Spatial Changes in Microbial Communities along Different Functional Zones of a Free-Water Surface Wetland

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
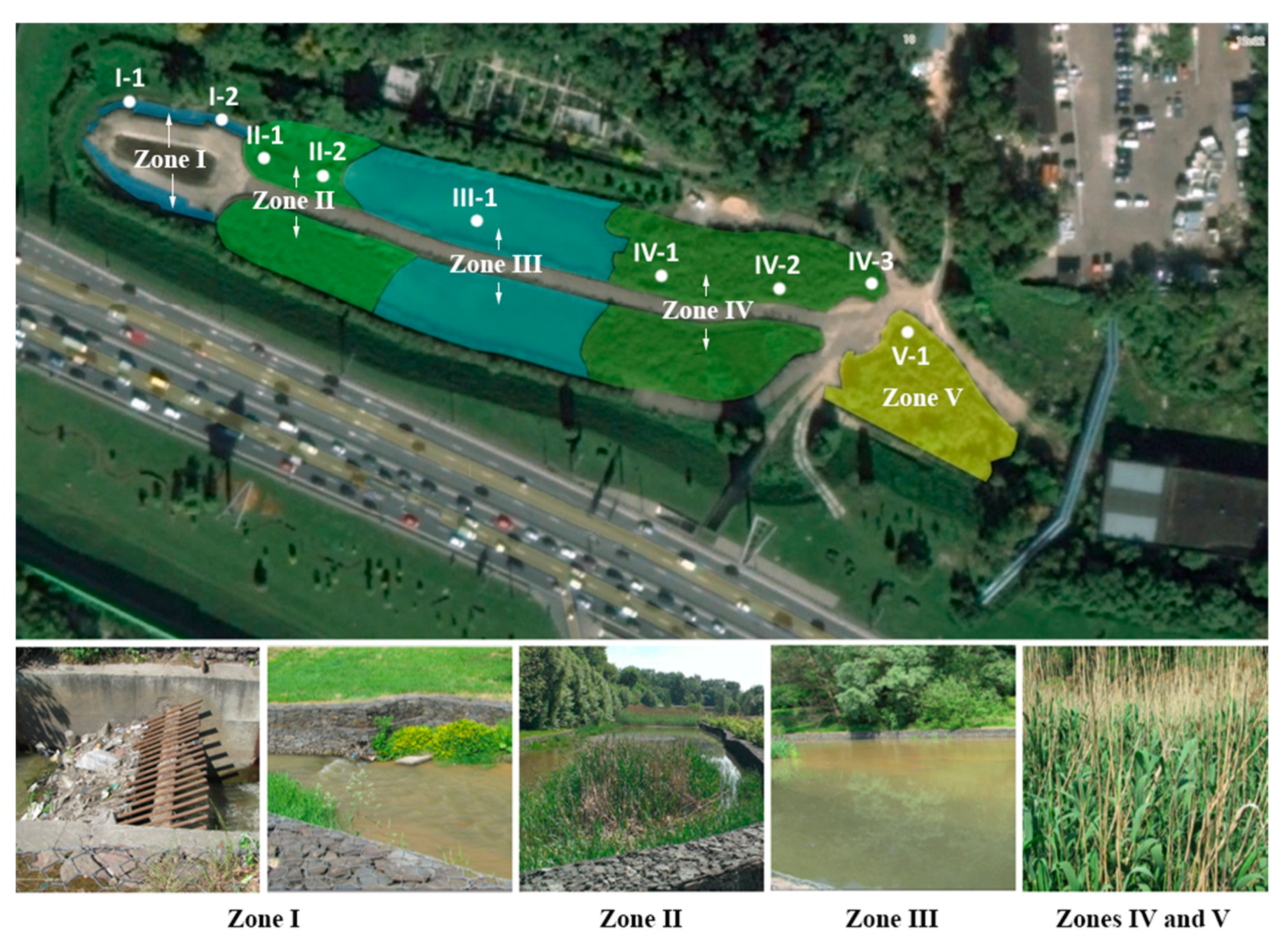
2.1. Constructed Wetland Design and Operation
2.2. Organic Contaminants Analysis
2.3. Particle-Size Distribution Analysis
2.4. Total DNA Extraction and Amplicon Sequencing
2.5. Statistics and Bioinformatics
3. Results
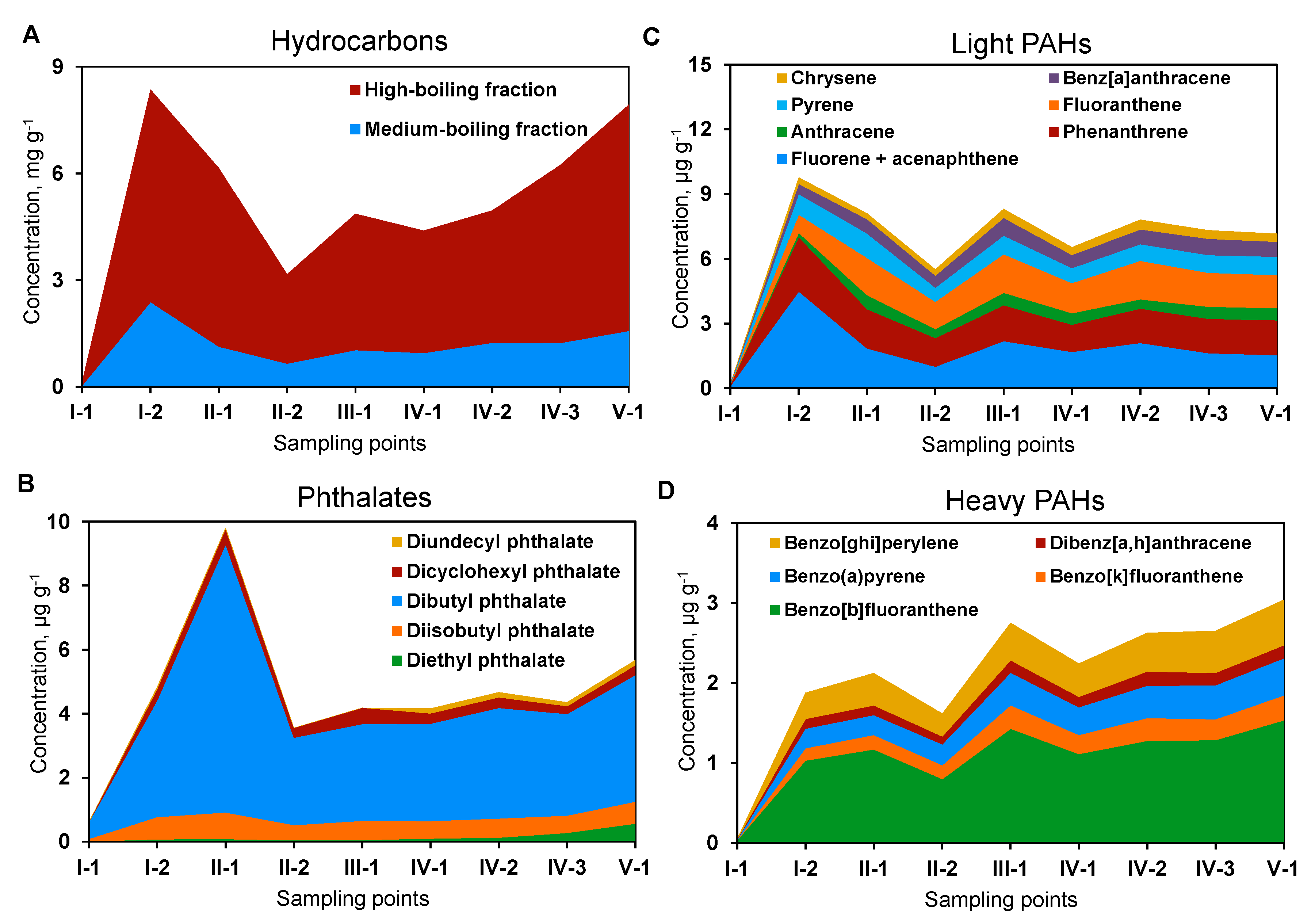
3.1. Chemical Composition of Organic Pollutants in the Sediments
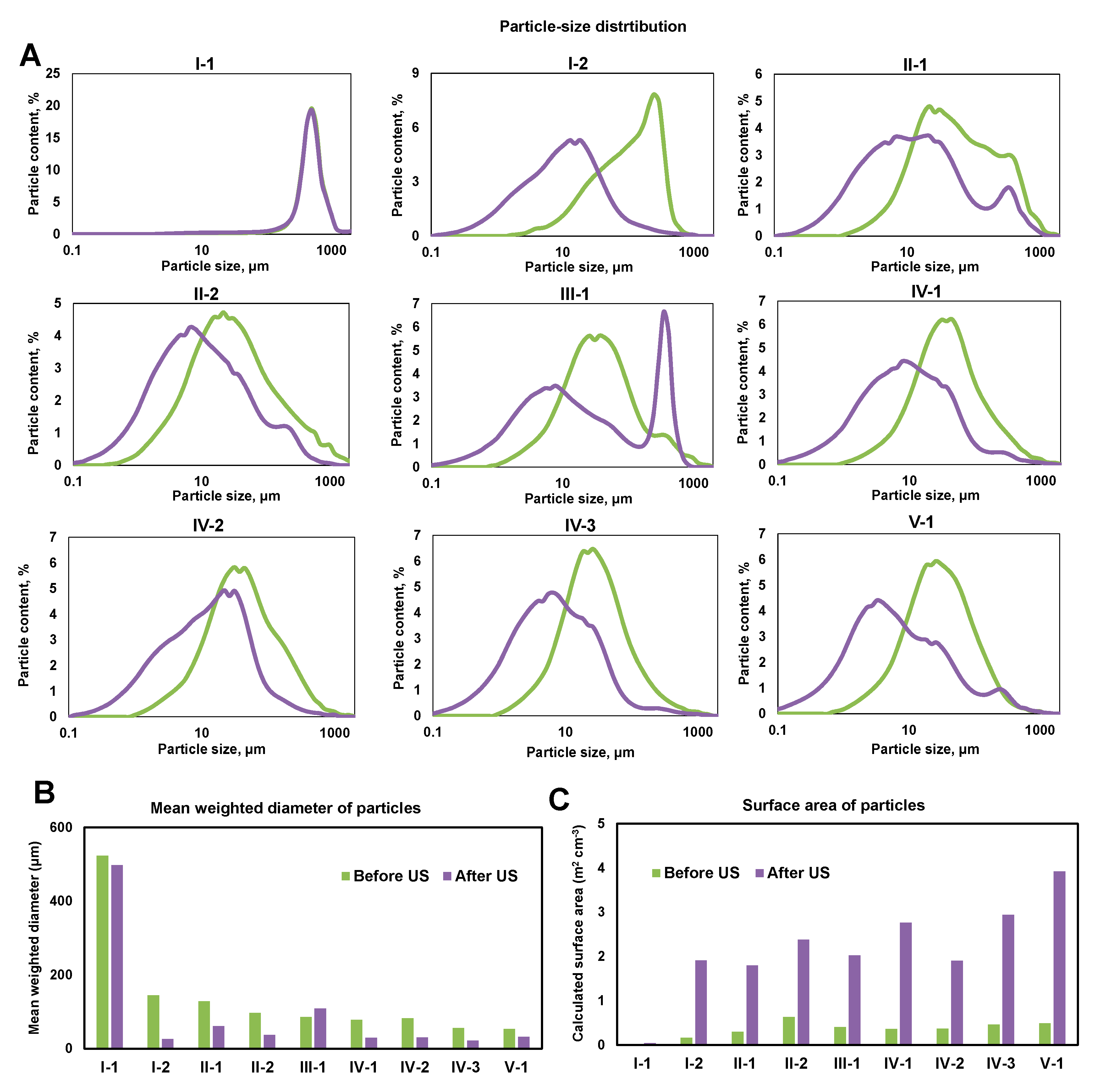
3.2. Particle-Size Distribution (PSD) of the Sediments
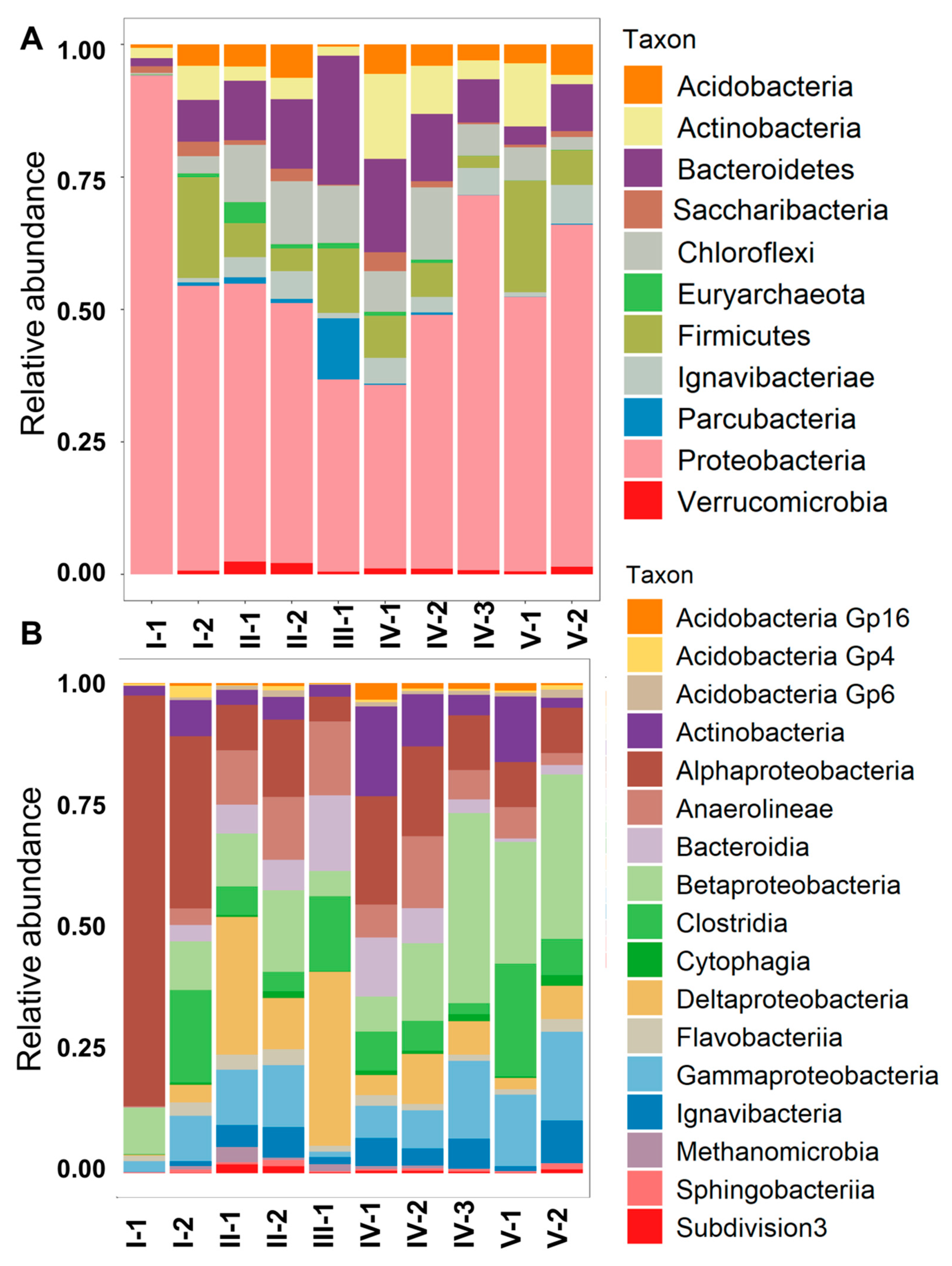
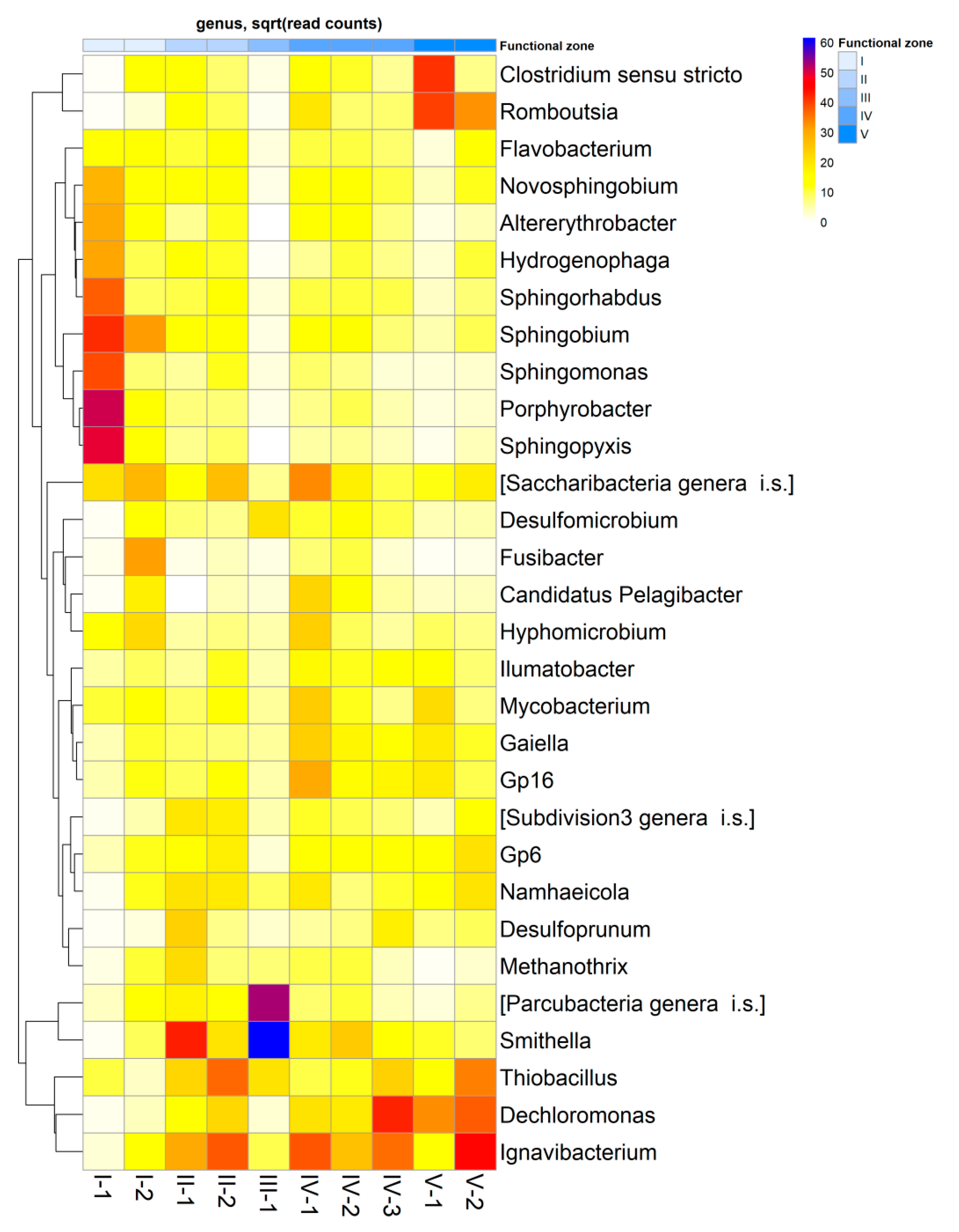
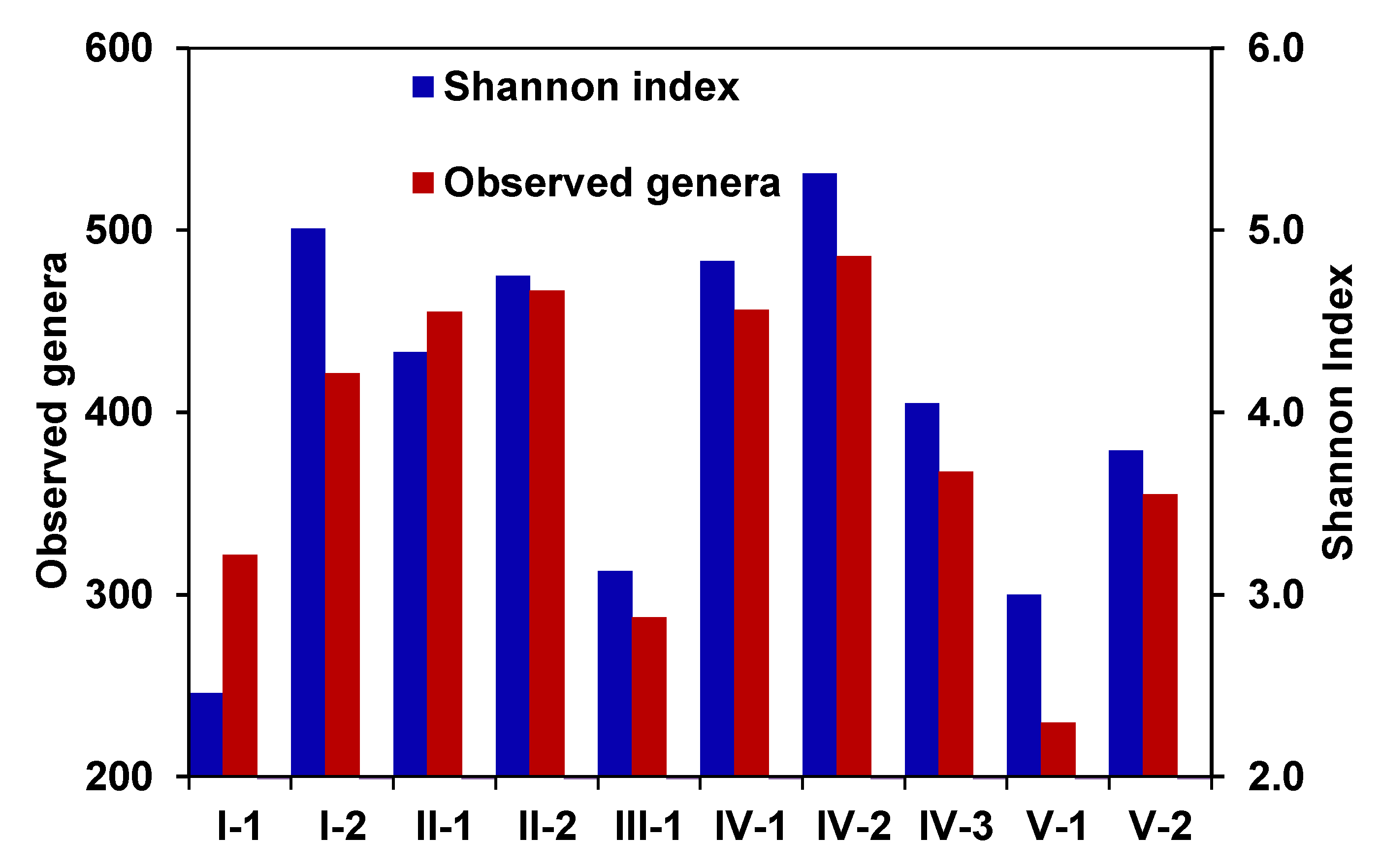
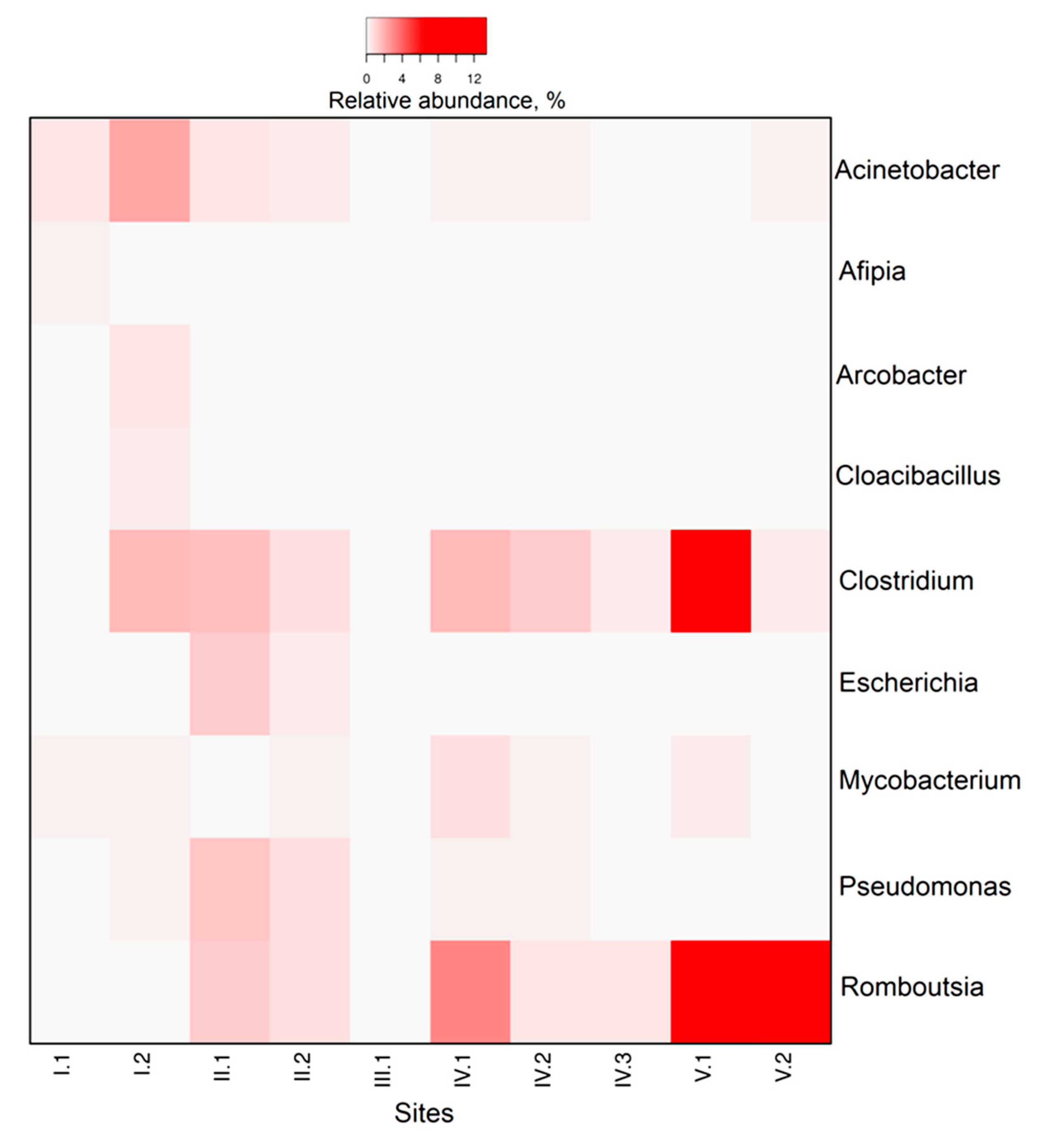
3.3. Microbial Community Composition and Diversity
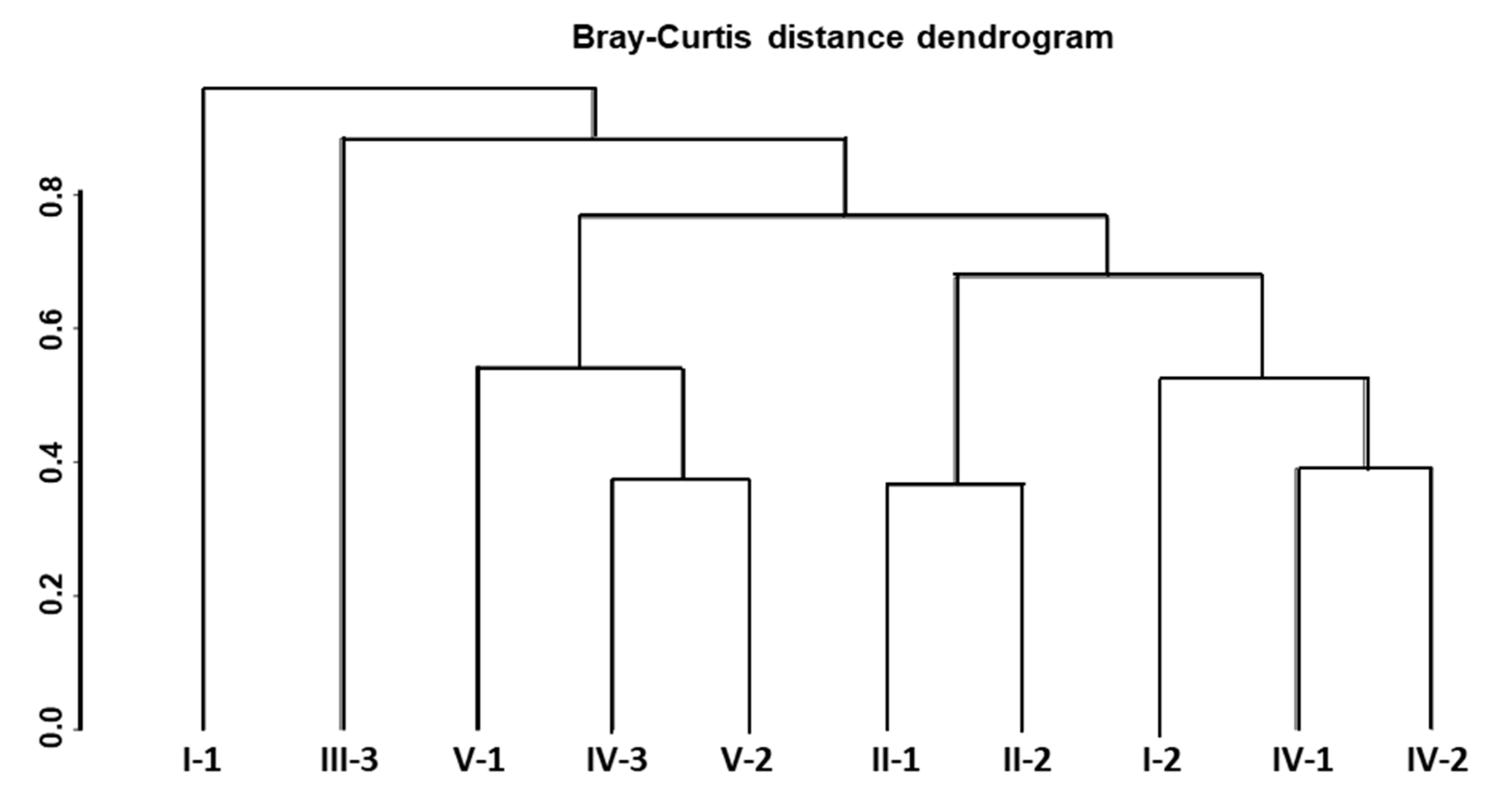
3.4. Diversity and Community Clustering
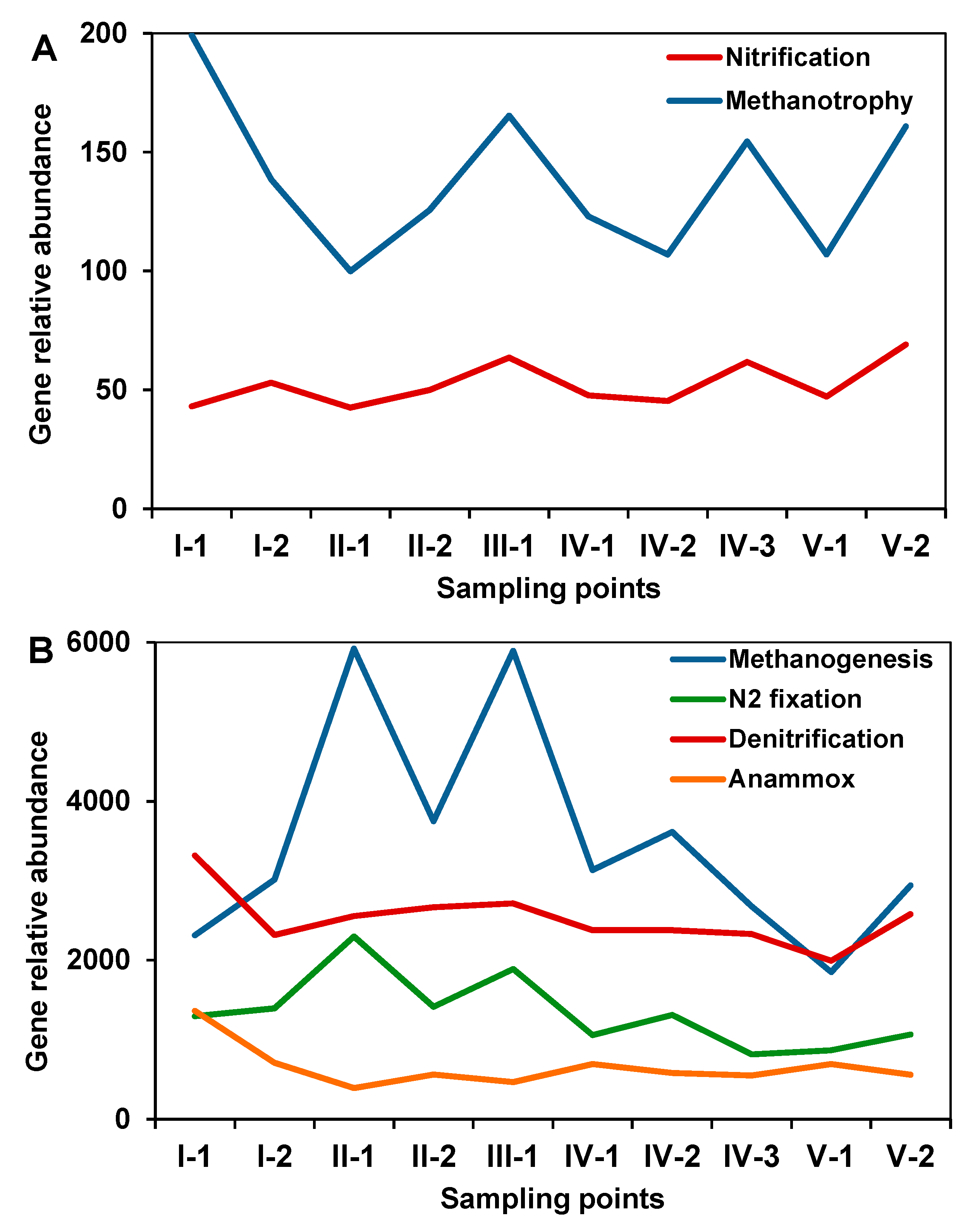
3.5. Prediction of Microbial Functional Profiles
3.6. Disinfection in the CW
4. Discussion
4.1. Functional Zones of the Constructed Wetland Are Characterized by Contrasting Physicochemical Properties of the Sediments
4.2. Functional Zones of the Constructed Wetland Are Characterized by Distinct Taxonomic and Functional Microbial Profiles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Carty, A.; Scholz, M.; Heal, K.; Gouriveau, F.; Mustafa, A. The universal design, operation and maintenance guidelines for farm constructed wetlands (FCW) in temperate climates. Bioresour. Technol. 2008, 99, 6780–6792. [Google Scholar] [CrossRef] [PubMed]
- Rai, U.N.; Tripathi, R.D.; Singh, N.K.; Upadhyay, A.K.; Dwivedi, S.; Shukla, M.K.; Mallick, S.; Singh, S.N.; Nautiyal, C.S. Constructed wetland as an ecotechnological tool for pollution treatment for conservation of Ganga river. Bioresour. Technol. 2013, 148, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, J.; Ngo, H.H.; Guo, W.; Hu, Z.; Liang, S.; Fun, J.; Liu, H. A review on the sustainability of constructed wetlands for wastewater treatment: Design and operation. Bioresour. Technol. 2015, 175, 594–601. [Google Scholar] [CrossRef]
- Scholz, M.; Lee, B.H. Constructed wetlands: A review. Int. J. Environ. Res. 2005, 62, 421–447. [Google Scholar] [CrossRef]
- Masi, F.; Martinuzzi, N. Constructed wetlands for the Mediterranean countries: Hybrid systems for water reuse and sustainable sanitation. Desalination 2007, 215, 44–55. [Google Scholar] [CrossRef]
- Vymazal, J. Constructed wetlands for wastewater treatment. Water 2010, 2, 530–549. [Google Scholar] [CrossRef] [Green Version]
- Vymazal, J. Constructed wetlands for treatment of industrial wastewaters: A review. Ecol. Eng. 2014, 73, 724–751. [Google Scholar] [CrossRef]
- Faulwetter, J.L.; Gagnon, V.; Sundberg, C.; Chazarenc, F.; Burr, M.D.; Brisson, J.; Camper, A.K.; Stein, O.R. Microbial processes influencing performance of treatment wetlands: A review. Ecol. Eng. 2009, 35, 987–1004. [Google Scholar] [CrossRef]
- Díaz, F.J.; Anthony, T.O.; Dahlgren, R.A. Agricultural pollutant removal by constructed wetlands: Implications for water management and design. Agric. Water Manag. 2012, 104, 171–183. [Google Scholar] [CrossRef]
