Tularemia Goes West: Epidemiology of an Emerging Infection in Austria


1
Department of Internal Medicine II, Infectious Diseases, Immunology, Rheumatology, Pneumology, Medical University of Innsbruck, 6020 Innsbruck, Austria
2
Institute for Veterinary Disease Control, Austrian Agency for Health and Food Safety (AGES), 2340 Mödling, Austria
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(10), 1597; https://doi.org/10.3390/microorganisms8101597
Submission received: 31 August 2020
/
Revised: 12 October 2020
/
Accepted: 13 October 2020
/
Published: 16 October 2020
(This article belongs to the Special Issue Epidemiology of Tularemia and Francisella tularensis)
{kind=link}
{kind=link}
Abstract
:The zoonotic disease tularemia is caused by the Gram-negative bacterium Francisella tularensis, with the two major subspecies tularensis and holarctica being responsible for infections in humans and animals. The F. tularensis subspecies holarctica is less virulent and prevalent in Europe and Asia. Over the last few centuries, few epidemic outbreaks and low numbers of infections have been registered in the eastern part of Austria, specifically in the provinces of Lower Austria, Burgenland, and Styria. The reported infections were mostly associated with hunting hares and the skinning of carcasses. Within the last decade, ticks have been identified as important vectors in Tyrol and served as first evidence for the spread of F. tularensis to Western Austria. In 2018, the pathogen was detected in hares in the provinces of Tyrol, Vorarlberg, and Salzburg. We presume that F. tularensis is now established in most regions of Austria, and that the investigation of potential host and vector animals should be spotlighted by public institutions. Tularemia in humans presents with various clinical manifestations. As glandular, ulceroglandular, and typhoidal forms occur in Austria, this infectious disease should be considered as a differential diagnosis of unknown fever.
1. Introduction
Tularemia is a zoonotic disease caused by Francisella tularensis (F. tularensis), a Gram-negative, facultative intracellular bacterium, which was first isolated from ground squirrels in Tulare County, (California, United States) by McCoy and Chapin in 1911 [1,2]. The first bacteriologically confirmed case was described by Wherry and Lamb in Ohio in 1914 [3].
The pathogen F. tularensis is currently divided into four subspecies: Francisella tularensis subsp. tularensis, Francisella tularensis subsp. holarctica, Francisella tularensis subsp. mediasiatica, and Francisella tularensis subsp. novicida [4]. Initially, F. tularensis subsp. novicida was classified as a distinct species, F. novicida, because of phenotypic differences and less fastidious growth requirements compared to F. tularensis [5]. In the 1980s, based on DNA–DNA hybridization experiments, it was then suggested to reassign F. novicida as a subspecies to F. tularensis because of a high degree of genetic relatedness [6]. To date, there has not been a clear decision on the correct nomenclature and both names are still in use [7].
The subspecies tularensis (type A) and holarctica (type B, former subsp. palaearctica) are the main causative agents for tularemia in humans and animals. The less virulent type B occurs throughout the Northern hemisphere, with predominance in Europe and Asia, and is further divided into biovar I (erythromycin sensitive, prevalent in Western Europe), biovar II (erythromycin resistant, prevalent in Northern and Eastern Europe), and biovar japonica (prevalent in Japan, China, and Turkey) [8,9,10,11]. In contrast to F. tularensis subsp. holarctica biovar I and II, biovar japonica, like F. tularensis subsp. tularensis, produces acid, not only in glucose-containing media, but also in glycerol-containing media. By amplifying variable-number tandem repeats (VNTRs), F. tularensis subsp. holarctica was first separated into five major clades (B.I, B.II, B.III, B.IV, and B.V) [12]. Later on, assays based on a canonical single nucleotide polymorphism (canSNP) and canonical insertions/deletions (INDELs), which circumvented the previous classification’s disadvantage for phylogenetic studies, led to a reduction to the four major clades: B.12 (B.I), corresponding to biovar II Eastern and Central European strains; B.4 (B.II), corresponding to North American strains; B.6 (B.IV), corresponding to biovar I Western European strains; and B.16 (B.V), corresponding to strains belonging to biovar japonica [10,13,14,15,16,17,18]. Recent reports of tularemia cases in humans and ring possums in Australia caused by F. tularensis subsp. holarctica have confirmed the presence of the pathogen also in the Southern hemisphere [19,20].
The highly virulent type A appears to be restricted to North America, and causes almost 70% of human cases on this continent [1,9,21]. Although some strains of F. tularensis subsp. tularensis were isolated from mice and fleas in the Danube region between 1978 and 1996, human tularemia infections in Europe have only been caused by F. tularensis subsp. holarctica so far [22,23]. The obtained type A isolates were later assigned to a laboratory strain by genome sequencing, and it was assumed that anthropogenic activities, such as the disposal of laboratory waste or the escape of infected animals, had led to an environmental contamination [24].
Infections with F. tularensis have been reported in a wide range of vertebrates, amphibians, fish, and invertebrates [25,26]. Furthermore, isolation of F. tularensis DNA in water and sediment indicates silent persistence in the environment, likely originating from dead affected animals or their excrements [23,27]. It has been experimentally proven that contaminated silt may remain infectious for up to two months [28]. According to the highly complex ecology of F. tularensis, transmission to humans may occur via different routes, such as direct contact with infected animals (e.g., during animal processing, through ingestion of uncooked meat, and animal bites), arthropod-borne (ticks, horseflies, and mosquitos) and through the consumption of contaminated water or inhalation of contaminated soil, as it may occur during farming works [23,29,30,31,32].
The European hare (Lepus europaeus), also known as the brown or field hare, is considered an important host of F. tularensis and a common vector for the pathogen’s transmission to humans in Europe [26,27,33]. In this species, variable clinical courses ranging from acute deadly septicema to protracted courses with only subacute lesions in various organs are reported. In case of chronic infection, European hares may serve as long-term reservoirs for F. tularensis, thus bearing a persistent risk of transmission to humans, either directly or via vectors [34,35,36]. Histopathological examinations demonstrated differences in the pathogenicity of clade B.FTNF002-00 (subgroup of B.6 and specific for Western Europe) and clade B.13 (subgroup of B.12 and specific for Central and Eastern Europe) in European brown hares [18,37]. While infections with strains of clade B.13 were reported to be associated with polyserositis, affecting the kidneys, pleura, and pericardium, histopathological findings in hares infected with B.FTNF002-00 have been almost invariably characterized by splenitis and hepatitis [34,37]. These results are in accordance with further observations in experimentally infected rats, showing significant differences in weight loss, mortality rate, and time to recovery between the two genotypes [38].
Rodents are very susceptible to F. tularensis, and commonly present with severe infection, leading to early death [39]. Nevertheless, experimental studies showed that infected voles may also show a protracted course of disease with chronic nephritis and bacteriuria, and could therefore also serve as a prolonged source of environmental contamination [40]. In the case of an epizootic event, water, soil, and foodstuffs may be contaminated through carcasses, faeces or urine from infected individuals [28,41]. Several large outbreaks of human tularemia have been reported due to ingestion of contaminated water, some of them supposedly associated with infected rodents, while others showed a demonstrable link [42,43,44,45]. Moreover, in Spain, a large pneumonic outbreak was associated with farming activities due to the inhalation of contaminated aerosols, as well as direct contact to common voles (Microtus arvalis) [46,47].
