
Abstract
Setd2 is the only enzyme that catalyzes histone H3 lysine 36 trimethylation (H3K36me3) on virtually all actively transcribed protein-coding genes, and this mechanism is evolutionarily conserved from yeast to human. Despite this widespread and conserved activity, Setd2 and H3K36me3 are dispensable for normal growth of yeast but are absolutely required for mammalian embryogenesis, such as oocyte maturation and embryonic vasculogenesis in mice, raising a question of how the functional requirements of Setd2 in specific developmental stages have emerged through evolution. Here, we explored this issue by studying the essentiality and function of Setd2 in zebrafish. Surprisingly, the setd2-null zebrafish are viable and fertile. They show Mendelian birth ratio and normal embryogenesis without vascular defect as seen in mice; however, they have a small body size phenotype attributed to insufficient energy metabolism and protein synthesis, which is reversable in a nutrition-dependent manner. Unlike the sterile Setd2-null mice, the setd2-null zebrafish can produce functional sperms and oocytes. Nonetheless, related to the requirement of maternal Setd2 for oocyte maturation in mice, the second generation of setd2-null zebrafish that carry no maternal setd2 show decreased survival rate and a developmental delay at maternal-to-zygotic transition. Taken together, these results indicate that, while the phenotypes of the setd2-null zebrafish and mice are apparently different, they are matched in parallel as the underlying mechanisms are evolutionarily conserved. Thus, the differential requirements of Setd2 may reflect distinct viability thresholds that associate with intrinsic and/or extrinsic stresses experienced by the organism through development, and these epigenetic regulatory mechanisms may serve as a reserved source supporting the evolution of life from simplicity to complexity.
Similar content being viewed by others

Introduction
While the life’s evolution from simplicity to complexity can benefit living beings on survival and reproduction, this transition also put them under tremendous pressures and stresses, such as demanded requirements of nutrient redistribution between cells and sexual propagation1,2. To effect this evolutionary transition, living beings must cope with these extra stresses by means of (i) acquiring mutations (including creation of new genes and regulatory elements) to increase genomic complexity1, and/or (ii) exploiting the potential of epigenetic programs which enables the organism to regulate gene expression more diversely and precisely2,3. Potentially relevant to the latter strategy is the notable enigma that some highly conserved and widespread epigenetic mechanisms, such as the methylation of histone H3 lysine 4 (H3K4) and lysine 36 (H3K36), are not essential for some lower species (e.g., yeast) growing in normal conditions, but they are absolutely required for the viability of higher organisms including mammalians4,5,6,7. It is thus conceivable that some epigenetic mechanisms, though paradoxically “non-essential”, are important for the organisms to survive some stress conditions, which provides a basis to support the evolution of life from simplicity to complexity.
H3K36 tri-methylation (H3K36me3) represents one of the most conserved epigenetic marks through the evolution of eukaryotes8, and it is associated with virtually all actively transcribed protein-coding genes9,10. An evolutionarily conserved histone methyltransferase that catalyzes H3K36me3 is responsible for establishing this association between H3K36me3 and the active genes, as it directly and specifically interacts with the elongating form of RNA polymerase II11,12,13,14,15. The yeast and mammalian version of this histone methyltransferase, named Set2 and SETD2, respectively, share a similar domain architecture containing the catalytic AWS-SET-PostSET domains and the Set2 RNA polymerase II-interacting (SRI) domain15,16. Both Setd2 and SETD2 have been shown to be the sole enzyme responsible for H3K36me3 in their hosts17,18,19, although in higher organisms there emerge several less conserved SET domain family members that likely catalyze H3K36me1 and H3K36me2, whose genomic distribution patterns do not resemble that of H3K36me320. The functions of Set2/SETD2 and H3K36me3 have been implicated in various aspects of gene regulation, including transcriptional elongation11,12,13,14,21,22, suppression of intragenic cryptic transcription23,24, nucleosome dynamics25, DNA repair26, DNA methylation27,28, N6-methyladenosine (m6A) mRNA modification29, and alternative mRNA splicing30. Furthermore, recent human cancer genomic studies have shown that somatic mutations, deletions, and dysregulated expression of SETD2, as well as altered H3K36me3 levels, are frequently identified in many types of cancers31,32,33,34,35, suggesting their pivotal roles in tumorigenesis and as potential therapeutic targets.
Despite such a high evolutionary conservation and the widely utilized functions of Set2/SETD2 and H3K36me3, loss of Set2 has little effect on the viability of yeasts in normal growth conditions; however, the set2-null yeast does show a slower growth in the condition of nutrition deprivation, compared with the wild-type36,37. In contrast, SETD2 is absolutely required for the viability and fertility of mammals, as a constitutive knockout of Setd2 in mice causes embryonic lethality due to defects in blood vessel development19, and Setd2 is also essential for spermiogenesis and oocyte development in mice38,39. Other tissue-specific studies in mice indicate that Setd2 regulates hematopoietic stem cell40,41 and bone marrow mesenchymal stem cell functions42, V(D)J recombination in lymphocytes43,44, as well as endodermal differentiation of embryonic stem cells45, though these functions may not necessarily be related to viability of the mice. These observations thus raise a question of how the functional requirements of H3K36me3 and Set2/SETD2 in the specific developmental stages have emerged through the evolution of life from simplicity to complexity. Here, we further explored this issue by using another vertebrate animal model, zebrafish, to investigate the essentiality and function of SETD2 in the development. It is surprising that setd2 knockout zebrafish is viable and fertile. However, a close comparison between these model organisms suggests that the apparently differential requirements of H3K36me3 and Set2/SETD2 in different organisms are actually well paralleled and likely associated with intrinsic and/or extrinsic stresses experienced by the organisms through development.
Results
setd2-null zebrafish is viable
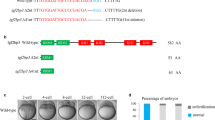
To disrupt the function of Setd2 in zebrafish, we designed CRISPR/Cas9 genome editing strategies to target the N-terminal end (mutation within exon 3) or the C-terminal SRI domain (mutation within exon 19) of zebrafish Setd2 protein (Fig. 1a). As a result, three mutant lines were generated. Sequencing results validated that two of them carried frame-shift mutations in exon 3 and therefore were named as E3fs1 and E3fs2, whereas the third carried a frame-shift mutation in exon 19 and was named as E19fs1 (Fig. 1a). We performed RT-qPCR to analyze setd2 mRNA levels in the embryos of the E3fs1 and E19fs1 lines, and the results showed a dramatic decrease of setd2 mRNA in the homozygous mutants of E3fs1 but not E19fs1 (Supplementary Fig. S1). These results suggest that the N-terminal frame-shift mutation (in exon 3) could induce nonsense-mediated mRNA decay (NMD), whereas the C-terminal frame-shift mutation (in exon 19) could not, probably due to the short distance between the frame-shit site and the natural stop codon. Therefore, the E19fs1 mutation likely gives rise to a truncated Setd2 without SRI domain, so that the E3fs1 and E19fs1 mutants represent different models in which Setd2 is either completely depleted or inhibited by disrupting the Setd2-RNA pol II interaction.