- Sundberg, C.; Tonderski, K.; Lindgren, P.E. Potential nitrification and denitrification and the corresponding composition of the bacterial communities in a compact constructed wetland treating landfill leachates. Water Sci. Technol. 2007, 56, 159–166. [Google Scholar] [CrossRef]
- Butterworth, E.; Richards, A.; Jones, M.; Brix, H.; Dotro, G.; Jefferson, B. Impact of aeration on macrophyte establishment in sub−surface constructed wetlands used for tertiary treatment of sewage. Ecol. Eng. 2016, 91, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, R.H.; Wallace, S. Treatment Wetlands, 2nd ed.; Lewis Publishers, CRC Press: New York, NY, USA, 2008; pp. 181–280. [Google Scholar]
- Braeckevelt, M.; Rokadia, H.; Imfeld, G.; Stelzer, N.; Paschke, H.; Kuschk, P.; Kastner, M.; Richnow, H.-H.; Weber, S. Assessment of in situ biodegradation of monochlorobenzene in contaminated groundwater treated in a constructed wetland. Environ. Pollut. 2007, 148, 428–437. [Google Scholar] [CrossRef]
- Matamoros, V.; Mujeriego, R.; Bayona, J.M. Trihalomethane occurrence in chlorinated reclaimed water at full−scale wastewater treatment plants in NE Spain. Water Res. 2007, 41, 3337–3344. [Google Scholar] [CrossRef]
- Masi, F.; Hamouri, B.E.; Abdel Shafi, H.; Baban, A.; Ghrabi, A.; Regelsberger, M. Treatment of segregated black/grey domestic wastewater using constructed wetlands in the Mediterranean basin: The zer0−m experience. Water Sci. Technol. 2010, 61, 97–105. [Google Scholar] [CrossRef]
- Wu, S.; Kuschk, P.; Brix, H.; Vymazal, J.; Dong, R. Development of constructed wetlands in performance intensifications for wastewater treatment: A nitrogen and organic matter targeted review. Water Res. 2014, 57, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Ávila, C.; Bayona, J.M.; Martín, I.; Salas, J.J.; García, J. Emerging organic contaminant removal in a full−scale hybrid constructed wetland system for wastewater treatment and reuse. Ecol. Eng. 2015, 80, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Xu, S.; Lu, S.; Qin, P.; Bi, B.; Ding, H.; Liu, Y.; Guo, X.; Liu, X. A review on removal of organophosphorus pesticides in constructed wetland: Performance, mechanism and influencing factors. Sci. Total Environ. 2019, 651, 2247–2268. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, V.; Hijosa, M.; Bayona, J.M. Assessment of the pharmaceutical active compounds removal in wastewater treatment systems at enantiomeric level. Ibuprofen and naproxen. Chemosphere 2009, 75, 200–205. [Google Scholar] [CrossRef]
- Verlicchi, P.; Zambello, E. How efficient are constructed wetlands in removing pharmaceuticals from untreated and treated urban wastewaters? A review. Sci. Total Environ. 2014, 470, 1281–1306. [Google Scholar] [CrossRef]
- Zhang, D.Q.; Jinadasa, K.B.S.N.; Gersberg, R.M.; Liu, Y.; Ng, W.J.; Tan, S.K. Application of constructed wetlands for wastewater treatment in developing countries–a review of recent developments (2000–2013). J. Environ. Manag. 2014, 141, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Mander, Ü.; Dotro, G.; Ebie, Y.; Towprayoon, S.; Chiemchaisri, C.; Nogueira, S.F.; Jamsranjav, B.; Kasak, K.; Truu, J.; Tournebiz, J.; et al. Greenhouse gas emission in constructed wetlands for wastewater treatment: A review. Ecol. Eng. 2014, 66, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, J.R.P.; Cabriales, J.J.P.; Vega, M.M. Identification and characterization of sulfur−oxidizing bacteria in an artificial wetland that treats wastewater from a tannery. Int. J. Phytoremed. 2008, 10, 359–370. [Google Scholar] [CrossRef]