Moreover, ticks appear to play a key role in the ecology of tularemia among arthropods, as they may carry pathogens over several years and life stages, thus maintaining enzootic tularemia foci between epizootic periods [27,48,49,50]. The genus Dermacentor reticulatus seems to be the most frequent carrier of F. tularensis in Central Europe [51,52,53].
In Sweden, mosquito-borne infections, which are related to the aquatic life cycle, are a common route for transmission of tularemia in humans and can lead to major outbreaks [54,55,56]. Studies have also provided evidence that mosquito larvae may be infected with F. tularensis via water, possibly by ingesting predatory protozoa [57].
Defined roles for the different potential host species in the ecology of tularemia are not yet sufficiently clarified, and may depend on geographical factors, as well as on the susceptibility and sensitivity of respective organisms to the pathogen [58]. Although recent observations provide evidence for differences in virulence among the specific lineages of F. tularensis subsp. holarctica, there is no clearly defined preference of certain lineages for particular host species [59]. It is assumed that most of the animals do not serve as amplifying hosts, increasing the basic reproduction number of F. tularensis, but are “incidental” dead-end hosts, and therefore may not play a role in the persistence and spread of the pathogen [58].
Transmission of tularemia from domestic animals to humans has not yet been documented in Europe, but reports of cat- and dog-related tularemia in humans exist in the United States. There, in a nationwide case study on human tularemia between 2006 and 2016, about 3% of infections were classified as canine-transmitted [60,61]. Furthermore, in a recent study, almost 50% of human tularemia cases in the United States were reported to be cat-associated [62]. Occasional outbreaks were observed in sheep during the lambing season, due to their feeble physical condition [49]. To date, there is no evidence for human-to-human transmission in Europe. However, due to its high virulence and its feasibility to be spread via aerosols, F. tularensis is classified as a category A potential agent for bioterrorism [23,63].
In humans, following a short incubation time of 3 to 5 days (maximum 2 to 3 weeks), tularemia may present with unspecific, flu-like symptoms, including fever, lymphadenopathy, headache, chills, myalgia, and arthralgia, and is therefore often misdiagnosed, especially in areas with low incidence [63,64]. The clinical presentation depends on the location of bacterial entry into the body. The six major clinical manifestations are comprised of ulceroglandular, glandular, oropharyngeal, oculoglandular, pneumonic, and typhoid forms. Ulceroglandular and glandular tularemia are acquired by direct contact with infected animals or are vector-borne, and are the most frequent clinical manifestations in European countries, covering more than 95% of human cases [23,63]. The pneumonic form is mostly related to farming activities, and is caused by the inhalation of aerosols, originating from carcasses of rodents and lagomorphs or infected dead animals [65].
The diagnosis of tularemia is confirmed via serological, PCR, and antigen testing, as well as from culturing [63]. Concerning therapy, aminoglycosides, tetracyclines, and fluorochinolones are the antimicrobial drugs of choice, due to well-established clinical efficacy and the lack of reports on resistance [66,67,68]. For patients with severe infection, aminoglycosides are the recommended therapy because of high cure and minimal relapse rates [69]. Due to the potential of F. tularensis as a bioterrorism agent, in recent decades increased research has been conducted to develop an efficient and safe vaccine. Among previous approaches, the live attenuated vaccine “Live Vaccine Strain” (LVS) was particularly promising, but has shown insufficient protection against strains of type A and has not been licensed in the United States or Europe, due to safety concerns regarding toxicity. Currently, promising research focusses on new genetically modified, live-attenuated vaccine strains and subunit vaccines with improved safety [70,71,72].
Tularemia is a notifiable disease in most European countries, meaning that both confirmed infection and death, as well as suspected cases, must be reported to the respective authorities. While Cyprus, Greece, Iceland, Ireland, Luxembourg, Malta, and the United Kingdom (only imported cases) are considered to be free of tularemia, the disease occurs in the other European countries with varying incidence [73,74]. Sweden and Norway recorded 40% in 2017 and 45% in 2018 of all reported cases of human tularemia across the member states of the European Union (EU) and the European Economic Area (EAA) [74,75].
2. Aims and Methods
The aim of this review is to focus on the epidemiology of tularemia in Austria using available data from the past and present. In addition to a literature search in the databases PubMed, VetMed, and UpToDate with a selection of reviews, case reports, prevalence studies, and guidelines, the annual statistics on human tularemia cases from 1990–2019, available from the Federal Ministry of Labour, Social Affairs, Health and Consumer Protection were included. The research also covered the website of the Austrian Agency for Health and Food Safety GmbH (AGES). Informed consent for publication of the cases and images was obtained from the Tyrolean patients.
3. Results
In Austria, direct or indirect contact with infected animals and tick bites represent the main transmission modes of tularemia to humans. In 2014, a nationwide cross-sectional serological analysis of 526 healthy adults from Austria found that 0.5% had detectable antibodies against F. tularensis [76]. Tularemia sporadically occurs in all counties of Austria, and until 2015 most cases were reported from the provinces of Lower Austria, Burgenland, Vienna, Styria, and Upper Austria, consequently regarded as endemic areas [23]. According to the predominant modes of transmission, the most frequent clinical manifestations of human tularemia in Austria include the ulceroglandular and the glandular forms. However, one case of a typhoid form with secondary pulmonary tularemia was recently reported in a forester from Lower Austria. The suspected route of infection in this case was the inhalation of contaminated dust during forest work [77]. Strains of F. tularensis detected in Austria are commonly assigned to the phylogenetic group B.13 (subclade of major clade B.12), corresponding to biovar II [10]. However, recently a strain belonging to biovar I was isolated from a person suffering from tularemia in the province of Tyrol, Western Austria [78].
3.1. Human Cases
3.1.1. Human Cases: Past
The first serologically confirmed human case of tularemia in Austria affected a woman in Mistelbach (Lower Austria) in 1934 [79]. An accumulation of human infections was observed from late November 1936 to early January 1937. Of approximately 200 affected persons in the districts of Mistelbach and Gänserndorf (Lower Austria, region north and north east of Vienna), serological confirmation was successfully performed in 94 cases. Transmission occurred during the skinning and disemboweling of hares [79]. A second cluster of human diseases was described between November 1945 and April 1946 in the same district. Human cases were also reported alongside the rivers Danube, March, and Thaya, comprising the northeastern regions of Lower Austria down near Vienna, which has remained the “natural focus” of tularemia in Austria. One year later, in 1947, sporadic human cases south of the Danube were documented. A major human tularemia epidemic occurred in 1959/1960 in sugar factories in Lower Austria, south of the Danube and close to the province of Burgenland. Following the inhalation of contaminated aerosols formed by dead, infected rodents entering the beet-washing process, a total of 577 human cases were recorded [80]. Further outbreaks of infections were observed after a strong proliferation of field mice (Microtus arvalis) promoted by favorable weather conditions like mild winter and hot summer temperatures [81]. Another major epidemic occurred in the late 1960s, with 170 confirmed cases of human tularemia.
