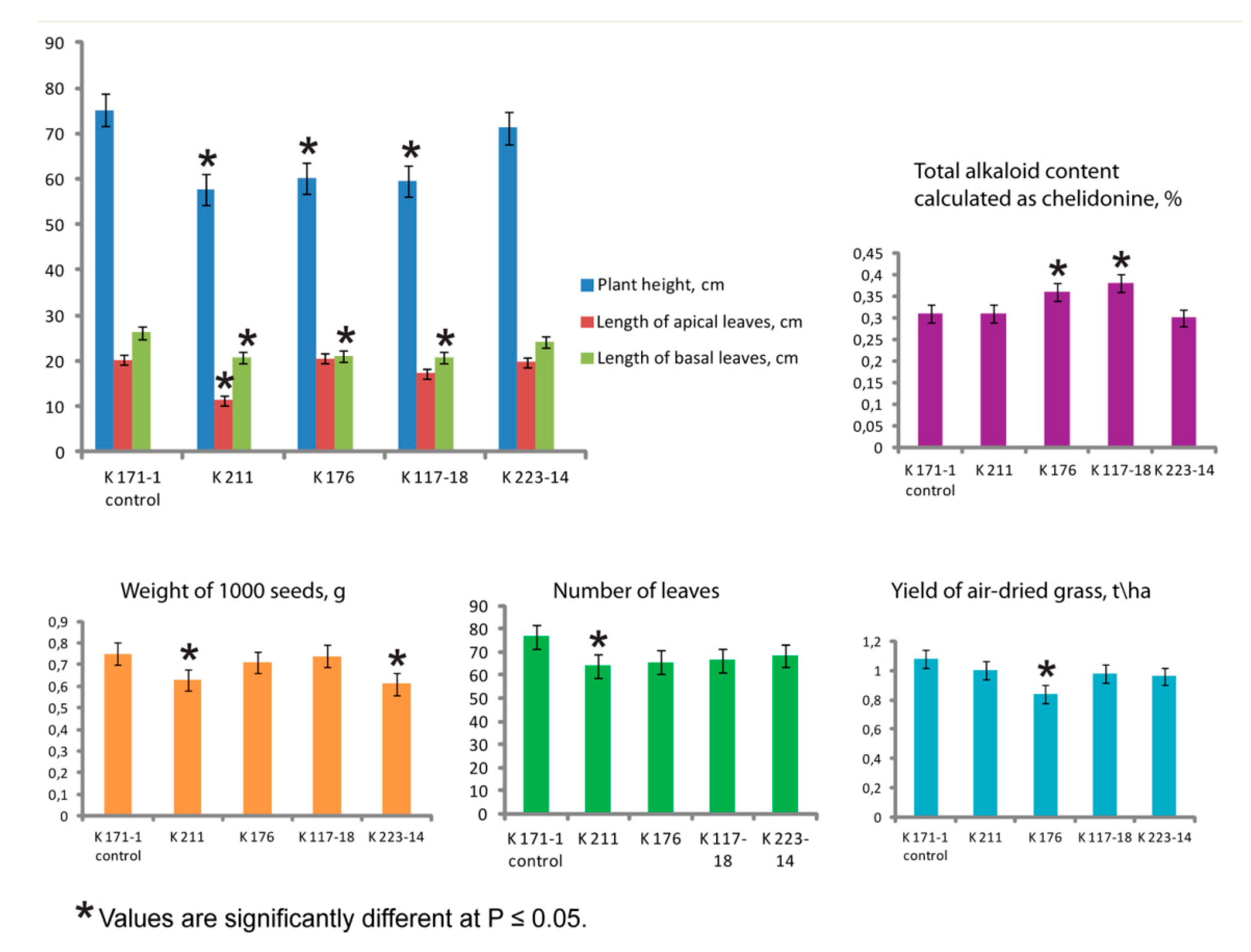
2.1. Agro-Morphological Peculiarities
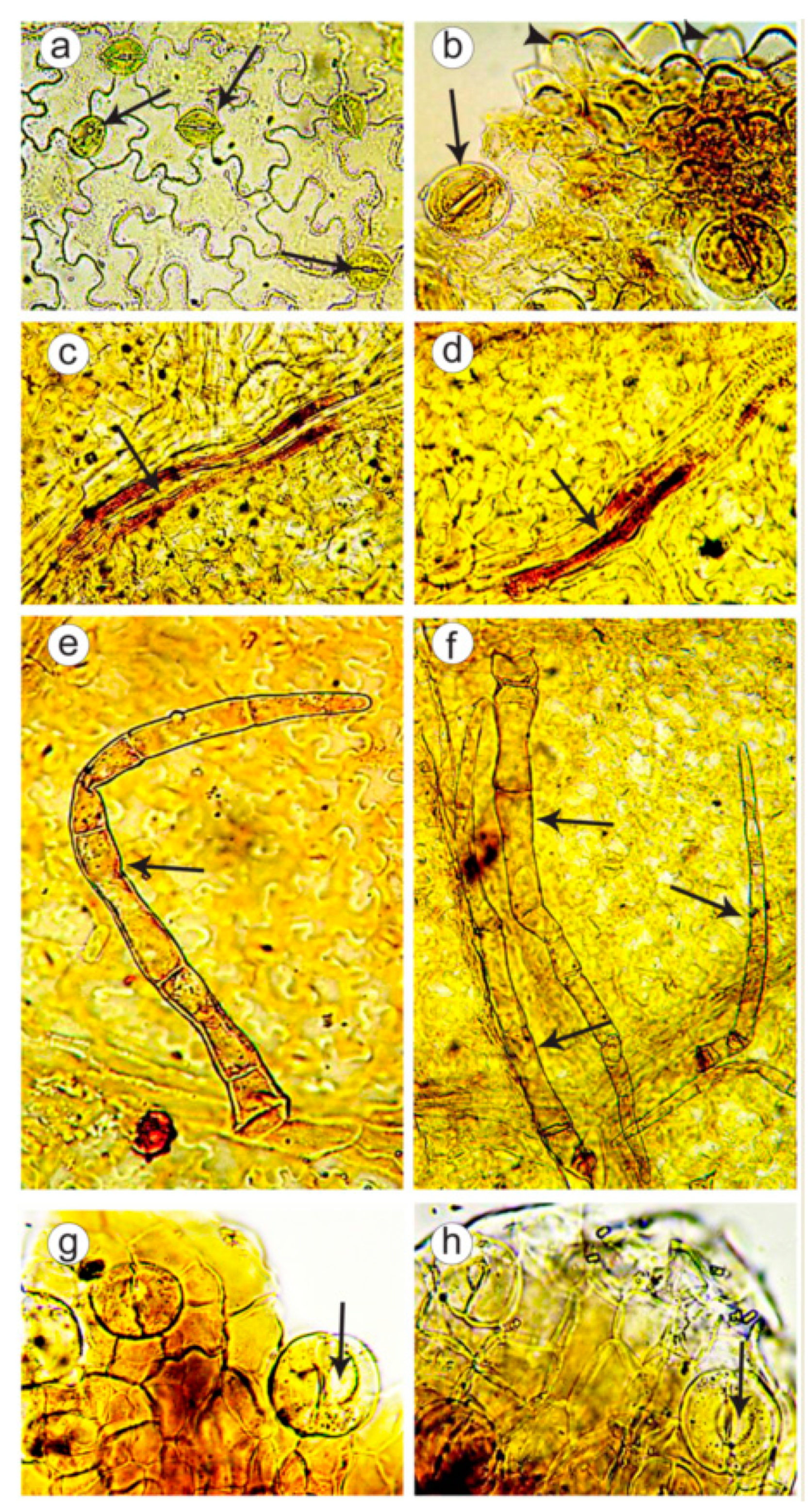
2.2. Microanatomical Analysis of Leaves and Flowers
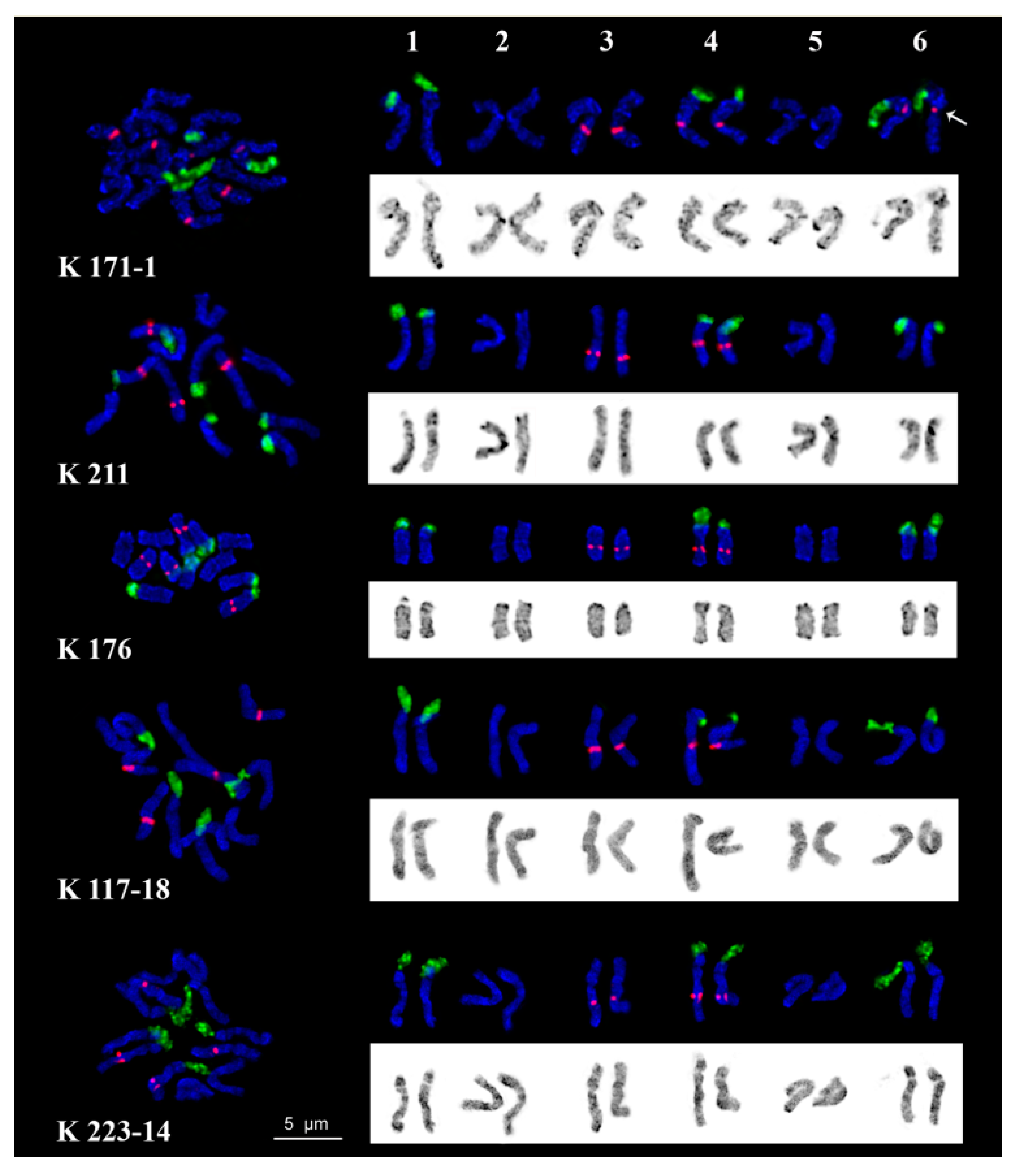
2.3. Chromosome Structure
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Bio-Morphological Characterization
4.3. Chemical Composition of the Grass
4.4. Chromosome Spread Preparation
4.5. FISH Procedure
4.6. DAPI Counterstaining
4.7. Analysis of Chromosomes
4.8. Microanatomical Analysis of Leaves and Flowers
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lai, P.K.; Roy, J. Antimicrobial and chemopreventive properties of herbs and spices. Curr. Med. Chem. 2004, 11, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K. Modern plant metabolomics: Advanced natural product gene discoveries, improved technologies, and future prospects. Nat. Prod. Rep. 2015, 32, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, S.; Jezierska-Domaradzka, A.; Wójciak-Kosior, M.; Sowa, I.; Junka, A.; Matkowski, A.M. Greater celandine’s ups and downs—21 centuries of medicinal uses of Chelidonium majus from the viewpoint of today’s pharmacology. Front. Pharmacol. 2018, 9, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilca, M.; Gaman, L.; Panait, E.; Stoian, I.; Atanasiu, V. Chelidonium majus—An Integrative Review: Traditional Knowledge versus Modern Findings. Forsch. Komplement. 2010, 17, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Kopytko, Y.F.; Dargaeva, T.D.; Sokol’slaya, T.A.; Grodnitskaya, E.I.; Kopnin, A.A. New methods for the quality control of a homeopathic matrix tincture of Greater celandine. Pharm. Chem. J. 2005, 39, 603–609. [Google Scholar] [CrossRef]
- Miao, F.; Yang, X.J.; Zhou, L.; Hu, H.J.; Zheng, F.; Ding, X.D.; Sun, D.M.; Zhou, C.D.; Sun, W. Structural modification of sanguinarine and chelerythrine and their antibacterial activity. Nat. Prod. Res. 2011, 25, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Cuong, T.D.; Hung, T.M.; Lee, I.; Na, M.; Kim, J.C.; Ryoo, S.; Lee, J.H.; Choi, J.S.; Woo, M.H.; et al. Alkaloids from Chelidonium majus and their inhibitory effects on LPS induced NO production in RAW264.7 cells. Bioorg. Med. Chem. Lett. 2011, 21, 6960–6963. [Google Scholar] [CrossRef]