- Krasnits, E.; Friedler, E.; Sabbah, I.; Beliavski, M.; Tarre, S.; Green, M. Spatial distribution of major microbial groups in a well established constructed wetland treating municipal wastewater. Ecol. Eng. 2009, 35, 1085–1089. [Google Scholar] [CrossRef]
- Ansola, G.; Arroyo, P.; de Miera, L.E.S. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total Environ. 2014, 473, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Truu, M.; Juhanson, J.; Truu, J. Microbial biomass, activity and community composition in constructed wetlands. Sci. Total Environ. 2009, 407, 3958–3971. [Google Scholar] [CrossRef] [PubMed]
- Morató, J.; Codony, F.; Sánchez, O.; Pérez, L.M.; García, J.; Mas, J. Key design factors affecting microbial community composition and pathogenic organism removal in horizontal subsurface flow constructed wetlands. Sci. Total Environ. 2014, 481, 81–89. [Google Scholar] [CrossRef]
- Alexandros, S.I.; Akratos, C.S. Removal of pathogenic bacteria in constructed wetlands: Mechanisms and efficiency. In Phytoremediation; Ansari, A., Gill, S., Gill, R., Lanza, G., Newman, L., Eds.; Springer: Cham, Switzerland, 2016; Volume 4, pp. 327–346. [Google Scholar]
- Yang, M.; Guo, Q.; Tong, T.; Li, N.; Xie, S.; Long, Y. Vegetation type and layer depth influence nitrite−dependent methane−oxidizing bacteria in constructed wetland. Arch. Microbiol. 2017, 199, 505–511. [Google Scholar] [CrossRef]
- Du, L.; Chen, Q.; Liu, P.; Zhang, X.; Wang, H.; Zhou, Q.; Xu, D.; Wu, Z. Phosphorus removal performance and biological dephosphorization process in treating reclaimed water by Integrated Vertical−flow Constructed Wetlands (IVCWs). Bioresour. Technol. 2017, 243, 204–211. [Google Scholar] [CrossRef]
- Zhong, F.; Wu, J.; Dai, Y.; Yang, L.; Zhang, Z.; Cheng, S.; Zhang, Q. Bacterial community analysis by PCR−DGGE and 454−pyrosequencing of horizontal subsurface flow constructed wetlands with front aeration. Appl. Microbiol. Biotechnol. 2015, 99, 1499–1512. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X.D.; Liu, Y.S.; Ying, G.G.; Liu, S.S.; He, L.Y.; Su, H.C.; Hu, L.X.; Chen, F.R.; Yang, Y.Q. Removal of antibiotics and antibiotic resistance genes from domestic sewage by constructed wetlands: Optimization of wetland substrates and hydraulic loading. Sci. Total Environ. 2016, 565, 240–248. [Google Scholar] [CrossRef]
- Weerakoon, G.M.P.R.; Jinadasa, K.B.S.N.; Herath, G.B.B.; Mowjood, M.I.M.; Van Bruggen, J.J.A. Impact of the hydraulic loading rate on pollutants removal in tropical horizontal subsurface flow constructed wetlands. Ecol. Eng. 2013, 61, 154–160. [Google Scholar] [CrossRef]
- Lin, Y.F.; Jing, S.R.; Lee, D.Y.; Wang, T.W. Nutrient removal from aquaculture wastewater using a constructed wetlands system. Aquaculture 2002, 209, 169–184. [Google Scholar] [CrossRef]
