QTL Mapping for Domestication-Related Characteristics in Field Cress (Lepidium campestre)—A Novel Oil Crop for the Subarctic Region




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Target Characteristics
2.2. Planting, DNA Isolation, and Genotyping
2.3. Phenotyping of Mapping Population
2.4. Phenotypic Data Analysis
2.5. SNP Data and QTL Analysis
2.6. Comparative Genomics/Candidate Gene Identification
2.7. SNP Genotypes Versus Trait Variation
3. Results
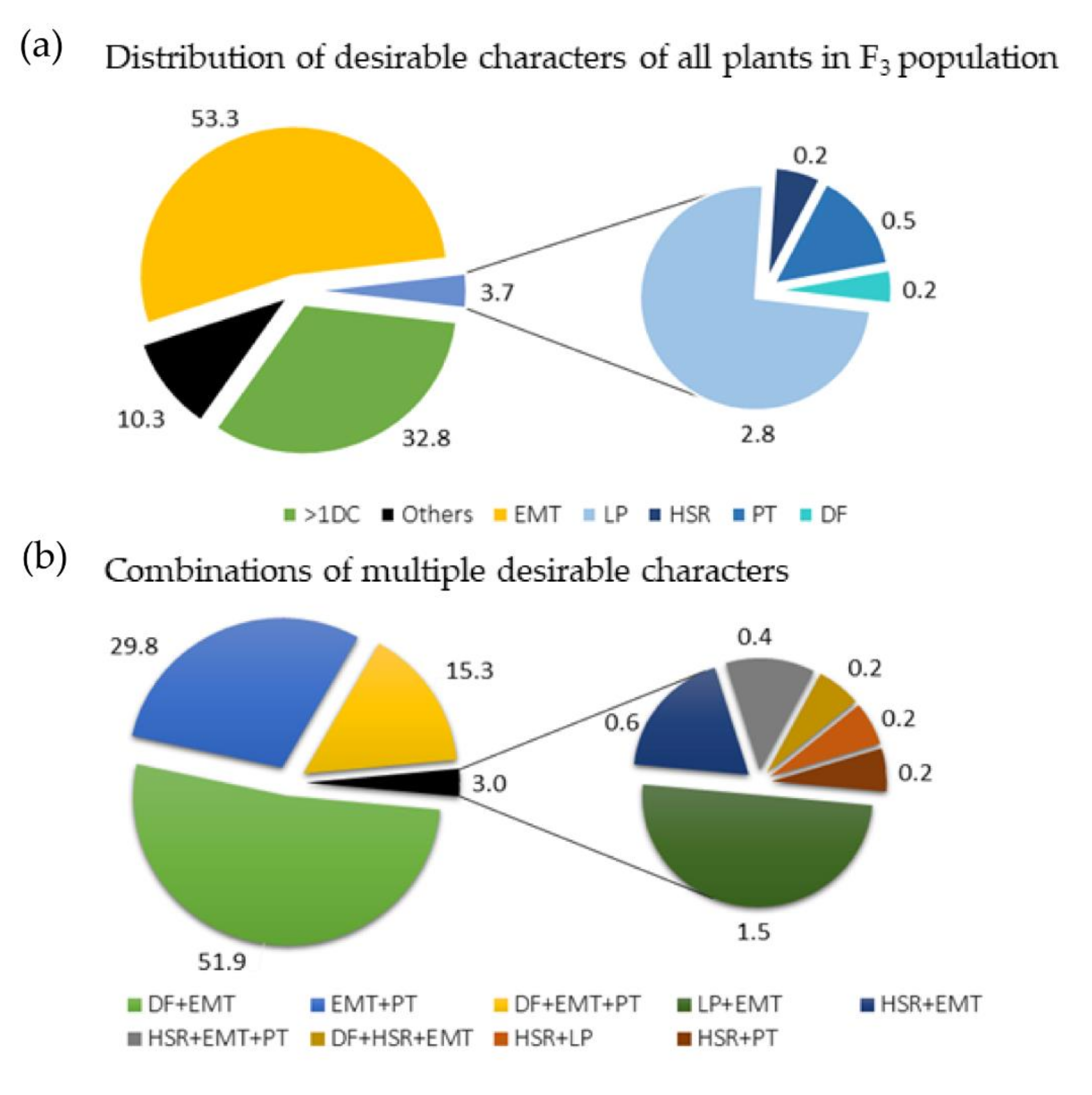
3.1. Analysis of Phenotypic Data of F2 and F3 Populations
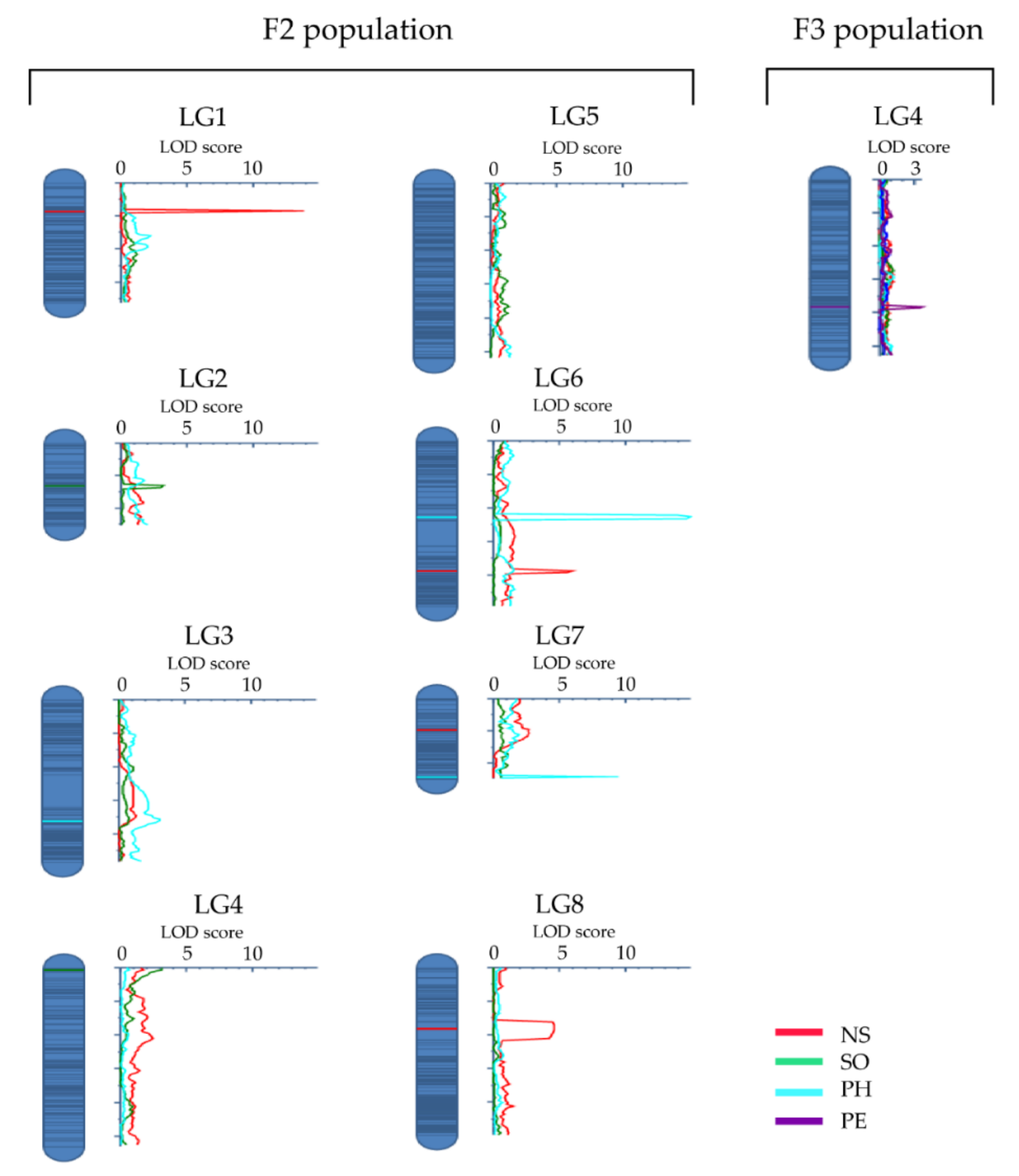
3.2. QTL Analysis
3.3. Comparative Analysis of the Genomic Regions Harboring the QTLs Using A. thaliana Genome
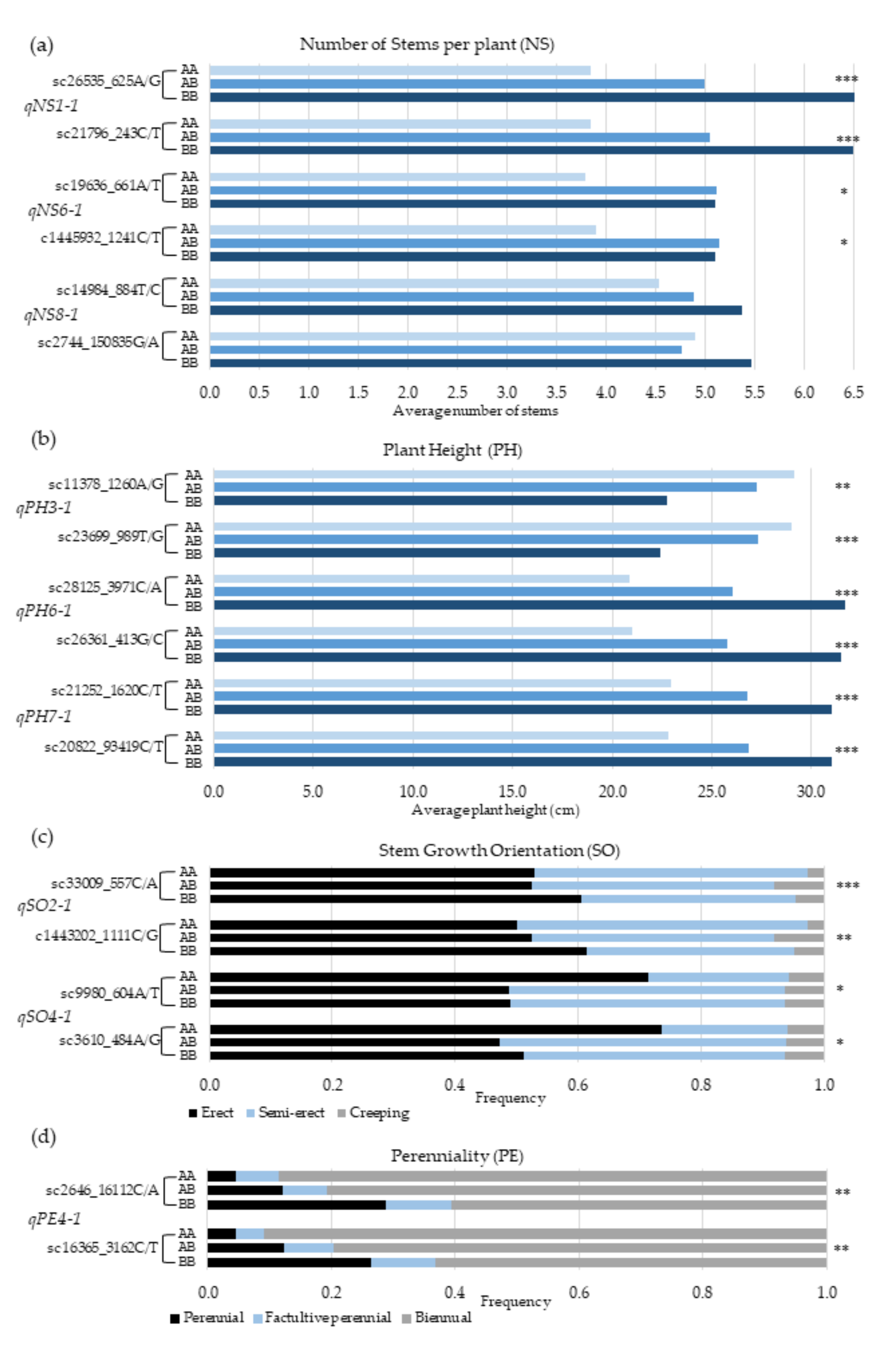
3.4. SNP Genotypes Versus Trait Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carlsson, A.S.; Yilmaz, J.L.; Green, A.G.; Stymne, S.; Hofvander, P. Replacing fossil oil with fresh oil—With what and for what? Eur. J. Lipid. Sci. Technol. 2011, 113, 812–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD-FAO. Agricultural Outlook 2019–2028; OECD Publishing: Paris, France, 2019; pp. 142–153. [Google Scholar]
- Fróna, D.; Szenderák, J.; Harangi-Rákos, M. The Challenge of Feeding the World. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef] [Green Version]
- Cox, T.S.; Glover, J.D.; Van Tassel, D.L.; Cox, C.M.; DeHaan, L.R. Prospects for developing perennial-grain crops. Bioscience 2006, 56, 649–659. [Google Scholar] [CrossRef]
- Glover, J.D.; Reganold, J.P.; Bell, L.W.; Borevitz, J.; Brummer, E.C.; Buckler, E.S.; Cox, C.M.; Cox, T.S.; Crews, T.E.; Culman, S.W.; et al. Agriculture. Increased food and ecosystem security via perennial grains. Science 2010, 328, 1638–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, C.; Willforss, J.; Lopes-Pinto, F.; Ortiz, R.; Geleta, M. Identification of genes regulating traits targeted for domestication of field cress (Lepidium campestre) as a biennial and perennial oilseed crop. BMC Genet. 