- Monavari, S.H.; Shahrabadi, M.S.; Keyvani, H.; Bokharaei-Salim, F. Evaluation of in vitro antiviral activity of Chelidonium majus L. against herpes simplex virus Type-1. Afr. J. Microbiol. Res. 2012, 6, 4360–4364. [Google Scholar] [CrossRef]
- Hou, Z.; Yang, R.; Zhang, C.; Zhu, L.F.; Miao, F.; Yang, X.J.; Zhou, L. 2(substituted phenyl)-3,4-dihydroisoquinolin2-iums as novel antifungal lead compounds: Biological evaluation and structure-activity relationships. Molecules 2013, 18, 10413–10424. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Kim, S.J.; Kim, M.C.; Jeon, Y.D.; Um, J.Y.; Hong, S.H. The therapeutic effect of chelidonic acid on ulcerative colitis. Biol. Pharm. Bull. 2012, 35, 666–671. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.J.; Bhattacharjee, N.; Khuda Bukhsh, A.R. Efficacy of a plant extract (Chelidonium majus L.) in combating induced hepatocarcinogenesis in mice. Food Chem. Toxicol. 2008, 46, 1474–1487. [Google Scholar] [CrossRef] [PubMed]
- Koriem, K.M.; Arbid, M.S.; Asaad, G.F. Chelidonium majus leaves methanol extract and its chelidonine alkaloid ingredient reduce cadmium-induced nephrotoxicity in rats. J. Nat. Med. 2013, 67, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Kaminskyy, V.O.; Lootsik, M.D.; Stoika, R.S. Correlation of the cytotoxic activity of four different alkaloids, from Chelidonium majus (greater celandine), with their DNA intercalating properties and ability to induce breaks in the DNA of NK/Ly murine lymphoma cells. Cent. Eur. J. Biol. 2006, 1, 2–15. [Google Scholar] [CrossRef]
- Philchenkov, A.; Kaminskyy, V.; Zavelevich, M.; Stoika, R. Apoptogenic activity of two benzophenanthridine alkaloids from Chelidonium majus L. does not correlate with their DNA damaging effects. Toxic. Vitro 2010, 22, 287–295. [Google Scholar] [CrossRef]
- Aljuraisy, Y.H.; Mahdi, N.K.; Al-Darraji, M.N.J. Cytotoxic effect of Chelidonium majus on cancer cell. Al-Anbar J. Veter. Sci. 2012, 5, 85–90. [Google Scholar]
- Amal, K.M.; Banerji, P. Chelidonium majus L. (Greater celandine)—A Review on its phytochemical and therapeutic perspectives. Int. J. Her. Med. 2015, 3, 10–27. [Google Scholar] [CrossRef]