- Sochan, A.; Polakowski, C.; Łagód, G. Impact of optical indices on particle size distribution of activated sludge measured by laser diffraction method. Ecol. Chem. Eng. 2014, 21, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, M.V.; Manucharova, N.A.; Krasnov, G.S.; Nikitin, D.A.; Stepanov, A.L. Biomass and taxonomic structure of microbial communities in soils of the right−bank basin of the Oka River. Eurasian Soil Sci. 2019, 52, 971–981. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next−generation sequencing−based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Qin, Z.; Zhang, D.; Hussain, J. Dissipation characteristics of pyrene and ecological contribution of submerged macrophytes and their biofilms−leaves in constructed wetland. Bioresour. Technol. 2018, 267, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Jin, M.; Zhou, X.; Chen, W.; Lu, D.; Zhang, Y.; Shao, X. Enhanced removal mechanism of iron carbon micro−electrolysis constructed wetland on C, N, and P in salty permitted effluent of wastewater treatment plant. Sci. Total Environ. 2019, 649, 21–30. [Google Scholar] [CrossRef]
- Qin, Z.; Zhao, Z.; Adam, A.; Li, Y.; Chen, D.; Mela, S.M.; Li, H. The dissipation and risk alleviation mechanism of PAHs and nitrogen in constructed wetlands: The role of submerged macrophytes and their biofilms−leaves. Environ. Int. 2019, 131, 104940. [Google Scholar] [CrossRef]
- Ruppelt, J.P.; Tondera, K.; Wallace, S.J.; Button, M.; Pinnekamp, J.; Weber, K.P. Assessing the role of microbial communities in the performance of constructed wetlands used to treat combined sewer overflows. Sci. Total Environ. 2020, 736, 139519. [Google Scholar] [CrossRef]
- Tan, X.; Yang, Y.L.; Li, X.; Zhou, Z.W.; Liu, C.J.; Liu, Y.W.; Yin, W.C.; Fan, X.Y. Intensified nitrogen removal by heterotrophic nitrification aerobic denitrification bacteria in two pilot−scale tidal flow constructed wetlands: Influence of influent C/N ratios and tidal strategies. Bioresour. Technol. 2020, 302, 122803. [Google Scholar] [CrossRef]
- Parikh, H.I.; Koparde, V.N.; Bradley, S.P.; Buck, G.A.; Sheth, N.U. MeFiT: Merging and filtering tool for illumina paired−end reads for 16S rRNA amplicon sequencing. BMC Bioinform. 2016, 17, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras−Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4 Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Jin, B.; Wilén, B.M.; Lant, P. A comprehensive insight into floc characteristics and their impact on compressibility and settleability of activated sludge. Chem. Eng. J. 2003, 95, 221–234. [Google Scholar] [CrossRef]
- Jin, B.; Wilén, B.M.; Lant, P. Impacts of morphological, physical and chemical properties of sludge flocs on dewaterability of activated sludge. Chem. Eng. J. 2004, 98, 115–126. [Google Scholar] [CrossRef]
- Kuśnierz, M. Scale of Small Particle Population in Activated Sludge Flocs. Water Air Soil Pollut. 2018, 229, 327. [Google Scholar] [CrossRef] [Green Version]
- Zita, A.; Hermansson, M. Effects of bacterial cell surface structures and hydrophobicity on attachment to activated sludge flocs. Appl. Environ. Microbiol. 1997, 63, 1168–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilén, B.-M.; Onuki, M.; Hermansson, M.; Lumley, D.; Mino, T. Microbial community structure in activated sludge floc analysed by fluorescence in situ hybridization and its relation to floc stability. Water Res. 2008, 42, 2300–2308. [Google Scholar]