2018, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Geleta, M.; Zhu, L.-H.; Stymne, S.; Lehrman, A. Domestication of Lepidium campestre as part of Mistra Biotech, a research programme focused on agro biotechnology for sustainable food. In Perennial Crops for Food Security, Proceedings of the FAO Expert Workshop, Rome, Italy, 28–30 August 2013; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; pp. 141–147. [Google Scholar]
- Adams, S.R.; Langton, F.A. Photoperiod and plant growth: A review. J. Hortic. Sci. Biotechnol. 2005, 80, 2–10. [Google Scholar] [CrossRef]
- Diepenbrock, W. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crop Res. 2000, 67, 35–49. [Google Scholar] [CrossRef]
- Kuroda, T.; Saitoh, K.; Mahmood, T.; Yanagawa, K. Differences in Flowering Habit between Determinate and Indeterminate Types of Soybean. Plant Prod. Sci. 1998, 1, 18–24. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, J.; Wang, X.; Liu, G.; Wang, H. Genetic architecture and mechanism of seed number per pod in rapeseed: Elucidated through linkage and near-isogenic line analysis. Sci. Rep. 2016, 6, 24124. [Google Scholar] [CrossRef] [Green Version]
- Salas Fernandez, M.G.; Becraft, P.W.; Yin, Y.; Lubberstedt, T. From dwarves to giants? Plant height manipulation for biomass yield. Trends Plant Sci. 2009, 14, 454–461. [Google Scholar] [CrossRef]
- Merker, A.; Eriksson, D.; Bertholdsson, N.-O. Barley yield increases with undersown Lepidium campestre. Acta Agric. Scand. Sect. B Soil Plant Sci. 2010, 60. [Google Scholar] [CrossRef]
- Ortiz, R.; Geleta, M.; Gustafsson, C.; Lager, I.; Hofvander, P.; Lofstedt, C.; Cahoon, E.B.; Minina, E.; Bozhkov, P.; Stymne, S. Oil crops for the future. Curr. Opin. Plant Biol. 2020. [Google Scholar] [CrossRef]