a Schematic diagram showing domain architecture of zebrafish Setd2 protein and mutated sites of the three mutant lines generated by CRISPR/Cas9 genome editing. The wild-type (WT) setd2 mRNA is referred to the NCBI Reference Sequence XM_009292184.3. Two mutants contain frame-shift mutations in exon 3 and are therefore named E3fs1 (c.215G>A216_217del; p.R72Qfs*9) and E3fs2 (c.217_244del; p.Q73fs*133), both of which disrupt the majority of the protein. The third mutant contains a frame-shift in exon 19, named E19fs1 (c.7910_7913GTGA>TTTC7914del; p.S2637Ifs*34), which disrupts the C-terminal SRI domain. Genomic sequences and deduced amino acids around targeting sequences in setd2 are shown. Red letters denote sgRNA-targeting sequences, while blue letters and dotted lines represent mutant sequences. b–d Genotype ratios of the WT, heterozygous (HE) and homozygous (HO) progenies, which are produced by self-cross of the three mutants, and their persistence through growth into adult. Note that they all fit the Mendelian Ratio. e Immunoblot analysis of the different H3K36 methylation states in the WT, HE and HO embryos of the E3fs1 and E19fs1 mutants. Total H3 was used as a loading control. Embryos at 72 hpf were analyzed. Each sample was loaded in 2-fold serial dilution to facilitate quantification. The immunoblot of the E3fs1 samples was actually analyzed in the same gels/explorations, but the images were rearranged just to fit the loading order in the lower panel. f Quantification of H3K36me3 levels in HE and HO relative to WT embryos of the E3fs1 and E19fs1 mutants.
We then analyzed the viability of the setd2 mutant zebrafish. In contrast to our previous finding that the homozygous Setd2 knockout mice are embryonic lethal19, it is surprising that self-cross of the heterozygous setd2 mutant zebrafish, for each of the three mutant lines, could produce wild-type, heterozygous and homozygous embryos, and their genotype frequencies perfectly fit the Mendelian ratio (Fig. 1b–d). Furthermore, a follow-up genotyping of each mutant line indicated that this ratio was maintained through the growth from birth to adult (Fig. 1b–d), suggesting that Setd2 is dispensable for embryonic development and growth of zebrafish.
Of special note, a phenomenon called genetic compensation has recently been proposed to explain some very mild or no phenotypes in gene knockout animal models, especially compared with the corresponding gene knockdown models46. An important mechanism underlying this phenomenon is transcriptional activation of homologous genes by an NMD-initiated, RNA fragment-mediated recruitment of the histone H3K4-methylating enzymes (e.g., the COMPASS complex) to the promoters of the homologous genes47,48. To test whether this mechanism is relevant to our setd2 mutant zebrafish, we first performed a Morpholino knockdown of Setd2. The results showed that, while the efficiency of Setd2 knockdown was verified by the significant inhibition of the EGFP reporter and the dramatic decrease of H3K36me3 (Supplementary Fig. S2a–c), the morphant embryos developed and grew normally (Supplementary Fig. S2b, d). We then performed RT-qPCR to examine whether the homologous genes of setd2 was upregulated in the E3fs1 mutant, which exhibited NMD as described above. The results showed that the mRNA levels of the setd2 homologues, including ash1l, nsd1a, nsd1b, nsd2 (whsc1) and nsd3 (whsc1l1), were not changed in the homozygous and heterozygous E3fs1 embryos, compared with the wild-type (Supplementary Fig. S3). Thus, these results suggest that the genetic compensation mechanisms may not be applicable to our observation of the non-essentiality of Setd2 in zebrafish viability.
Next, to determine the H3K36 methylation levels in the setd2 mutant zebrafish, we performed immunoblot analysis of the embryos at 72 hour post fertilization (hpf) with methylation state-specific antibodies. The results showed that H3K36me3 was dramatically decreased in both E3fs1 and E19fs1 homozygous mutants, compared with the wild-type and heterozygous, whereas their H3K36me1 and H3K36me2 remained intact (Fig. 1e, f). These patterns are exactly the same as what we previously observed in the Setd2 knockout mouse embryos19. The loss of H3K36me3 provides another line of evidence to exclude the genetic compensation effect and, in combination with previous studies, these results demonstrate that, while the specific role of Setd2 as the major enzyme for H3K36me3 is evolutionarily conserved, the functions of Setd2 and H3K36me3 are differentially required for the viability of different vertebrate animals.
Unlike mouse, zebrafish vascular development is not impaired by loss of setd2
Formation of blood vessels is a critical event both in the evolution from simple to complex species1 and in the development from the earliest non-vascular embryo, which receives nutrition by diffusion, to the highly vascular organism capable of quickly redistributing nutrition throughout the body49. Consistent with this notion, blood vessel development is induced, and tightly regulated, by various nutrient stresses such as hypoxia and amino acid starvation. In mammals, Setd2 plays an important role in this process, and loss of Setd2 leads to embryonic lethal at embryonic day (E) 10.5–11.5 due to a failure of embryonic vascular remodeling19. In sharp contrast, the homozygous setd2 mutant zebrafish can grow through the embryogenetic stage without apparent abnormalities; whole-mount in situ mRNA hybridization analysis with organ-specific markers indicated that their major organs, including liver (marked by lfabp), intestine (ifabp), thymus (rag1), heart (cmlc2), pancreas (trypsin), and pancreatic islet (insulin), are completely normal in the shape, size, location and developing stage (Fig. 2a) (analyses of the three mutant lines produced similar results; herein shown is E3fs1, which will be used as the default setd2 loss-of-function model in the following parts of the manuscript).

a Normal development of the major organs as indicated by whole mount in situ mRNA hybridization with specific markers (written in parentheses) in the embryos of indicated genotypes at 3 dpf. The numbers of embryos used in each staining experiment and of those showed the presented patterns are labelled as denominators and numerators, respectively. b Normal vascular development as indicated by confocal microscopy images of flk1:EGFP-labelled blood vessels in the trunk of the heterozygous and homozygous setd2 mutants at 50 and 72 hpf. c RT-qPCR analysis of the expression levels of representative vasculogenic genes, whose mouse orthologs have been known to be up- and downregulated in Setd2 knockout mice19. The analyzed setd2 mutant zebrafish embryos were at 72 hpf. In b, c, the embryos were produced by a cross between heterozygous and homozygous zebrafish so that the WT could only be analyzed separately and therefore was not shown. **P < 0.01; n.s., not significant.
We then closely analyzed the blood vessel development in the setd2-null embryos. An important tool for studying zebrafish blood vessels is the transgenic line Tg(flk1:EGFP), in which the blood vessels are labelled with the enhanced green fluorescent protein (EGFP) driven by the vascular endothelial cell-specific flk1 promoter50. Therefore, we crossed between this Tg(flk1:EGFP) line and our setd2-null mutant, and fluorescent microscope imaging of the EGFP-expressing embryos at different stages showed that the loss of Setd2 did not impair the vascular development in zebrafish (Fig. 2b). To further clarify this issue at molecular level, we performed RT-qPCR to analyze the vasculogenic genes whose mouse orthologues had been known to be up- or downregulated in the Setd2 knockout mice19. The results showed that none of these genes were altered by loss of Setd2 in zebrafish (Fig. 2c), thus further supporting that, unlike mouse, zebrafish does not require Setd2 function in their vascular development.