- Semenov, M.V.; Manucharova, N.A.; Stepanov, A.L. Distribution of metabolically active prokaryotes (Archaea and Bacteria) throughout the profiles of chernozem and brown semidesert soil. Eurasian Soil Sci. 2016, 49, 217–225. [Google Scholar] [CrossRef]
- Loeppmann, S.; Semenov, M.; Kuzyakov, Y.; Blagodatskaya, E. Shift from dormancy to microbial growth revealed by RNA: DNA ratio. Ecol. Indic. 2018, 85, 603–612. [Google Scholar] [CrossRef]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The ecology of Acidobacteria: Moving beyond genes and genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; van Bruggen, A.H. Long−term fertilization rather than plant species shapes rhizosphere and bulk soil prokaryotic communities in agroecosystems. Appl. Soil Ecol. 2020, 154, 103641. [Google Scholar] [CrossRef]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Hentrissat, B.; Coutinho, B.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, M.; Blagodatskaya, E.; Stepanov, A.; Kuzyakov, Y. DNA−based determination of soil microbial biomass in alkaline and carbonaceous soils of semi−arid climate. J. Arid. Environ. 2018, 150, 54–61. [Google Scholar] [CrossRef]
- Balkwill, D.L.; Fredrickson, J.K.; Romine, M.F. Sphingomonas and related genera. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 7, pp. 605–629. [Google Scholar]
- Verma, H.; Kumar, R.; Oldach, P.; Sangwan, N.; Khurana, J.P.; Gilbert, J.A.; Lal, R. Comparative genomic analysis of nine Sphingobium strains: Insights into their evolution and hexachlorocyclohexane (HCH) degradation pathways. BMC Genom. 2014, 15, 1014. [Google Scholar] [CrossRef] [Green Version]
- Godoy, F. Sphingopyxischilensis sp. nov.; a chlorophenol−degrading bacterium that accumulates polyhydroxyalkanoate, and transfer of Sphingomonas alaskensis to Sphingopyxis alaskensis comb. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 473–477. [Google Scholar] [CrossRef]
- Schmidt, G.; Tischler, D.; Oelschlägel, M.; Kalinowski, J.; Rückert, C.; Schlömann, M. Sphingopyxis fribergensis sp. nov., a soil bacterium with the ability to degrade styrene and phenylacetic acid. Int. J. Syst. Evol. Microbiol. 2015, 65, 3008–3015. [Google Scholar]
- Liu, Z.-P.; Wang, B.-J.; Liu, Y.-H.; Liu, S.-J. Novosphingobium taihuense sp. nov., a novel aromatic−compound−degrading bacterium isolated from Taihu Lake, China. Int. J. Syst. Evol. Microbiol. 2005, 55, 1229–1232. [Google Scholar] [CrossRef]
- Hiraishi, A.; Yonemitsu, Y.; Matsushita, M.; Shin, Y.; Kuraishi, H.; Kawahara, K. Characterization of Porphyrobacter sanguineus sp. nov., an aerobic bacteriochlorophyll−containing bacterium capable of degrading biphenyl and dibenzofuran. Arch. Microbiol. 2002, 178, 45–52. [Google Scholar] [CrossRef]