- Lee, J.Y.; Mummenhoff, K.; Bowman, J.L. Allopolyploidization and evolution of species with reduced floral structures in Lepidium L. (Brassicaceae). Proc. Natl. Acad. Sci. USA 2002, 99, 16835–16840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geleta, M.; Gustafsson, C.; Glaubitz, J.C.; Ortiz, R. High-Density Genetic Linkage Mapping of Lepidium Based on Genotyping-by-Sequencing SNPs and Segregating Contig Tag Haplotypes. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.H.; Zhang, L.Y.; Wang, J.K. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ribaut, J.M.; Li, Z.; Wang, J. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations. Theor. Appl. Genet. 2008, 116, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.P.; Simon, E.J.; Trotochaud, A.E.; Clark, S.E. POLTERGEIST functions to regulate meristem development downstream of the CLAVATA loci. Development 2000, 127, 1661–1670. [Google Scholar]
- Goda, H.; Sawa, S.; Asami, T.; Fujioka, S.; Shimada, Y.; Yoshida, S. Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2004, 134, 1555–1573. [Google Scholar] [CrossRef] [Green Version]
- Econopouly, B.F.; Van Veldhuizen, B.M.; Lyon, S.R.; Killilea, D.W.; Zhang, M.C.; Jones, S.S. Early maturing spring wheat in Nordic wildtype NAM-B1 germplasm for short-duration alternative wheat-producing regions. Plant Genet. Resour. C 2019, 17, 352–361. [Google Scholar] [CrossRef]
- Makinde, O.M.; Makinde, B.O. Phenotypic Analysis of Seed Yield and Yield Components in Cowpea (Vigna unguiculata; L.). Plant Breed. Biotech. 2016, 4, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Uva, R.H.; Neal, J.C.; DiTomaso, J.M. Weeds of the Northeast; Comstock Pub. Associates: Ithaca, NY, USA, 1997. [Google Scholar]
- Günster, A. Variability in life history parameters of four serotinous plants in the Namib Desert. Vegetatio 1994, 114, 149–160. [Google Scholar]