- Colombo, M.L.; Bugatti, C.; Mossa, A.; Pescalli, N.; Piazzoni, L.; Pezzoni, G.; Menta, E.; Spinelli, S.; Johnson, F.; Gupta, R.C.; et al. Cytotoxicity evaluation of natural coptisine and synthesis of coptisine from berberine. Farmaco 2001, 56, 403–409. [Google Scholar] [CrossRef]
- Gansauge, F.; Ramadani, M.; Pressmar, J.; Gansauge, S.; Muehling, B.; Stecker, K.; Cammerer, G.; Leder, G.; Beger, H.G. NSC-631570 (Ukrain) in the palliative treatment of pancreatic cancer. Results of a phase II trial. Langenbeck’s Arch. Surg. 2002, 386, 570–574. [Google Scholar] [CrossRef]
- Ernst, E.; Schmidt, K. Ukrain—A new cancer cure? A systematic review of randomised clinical trials. BCM Cancer 2005, 5, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Pourmazaheri, H.; Soorni, A.; Kohnerouz, B.B.; Dehaghi, N.K.; Kalantar, E.; Omidi, M.; Naghavi, M.R. Comparative analysis of the root and leaf transcriptomes in Chelidonium majus L. PLoS ONE 2019, 14, e0215165. [Google Scholar] [CrossRef]
- Krahulcová, A. Cytotaxonomic study of Chelidonium majus L. Folia Geobot. Phytotax. 1982, 17, 238–270. [Google Scholar] [CrossRef]
- Chepinoga, V.V.; Gnutikov, A.A.; Lubogoschinsky, P.I. Chromosome numbers of some vascular plant species from the South Baikal Siberia. Bot. Pacif. 2012, 1, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Seledets, V.P.; Probatova, N.S. Ecological differentiation in Chelidonium (Papaveraceae). Sch. Not. Transbaikal. St. Univer. Ser. Biol. 2018, 1, 41–49. [Google Scholar] [CrossRef]
- Narae, Y.U.N.; Sang-Hun, O.H. Phylogenetic relationships of Coreanomecon (Papaveraceae: Papaveroideae), an endemic genus in Korea, using DNA sequences. Korean J. Pl. Taxon. 2018, 48, 289–300. [Google Scholar] [CrossRef]
- Wu, J.; He, J.; Peng, L.; Wang, A.; Zhao, L. The complete chloroplast genome sequence of Chelidonium majus (Papaveraceae). Mitoch. DNA 2019, 4, 1206–1207. [Google Scholar] [CrossRef] [Green Version]
- Sobirova, I.S. Variability of the composition of celandine alkaloids from different growing places. Pharmacy 1991, 5, 37–40. [Google Scholar]