- Zhang, Y.; Carvalho, P.N.; Lv, T.; Arias, C.; Brix, H.; Chen, Z. Microbial density and diversity in constructed wetland systems and the relation to pollutant removal efficiency. Water Sci. Technol. 2016, 73, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Balkwill, D.L.; Aldrich, H.C.; Drake, G.R.; Boone, D.R. Characterization of the anaerobic propionate−degrading syntrophs Smithella propionica gen. nov., sp. nov. and Syntrophobacter wolinii. Int. J. Syst. Bacteriol. 1999, 49, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Iino, T.; Mori, K.; Uchino, Y.; Nakagawa, T.; Harayama, S.; Suzuki, K. Ignavibacterium album gen. nov., sp. nov., a moderately thermophilic anaerobic bacterium isolated from microbial mats at a terrestrial hot spring and proposal of Ignavibacteria classis nov.; for a novel lineage at the periphery of green sulfur bacteria. Int. J. Syst. Evol. Microbiol. 2010, 60, 1376–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Xu, X.; Zhu, L. Structure and function of the microbial consortia of activated sludge in typical municipal wastewater treatment plants in winter. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Beller, H.R.; Chain, P.S.; Letain, T.E.; Chakicherla, A.; Larimer, F.W.; Richardson, P.M.; Coleman, M.A.; Wood, A.P.; Kelly, D.P. The genome sequence of the obligately chemolithoautotrophic, facultatively anaerobic bacterium Thiobacillus denitrificans. J. Bacteriol. 2006, 188, 1473–1488. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.C.; Stegen, J.C. The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 2015, 6, 713. [Google Scholar] [CrossRef] [Green Version]
- Bédard, C.; Knowles, R. Physiology, biochemistry, and specific inhibitors of CH4, NH4+, and CO oxidation by methanotrophs and nitrifiers. Microbiol. Rev. 1989, 53, 68–84. [Google Scholar] [CrossRef]
- Thrash, J.C.; Pollock, J.; Torok, T.; Coates, J.D. Description of the novel perchlorate−reducing bacteria Dechlorobacter hydrogenophilus gen. nov., sp. nov. and Propionivibrio militaris, sp. nov. Appl. Microbiol. Technol. 2010, 86, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Salinero, K.; Keller, K.; Feil, W.S.; Feil, H.; Trong, S.; Di Bartolo, G.; Lapidus, A. Metabolic analysis of the soil microbe Dechloromonas aromatica str. RCB: Indications of a surprisingly complex life−style and cryptic anaerobic pathways for aromatic degradation. BMC Genom. 2009, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Bond, P.L.; Hugenholtz, P.; Keller, J.; Blackall, L.L. Bacterial community structures of phosphate−removing and non−phosphate−removing activated sludges from sequencing batch reactors. Appl. Environ. Microbiol. 1995, 61, 1910–1916. [Google Scholar] [CrossRef] [Green Version]
- Godon, J.J.; Zumstein, E.; Dabert, P.; Habouzit, F.; Moletta, R. Molecular microbial diversity of an anaerobic digestor as determined by small−subunit rDNA sequence analysis. Appl. Environ. Microbiol. 1997, 63, 2802–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Xie, S.; Sun, W.; Li, X.; Cupples, A.M. Identification of a novel toluene−degrading bacterium from the candidate phylum TM7, as determined by DNA stable isotope probing. Appl. Environ. Microbiol. 2009, 75, 4644–4647. [Google Scholar] [CrossRef] [Green Version]