3.1.2. Human Cases: Present
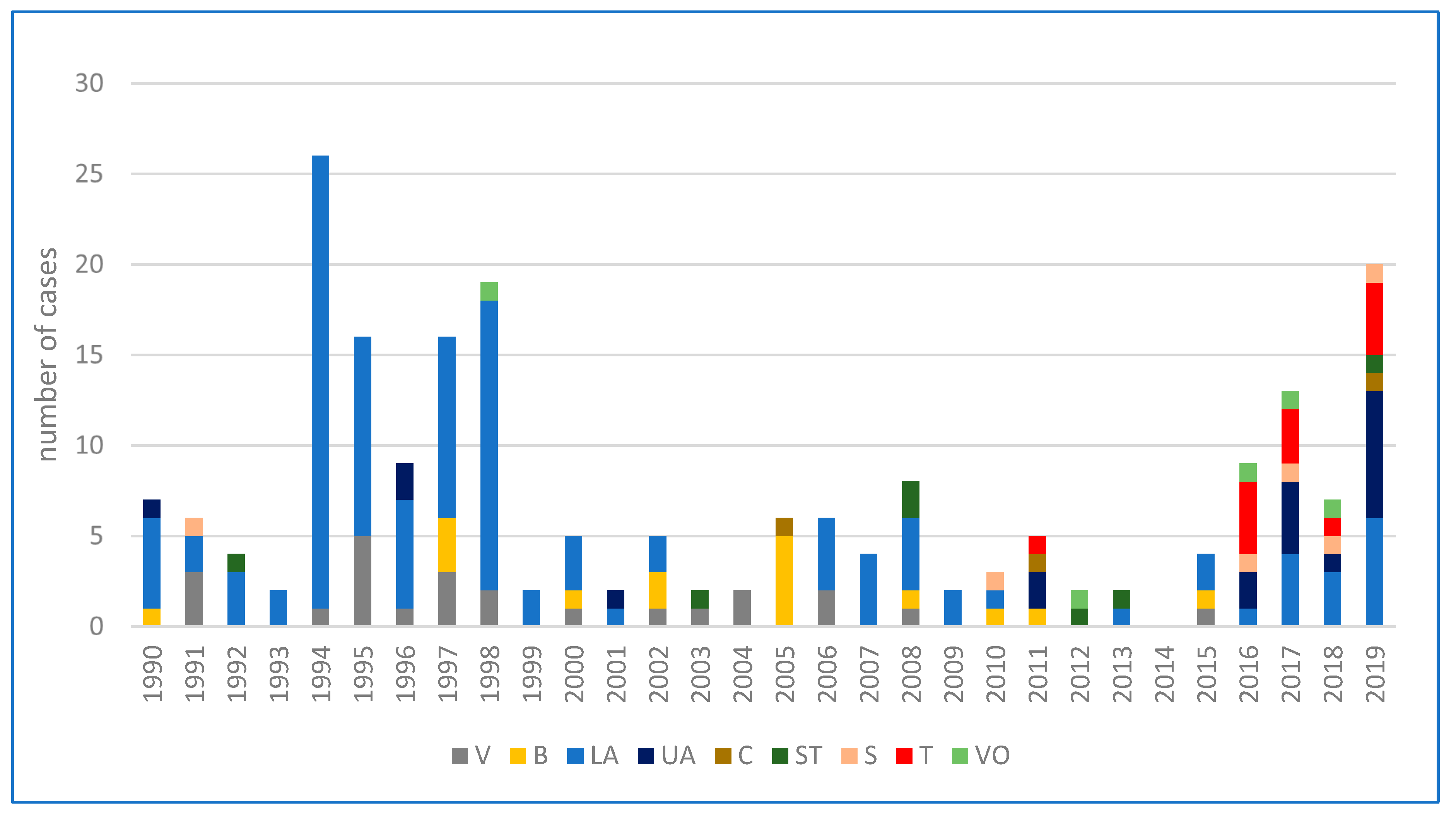
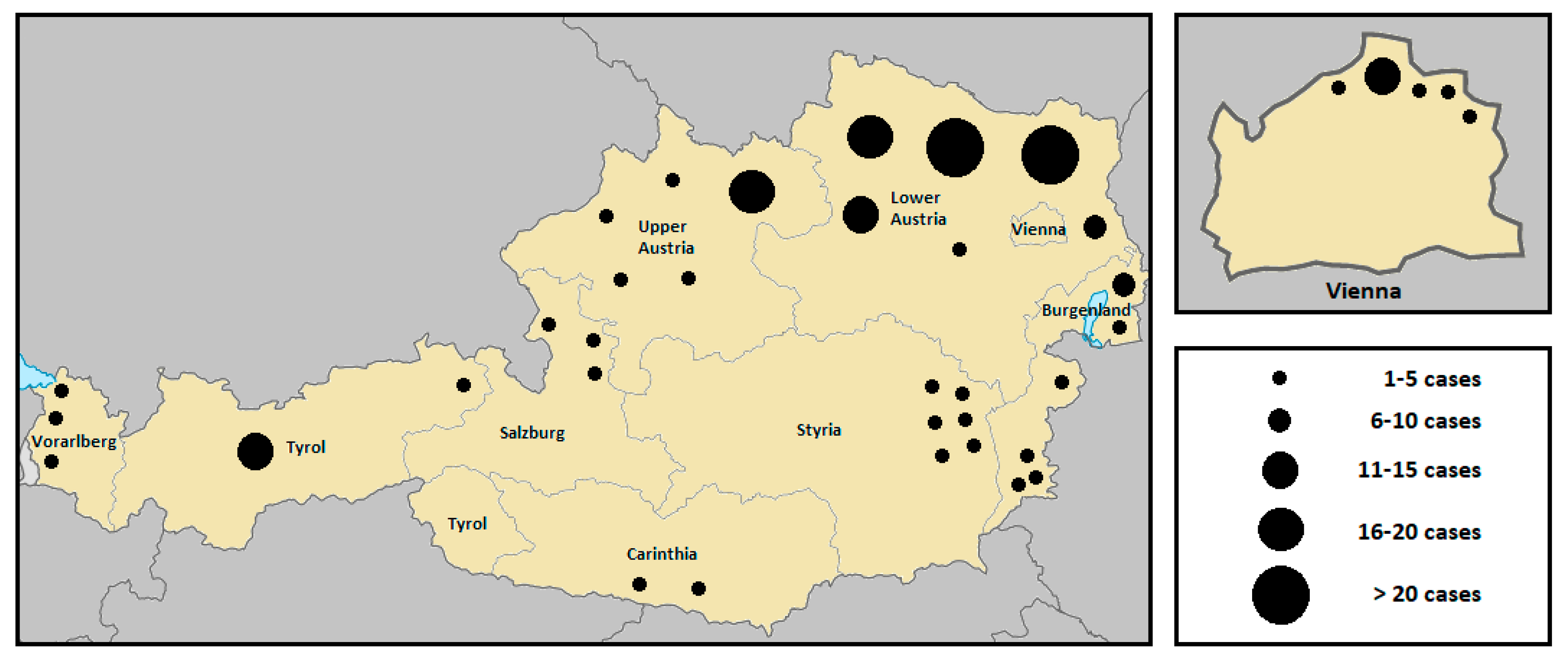
According to the Federal Ministry of Labour, Social Affairs, Health and Consumer Protection, a total of 214 human infections with tularemia were reported in Austria over the last thirty years. Subject to a wide range of variation (from 0 to 26), the annual average incidence was 7 cases per year [82]. While the number of human infections was continuously decreasing during the years 1990 to 1993, an accumulation was then observed in 1994 (26 cases), 1995 (16 cases), 1997 (16 cases), and 1998 (19 cases), with the largest proportion occurring in the endemic area of the Danube region, especially in the province of Lower Austria. The increased incidence was reported to be associated with two simultaneously occurring field hare epizootics in the same area [83,84]. Between 1999 and 2013, fewer than 10 infections per year were consistently reported. In 2014, no human cases of tularemia were notified, while in the following years, a fluctuation of the incidence rate was observed again. A significantly above-average number of cases was recorded in 2017 (13 cases) and 2019 (20 cases). The number of cases over the last three decades, broken down by individual federal provinces, are depicted in Figure 1. A geographical summative representation of tularemia cases in humans in the various Austrian provinces from 1990 to 2019 is shown in Figure 2. Interestingly, in recent years, several infections were reported from Western Austria, especially in the province of Tyrol, where only isolated cases of human tularemia had occurred before. In 2016, a small series of tick-borne, ulceroglandular tularemia in humans was registered in Tyrol [78]. Later in 2018, the first two human cases of ulceroglandular tularemia after direct contact with an infected hare were confirmed in a hunter and a butcher near Innsbruck, Tyrol (Supplementary Materials, Figure S1).