- Colombo, M.L.; Bosissio, E. Pharmacological activities of Chelidonium majus L. (Papaveraceae). Pharm. Res. 1996, 33, 127–134. [Google Scholar] [CrossRef]
- Kashin, A.S.; Mashurchak, M.V.; Mashurchak, N.V. Morphometric features, productivity and age composition of Chelidonium majus (Papaveraceae) populations. Rast. Res. 2006, 42, 36–42. [Google Scholar]
- Baeva, V.M. Polymorphism of medicinal plants. Pharmacy 2005, 5, 40–42. [Google Scholar]
- Pavel, A.; Butnaru, M. Karyotype and idiogram in Chelidonium majus L. J. Exp. Mol. Biol. 2003, 4, 102–107. [Google Scholar]
- Vorniceanu, C.; Bara, I.I.; Morarıu, A.; Costica, N. The study of mitotic chromosomes at the Glaucium flavum Cr. (2n = 12) and Chelidonium majus L. (2n = 12) species. An. Stiintifice Univ. Alexandru Ioan Cuza Iasi Sec. II a. Genet. Biol. Mol. 2004, 5, 1. [Google Scholar]
- Kaplan, D.R. The science of plant morphology: Definition, history, and role in modern biology. Am. J. Bot. 2001, 88, 1711–1741. [Google Scholar] [CrossRef] [PubMed]
- Lotova, L.I. Botany: Morphology and Anatomy of Higher Plants; URSS Editorial: Moscow, Russia, 2001. [Google Scholar]
- Khazieva, F.M.; Morozov, A.I. Results and peculiarities of the breeding of medicinal plants in Russian Research Institute of Medicinal and Aromatic Plants. Pomic. Small Fruits Cult. Russ. 2019, 58, 265–272. [Google Scholar]
- Anderson, N.O. Flower Breeding and Genetics; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Samatadze, T.E.; Zoshchuk, S.A.; Hazieva, F.M.; Yurkevich, O.Y.; Svistunova, N.Y.; Morozov, A.I.; Amosova, A.V.; Muravenko, O.V. Phenotypic and molecular cytogenetic variability in calendula (Calendula officinalis L.) cultivars and mutant lines obtained via chemical mutagenesis. Sci. Rep. 2019, 9, 9155. [Google Scholar] [CrossRef] [Green Version]
- Maisuradze, N.I.; Kiselev, V.P.; Cherkasov, O.A. Methods for the Introduction of Medicinal Plants; Medprom: Moscow, Russia, 1984. [Google Scholar]
- Saposhnikov, S.A. Agro-Climatic Guide to the Moscow Region; Hydrometeoizdat: Saint Petersburg, Russia, 1967. [Google Scholar]
- Zarzycki, K.; Trzcińska-Tacik, H.; Różański, W.; Szeląg, Z.; Wołek, J.; Korzeniak, U. Ecological Indicator Values of Vascular Plants of Poland; Polish Academy of Sciences: Krakow, Poland, 2002. [Google Scholar]