- Friedman, J.; Rubin, M.J. All in good time: Understanding annual and perennial strategies in plants. Am. J. Bot. 2015, 102, 497–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vico, G.; Manzoni, S.; Nkurunziza, L.; Murphy, K.; Weih, M. Trade-offs between seed output and life span—A quantitative comparison of traits between annual and perennial congeneric species. New Phytol. 2016, 209, 104–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeHaan, L.; Christians, M.; Crain, J.; Poland, J. Development and Evolution of an Intermediate Wheatgrass Domestication Program. Sustainability 2018, 10, 1499. [Google Scholar] [CrossRef] [Green Version]
- Glover, J. The necessity and possibility of perennial grain production systems. Renew. Agric. Food Syst. 2005, 20, 1–4. [Google Scholar] [CrossRef]
- Kauschmann, A.; Jessop, A.; Koncz, C.; Szekeres, M.; Willmitzer, L.; Altmann, T. Genetic evidence for an essential role of brassinosteroids in plant development. Plant J. 1996, 9, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Clouse, S.D. Molecular genetic studies confirm the role of brassinosteroids in plant growth and development. Plant J. 1996, 10, 1–8. [Google Scholar] [CrossRef]
- Song, S.K.; Lee, M.M.; Clark, S.E. POL and PLL1 phosphatases are CLAVATA1 signaling intermediates required for Arabidopsis shoot and floral stem cells. Development 2006, 133, 4691–4698. [Google Scholar] [CrossRef] [Green Version]
- Baurle, I.; Laux, T. Regulation of WUSCHEL transcription in the stem cell niche of the Arabidopsis shoot meristem. Plant Cell 2005, 17, 2271–2280. [Google Scholar] [CrossRef] [Green Version]
- Brand, U.; Grunewald, M.; Hobe, M.; Simon, R. Regulation of CLV3 expression by two homeobox genes in Arabidopsis. Plant Physiol. 2002, 129, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Description | Measurement/Count/Category | Percentage of Each Category as Measured in F2 or F3 Population |