3.2. F. tularensis in Animals
In Austria, F. tularensis has been detected in hares, wild rabbits, mice, and ticks [51,53,83,84,85]. Due to their status as common vectors for tularemia, prevalence studies have been performed most frequently in field hares.
3.2.1. F. tularensis in Hares
It is assumed that transmission to hares may occur as a consequence to preceding mouse epizootics [86]. In 1936, a field hare at the border of former Czechoslovakia tested positive for tularemia for the first time [85]. In 2003, Winkelmayer et al. examined a total of 311 serum samples derived from clinically healthy European field hares from six Lower Austrian districts (northeastern region comprising the “natural focus”) for antibodies against F. tularensis. Of these serum samples, 7.1% tested positive, which is in good agreement with earlier results by Höflechner-Pöltl et al. in 2000 (positive detection in 4.5%). However, it must be noted that the latter study only included clinically ill animals and animals killed in accidents [87,88]. Between 1994 and 2005, a total of 271 hares tested positive for F. tularensis in Lower Austria, Burgenland, and Styria [89].
3.2.2. F. tularensis in Foxes
In 2007, F. tularensis was detected for the first time in the mandibular lymph nodes of red foxes (Vulpes vulpes) from northern Burgenland that were collected by hunters as part of the rabies control program [90]. Between 2007 and 2008, Hofer and colleagues investigated mandibular lymph nodes of 903 red foxes from the provinces of Lower Austria, Burgenland, Upper Austria, and Styria. Infection with F. tularensis was detected in only 10 animals, and the distribution of the positive findings corresponded to the known endemic areas in Eastern Austria [91].
3.2.3. F. tularensis in Ticks
Experimental animal transmission tests for F. tularensis were performed with ticks from Austria in 1998. In this study, 2.8% of Dermacentor reticulatus ticks from Lower Austria transmitted tularemia to mice [51]. Furthermore, between 1991 and 1997, up to 1.3% of Dermacentor reticulatus ticks collected in regions of northeastern Austria were infected with F. tularensis [53]. In a study in Baden-Württemberg (Germany), which geographically borders Western Austria, a 16S rRNA-gene screening by PCR of 95 pools of Ixodes ricinus ticks showed a positive signal in 8.4% [92]. Transmission of tularemia via ticks to humans was first reported in Western Austria [78].
3.2.4. F. tularensis in Domestic Animals
There is only limited data in terms of tularemia in domestic animals in Austria. An examination of 80 clinically healthy dogs, used for hunting in the endemic area of Lower Austria, showed a seroprevalence for F. tularensis in 6.25% [93]. According to our literature search, no human infections transmitted by domestic animals have been reported in Austria so far.
4. Discussion
Tularemia is a rare infection in humans in Austria. Low numbers have been reported throughout the last 85 years, though sporadic epidemic outbreaks have occurred, all of them originating from the North and Northeast of Austria. From the districts north of the Danube in Lower Austria, F. tularensis spread further south to Burgenland and Styria; thus, for many years, this region of Austria was regarded the “natural or endemic focus” of tularemia. Since then, hares have been considered to play an important role in transmission of tularemia in Austria, and humans have been assumed to be infected when skinning and handling the carcasses [79]. Since 1991, an official registry of human infections is accessible in Austria. While some cases of active natural endemic areas in Eastern Austria are continuously being reported, until recently, single (likely imported) human cases were diagnosed in Western Austria (Vorarlberg, Tyrol, and Salzburg), which had been regarded as free of tularemia in the past. In 2016, a series of human tularemia cases were reported from Tyrol, and all these infections were transmitted via tick bites, a way of transmission that had not been reported before in Austria [78]. Thus, the question arises whether ticks were underestimated vectors in the endemic regions around Danube, March, and Thaya and had moved from east to west. Alternatively, a new endemic area in Western Austria could have evolved due to the migration of infected ticks from the north (Germany) and west (Switzerland), where they had previously been described as the main vectors of tularemia [94,95]. Interestingly, F. tularensis subsp. holarctica biovar I was identified in one Tyrolean patient for the first time in Austria [78]. The distribution of biovar I and II in Western Austria is in line with the prevalent data on the coexistence of both biovars in Germany and Switzerland, and demonstrates an epidemiological spread of different clades of F. tularensis subsp. holarctica in these countries [96,97,98].
In 2016, based on genetic and geographical analyses, Dwibedi et al. demonstrated that F. tularensis had spread from east to west within Europe [18]. Environmental and climatic factors seem to play a key role in the incidence of tularemia. In 2009, Deutz and colleagues investigated the impact of climate and weather on the geographical distribution of tularemia in Austria, and demonstrated a positive correlation to mild winters, cool springs, and a high precipitation amount in summer. They calculated a 3.5-fold increase of the tularemia endemic area in Austria, with an expansion from east to west by 2035 due to global warming [89]. An important contributing factor is the effect of climate change on population density, as well as the susceptibility of field hares (especially young animals) to pathogens, as these still play a key role in Austria as natural hosts for F. tularensis. In turn, weakening of young animals makes them easier prey for natural enemies, such as the red fox, thus favoring transmission between animals of different species with a greater range of movement and concomitant spread of tularemia [89]. The reported human infections caused by the handling of a hunted hare indicate that this species has now become an established reservoir for F. tularensis in Western Austria. From April 2018 to May 2020, the pathogen was detected in 38 European brown hares, sent to the Institute for Veterinary Disease Control of the Austrian Agency for Health and Food Safety (AGES) in Mödling by official veterinarians, indicating epizootics in hares in the federal states Salzburg, Tyrol, and Vorarlberg. Thirty cases of tularemia from hares were found in Salzburg, seven in Vorarlberg, and one in Tyrol (unpublished data). These data suggest that the expansion of the F. tularensis habitat, which was estimated to not be reached before 2035, has now already been attained [89]. Prevalence data of F. tularensis in Austrian ticks, as well as in mosquitos or horse flies, are very scarce or completely absent. As these are known potential vectors of tularemia to humans, further investigation is required. In order to improve the epidemiologic knowledge of tularemia in wild animals and the associated risks to humans, long-term systemic monitoring of known natural reservoir animals will be essential in the future.