- Tomē, F.; Colombo, M.L. Distribution of alkaloids in Chelidonium majus and factors affecting their accumulation. Phytochemistry 1995, 40, 37–39. [Google Scholar] [CrossRef]
- Seidler-Łożykowska, K.; Kędzia, B.; Bocianowski, J.; Gryszczyńska, A.; Łowicki, Z.; Opala, B.; Pietrowiak, A. Content of alkaloids and flavonoids in celandine (Chelidonium majus L.) herb at the selected developmental phases. Acta. Sci. Pol. Hort. Cult. 2016, 15, 161–172. [Google Scholar]
- Ministry of Healthcare of the Russian Federation. State Pharmacopoeia of the Russian Federation, 14th ed.; SCAMP: Moscow, Russia, 2018. [Google Scholar]
- Council of Europe. European Pharmacopoeia; Council of Europe: Strasbourg, France, 1997. [Google Scholar]
- Ramos, E.; Fernández, D.S. Classification of leaf epidermis microphotographs using texture features. Ecol. Inf. 2009, 4, 177–181. [Google Scholar] [CrossRef]
- Cope, J.S.; Corney, D.; Clark, J.Y.; Remagnino, P.; Wilkin, P. Plant species identification using digital morphometrics: A review. Expert Sys. Appl. 2012, 39, 7562–7573. [Google Scholar] [CrossRef]
- Sowa, I.; Zielinska, S.; Sawicki, J.; Bogucka-Kocka, A.; Staniak, M.; Bartusiak-Szczesniak, E.; Podolska-Fajks, M.; Kocjan, R.; Wójciak-Kosior, M. Systematic evaluation of chromatographic parameters for isoquinoline alkaloids on XB-C18 core-shell column using different mobile phase compositions. J. Anal. Methods Chem. 2018. [Google Scholar] [CrossRef] [Green Version]
- Lavinscky, M.P.; Silva, G.S.; Souza, M.M.; Melo, C.A.F. Contributions of classical and molecular cytogenetic in meiotic analysis and pollen viability for plant breeding. Genet. Mol. Res. 2017, 16, 1–15. [Google Scholar] [CrossRef]
- Harrison, C.J.; Alvey, E.; Henderson, I.R. Meiosis in flowering plants and other green organisms. J. Exp. Bot. 2010, 61, 2863–2875. [Google Scholar] [CrossRef] [Green Version]
- Lázaro-Diéguez, F.; Ispolatov, I.; Müsch, A. Cell shape impacts on the positioning of the mitotic spindle with respect to the substratum. Mol. Biol. Cell 2015, 26, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Baptista-Giacomelli, F.R.; Pagliarini, M.S.; Almeida, J.L. Meiotic behavior in several Brazilian oat cultivars (Avena sativa L.). Cytologia 2000, 65, 371–378. [Google Scholar] [CrossRef]