|---|---|---|---|
| PH a | Height of each plant measured in cm at maturity | measurement in cm | NA |
| NS b | Number of basal branches (tillers) of each plant at maturity | count | NA |
| SO | Growth orientation of the central stem with reference to the surface | 1 = erect (ca. 90°) 2 = semi-erect (≥45°) 3 = creeping (<10°) | 53.1 38.5 8 |
| FH | The ability of plants to continue flowering after the original group of inflorescence matures | 1 = determinate 2 = moderately indeterminate 3 = indeterminate | 22 34 43 |
| ER | Determined based on number of days from end of winter to plant maturity | 1 = early (90–100 days) 2 = late (115–125 days) | 86 14 |
| SY | Determined based on the weight of seeds harvested from each plant | 1 = low (<5 gm) 2 = medium (5–10 gm) 3 = medium-high (11–20 gm) 4 = high (>20 gm) | 52.9 35.7 8.1 3.3 |
| SHR c | Determined based on the percentage of seeds lost at full maturity after exposure to heavy rain and strong wind | 1 = high resistance (<10% seed loss) 2 = moderate resistance (10–30% seed loss) 3 = susceptible (>30% of seed loss) | 0.7 12.3 87 |
| PE | The ability of the plants to ratoon and growth to full maturity after first harvest | 1 = perennial ** 2 = facultative perennial *** 3 = biennial | 15.6 13.9 70.5 |
| Characteristic | Category | r | p-Value |
|---|---|---|---|
| FH | determinate | 0.24 | <0.001 |
| moderately indeterminate | 0.06 | 0.340 | |
| indeterminate | 0.27 | <0.001 | |
| combined | 0.23 | <0.001 | |
| ER | early | 0.31 | <0.001 |
| late | 0.31 | <0.001 | |
| combined | 0.70 | <0.001 | |
| SY | high | 0.31 | <0.001 |
| medium-high | 0.19 | 0.001 | |
| medium | 0.16 | 0.006 | |
| low | 0.26 | <0.001 | |
| combined | 0.47 | <0.001 | |
| SHR | high resistance | 0.01 | 0.808 |
| moderate resistance | 0.25 | <0.001 | |
| susceptible | 0.25 | <0.001 | |
| combined | 0.80 | <0.001 | |