Nutrition-dependent, reversable smaller body size of setd2-null zebrafish
While the setd2 mutant zebrafish embryos have normal blood vessel development and organogenesis, the adult homozygous growing in the regular culture condition were found to have notably shorter body length, but without any developmental retardation at specific stage, compared with the heterozygous and wild-type siblings (Fig. 3a). This difference could be significantly measurable after 45 day post fertilization (dpf) (Fig. 3b) and, accordingly, the body weight of the homozygous were also lower than the heterozygous and wild-type siblings, which was started to be significantly measurable by an analytical balance as early as 25 dpf (Fig. 3c). Consistent with previous reports that the domesticated zebrafish strains (e.g., the herein used Tübingen strain) lack the natural sex-determination system51, and that their rapid growth can increase the proportion of female52, we found that the homozygous setd2 mutants had much smaller number of female under regular culture condition. Nonetheless, there was no obvious difference in food intaking or swimming behaviors between the mutants and the wild-type, as indicated by comparable frequencies of bite, areas of swimming, and involuntary tail movements, although these measurements could not exclude other ethological differences such as ability or willingness of competition, which may play a role in these phenotypes.

a Photographs of representative wild-type and setd2 mutant zebrafish growing in regular culture condition. b, c Quantification and statistical analysis of body length and body weight of the wild-type and setd2 mutant zebrafish of indicated ages. d Gene set enrichment analysis (GSEA) of RNA-seq data of the embryos at 36 hpf, showing that the genes related to energy metabolism, protein translation, and NMD are depleted in the homozygous but not the heterozygous setd2 mutant embryos, compared with the wild-type. e Activation of the PI3K-AKT-mTOR signaling pathway in the homozygous setd2 mutant embryos. f Quantification of total protein levels of individual dechorionated and deyolked embryos at early ages. g Rescue of the small body size phenotype by a separate feeding strategy started at 55 hpf. h Photographs of representative wild-type and setd2 mutant zebrafish growing in the separate feeding condition as in g. In a, h, scale bars = 1 cm. In b, c, f and g, data are showed as means ± SD; two-tailed t-test; ***P < 0.001; **P < 0.01; n.s., not significant.
To understand the underlying mechanism of how the setd2 deficiency causes this small body size phenotype, we performed RNA-seq analysis of the homozygous, heterozygous and wild-type littermate embryos at 36 hpf. The age of 36 hpf was chosen because at such an early stage the mutant phenotype has not emerged, and the nutrition supplied by the yolk is sufficient to support the growth of the embryo, so that the altered molecular mechanism at this stage may function as a cause rather than a consequence of the phenotype. Indeed, our gene set enrichment analysis (GSEA) of the RNA-seq data revealed that the genes associated with oxidative phosphorylation and protein translation are significantly depleted in the homozygous, but not the heterozygous, setd2 mutant embryos, compared with the wild-type (Fig. 3d, left and middle). Meanwhile, as it has been shown that inhibition of NMD acts as an important compensatory response to amino acid starvation53, we indeed found that the NMD-associated genes were also significantly downregulated in the setd2-null embryos (Fig. 3d, right), which, as reported53, could promote amino acid homeostasis by selectively retaining the transcripts with amino acid metabolism functions. Thus, these results suggest that the decreased activities of energy metabolism and protein synthesis have already been evident in the setd2-null embryos at early stage. Furthermore, we also found that the PI3K/AKT/mTOR signaling pathway was considerably activated in the setd2-null embryos (Fig. 3e), which possibly could improve the survival and growth of the setd2-null embryos that were suffering insufficient energy metabolism and protein synthesis, likely through the capacity of the PI3K/AKT/mTOR axis for sensing and responding to these stresses54. Interestingly, a direct cross-species comparison of the gene expression regulated by loss of Set2/Setd2 showed similar patterns in the yeasts under nutrient stress and the zebrafish and mouse embryos (Supplementary Fig. S4 and data not shown), which suggests an evolutionary conservation of these mechanisms.
Based on these possible mechanisms, we made two hypotheses: (i) a reduction of protein synthesis could be an early symptom that associates with the smaller body size of the setd2-null embryos; and (ii) an increase of nutrition supply may rescue, at least to a certain degree, this mutant phenotype. To test these hypotheses, we first quantified the total protein levels of individual embryos at different stages and compared between the setd2-null embryos and the siblings. As a result, it was as early as 4 dpf when the significantly different total protein levels of individual embryos were detectable (Fig. 3f), which suggests that the reduced protein synthesis indeed occurs very early, although it seems not severely affect the embryogenesis. We then performed a separate culturing and feeding strategy to increase nutrition and to prevent competition between the embryos. The results showed that this strategy largely rescued the small body size phenotype of the setd2-null embryos. Furthermore, a decrease of rearing density in mixed culture could also diminish the body size difference of the homozygous setd2 mutants compared with the wild-type and the heterozygous siblings (Supplementary Fig. S5). Meanwhile, the proportion of female of the homozygous setd2 mutants was also increased in these conditions. To explore the mechanism underlying this rescue effect, we performed RNA-seq analysis of the livers of the rescued and the control zebrafish. The results revealed that the genes associated with metabolism and protein synthesis were upregulated (Supplemental Fig. 6 and data not shown), which suggests an overactivation of these pathways in the setd2-null zebrafish that received increased nutrition. These results may explain why the small body size phenotype can be rescued in these conditions. Notably, the liver was chosen for this analysis because it provides relatively homogenous tissue and is easily obtainable. However, the upregulation of the metabolism- and protein synthesis-related genes is unlikely restricted to liver but possibly occurs in many tissue/cell types. Lastly, It is also notable that, even though the separate culture was applied at a late timepoint (e.g., 55 dpf) when the mutant phenotype had already been established, this phenotype could still be rescued (Fig. 3g, h). Taken together, these results strongly suggest that the small body size phenotype of the setd2-null zebrafish is likely associated with metabolic mechanisms, and that this phenotype is reversable and dependent on the nutrition supply.
setd2-null zebrafish are fertile in both male and female
We then investigated the reproductive functions of the setd2-null zebrafish. In contrast to the previous findings that the mouse Setd2 is required for sperm and oocyte development38,39,55, and that the germ cell-specific Setd2 knockout mice are sterile38,39, we surprisingly observed that both male and female homozygous setd2-null zebrafish could produce a number of progenies, suggesting that their reproductive functions should remain intact. To clarify this issue closely, we examined the reproductive processes of the setd2-null zebrafish ranging from germ cell development to fertilization. We first analyzed the primordial germ cells (PGCs), the precursors of both sperm and oocyte, by whole mount in situ mRNA hybridization with the germ cell-specific marker vasa (also known as ddx4)56,57. The results showed that the formation, migration, and number of PGCs in the homozygous setd2-null zebrafish were normal, as compared with those in the sibling controls (Fig. 4a). We then performed histological analysis of their testes and ovaries. Unlike the spermiogenic arrest caused by Setd2 deficiency in mice38, our H&E staining of the testes of the adult setd2-null zebrafish showed that the lobular cavities were filled with morphologically normal mature sperms, which aligned tightly and in proper order with their precursor cells (Fig. 4b). Quantification of the different cell types showed normal percentages of the spermatogonia, spermatocytes and spermatozoa in the testes (Fig. 4c). Meanwhile, H&E staining of the ovaries of the setd2-null zebrafish also showed normal morphology and percentages of the oocytes at different stages, compared with the sibling controls (Fig. 4d, e). Furthermore, given that some germ cell-specific genes have been shown to be expressed in specific stages of spermatogenesis, we performed RT-qPCR analysis of several of these genes, including amh (Sertoli cells)58, ziwi (also known as piwil1; type A spermatogonia)59, sycp3 (spermatocytes)60, cyp11c1 (Leydig cells)61,62, as well as the germ cell markers vasa56,57 and dnd63, in the adult testes. The results showed that all these genes were expressed normally in the setd2-null zebrafish compared with the sibling controls (Fig. 4f). Lastly, to determine whether the setd2 loss could affect the function of the sperms and oocytes, the homozygous and heterozygous setd2 mutant, as well as the wild-type, sperms and oocytes were subjected to in vitro fertilization with wild-type oocytes and sperms, respectively, and the successful fertilization rate was indicated by formation of blastodisc. As a result, for both sperms and oocytes, the groups of different genotypes showed similar fertilization rates (Fig. 4g, h), thus suggesting that the setd2-null sperms and oocytes are fully functional in the fertilization.