5. Conclusions
Previous studies on the prevalence of F. tularensis in field hares, ticks, and to a lesser extent, red foxes have been limited to the known endemic areas in Eastern Austria, and therefore allow only a very limited estimation of the actual natural spread of tularemia in Austria. Given the knowledge that epizootics precede epidemics of tularemia in humans, long-term systemic surveillance of natural foci will be essential to monitor the distribution and prevalence of F. tularensis, in order to anticipate possible risks of outbreaks and to take appropriate preventative measures [53]. This requires the broad screening of potential reservoir animals for F. tularensis, in order to get a real image of distribution of tularemia in Austria, which can only be successful through close cooperation of humans, veterinary health professionals, and authorities. It is furthermore important to raise awareness of tularemia among the general public, particularly for risk groups such as hunters, foresters, butchers, and taxidermists. Finally, doctors must be trained to be aware of this infection with its various forms of presentation [99].
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2607/8/10/1597/s1, Figure S1: Ulceroglandular infection with F. tularensis originating from an infected brown hare (a) and ulcer in the palm of patient 1 (b) acquired in Tyrol in 2018.
Author Contributions
Conceptualization, S.S. and R.B.-W.; methodology, S.S. and R.B.-W.; validation, S.S., A.S., E.H. and R.B.-W.; resources, S.S., A.S., E.H., G.W. and R.B.-W.; writing—original draft preparation, S.S. and R.B.-W.; writing—review and editing, E.H., G.W. and R.B.-W.; supervision, R.B.-W.; funding acquisition, G.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sjöstedt, A. Tularemia: History, epidemiology, pathogen physiology, and clinical manifestations. Ann. N. Y. Acad. Sci. 2007, 1105, 1–29. [Google Scholar] [CrossRef] [PubMed]
- McCoy, G.W.; Chapin, C.W. Further observations on a plague-like disease of rodents with a preliminary note on the causative agent, Bacterium tularense. J. Infect. Dis. 1912, 10, 61–72. [Google Scholar] [CrossRef]
- Wherry, W.B.; Lamb, B.H. Infection of man with Bacterium tularense. 1914. J. Infect. Dis. 2004, 189, 1321–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöstedt, A.B. Francisella. Proteobacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Stanley, J.T., Garrity, G.M., Eds.; Part B; Springer: New York, NY, USA, 2005; Volume 2, pp. 200–210. [Google Scholar]
- Larson, C.L.; Wicht, W.; Jellison, W.L. A new organism resembling P. tularensis isolated from water. Public Health Rep. 1955, 70, 253–258. [Google Scholar] [CrossRef]
- Hollis, D.G.; Weaver, R.E.; Steigerwalt, A.G.; Wenger, J.D.; Moss, C.W.; Brenner, D.J. Francisella philomiragia comb. nov. (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease. J. Clin. Microbiol. 1989, 27, 1601–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingry, L.C.; Petersen, J.M. Comparative review of Francisella tularensis and Francisella novicida. Front. Cell. Infect. Microbiol. 2014, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Tomaso, H.; Al Dahouk, S.; Hofer, E.; Splettstoesser, W.D.; Treu, T.M.; Dierich, M.P.; Neubauer, H. Antimicrobial susceptibilities of Austrian Francisella tularensis holarctica biovar II strains. Int. J. Antimicrob. Agents 2005, 26, 279–284. [Google Scholar] [CrossRef]
- Ellis, J.; Oyston, P.C.; Green, M.; Titball, R.W. Tularemia. Clin. Microbiol. Rev. 2002, 15, 631–646. [Google Scholar] [CrossRef] [Green Version]
- Gyuranecz, M.; Birdsell, D.N.; Splettstoesser, W.; Seibold, E.; Beckstrom-Sternberg, S.M.; Makrai, L.; Fodor, L.; Fabbi, M.; Vicari, N.; Johansson, A.; et al. Phylogeography of Francisella tularensis subsp. holarctica, Europe. Emerg. Infect. Dis. 2012, 18, 290–293. [Google Scholar] [CrossRef] [Green Version]
- Ol’sufev, N.G.; Meshcheriakova, I.S. Intraspecific taxonomy of tularemia agent Francisella tularensis McCoy et Chapin. J. Hyg. Epidemiol. Microbiol. Immunol. 1982, 20, 291–299. [Google Scholar]
- Johansson, A.; Farlow, J.; Larsson, P.; Dukerich, M.; Chambers, E.; Byström, M.; Fox, J.; Chu, M.; Forsman, M.; Sjöstedt, A.; et al. Worldwide genetic relationships among Francisella tularensis isolates determined by multiple-locus variable-number tandem repeat analysis. J. Bacteriol. 2004, 186, 5808–5818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, P.; Svensson, K.; Karlsson, L.; Guala, D.; Granberg, M.; Forsman, M.; Johanssont, A. Canonical insertion-deletion markers for rapid DNA typing of Francisella tularensis. Emerg. Infect. Dis. 2007, 13, 1725–1732. [Google Scholar] [CrossRef]
- Svensson, K.; Granberg, M.; Karlsson, L.; Neubauerova, V.; Forsman, M.; Johansson, A. A real-time PCR array for hierarchical identification of Francisella isolates. PLoS ONE 2009, 4, e8360. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.; Svensson, K.; Lindgren, P.; Byström, M.; Sjödin, A.; Forsman, M.; Johansson, A. The phylogeographic pattern of Francisella tularensis in Sweden indicates a Scandinavian origin of Eurosiberian tularaemia. Environ. Microbiol. 2013, 15, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Fujita, O.; Uda, A.; Hotta, A.; Okutani, A.; Inoue, S.; Tanabayashi, K.; Yamada, A. Genetic diversity of Francisella tularensis subspecies holarctica strains isolated in Japan. Microbiol. Immunol. 2008, 52, 270–276. [Google Scholar] [CrossRef]
- Vogler, A.J.; Birdsell, D.; Price, L.B.; Bowers, J.R.; Beckstrom-Sternberg, S.M.; Auerbach, R.K.; Beckstrom-Sternberg, J.S.; Johansson, A.; Clare, A.; Buchhagen, J.L.; et al. Phylogeography of Francisella tularensis: Global expansion of a highly fit clone. J. Bacteriol. 2009, 191, 2474–2484. [Google Scholar] [CrossRef] [Green Version]
- Dwibedi, D.; Birdsell, D.; Lärkeryd, A.; Myrtennäs, K.; Öhrman, C.; Nilsson, E.; Karlsson, E.; Hochhalter, C.; Rivera, A.; Maltinsky, S.; et al. Long-range dispersal moved Francisella tularensis into Western Europe from the East. Microb. Genom. 2016, 2, e000100. [Google Scholar] [CrossRef]
- Jackson, J.; McGregor, A.; Cooley, L.; Ng, J.; Brown, M.; Ong, C.W.; Darcy, C.; Sintchenko, V. Francisella tularensis subspecies holarctica, Tasmania, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 1484–1486. [Google Scholar] [CrossRef] [Green Version]
- Eden, J.S.; Rose, K.; Ng, J.; Shi, M.; Wang, Q.; Sintchenko, V.; Holmes, E.C. Francisella tularensis ssp. holarctica in Ringtail Possums, Australia. Emerg. Infect. Dis. 2017, 23, 1198–1201. [Google Scholar] [CrossRef] [Green Version]