- Sheidai, M.; Koobaz, P.; Zehzad, B. Meiotic studies of some Avena, L. species and populations. Iran. J. Sci. 2003, 14, 121–131. [Google Scholar]
- Ghaffari, S.M. Occurrence of diploid and polyploid microspores in Sorghum bicolor (Poaceae) is the result of cytomixis. Afr. J. Biotech. 2006, 5, 1450–1453. [Google Scholar]
- Fadaei, F.; Sheidai, M.; Asadi, M. Cytological study of the genus Arenaria L. (Caryophyllaceae). Caryologia 2010, 63, 149–156. [Google Scholar] [CrossRef]
- Kumar, S.; Jeelani, S.M.; Rani, S.; Gupta, R.C.; Kumari, S. Cytology of five species of subfamily Papaveroideae from the Western Himalayas. Protoplasma 2012, 250, 307–316. [Google Scholar] [CrossRef]
- Kumari, K.; Saggoo, M.I.S. Male meiosis in two morphotypes of Melica persica Kunth (Poaceae) from Himachal Pradesh, India. Cytologia 2017, 81, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Kaur, D.; Singhal, V.K. Meiotic abnormalities affect genetic constitution and pollen viability in dicots from Indian cold deserts. BMC Plant Biol. 2019, 19, 10–19. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, C.; Yue, H.; Yang, R. Meiotic behavior and pollen fertility of five species in the genus Epimedium. Afr. J. Biotech. 2011, 10, 16189–16192. [Google Scholar] [CrossRef]
- Barros e Silva, A.E.; Guerra, M. The meaning of DAPI bands observed after C-banding and FISH procedures. Biotech. Histochem. 2010, 85, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Kovařík, A.; Leitch, A.R.; Garnatje, T. Cytogenetic features of rRNA genes across land plants: Analysis of the plant rDNA database. Plant J. 2017, 89, 1020–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muravenko, O.V.; Amosova, A.V.; Samatadze, T.E.; Popov, K.V.; Poletaev, A.I.; Zelenin, A.V. 9-aminoacridin- an efficient reagent to improve human and plant chromosome banding patterns and to standardize chromosome image analysis. Cytometry 2003, 51, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Lavania, U.C.; Kushwaha, J.; Lavania, S.; Basu, S. Chromosomal localization of rDNA and DAPI bands in solanaceous medicinal plant Hyoscyamus niger L. J. Genet. 