a Whole mount in situ hybridization with vasa showing the PGCs in the homozygous (HO) and heterozygous (HE) setd2 mutants at indicated stages. b H&E staining of the testes of adult setd2 mutant zebrafish at 3.5 month-post-fertilization (mpf). The boxed areas in the upper panels are magnified in the lower ones. Red and green arrows and red arrowheads denote spermatogonia, spermatocyte, and spermatozoa, respectively. c Quantification of the indicated cell types showing normal percentages of the spermatogonia, spermatocyte and spermatozoa in the testes of the setd2-null zebrafish. d H&E staining of the ovaries of adult setd2 mutant zebrafish at 3.5 mpf. The boxed areas in the upper panels are magnified in the lower ones. Red arrows denote the oocytes at primary growth stage (PG). PV, previtellogenic stage; EV, early vitellogenic stage; FG, full-grown stage. e Quantification of the indicated developmental stages, showing their normal pecentages in the total follicles. f RT-qPCR analysis of the germ cell-specific genes in the adult testes of setd2 mutant zebrafish. g, h The ratio of successful fertilization by setd2-null male (sperm) and female (oocyte) with wild-type oocyte and sperm, respectively, in the in vitro fertilization assay. In e–h, data are showed as means ± SD. n.s., not significant.
Maternal setd2 regulates survival rate of zebrafish embryos
We then examined whether the second generation of setd2-null zebrafish has any defect in their development. Interestingly, although a large number of the second generation of progenies could be produced by the setd2-null parents, we observed that, if the oocytes were derived from a homozygous setd2 mutant female, the fertilized embryos (i.e., the second generation) would have a decreased survival rate, which was regardless of the genotypes of the embryos as homozygous or heterozygous, even if the paternal genome was wild-type (Fig. 5a). In contrast, this decrease of survival rate was not seen in the mating between homozygous setd2 mutant sperms and wild-type oocytes (Fig. 5b). This finding was further validated by a comparison of four mating relationships between the heterozygous and homozygous setd2 mutants, which confirmed that, for example, the mating between homozygous oocytes and heterozygous sperms (HO♀ × HE♂), but not the opposite mating relationship (HE♀ × HO♂), led to the decreased survival rate (Fig. 5c).

a Decrease of survival rate of the embryos produced by setd2-null (homozygous setd2 mutant) female and wild-type male zebrafish. b Normal survival rate of the embryos produced by setd2-null male and wild-type female zebrafish. c A direct comparison among the four mating relationships between the homozygous and heterozygous parents. Note that only the setd2-null female could determine the survival rate, regardless of the mating patterners. d Allele-specific RT-qPCR analysis of the wild-type and mutant setd2 mRNAs along the development of the embryos of indicated genotypes. The red and green arrows denote the degradation of maternally deposited, mutant and wild-type alleles of setd2 mRNAs in the wild-type and homozygous setd2 mutant embryos, respectively. e Immunoblot analysis of H3K36me3 levels in the heterozygous and homozygous setd2 mutant embryos that carry maternal setd2. Blots of histone H3 were used as loading controls. Each sample was loaded in 2-fold serial dilution. Note the gradual decrease of H3K36me3 along the 72 h of development. f Immunoblot analysis of H3K36me3 in the homozygous setd2 mutant embryos in the absence of maternal setd2. Note that the control heterozygous embryos were impossible to be the littermate, therefore being labelled as “HE*”, because no heterozygous progeny could be produced by homozygous parents. Instead, the control heterozygous embryos were produced by crossing between homozygous male and wild-type female, and their H3K36me3 levels had been confirmed to be comparable with the “HE” use in e. g Quantification of the H3K36me3 levels in the indicated embryos based on the immunoblot results as shown in e, f.
A possible explanation of these observations is that setd2 is maternally deposited in the oocytes, which must play an important role in supporting a high survival rate of the fertilized embryos. Indeed, our previous genome-wide developmental expression profiling studies have identified setd2 as one of the maternal histone methyltransferases in zebrafish64. As the herein generated setd2 mutant zebrafish carry maternal setd2 mRNAs that are distinguishable from the zygotic ones by allele-specific RT-qPCR (i.e., the wild-type embryos carry maternally deposited mutant setd2 mRNA, and the homozygous setd2 mutant embryos carry wild-type mRNA), they provide an ideal model to further verify the deposition, function, and degradation of the maternal setd2 mRNAs during embryonic development. To this end, we designed PCR primers that could specifically detect the wild-type or the mutant setd2 mRNAs (Supplementary Fig. S7) and analyzed the embryos at different stages. The results showed that the maternal setd2 mRNA, both wild-type and mutant, were indeed present in the embryos and were significantly degraded by 6 hpf (Fig. 5d). Furthermore, immunoblot analysis of H3K36me3 levels in the setd2 mutant embryos also showed a trend toward decrease of H3K36me3 in the homozygous embryos that carried maternal setd2 mRNA and protein, and this decrease was much slower than the setd2 mRNA degradation (Fig. 5e). In contrast, in the embryos that were produced by homozygous parents and thus carried no maternal setd2 mRNA/protein, the H3K36me3 was quickly decreased (Fig. 5f). Therefore, this comparison suggests that, compared with the actively degraded setd2 mRNA, it takes more time to degrade the maternally deposited or maternal mRNA-translated Setd2 protein. Taken together, these results are strongly supportive of the existence and the potential function of the maternal setd2 in the embryonic development.
Loss of maternal setd2 causes a developmental delay at maternal-to-zygotic transition
Previous mouse studies have shown that the maternal Setd2 is absolutely required for proper zygotic genome activation (ZGA), and maternal depletion of Setd2 causes an arrest of development at 1-cell stage when the ZGA is initiating39. In zebrafish, ZGA occurs when the embryo generates ~1000 (1k) cells at 3 hpf, which is called maternal-to-zygotic transition (MZT). The apparently normal embryogenesis of many embryos that lack maternal setd2 (i.e., the second-generation progenies produced by setd2-null female; herein written as setd2mNull) indicated that the loss of maternal setd2 in zebrafish does not cause a complete arrest at MZT. However, we found that, while the setd2mNull embryos could keep the pace of the wild-type embryos to develop to the 1k-cell stage at 3 hpf, they exhibited a short delay afterward (Fig. 6a). In particular, when the wild-type embryos reached the oblong stage at 3.7 hpf, the setd2mNull embryos still remained at the high stage (Fig. 6a) and, subsequently, the setd2mNull embryos consistently showed a significant delay equivalent to 0.5–1 h of normal development65, compared with the wild-type embryos (Fig. 6a). These results suggest that this developmental delay of the setd2mNull embryos is tightly associated with MZT. Notably, a similar phenotype has been observed in the zebrafish embryos that lack maternal Ythdf2, and m6A-binding protein66, thus suggesting a potential link between Setd2/H3K36me3 and m6A mRNA modification in the regulation of MZT.