| PE | perennial | 0.13 | 0.037 |
| facultative perennial | 0.13 | 0.026 | |
| biennial | 0.26 | <0.001 | |
| combined | 0.56 | <0.001 |
| PH | NS | SO | FH | SY | SHR | ER | |
|---|---|---|---|---|---|---|---|
| NS | 0.26 a*** | ||||||
| SO | −0.15 a** | 0.02 a | |||||
| FH | 0.12 b* | 0.02 b | 0.02 b | ||||
| SY | 0.11 b | 0.004 b | 0.02 b | −0.34 c*** | |||
| SHR | 0.09 b | 0.09 b | 0.05 b | −0.04 c | 0.18 c** | ||
| ER | 0.05 b | 0.06 b | 0.02 b | 0.37 c*** | −0.66 c*** | −0.26 c*** | |
| PE | 0.02 | 0.09 b | 0.03 b | 0.14 c* | −0.42 c*** | −0.07 c | 0.25 c*** |
| Trait | QTL a | LG | Position (cM) | QTL Region (cM) | Flanking Marker Loci b | LOD | PVE (%) | AtChr c | Start (nt) | End (nt) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Left | Right | ||||||||||
| NS | qNS1-1 | 1 | 17.00 | 16.13–17.93 | sc26535_625A/G | sc21796_243C/T | 13.04 | 11.81 | 1 | 6630234 | 6816262 |
| SO | qSO2-1 | 2 | 26.00 | 25.80–27.17 | sc33009_557C/A | c1443202_1111C/G | 3.21 | 1.93 | NA | NA | NA |
| PH | qPH3-1 | 3 | 72.00 | 71.33–72.85 | sc11378_1260A/G | sc23699_989T/G | 3.04 | 3.16 | 3 | 4211673 | 4582690 |
| SO | qSO4-1 | 4 | 1.00 | 0.00–1.38 | sc9980_604A/T | sc3610_484A/G | 3.16 | 1.84 | 2 | 19096856 | 19679860 |
| PE | qPE4-1 | 4 | 76.00 | 75.12–77.94 | sc2646_16112C/A | sc16365_3162C/T | 3.64 | 7.23 | NA | NA | NA |
| PH | qPH6-1 | 6 | 45.00 | 44.86–46.38 | sc28125_3971C/A | sc26361_413G/C | 14.89 | 16.94 | 5 | 20689846 | 20845218 |
| NS | qNS6-1 | 6 | 77.00 | 76.48–78.14 | sc19636_661A/T | c1445932_1241C/T | 5.95 | 5.06 | 5 | 23044365 | 23311700 |
| PH | qPH7-1 | 7 | 48.00 | 47.89–48.08 | sc21252_1620C/T | sc20822_9341C/T | 8.66 | 9.39 | 5 | 15556187 | 15708117 |
| NS | qNS8-1 | 8 | 36.00 | 31.95–42.77 | sc14984_884T/C | sc2744_150835G/A | 4.63 | 4.42 | 5 | 9100466 | 24634605 |
| Phenotypic Class | Geno-Type | Characteristic | Number of Stems Per Plant (NS) | p Value | ||