- Alou, M.T.; Ndongo, S.; Frégère, L.; Labas, N.; Andrieu, C.; Richez, M.; Couderc, C.; Baudoin, J.-P.; Abrahão, J.; Brah, S.; et al. Taxonogenomic description of four new Clostridium species isolated from human gut: ‘Clostridium amazonitimonense’, ‘Clostridium merdae’, ‘Clostridium massilidielmoense’ and ‘Clostridium nigeriense’. New Microbes New Infect. 2018, 21, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Shchegolkova, N.M.; Krasnov, G.S.; Belova, A.A.; Dmitriev, A.A.; Kharitonov, S.L.; Klimina, K.M.; Melnikova, N.V.; Kudryavtseva, A.V. Microbial Community Structure of Activated Sludge in Treatment Plants with Different Wastewater Compositions. Front. Microbiol. 2016, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Moir, A.; Cooper, G. Spore germination. In The Bacterial Spore: From Molecules to Systems; Driks, A., Eichenberger, P., Eds.; ASM Press: Washington, DC, USA, 2016; pp. 217–236. [Google Scholar]
- Gerritsen, J.; Hornung, B.; Ritari, J.; Paulin, L.; Rijkers, G.T.; Schaap, P.J.; de Vos, W.M.; Smidt, H. A comparative and functional genomics analysis of the genus Romboutsia provides insight into adaptation to an intestinal lifestyle. BioRxiv 2019, 845511. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.J.; Lacey, J.A. Genomics of the Pathogenic Clostridia. Microbiol. Spectrum. 2019, 7, GPP3-0033-2018. [Google Scholar] [CrossRef] [PubMed]
- Ricaboni, D.; Mailhe, M.; Khelaifia, S.; Raoult, D.; Million, M. Romboutsia timonensis, a new species isolated from human gut. New Microbes New Infect. 2016, 12, 6–7. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of Wastewater | Range of Values during the Year |
|---|---|
| Temperature, °C | 17.0–26.0 |
| O2 (dissolved), mg L−1 | 2.1–8.3 |
| pH | 5.8–10.4 |
| Electrical conductivity, µS cm−1 | 576–4201 |
| NO3−, mg L−1 | 4.97–11.17 |
| NO2−, mg L−1 | 0.095–0.345 |
| NH4+, mg L−1 | 0.05–2.23 |
| Total N , mg L−1 | 1.68–3.94 |
| Fe2+, mg L−1 | 0.016–0.094 |
| S2−, mg L−1 | 0.0013–0.0215 |
| SO42−, mg L−1 | 70.0–90.2 |
| PO43−, mg L−1 | 0.19–1.78 |
| Cl−, mg L−1 | 48.5–50.7 |
| Suspended substances (SS)mg L−1 | 9.0–180.0 |
| Chemical oxygen demand (COD), mg L−1 | 10.6–83.8 |
| Sample ID | Zone Index | Zone | Volume, m3 | Sediment | Oxygen Conditions | Dominant Processes |
|---|---|---|---|---|---|---|
| I-1 | I | Sand traps and grids | 280 | Washed sand | Aerobic | Mechanical retention, sedimentation of coarse suspended substances, organic matter (OM) oxidation; |
| I-2 | Sand | |||||
| II-1 | II | Phytofilters | 2500 | Silty sand | Aerobic | OM oxidation, nitrification; |
| II-2 | Rooted sludge | |||||
| III-1 | III | Sedimentation tank | 3100 | Bottom sludge | Anaerobic | Anaerobic oxidation of OM and sulfides, denitrification, anammox; |
| IV-1 | IV | Phyto-treatment | 580 | Rooted sludge | Aerobic, locally anaerobic | All previous + biomass assimilation, nitrification; |
| IV-2 | Sludge | |||||
| IV-3 | Sludge | |||||
| V-1 | V | Additional phyto-treatment | 730 | Rooted sludge | Aerobic, locally anaerobic | Biomass assimilation, OM oxidation, nitrification |
| V-2 | Rooted sludge |
| Taxon | Average Diameter | Surface Area | Diameter > 1000 µm | Diameter 100–1000 µm | Diameter 10–100 µm | Diameter 1–10 µm | Diameter < 1 µm |
|---|---|---|---|---|---|---|---|
| Phyla | |||||||
| Acidobacteria | −0.41 | 0.46 | −0.21 | −0.37 | 0.33 | 0.41 | 0.49 |
| Bacteroidetes | −0.45 | 0.36 | −0.25 | −0.47 | 0.49 | 0.34 | 0.13 |
| Chloroflexi | −0.60 | 0.66 | −0.30 | −0.65 | 0.61 | 0.64 | 0.42 |