a A comparison between the wild-type (WT) and the maternal setd2-depleted (setd2mNull) embryos of indicated ages. The actual developing timepoints (in hpf) of the embryos are written on top, whereas the morphological stages are labelled in the lower right corners of the images. The red vertical bar denotes the MZT, which occurred after 3 hpf or 1k-cell stage, and the blue arrow denotes the developmental delay after MZT, as indicated by asynchronous division of the embryos. The numbers of embryos analyzed at each time point and of those showed the presented developmental stage are labelled as denominators and numerators, respectively. b Different somite numbers of the setd2mNull embryos compared with control embryos carrying maternal setd2. The control embryos with different genotypes were produced by self-crossing of heterozygous setd2 mutant zebrafish. The somites were labelled by whole mount in situ mRNA hybridization with myod. c Ratios of the setd2mNull and control embryos having indicated numbers of somites. The numbers labelled on the top denote the numbers of embryos analyzed for each group in b, c.
To further confirm this phenotype, and to exclude the possibility that the loss of zygotic setd2 (i.e., in the homozygous setd2 mutant) may also contribute to this developmental delay, we compared the setd2mNull embryos and the embryos produced by self-crossing of the heterozygous setd2 mutant; we also performed whole-mount in situ hybridization analysis of myod, which is specifically expressed in the somites and thus can be used to define developmental stages67. The results showed that, in the presence of maternal setd2, the wild-type, heterozygous and homozygous setd2 mutant embryos at 14.5 hpf could normally reach 9-somite stage, whereas most of the setd2mNull embryos remained at 7-somite stage (Fig. 6b, c). Notably, these statistics (i.e., almost all setd2mNull embryos undergoing developmental delay at MZT) are different from the above-described decreased survival rate (~50%), suggesting that the MZT-associated developmental delay and the decrease of survival rate are distinct phenotypes. Taken together, these results identify a specific role of the maternal setd2 in regulation of MZT, which is independent of whether setd2 is present in or absent from the zygotic genome.
Discussion
In this study, to gain a better understanding of the physiological function of Set2/Setd2 and H3K36me3 in an evolutionary context, we generated the setd2 mutant zebrafish and analyzed their phenotypes in a comparison with those of the yeast and mouse models. Given the high conservation of genetic and epigenetic regulatory mechanisms in vertebrate animals, it is surprising that the setd2-null zebrafish seem to have dramatically different phenotypes, especially in their viability and fertility, compared with the Setd2-null mice. However, our close studies reveal some interesting phenotypes in zebrafish, including the nutrition-dependent, reversable small body size and the developmental delay at MZT, which could be mechanistically connected with the known phenotypes in yeast and mice. These findings suggest that, despite the apparently different phenotypes in these species, the functions and mechanisms of Set2/Setd2 and H3K36me3 are actually conserved through the evolution of eukaryotes, even though they sometimes are non-essential for normal growth. Further, the seemingly different requirements of Set2/Setd2 and H3K36me3 in the development and reproduction of these species is likely attributed to the different levels of stresses associated with these functions that can be tolerated by the organism. Therefore, based on these studies, we propose a working model (Fig. 7) to explain the apparently different, but actually paralleled, phenotypes of the Set2/Setd2-null yeast, zebrafish and mouse.

The blue curves denote the levels of stresses that the organisms experience during normal development. The pink curve for yeast denotes an increased level of stress suffered by the yeast growing in nutrient depletion condition. The peaks of these curves are aligned vertically into two groups because they likely share conserved mechanisms in both nutrition and reproduction regulations. The red dashed lines denote the viability thresholds of different species when Set2/Setd2 is lost, and, therefore, the gray regions above the thresholds mean that only in this region where the Set2/Setd2-mediated epigenetic regulation is required. Therefore, if the peaks of the blue/pink curves reach above the red dashed lines, it means that the levels of stress are too high to be tolerated by the set2/setd2-null organism, thereby leading to lethality. In the cases of zebrafish, it is proposed that, as the stresses just reach (but not above) the viability threshold, they cause tolerable and apparently minor (but clearly significant) defects in the setd2-null zebrafish, thus explaining the non-requirement of setd2 in zebrafish development.
First, by comparing the three species, this model explains why the different phenotypes can be matched in parallel (Fig. 7, aligned peaks of the curves). In yeast, a phenotype of the Set2-null mutant is the growth defect under nutrient depletion but otherwise developing normally (in regular culture condition)36,37, suggesting a stress-related function of Set2. In the setd2-null zebrafish, the herein identified reversable phenotype of small body size also represents a growth problem and it is entirely dependent on nutrition supply, thus highly reminiscent of the yeast phenotype. Furthermore, our gene expression analysis reveals a decrease of the genes associated with energy metabolism and protein synthesis in the setd2-null embryos at early stages, thus providing a possible mechanism underlying this phenotype. Interestingly, these mechanisms seem conserved in yeast and vertebrates. In the Setd2-null mouse, although the phenotype in vasculogenesis19 seems unique and is not seen in zebrafish, it is probably attributed to the incompetence of the mutant mouse embryos in coping with the intrinsic nutrient stress. Indeed, as reported in our previous study19, a small body size phenotype actually is also observed in the Setd2-null mouse embryos at E8.5, and this phenotype is prior to the vasculogenic defect (at E10.5). Furthermore, recent studies showed that nutrient stress, such as amino acid restriction, can act as a proangiogenic trigger in mice68. Thus, the nutrient stress, small body size and vasculogenic defect can be mechanistically interconnected, suggesting the function and mechanism of Set2/Setd2 in regulating these phenotypes are evolutionarily conserved. Similarly, another group of paralleled phenotypes are that both Setd2-null zebrafish and mouse embryos show defects at the stage of MZT (or ZGA), which also suggest evolutionarily conserved functions and mechanisms of Setd2 in the regulation of reproduction.
Second, this model explains why the requirements of Set2/Setd2 for the viability of these species are different. Based on the present and previous studies, we use the concept of “viability threshold”, which represents the ability of an organism to tolerate certain factors such as stresses, to address this issue (Fig. 7, red dashed lines). While sharing many evolutionarily conserved molecular and cellular mechanisms, different species has distinct physiological structures and living environments, and therefore their viability thresholds can be widely variable69. For example, it has been known that the zebrafish embryos can obtain nutrition from the yolk by passive diffusion, so that they can tolerate higher degree of nutrient stress and have less dependence on the blood vessels to obtain and redistribute nutrition during early embryogenesis70. In contrast, the mouse embryos do not have a yolk as a source of nutrition, and they must rely on the development of a complex vascular system to obtain nutrition from the mother69. This difference may explain the apparently different phenotypes of setd2-null zebrafish and mouse embryos in the body size, blood vessel development and embryonic viability, which further supports that Setd2 and the related epigenetic regulatory mechanisms are important for the organisms to properly respond to nutrient stress. Another example is about the different phenotypes of the maternal setd2-depleted zebrafish and mouse embryos at the stage of MZT — while the loss of maternal Setd2 leads to a complete arrest of the mouse embryos, it seems to cause only a short delay of zebrafish embryos at this stage. It is conceivable that these phenotypes are regulated, at least partially, by conserved mechanisms of Setd2 in reprogramming the epigenome, as recently observed in mice39, and that the difference in the phenotypes is possibly related to some specific features of these species. In this regard, an important feature of zebrafish embryos is that the MZT is equivalent to 1000-cell stage, relative to mouse MZT at 2-cell stage71. Furthermore, epigenetic reprogramming of parental genomes in zebrafish also show some differences from that of mice72,73,74. Thus, although the detailed mechanism remains to be clarified, these morphological and mechanistic differences between zebrafish and mouse embryos may be directly relevant to the different requirements of maternal Setd2 for the embryos to complete MZT.