2010, 89, 493–496. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Dias, A.L. ·Interindividual size heteromorphism of NOR and chromosomal location of 5S rRNA genes in Iheringichthys labrosus. Biol. App. Sci. 2007, 50, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Tutois, S.; Cloix, C.; Mathieu, O.; Cuvillier, C.; Tourmente, S. Analysis of 5S rDNA loci among Arabidopsis ecotypes and subspecies. Gen. Lett. 2002, 3, 115–122. [Google Scholar] [CrossRef]
- Simon, L.; Rabanal, F.A.; Dubos, T.; Oliver, C.; Lauber, D.; Poulet, A.; Vogt, A.; Mandlbauer, A.; Le Goff, S.; Sommer, A.; et al. Genetic and epigenetic variation in 5S ribosomal RNA genes reveals genome dynamics in Arabidopsis thaliana. Nucleic Acids Res. 2018, 46, 3019–3033. [Google Scholar] [CrossRef] [Green Version]
- Malankina, E.L.; Tsitsilin, A.N. Medicinal and Essential Oil Plants: Textbook; INFRA-M: Moscow, Russia, 2016. [Google Scholar]
- Beideman, I.N. The Method of Study of Phenology of Plants in Plant Communities; USSR Academy of Sciences: Moscow, Russia, 1974. [Google Scholar]
- Dospehov, B.A. Methods of Field Experience. (With the Basics of Statistical Processing of Research Results); Agropromizdat: Moscow, Russia, 1985. [Google Scholar]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Dyer, T.A. Sequence organization of the repeating units in the nucleus of wheat which contain 5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4865. [Google Scholar] [CrossRef] [Green Version]
- Amosova, A.V.; Zemtsova, L.V.; Yurkevich, O.Y.; Zhidkova, E.N.; Książczyk, T.; Shostak, N.G.; Muravlev, A.A.; Artemyeva, A.M.; Samatadze, T.E.; Zoshchuk, S.A.; et al. Genomic changes in generations of synthetic rapeseed-like allopolyploid grown under selection. Euphytica 2017, 213, 217–229. [Google Scholar] [CrossRef]
- Dolgova, A.A.; Ladygina, E.Y. Manual of Practical Pharmacognosy; Meditsina: Moscow, Russia, 2003. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Vegetative Parameters | Accession Number | ||||
|---|---|---|---|---|---|
| K 171-1 Control | K 211 | K 176 | K 117-18 | K 223-14 | |