- Staples, J.E.; Kubota, K.A.; Chalcraft, L.G.; Mead, P.S.; Petersen, J.M. Epidemiologic and molecular analysis of human tularemia, United States, 1964–2004. Emerg. Infect. Dis. 2006, 12, 1113–1118. [Google Scholar] [CrossRef]
- Gurycová, D. First isolation of Francisella tularensis subsp. tularensis in Europe. Eur. J. Epidemiol. 1998, 14, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Gyuranecz, M. Tularaemia: Clinical aspects in Europe. Lancet Infect. Dis. 2016, 16, 113–124. [Google Scholar] [CrossRef]
- Chaudhuri, R.R.; Ren, C.P.; Desmond, L.; Vincent, G.A.; Silman, N.J.; Brehm, J.K.; Elmore, M.J.; Hudson, M.J.; Forsman, M.; Isherwood, K.E.; et al. Genome sequencing shows that European isolates of Francisella tularensis subspecies tularensis are almost identical to US laboratory strain Schu S4. PLoS ONE 2007, 2, e352. [Google Scholar] [CrossRef] [PubMed]
- Gyuranecz, M.; Rigó, K.; Dán, A.; Földvári, G.; Makrai, L.; Dénes, B.; Fodor, L.; Majoros, G.; Tirják, L.; Erdélyi, K. Investigation of the ecology of Francisella tularensis during an inter-epizootic period. Vector Borne Zoonotic Dis. 2011, 11, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Mörner, T. The ecology of tularaemia. Rev. Sci. Tech. 1992, 11, 1123–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hestvik, G.; Warns-Petit, E.; Smith, L.A.; Fox, N.J.; Uhlhorn, H.; Artois, M.; Hannant, D.; Hutchings, M.R.; Mattsson, R.; Yon, L.; et al. The status of tularemia in Europe in a one-health context: A review. Epidemiol. Infect. 2015, 143, 2137–2160. [Google Scholar] [CrossRef] [Green Version]
- Parker, R.R.; Steinhaus, E.A.; Kohls, G.M.; Jellison, W.L. Contamination of natural waters and mud with Pasteurella tularensis and tularemia in beavers and muskrats in the northwestern United States. Bull. Natl. Inst. Health 1951, 193, 1–161. [Google Scholar]
- Willke, A.; Meric, M.; Grunow, R.; Sayan, M.; Finke, E.J.; Splettstößer, W.; Seibold, E.; Erdoğan, S.; Ergonul, O.; Yumuk, Z.; et al. An outbreak of oropharyngeal tularaemia linked to natural spring water. J. Med. Microbiol. 2009, 58, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Matyas, B.T.; Nieder, H.S.; Telford, S.R. Pneumonic tularemia on Martha’s Vineyard: Clinical, epidemiologic, and ecological characteristics. Ann. N. Y. Acad. Sci. 2007, 1105, 351–377. [Google Scholar] [CrossRef]
- Reintjes, R.; Dedushaj, I.; Gjini, A.; Jorgensen, T.R.; Cotter, B.; Lieftucht, A.; D’Ancona, F.; Dennis, D.T.; Kosoy, M.A.; Mulliqi-Osmani, G.; et al. Tularemia outbreak investigation in Kosovo: Case control and environmental studies. Emerg. Infect. Dis. 2002, 8, 69–73. [Google Scholar] [CrossRef]
- Chitadze, N.; Kuchuloria, T.; Clark, D.V.; Tsertsvadze, E.; Chokheli, M.; Tsertsvadze, N.; Trapaidze, N.; Lane, A.; Bakanidze, L.; Tsanava, S.; et al. Water-borne outbreak of oropharyngeal and glandular tularemia in Georgia: Investigation and follow-up. Infection 2009, 37, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Hopla, C.E. The ecology of tularemia. Adv. Vet. Sci. Comp. Med. 1974, 18, 25–53. [Google Scholar]
- Gyuranecz, M.; Szeredi, L.; Makrai, L.; Fodor, L.; Mészáros, A.R.; Szépe, B.; Füleki, M.; Erdélyi, K. Tularemia of European Brown Hare (Lepus europaeus): A pathological, histopathological, and immunohistochemical study. Vet. Pathol. 2010, 47, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Decors, A.; Lesage, C.; Jourdain, E.; Giraud, P.; Houbron, P.; Vanhem, P.; Madani, N.; Madani, M. Outbreak of tularaemia in brown hares (Lepus europaeus) in France, January to March 2011. Euro Surveill. 2011, 16, 19913. [Google Scholar] [PubMed]
- Hestvik, G.; Uhlhorn, H.; Södersten, F.; Åkerström, S.; Karlsson, E.; Westergren, E.; Gavier-Widén, D. Tularaemia in European Brown Hares (Lepus europaeus) and Mountain Hares (Lepus timidus) Characterized by Histopathology and Immunohistochemistry: Organ Lesions and Suggestions of Routes of Infection and Shedding. J. Comp. Pathol. 2017, 157, 103–114. [Google Scholar] [CrossRef]
- Origgi, F.C.; Pilo, P. Francisella Tularensis Clades B.FTN002-00 and B.13 Are Associated with Distinct Pathology in the European Brown Hare (Lepus europaeus). Vet. Pathol. 2016, 53, 1220–1232. [Google Scholar] [CrossRef] [Green Version]
- Kreizinger, Z.; Erdélyi, K.; Felde, O.; Fabbi, M.; Sulyok, K.M.; Magyar, T.; Gyuranecz, M. Comparison of virulence of Francisella tularensis ssp. holarctica genotypes B.12 and B.FTNF002-00. BMC Vet. Res. 2017, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Downs, C.M.; Coriell, L.L. Studies on tularemia; the comparative susceptibility of various laboratory animals. J. Immunol. 1947, 56, 217–228. [Google Scholar]
- Bell, J.F.; Stewart, S.J. Chronic shedding tularemia nephritis in rodents: Possible relation to occurrence of Francisella tularensis in lotic waters. J. Wildl. Dis. 1975, 11, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Karpoff, S.P.; Antonoff, N.I. The Spread of Tularemia through Water, as a New Factor in Its Epidemiology. J. Bacteriol. 1936, 32, 243–258. [Google Scholar] [CrossRef] [Green Version]
- Berdal, B.P.; Mehl, R.; Haaheim, H.; Løksa, M.; Grunow, R.; Burans, J.; Morgan, C.; Meyer, H. Field detection of Francisella tularensis. Scand. J. Infect. Dis. 2000, 32, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Brantsaeter, A.B.; Krogh, T.; Radtke, A.; Nygard, K. Tularaemia outbreak in northern Norway. Eurosurveillance 2007, 12, E070329.2. [Google Scholar] [CrossRef] [PubMed]
- Akalin, H.; Helvaci, S.; Gedikoğlu, S. Re-emergence of tularemia in Turkey. Int. J. Infect. Dis. 2009, 13, 547–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantardjiev, T.; Ivanov, I.; Velinov, T.; Padeshki, P.; Popov, B.; Nenova, R.; Mincheff, M. Tularemia outbreak, Bulgaria, 1997–2005. Emerg. Infect. Dis. 2006, 12, 678–680. [Google Scholar] [CrossRef]
- Allue, M.; Sopeña, C.R.; Gallardo, M.T.; Mateos, L.; Vian, E.; Garcia, M.J.; Ramos, J.; Berjon, A.C.; Viña, M.C.; Garcia, M.P.; et al. Tularaemia outbreak in Castilla y León, Spain, 2007: An update. Eurosurveillance 2008, 13, 18948. [Google Scholar]
- Martín, C.; Gallardo, M.T.; Mateos, L.; Vián, E.; García, M.J.; Ramos, J.; Berjón, A.C.; del Carmen Viña, M.; García, M.P.; Yáñez, J.; et al. Outbreak of tularaemia in Castilla y León, Spain. Eurosurveillance 2007, 12, E071108.1. [Google Scholar] [CrossRef]
- Keim, P.; Johansson, A.; Wagner, D.M. Molecular epidemiology, evolution, and ecology of Francisella. Ann. N. Y. Acad. Sci. 2007, 1105, 30–66. [Google Scholar] [CrossRef]
- Friend, M. Tularemia; No. 1297; US Geological Survey: Reston, VA, USA, 2006.