Lastly, by establishing the connection between Set2/Setd2 and the abilities of the organisms to cope with stresses, this model reemphasizes the important role of the epigenetic programs as a reserved source supporting the evolution of living beings from simplicity to complexity2,75. In the evolution, vasculogenesis and sexual reproduction represent two major advantages for multicellular organisms to obtain and redistribute nutrition and to maintain genetic stability and diversity1. It is interesting that Set2/Setd2 is involved into both processes. This is probably because the emergence of these processes had relied on the existing regulatory mechanisms capable of dealing with extra stresses, such as Set2/Setd2 and related epigenetic mechanisms. As such, the role of Set2/Setd2 should be changed from non-essential to essential for some specific developmental processes, as well as the viability. Also related to this notion, the epigenetic regulators (e.g., the homologues of Set2/Setd2) have been greatly expanded in the numbers, specificities, and regulatory mechanisms through eukaryotic evolution, and they play nonredundant roles in many aspects of development64,75. Taken together, these results and analysis suggest that the epigenetic regulatory system may serve as a buffer not only for coping with intrinsic and/or extrinsic stresses but also for the evolution of life from simplicity to complexity.
Materials and methods
Zebrafish strains
The zebrafish wild-type Tübingen strain (ZFIN ID: ZDB-GENO-990623-3) and the transgenic zebrafish line Tg(flk1:EGFP)50 (ZFIN ID: ZDB-ALT-050916-14) were used in this study. All zebrafish husbandry was achieved under standard conditions (temperature: 28.5 °C; pH: 7.0–8.0; light cycle: 12 h-on/12 h-off). The embryos were maintained in egg water (containing 60 μg/mL sea salt and 0.2% methylene blue) at 28.5 °C and 0.045% N-Phenylthiourea (PTU; Sigma-Aldrich) was performed to prevent pigmentation.
Establishment of setd2 mutant lines by the CRISPR/Cas9 system
The genomic sequence of zebrafish setd2 gene was obtained from the National Center for Biotechnology Information. An online tool (ZiFit Targeter, http://zifit.partners.org/ZiFiT/Disclaimer.aspx) was used to design guide RNA (gRNA) target sites in exon 3 and exon 19 containing 20-base pair (bp) target sequences (E3: cgtcaaaatgtgtttctacc; E19: ggcagacacgtccagtgagc). The predicted sequence was cloned into p-T7-gRNA vector backbone to generate the guide RNA expression construct. The specific single-guide RNAs (sgRNAs) was produced by in vitro transcription with the mMESSAGE mMACHINE SP6 Transcription Kit (Thermo Fisher, AM1340). The sgRNA (10 ng/μL) and Cas9 protein (200 ng/mL) (EnGen Cas9 NLS; New England Biolabs, M0646T) were co-injected into zebrafish embryos at the one-cell stage. A dozen of injected embryos were subjected to DNA extraction and genotyping at 24 hpf. The rest of founders (F0) were raised to adulthood and the individuals carrying mutations were identified by genotyping on caudal fin with sequencing. The F0 founders bearing mosaic mutations at the target sites were then outcrossed with WT fish to produce F1 heterozygotes. The F1 heterozygotes harboring same frame-shift mutation were incrossed to generate F2 homozygous mutants.
Protein quantification
To compare the total amount of embryos protein at different time points during the early development, every embryo was dechorionated and pipetted into standard protein RIPA lysis buffer for sonication. Then six embryos with same genotype were merged to perform BCA protein assay kit (Thermo Scientific, 23227). Three biological replicates were measured for each time point to determine the average protein amount.
Immunoblot
Zebrafish embryos were deyolked as described previously76 and the proteins were extracted by RIPA buffer (Beyotime Biotechnology, P0013D) containing appropriate protease inhibitors (MedChemExpress, HY-K0010) with sonication. Boiled lysates were first separated with SDS-PAGE and transferred onto PVDF membranes. The separated proteins were immunoblotted with antibodies for H3K36me1 (Abcam, ab9049), H3K36me2 (Abcam, ab9048), H3K36me3 (Abcam, ab9050) and H3 (Cell Signaling Technology, 4499S).
Live imaging of blood vessels
To observe the vasculogenesis of setd2 mutants, the transgenic zebrafish line Tg(flk1:EGFP)50 was crossed with the setd2 mutants. Live embryos were anesthetized with 0.03% Ethyl 3-aminobenzoate methanesulfonate salt (Sigma-Aldrich, A5040) and mounted in 1% low melting agarose (Sangon Biotech, A600015). Fluorescence images were captured with a scanning confocal microscope (Olympus, FV1000) processed with Image-Pro Plus 6.0 (Media Cybernetics).
Whole-mount mRNA in situ hybridization
The PGCs were labelled with vasa; and the somites were labelled with myod. The major organs were labelled specific markers including ifabp, lfabp, cmlc2, rag1, trypsin, and insulin. Whole-mount mRNA in situ hybridization was performed as previously described77,78. Anti-digoxigenin Fab fragment antibodies conjugated to alkaline phosphatase (Roche, 1277073) were used for detection of the RNA probes, and this was followed by NBT/BCIP color reaction (Vector Laboratories, SK-5400).
RNA isolation and quantitative reverse transcription PCR (RT-qPCR)
Total RNA was isolated with TRIzol reagent (Ambion, 15596018) according to the manufacturer’s instructions. The amount and purity of the RNA were determined by spectrophotometry and agarose gel electrophoresis. After treatment with RNase-free DNase, total RNA was reverse transcribed using random hexamers and oligo(dT) primers (Takara, RR037Q). qPCR was carried out on an ABI Prism 7900HT Sequence Detection System (Applied Biosystems) using the SYBR Green Real-Time PCR Master Mix kit (TOYOBO, QPK-201). The expression levels of target genes were normalized against the internal control β-actin (actb1) gene.
RNA-seq and bioinformatic analysis
Total RNA was extracted from whole embryos at 36 hpf using TRIzol (Ambion, 15596018). Library construction with fragmented mRNA was performed with TruSeq stranded mRNA library prep kit (Illumina). Sequencing was carried out on Illumina HiSeq 2000 according to the manufacturer’s instructions. The deep-sequencing data were mapped to zebrafish genome version 10 (GRCz10) with Tophat and Cufflink (v2.2.1) was used to calculate the FPKM of each gene to represent their mRNA expression level. Gene Set Enrichment Analysis (GSEA) was performed to identify regulated signaling pathways and to interpret gene expression patterns globally.
Histological analysis and morphometric measurement
After anesthetized with 0.03% Ethyl3-aminobenzoate methanesulfonate salt (Tricaine), the gonadal tissues were dissected fixed overnight in 4% paraformaldehyde (Servicebio, G1101-20) and then subjected to paraffin embedding. Serial sections (4 μm) were cut, deparaffinized, and stained with H&E. All genotypes zebrafish collected for morphometric analysis, including somatic length and body weight, were measured from juvenile to adult at a 10-day interval. Meanwhile, different genotype raising in same tank were isolated feeding at 55 dpf, as morphometric record continued.