| Number of flower spikes | 10.7 ± 0.7 | 8.0 ± 0.5 | 10.6 ± 0.6 | 9.8 ± 0.6 | 11.7 ± 0.9 |
| Number of flowers per plant | 121.5 ± 7.2 | 118.0 ± 7.0 | 120.5 ± 7.2 | 117.5 ± 6.3 | 125.5 ± 6.7 |
| Flower diameter, cm | 1.43 ± 0.08 | 1.58 ± 0.08 | 1.44 ± 0.08 | 1.37 ± 0.07 | 1.49 ± 0.06 |
| Number of petals in a flower | 4.0 ± 0.0 | 24.6 ± 1.56 | 4.0 ± 0.0 | 4.0 ± 0.0 | 4.0 ± 0.0 |
| Number of seeds per fruit | 50.2 ± 3.5 | 51.0 ± 3.6 | 50.1 ± 3.2 | 48.2 ± 3.1 | 51.2 ± 3.2 |
| Length of fruits, cm | 4.5 ± 0.3 | 4.0 ± 0.2 | 4.3 ± 0.2 | 4.2 ± 0.2 | 4.4 ± 0.3 |
| Seed yield, kg/ha | 18.6 ± 1.4 | 19.1 ± 1.7 | 18.2 ± 1.5 | 17.5 ± 1.5 | 19.9 ± 1.7 |
| Accession Number | Origin | Developmental Characteristics |
|---|---|---|
| K 171-1 control | Moscow region, RF (55°33′52″ N, 37°35′30″ E) | normal-flowered |
| K 211 | Moscow region, RF (55°33′52″ N, 37°35′30″ E) | double-flowered |
| K 176 | Prioksko-Terrasny Nature Reserve, RF (54°54′53″ N, 37°34′19″ E) | normal-flowered |
| K 117-18 | Petrozavodsk region, RF (61°50′57″ N, 34°19′54″ E) | normal-flowered |
| K 223-14 | Minsk region, Belarus (54°33′57″ N, 29°09′17″ E) | normal-flowered |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samatadze, T.E.; Yurkevich, O.Y.; Hazieva, F.M.; Konyaeva, E.A.; Morozov, A.I.; Zoshchuk, S.A.; Amosova, A.V.; Muravenko, O.V. Agro-Morphological, Microanatomical and Molecular Cytogenetic Characterization of the Medicinal Plant Chelidonium majus L. Plants 2020, 9, 1396. https://doi.org/10.3390/plants9101396
Samatadze TE, Yurkevich OY, Hazieva FM, Konyaeva EA, Morozov AI, Zoshchuk SA, Amosova AV, Muravenko OV. Agro-Morphological, Microanatomical and Molecular Cytogenetic Characterization of the Medicinal Plant Chelidonium majus L. Plants. 2020; 9(10):1396. https://doi.org/10.3390/plants9101396
Chicago/Turabian StyleSamatadze, Tatiana E., Olga Y. Yurkevich, Firdaus M. Hazieva, Elena A. Konyaeva, Alexander I. Morozov, Svyatoslav A. Zoshchuk, Alexandra V. Amosova, and Olga V. Muravenko. 2020. "Agro-Morphological, Microanatomical and Molecular Cytogenetic Characterization of the Medicinal Plant Chelidonium majus L." Plants 9, no. 10: 1396. https://doi.org/10.3390/plants9101396