- Zellner, B.; Huntley, J.F. Ticks and Tularemia: Do We Know What We Don’t Know? Front. Cell. Infect. Microbiol. 2019, 9, 146. [Google Scholar] [CrossRef]
- Hubálek, Z.; Sixl, W.; Halouzka, J. Francisella tularensis in Dermacentor reticulatus ticks from the Czech Republic and Austria. Wiener Klin. Wochenschr. 1998, 110, 909–910. [Google Scholar]
- Gurycová, D.; Kocianová, E.; Výrosteková, V.; Rehácek, J. Prevalence of ticks infected with Francisella tularensis in natural foci of tularemia in western Slovakia. Eur. J. Epidemiol. 1995, 11, 469–474. [Google Scholar] [CrossRef]
- Gurycová, D.; Výrosteková, V.; Khanakah, G.; Kocianová, E.; Stanek, G. Importance of surveillance of tularemia natural foci in the known endemic area of Central Europe, 1991–1997. Wien. Klin. Wochenschr. 2001, 113, 433–438. [Google Scholar] [PubMed]
- Eliasson, H.; Lindbäck, J.; Nuorti, J.P.; Arneborn, M.; Giesecke, J.; Tegnell, A. The 2000 tularemia outbreak: A case-control study of risk factors in disease-endemic and emergent areas, Sweden. Emerg. Infect. Dis. 2002, 8, 956–960. [Google Scholar] [CrossRef] [PubMed]
- Desvars, A.; Furberg, M.; Hjertqvist, M.; Vidman, L.; Sjöstedt, A.; Rydén, P.; Johansson, A. Epidemiology and ecology of tularemia in Sweden, 1984–2012. Emerg. Infect. Dis. 2015, 21, 32–39. [Google Scholar] [CrossRef]
- Rydén, P.; Björk, R.; Schäfer, M.L.; Lundström, J.O.; Petersén, B.; Lindblom, A.; Forsman, M.; Sjöstedt, A.; Johansson, A. Outbreaks of tularemia in a boreal forest region depends on mosquito prevalence. J. Infect. Dis. 2012, 205, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathisen, P.; Andersson, A.C.; Backman, S.; Lundstrom, J.; Kouppa, K.; Schafer, M.; Thelaus, J.; Forsman, M. Association and uptake of Francisella tularensis subspecies holarctica into mosquitoes, in Abstract P2-02. In Proceedings of the 6th International Conference on Tularemia, Berlin, Germany, 13–16 September 2009. [Google Scholar]
- Telford, S.R.; Goethert, H.K. Toward an understanding of the perpetuation of the agent of tularemia. Front. Microbiol. 2010, 1, 150. [Google Scholar] [CrossRef] [Green Version]
- Pilo, P. Phylogenetic Lineages of Francisella tularensis in Animals. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef]
- Kwit, N.A.; Schwartz, A.; Kugeler, K.J.; Mead, P.S.; Nelson, C.A. Human tularaemia associated with exposure to domestic dogs-United States, 2006–2016. Zoonoses Public Health 2019, 66, 417–421. [Google Scholar] [CrossRef]
- Capellan, J.; Fong, I.W. Tularemia from a cat bite: Case report and review of feline-associated tularemia. Clin. Infect. Dis. 1993, 16, 472–475. [Google Scholar] [CrossRef]
- Larson, M.A.; Fey, P.D.; Hinrichs, S.H.; Iwen, P.C. Francisella tularensis bacteria associated with feline tularemia in the United States. Emerg. Infect. Dis. 2014, 20, 2068–2071. [Google Scholar] [CrossRef]
- WHO. WHO Guidelines on Tularaemia; WHO: Geneva, Switzerland, 2007. Available online: https://www.cdc.gov/tularemia/resources/whotularemiamanual.pdf (accessed on 15 August 2020).
- Ernst, M.; Pilo, P.; Fleisch, F.; Glisenti, P. Tularemia in the Southeastern Swiss Alps at 1700 m above sea level. Infection 2015, 43, 111–115. [Google Scholar] [CrossRef]
- Tärnvik, A.; Berglund, L. Tularaemia. Eur. Respir. J. 2003, 21, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, H.S.; Miller, L.; Halasohoris, S.; Purcell, B.K. Antibiotic Susceptibilities of Francisella tularensis Determined by Broth Microdilution following CLSI Methods. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgi, E.; Schacht, E.; Scholz, H.C.; Splettstoesser, W.D. Standardized broth microdilution antimicrobial susceptibility testing of Francisella tularensis subsp. holarctica strains from Europe and rare Francisella species. J. Antimicrob. Chemother. 2012, 67, 2429–2433. [Google Scholar] [CrossRef]
- Urich, S.K.; Petersen, J.M. In vitro susceptibility of isolates of Francisella tularensis types A and B from North America. Antimicrob. Agents. Chemother. 2008, 52, 2276–2278. [Google Scholar] [CrossRef] [Green Version]
- Enderlin, G.; Morales, L.; Jacobs, R.F.; Cross, J.T. Streptomycin and alternative agents for the treatment of tularemia: Review of the literature. Clin. Infect. Dis. 1994, 19, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Marohn, M.E.; Barry, E.M. Live attenuated tularemia vaccines: Recent developments and future goals. Vaccine 2013, 31, 3485–3491. [Google Scholar] [CrossRef] [Green Version]
- Abraham, A.; Ostroff, G.; Levitz, S.M.; Oyston, P.C.F. A novel vaccine platform using glucan particles for induction of protective responses against Francisella tularensis and other pathogens. Clin. Exp. Immunol. 2019, 198, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Horwitz, M.A. Live Attenuated Tularemia Vaccines for Protection against Respiratory Challenge with Virulent F. tularensis subsp. tularensis. Front. Cell. Infect. Microbiol. 2018, 8, 154. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Annual Epidemiological Report: Reporting on 2009 Surveillance Data and 2010 Epidemic Intelligence Data; European Centre for Disease Prevention and Control: Solna, Sweden, 2011; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/1111_SUR_Annual_Epidemiological_Report_on_Communicable_Diseases_in_Europ....pdf (accessed on 15 September 2020).