Data availability
RNA-seq data are accessible through the Gene Expression Omnibus (GEO) accession code GSE151238.
References
King, N. The unicellular ancestry of animal development. Dev. Cell 7, 313–325 (2004).
Turner, B. M. Epigenetic responses to environmental change and their evolutionary implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364, 3403–3418 (2009).
Cavalli, G. & Heard, E. Advances in epigenetics link genetics to the environment and disease. Nature 571, 489–499 (2019).
Lee, J. S. & Shilatifard, A. A site to remember: H3K36 methylation a mark for histone deacetylation. Mutat. Res 618, 130–134 (2007).
Henikoff, S. & Shilatifard, A. Histone modification: cause or cog? Trends Genet 27, 389–396 (2011).
Howe, F. S., Fischl, H., Murray, S. C. & Mellor, J. Is H3K4me3 instructive for transcription activation? Bioessays 39, 1–12 (2017).
Crump, N. T. & Milne, T. A. Why are so many MLL lysine methyltransferases required for normal mammalian development? Cell Mol. Life Sci. 76, 2885–2898 (2019).
Kouzarides, T. Histone methylation in transcriptional control. Curr. Opin. Genet Dev. 12, 198–209 (2002).
Pokholok, D. K. et al. Genome-wide map of nucleosome acetylation and methylation in yeast. Cell 122, 517–527 (2005).
Barski, A. et al. High-resolution profiling of histone methylations in the human genome. Cell 129, 823–837 (2007).
Li, J., Moazed, D. & Gygi, S. P. Association of the histone methyltransferase Set2 with RNA polymerase II plays a role in transcription elongation. J. Biol. Chem. 277, 49383–49388 (2002).
Li, B., Howe, L., Anderson, S., Yates, J. R. 3rd & Workman, J. L. The Set2 histone methyltransferase functions through the phosphorylated carboxyl-terminal domain of RNA polymerase II. J. Biol. Chem. 278, 8897–8903 (2003).
Xiao, T. et al. Phosphorylation of RNA polymerase II CTD regulates H3 methylation in yeast. Genes Dev. 17, 654–663 (2003).
Krogan, N. J. et al. Methylation of histone H3 by Set2 in Saccharomyces cerevisiae is linked to transcriptional elongation by RNA polymerase II. Mol. Cell Biol. 23, 4207–4218 (2003).
Sun, X. J. et al. Identification and characterization of a novel human histone H3 lysine 36-specific methyltransferase. J. Biol. Chem. 280, 35261–35271 (2005).
Kizer, K. O. et al. A novel domain in Set2 mediates RNA polymerase II interaction and couples histone H3 K36 methylation with transcript elongation. Mol. Cell Biol. 25, 3305–3316 (2005).
Strahl, B. D. et al. Set2 is a nucleosomal histone H3-selective methyltransferase that mediates transcriptional repression. Mol. Cell Biol. 22, 1298–1306 (2002).
Edmunds, J. W., Mahadevan, L. C. & Clayton, A. L. Dynamic histone H3 methylation during gene induction: HYPB/Setd2 mediates all H3K36 trimethylation. EMBO J. 27, 406–420 (2008).
Hu, M. et al. Histone H3 lysine 36 methyltransferase Hypb/Setd2 is required for embryonic vascular remodeling. Proc. Natl Acad. Sci. USA 107, 2956–2961 (2010).
Wagner, E. J. & Carpenter, P. B. Understanding the language of Lys36 methylation at histone H3. Nat. Rev. Mol. Cell Biol. 13, 115–126 (2012).
Joshi, A. A. & Struhl, K. Eaf3 chromodomain interaction with methylated H3-K36 links histone deacetylation to Pol II elongation. Mol. Cell 20, 971–978 (2005).
Keogh, M. C. et al. Cotranscriptional set2 methylation of histone H3 lysine 36 recruits a repressive Rpd3 complex. Cell 123, 593–605 (2005).
Carrozza, M. J. et al. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell 123, 581–592 (2005).
Neri, F. et al. Intragenic DNA methylation prevents spurious transcription initiation. Nature 543, 72–77 (2017).
Venkatesh, S. et al. Set2 methylation of histone H3 lysine 36 suppresses histone exchange on transcribed genes. Nature 489, 452–455 (2012).
Li, F. et al. The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSalpha. Cell 153, 590–600 (2013).
Baubec, T. et al. Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature 520, 243–247 (2015).
Morselli, M. et al. In vivo targeting of de novo DNA methylation by histone modifications in yeast and mouse. Elife 4, e06205 (2015).
Huang, H. et al. Histone H3 trimethylation at lysine 36 guides m(6)A RNA modification co-transcriptionally. Nature 567, 414–419 (2019).
Luco, R. F. et al. Regulation of alternative splicing by histone modifications. Science 327, 996–1000 (2010).
Dalgliesh, G. L. et al. Systematic sequencing of renal carcinoma reveals inactivation of histone modifying genes. Nature 463, 360–363 (2010).
Zhu, X. et al. Identification of functional cooperative mutations of SETD2 in human acute leukemia. Nat. Genet 46, 287–293 (2014).
Chen, B. Y. et al. SETD2 deficiency accelerates MDS-associated leukemogenesis via S100a9 in NHD13 mice and predicts poor prognosis in MDS. Blood 135, 2271–2285 (2020).
Patnaik, M. M. & Abdel-Wahab, O. SETD2-linking stem cell survival and transformation. Cell Res 28, 393–394 (2018).
Hu, M., Hu, M., Zhang, Q., Lai, J. & Liu, X. SETD2, an epigenetic tumor suppressor: a focused review on GI tumor. Front Biosci. (Landmark Ed.) 25, 781–797 (2020).
Morris, S. A. et al. Histone H3 K36 methylation is associated with transcription elongation in Schizosaccharomyces pombe. Eukaryot. Cell 4, 1446–1454 (2005).
McDaniel, S. L. et al. H3K36 Methylation Regulates Nutrient Stress Response in Saccharomyces cerevisiae by Enforcing Transcriptional Fidelity. Cell Rep. 19, 2371–2382 (2017).
Zuo, X. et al. The histone methyltransferase SETD2 is required for expression of acrosin-binding protein 1 and protamines and essential for spermiogenesis in mice. J. Biol. Chem. 293, 9188–9197 (2018).
Xu, Q. et al. SETD2 regulates the maternal epigenome, genomic imprinting and embryonic development. Nat. Genet 51, 844–856 (2019).
Zhang, Y. L. et al. Setd2 deficiency impairs hematopoietic stem cell self-renewal and causes malignant transformation. Cell Res 28, 476–490 (2018).
Zhou, Y. et al. Setd2 regulates quiescence and differentiation of adult hematopoietic stem cells by restricting RNA polymerase II elongation. Haematologica 103, 1110–1123 (2018).
Wang, L. et al. H3K36 trimethylation mediated by SETD2 regulates the fate of bone marrow mesenchymal stem cells. PLoS Biol. 16, e2006522 (2018).
Ji, Z. et al. The histone methyltransferase Setd2 is indispensable for V(D)J recombination. Nat. Commun. 10, 3353 (2019).
Chu, S. H. et al. Loss of H3K36 methyltransferase SETD2 impairs V(D)J recombination during lymphoid development. iScience 23, 100941 (2020).