|---|---|---|---|---|---|---|
| Marker | sc26535_625A/G | sc19636_661A/T | sc14984_884T/C | |||
| QTL | qNS1-1 | qNS6-1 | qNS8-1 | |||
| ≥10 stems | AA | 13 | 4 | 17 | <0.01 | |
| BB | 46 | 29 | 25 | |||
| Single stem | AA | 56 | 28 | 28 | <0.05 | |
| BB | 8 | 13 | 18 | |||
| Characteristic | Plant Height (PH) | |||||
| QTL | qPH3-1 | qPH6-1 | qPH7-1 | |||
| Marker | sc11378_1260A/G | sc28125_3971C/A | sc21252_1620C/T | |||
| >30cm | AA | 27 | 3 | 9 | <0.05 | |
| BB | 15 | 48 | 34 | |||
| <20 cm | AA | 20 | 38 | 44 | <0.01 | |
| BB | 39 | 11 | 11 | |||
| Characteristic | Stem growth orientation (SO) | |||||
| QTL | qSO2-1 | qSO4-1 | ||||
| Marker | sc33009_557C/A | sc9980_604A/T | ||||
| Erect | AA | 15 | 37 | <0.01 | ||
| BB | 38 | 18 | ||||
| Creeping | AA | 20 | 33 | <0.01 | ||
| BB | 27 | 17 | ||||
| Characteristic | Perenniality (PE) | |||||
| QTL | qPE4-1 | |||||
| Marker | sc2646_16112C/A | |||||
| Perennial | AA | 14 | <0.01 | |||
| BB | 34 | |||||
| Biennial | AA | 19 | <0.05 | |||
| BB | 24 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammenhag, C.; Saripella, G.V.; Ortiz, R.; Geleta, M. QTL Mapping for Domestication-Related Characteristics in Field Cress (Lepidium campestre)—A Novel Oil Crop for the Subarctic Region. Genes 2020, 11, 1223. https://doi.org/10.3390/genes11101223
Hammenhag C, Saripella GV, Ortiz R, Geleta M. QTL Mapping for Domestication-Related Characteristics in Field Cress (Lepidium campestre)—A Novel Oil Crop for the Subarctic Region. Genes. 2020; 11(10):1223. https://doi.org/10.3390/genes11101223
Chicago/Turabian StyleHammenhag, Cecilia, Ganapathi Varma Saripella, Rodomiro Ortiz, and Mulatu Geleta. 2020. "QTL Mapping for Domestication-Related Characteristics in Field Cress (Lepidium campestre)—A Novel Oil Crop for the Subarctic Region" Genes 11, no. 10: 1223. https://doi.org/10.3390/genes11101223