| Firmicutes | −0.46 | 0.18 | −0.64 | −0.33 | 0.39 | 0.15 | 0.02 |
| Proteobacteria | 0.79 | −0.50 | 0.74 | 0.71 | −0.76 | −0.47 | −0.14 |
| Verrucomicrobia | −0.40 | 0.51 | −0.15 | −0.40 | 0.34 | 0.49 | 0.49 |
| Acidobacteria | −0.41 | 0.46 | −0.21 | −0.37 | 0.33 | 0.41 | 0.49 |
| Classes | |||||||
| Acidobacteria Gp17 | −0.37 | 0.55 | −0.14 | −0.38 | 0.29 | 0.54 | 0.54 |
| Acidobacteria Gp18 | −0.27 | 0.44 | 0.00 | −0.30 | 0.23 | 0.41 | 0.49 |
| Acidobacteria Gp23 | −0.31 | 0.65 | −0.02 | −0.37 | 0.24 | 0.64 | 0.76 |
| Acidobacteria Gp3 | −0.22 | 0.44 | 0.05 | −0.20 | 0.11 | 0.39 | 0.65 |
| Acidobacteria Gp6 | −0.48 | 0.64 | −0.23 | −0.50 | 0.42 | 0.61 | 0.60 |
| Acidobacteria Gp7 | −0.33 | 0.60 | 0.00 | −0.35 | 0.24 | 0.56 | 0.76 |
| α−Proteobacteria | 0.98 | −0.75 | 0.87 | 0.93 | −0.94 | −0.75 | −0.24 |
| Anaerolineae | −0.58 | 0.66 | −0.28 | −0.63 | 0.58 | 0.63 | 0.44 |
| Sphingobacteriia | −0.04 | 0.38 | 0.21 | −0.02 | −0.10 | 0.33 | 0.75 |
| Verrucomicrobiae | −0.64 | 0.72 | −0.42 | −0.65 | 0.59 | 0.70 | 0.56 |
| Genera | |||||||
| Altererythrobacter | 0.97 | −0.70 | 0.91 | 0.90 | −0.92 | −0.70 | −0.20 |
| Cloacibacterium | 0.71 | −0.83 | 0.42 | 0.86 | −0.81 | −0.86 | −0.36 |
| Dechloromonas | −0.44 | 0.53 | −0.34 | −0.54 | 0.50 | 0.58 | 0.15 |
| Flavobacterium | 0.65 | −0.54 | 0.60 | 0.75 | −0.77 | −0.58 | 0.05 |
| Gp17 | −0.37 | 0.55 | −0.14 | −0.38 | 0.29 | 0.54 | 0.54 |
| Gp18 | −0.27 | 0.44 | 0.00 | −0.30 | 0.23 | 0.41 | 0.49 |
| Gp6 | −0.48 | 0.64 | −0.23 | −0.50 | 0.42 | 0.61 | 0.60 |
| Gp7 | −0.33 | 0.60 | 0.00 | −0.35 | 0.24 | 0.56 | 0.76 |
| Hydrogenophaga | 0.99 | −0.70 | 0.92 | 0.91 | −0.94 | −0.69 | −0.19 |
| Ignavibacterium | −0.44 | 0.55 | −0.17 | −0.50 | 0.45 | 0.55 | 0.38 |
| Novosphingobium | 0.97 | −0.68 | 0.93 | 0.90 | −0.94 | −0.68 | −0.14 |
| Reyranella | −0.24 | 0.63 | 0.11 | −0.30 | 0.16 | 0.59 | 0.89 |
| Sphingobium | 0.96 | −0.84 | 0.79 | 0.98 | −0.98 | −0.84 | −0.29 |
| Sphingomonas | 0.98 | −0.68 | 0.92 | 0.89 | −0.92 | −0.67 | −0.18 |
| Sphingopyxis | 0.99 | −0.71 | 0.90 | 0.90 | −0.93 | −0.70 | −0.22 |
| Sphingorhabdus | 0.98 | −0.68 | 0.93 | 0.88 | −0.92 | −0.66 | −0.18 |
| Thiobacillus | −0.20 | 0.65 | 0.19 | −0.31 | 0.15 | 0.64 | 0.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, M.V.; Krasnov, G.S.; Rybka, K.Y.; Kharitonov, S.L.; Zavgorodnyaya, Y.A.; Yudina, A.V.; Shchegolkova, N.M. Spatial Changes in Microbial Communities along Different Functional Zones of a Free-Water Surface Wetland. Microorganisms 2020, 8, 1604. https://doi.org/10.3390/microorganisms8101604
Semenov MV, Krasnov GS, Rybka KY, Kharitonov SL, Zavgorodnyaya YA, Yudina AV, Shchegolkova NM. Spatial Changes in Microbial Communities along Different Functional Zones of a Free-Water Surface Wetland. Microorganisms. 2020; 8(10):1604. https://doi.org/10.3390/microorganisms8101604
Chicago/Turabian StyleSemenov, Mikhail V., George S. Krasnov, Ksenia Y. Rybka, Sergey L. Kharitonov, Yulia A. Zavgorodnyaya, Anna V. Yudina, and Nataliya M. Shchegolkova. 2020. "Spatial Changes in Microbial Communities along Different Functional Zones of a Free-Water Surface Wetland" Microorganisms 8, no. 10: 1604. https://doi.org/10.3390/microorganisms8101604