Zhang, Y. et al. H3K36 histone methyltransferase Setd2 is required for murine embryonic stem cell differentiation toward endoderm. Cell Rep. 8, 1989–2002 (2014).
El-Brolosy, M. A. & Stainier, D. Y. R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet 13, e1006780 (2017).
El-Brolosy, M. A. et al. Genetic compensation triggered by mutant mRNA degradation. Nature 568, 193–197 (2019).
Ma, Z. et al. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature 568, 259–263 (2019).
Conway, E. M., Collen, D. & Carmeliet, P. Molecular mechanisms of blood vessel growth. Cardiovasc Res 49, 507–521 (2001).
Jin, S. W., Beis, D., Mitchell, T., Chen, J. N. & Stainier, D. Y. Cellular and molecular analyses of vascular tube and lumen formation in zebrafish. Development 132, 5199–5209 (2005).
Wilson, C. A. et al. Wild sex in zebrafish: loss of the natural sex determinant in domesticated strains. Genetics 198, 1291–1308 (2014).
Lawrence, C., Ebersole, J. P. & Kesseli, R. V. Rapid growth and out-crossing promote female development in zebrafish (Danio rerio). Environ. Biol. Fishes 81, 239–246 (2007).
Mendell, J. T., Sharifi, N. A., Meyers, J. L., Martinez-Murillo, F. & Dietz, H. C. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat. Genet 36, 1073–1078 (2004).
Laplante, M. & Sabatini, D. M. mTOR signaling in growth control and disease. Cell 149, 274–293 (2012).
Li, C. et al. Histone methyltransferase SETD2 is required for meiotic maturation in mouse oocyte. J. Cell Physiol. 234, 661–668 (2018).
Olsen, L. C., Aasland, R. & Fjose, A. A vasa-like gene in zebrafish identifies putative primordial germ cells. Mech. Dev. 66, 95–105 (1997).
Yoon, C., Kawakami, K. & Hopkins, N. Zebrafish vasa homologue RNA is localized to the cleavage planes of 2- and 4-cell-stage embryos and is expressed in the primordial germ cells. Development 124, 3157–3165 (1997).
Skaar, K. S. et al. Proteolytically activated, recombinant anti-mullerian hormone inhibits androgen secretion, proliferation, and differentiation of spermatogonia in adult zebrafish testis organ cultures. Endocrinology 152, 3527–3540 (2011).
Tan, C. H. et al. Ziwi, the zebrafish homologue of the Drosophila piwi: co-localization with vasa at the embryonic genital ridge and gonad-specific expression in the adults. Mech. Dev. 119(Suppl 1), S221–S224 (2002).
Ozaki, Y., Saito, K., Shinya, M., Kawasaki, T. & Sakai, N. Evaluation of Sycp3, Plzf and Cyclin B3 expression and suitability as spermatogonia and spermatocyte markers in zebrafish. Gene Expr. Patterns 11, 309–315 (2011).
Zheng, Q. et al. Loss of Cyp11c1 causes delayed spermatogenesis due to the absence of 11-ketotestosterone. J. Endocrinol. 244, 487–499 (2020).
Zhang, Q., Ye, D., Wang, H., Wang, Y. & Sun, Y. Zebrafish cyp11c1 knockout reveals the roles of 11-ketotestosterone and cortisol in sexual development and reproduction. Endocrinology 161, bqaa048 (2020).
Weidinger, G. et al. dead end, a novel vertebrate germ plasm component, is required for zebrafish primordial germ cell migration and survival. Curr. Biol. 13, 1429–1434 (2003).
Sun, X. J. et al. Genome-wide survey and developmental expression mapping of zebrafish SET domain-containing genes. PLoS ONE 3, e1499 (2008).
Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. & Schilling, T. F. Stages of embryonic development of the zebrafish. Dev. Dyn. 203, 253–310 (1995).
Zhao, B. S. et al. m(6)A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature 542, 475–478 (2017).
Weinberg, E. S. et al. Developmental regulation of zebrafish MyoD in wild-type, no tail and spadetail embryos. Development 122, 271–280 (1996).
Longchamp, A. et al. Amino Acid Restriction Triggers Angiogenesis via GCN2/ATF4 Regulation of VEGF and H2S Production. Cell 173, 117–129 (2018).
Copp, A. J. Death before birth: clues from gene knockouts and mutations. Trends Genet 11, 87–93 (1995).
Stainier, D. Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet 2, 39–48 (2001).
Tadros, W. & Lipshitz, H. D. The maternal-to-zygotic transition: a play in two acts. Development 136, 3033–3042 (2009).
Wu, S. F., Zhang, H. & Cairns, B. R. Genes for embryo development are packaged in blocks of multivalent chromatin in zebrafish sperm. Genome Res 21, 578–589 (2011).
Jiang, L. et al. Sperm, but not oocyte, DNA methylome is inherited by zebrafish early embryos. Cell 153, 773–784 (2013).
Potok, M. E., Nix, D. A., Parnell, T. J. & Cairns, B. R. Reprogramming the maternal zebrafish genome after fertilization to match the paternal methylation pattern. Cell 153, 759–772 (2013).
Huang, S. Inverse relationship between genetic diversity and epigenetic complexity. Nature Precedings, https://doi.org/10.1038/npre.2009.1751.2 (2009).
Link, V., Shevchenko, A. & Heisenberg, C. P. Proteomics of early zebrafish embryos. BMC Dev. Biol. 6, 1 (2006).
Song, H. D. et al. Hematopoietic gene expression profile in zebrafish kidney marrow. Proc. Natl Acad. Sci. USA 101, 16240–16245 (2004).
Thisse, C. & Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 3, 59–69 (2008).
Acknowledgements
We thank Dr. Jin-Biao Ma (Fudan University) for helpful discussions. This work was supported by the National Key Research and Development Plan of China 2018YFA0107802 (X.-J.S.), 2018YFC1004500 (B.L.), the National Natural Science Foundation of China (NSFC) General Program 81670094, 81470316 (X.-J.S.), 31872817 (B.L.), the Shanghai Municipal Education Commission-Gaofeng Clinical Medicine Grant 20152506 (X.-J.S.), Shanghai Collaborative Innovation Program on Regenerative Medicine and Stem Cell Research 2019CXJQ01 (S.-J.C. and X.-J.S.), Innovative Research Team of High-Level Local Universities in Shanghai (W.-L.Z. and X.-J.S.), the Samuel Waxman Cancer Research Foundation and the Shanghai Guangci Translational Medical Research Development Foundation. X.-J.S. was supported by the 1000 Talents Program for Young Scholars.
Author information
Authors and Affiliations
Contributions
D.-J.L., and X.-J.S. designed research; D.-J.L., F.Z., Y.C., Y.J., Y.-L.Z., S.-B.C., Y.-Y.X., and Q.-H.H. performed research; W.-L.Z., and L.W. contributed new reagents/analytic tools; D.-J.L., P.-F.X., Z.C., S.-J.C., B.L., A.Z., and X.-J.S. analyzed data; D.-J.L., and X.-J.S. wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, DJ., Zhang, F., Chen, Y. et al. setd2 knockout zebrafish is viable and fertile: differential and developmental stress-related requirements for Setd2 and histone H3K36 trimethylation in different vertebrate animals. Cell Discov 6, 72 (2020). https://doi.org/10.1038/s41421-020-00203-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41421-020-00203-8