- European Centre for Disease Prevention and Control. Tularaemia Annual Epidemiological Report for 2018; European Centre for Disease Prevention and Control: Solna, Sweden, 2018; Available online: https://www.ecdc.europa.eu/en/publications-data/tularaemia-annual-epidemiological-report-2018#no-link (accessed on 24 August 2020).
- European Centre for Disease Prevention and Control. Tularaemia—Annual Epidemiological Report for 2017; European Centre for Disease Prevention and Control: Solna, Sweden, 2019; Available online: https://www.ecdc.europa.eu/en/publications-data/tularaemia-annual-epidemiological-report-2017 (accessed on 24 August 2020).
- Tobudic, S.; Nedomansky, K.; Poeppl, W.; Müller, M.; Faas, A.; Mooseder, G.; Allerberger, F.; Stanek, G.; Burgmann, H. Seroprevalence for Coxiella burnetii, Francisella tularensis, Brucella abortus and Brucella melitensis in Austrian adults: A cross-sectional survey among military personnel and civilians. Ticks Tick Borne Dis. 2014, 5, 315–317. [Google Scholar] [CrossRef]
- Antonitsch, L.; Weidinger, G.; Stanek, G.; Markowicz, M. Francisella tularensis as the cause of protracted fever. BMC Infect. Dis. 2020, 20, 327. [Google Scholar] [CrossRef]
- Schroll, A.; Theurl, I.; Georgi, E.; Zange, S.; Rettenbacher, T.; Bellmann-Weiler, R.; Weiss, G. Newly emerging ulceroglandular tularaemia in Western Austria. Ticks Tick Borne Dis. 2018, 9, 1331–1333. [Google Scholar] [CrossRef] [PubMed]
- Puntigam, F. Thorakale Formen im Seuchengeschehen der Tularämie in Österreich. Wiener Klinische Wochenschrift 1960, 72, 813–816. [Google Scholar] [PubMed]
- Puntigam, F. Erkrankungen an thorakalen Formen der Tularämie bei Arbeitnehmern in Zuckerfabriken. Zeitschrift für Hygiene und Infektionskrankheiten 1960, 147, 162–168. [Google Scholar] [CrossRef]
- Jusatz, H.J. Dritter Bericht über das Vordringen der Tularämie nach Mittel- und Westeuropa über den Zeitraum von 1950–1960. Zeitschrift für Hygiene und Infektionskrankheiten 1961, 148, 69–93. [Google Scholar] [CrossRef]
- Federal Ministry of Labour, Social Affairs, Health and Consumer Protection. Jahresstatistik Meldepflichtiger Infektionskrankheiten 1990–2019. Available online: https://www.sozialministerium.at/Themen/Gesundheit/Uebertragbare-Krankheiten/Statistiken-und-Fallzahlen.html (accessed on 10 August 2020).
- Hofer, E.; Schildorfer, H.; Flatscher, J.; Müller, M. Zum Nachweis der Tularämie bei Feldhasen (Lepus europaeus) in Österreich. Wien. Tierarztl. Mon. 1997, 84, 301–306. [Google Scholar]
- Steineck, T.; Hofer, E. Zum Vorkommen der Tularämie in Österreich. In Verhandlungsbericht des 39. Internationalen Symposiums über die Erkrankungen der Zoo- und Wildtiere; Akademie-Verlag: Wien, Austria, 1999; pp. 263–269. [Google Scholar]
- David, H. Zum Auftreten der Tularämie in Österreich. Wien. Klin. Wochenschr. 1937, 50, 459–462. [Google Scholar]
- Michalka, J. Die Tularämie in Österreich. Wien. Klin. Wochenschr. 1960, 47, 341–348. [Google Scholar]
- Höflechner-Pöltl, A.; Hofer, E.; Awad-Masalmeh, M.; Müller, M.; Steineck, T. Prevalence of tularaemia and brucellosis in European brown hare (Lepus europaeus) and red fox (Vulpes vulpes) in Austria. Tierärztliche Umsch. 2000, 55, 264–268. [Google Scholar]
- Winkelmayer, R.; Vodnansky, M.; Paulsen, P.; Gansterer, A.; Treml, F. Explorative study on the seroprevalence of Brucella-, Francisella- and Leptospira antibodies in the European hare (Lepus europaeus Pallas) of the Austrian-Czech border region. Wien. Tierarztl. Mon. 2005, 92, 131–135. [Google Scholar]
- Deutz, A.; Guggenberger, T.; Gasteiner, J.; Steineck, T.; Bago, Z.; Hofer, E.; Auer, I.; Böhm, R. Untersuchungen zur Verbreitung der Tularämie in Österreich unter dem Aspekt des Klimawandels. Wien. Tierarztl. Mon. 2009, 96, 107–113. [Google Scholar]
- Hofer, E.; Revilla-Fernandez, S.; Plicka, H.; Reisp, K.; Richter, S.; Müller, M.; Fink, M.; Romanek, B.; Bago, Z.; Weikel, J.; et al. Erstmalige Isolierung von Francisella tularensis subsp. holarctica Biovar II und Brucella suis Biovar 2 beim Rotfuchs in Österreich. Available online: https://www.ages.at/themen/krankheitserreger/tularaemie-hasenpest/ (accessed on 25 August 2020).
- Hofer, E.; Reisp, K.; Winter, P.; Köfer, J. Zur Überwachung der österreichischen Tularämie- und Brucellose-Naturherde mit dem Rotfuchs (Vulpes vulpes) als Bioindikator. Available online: https://www.ages.at/themen/krankheitserreger/tularaemie-hasenpest/ (accessed on 25 August 2020).
- Gehringer, H.; Schacht, E.; Maylaender, N.; Zeman, E.; Kaysser, P.; Oehme, R.; Pluta, S.; Splettstoesser, W.D. Presence of an emerging subclone of Francisella tularensis holarctica in Ixodes ricinus ticks from south-western Germany. Ticks Tick Borne Dis. 2013, 4, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Posautz, A.; Gyuranecz, M.; Dénes, B.; Knauer, F.; Dier, H.; Walzer, C. Seroprevalence of Francisella tularensis in Austrian Hunting Dogs. Vector Borne Zoonotic Dis. 2018, 18, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Borde, J.P.; Zange, S.; Antwerpen, M.H.; Georgi, E.; von Buttlar, H.; Kern, W.V.; Rieg, S. Five cases of vector-borne Francisella tularensis holarctica infections in south-western Germany and genetic diversity. Ticks Tick Borne Dis. 2017, 8, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, M.; Altpeter, E.; Pilo, P.; Gygli, S.M.; Beuret, C.; Foucault, F.; Ackermann-Gäumann, R.; Karrer, U.; Jacob, D.; Grunow, R.; et al. Population Genomics of Francisella tularensis subsp. holarctica and its Implication on the Eco-Epidemiology of Tularemia in Switzerland. Front. Cell. Infect. Microbiol. 2018, 8, 89. [Google Scholar] [CrossRef]
- Origgi, F.C.; Frey, J.; Pilo, P. Characterisation of a new group of Francisella tularensis subsp. holarctica in Switzerland with altered antimicrobial susceptibilities, 1996 to 2013. Eurosurveillance 2014, 19. [Google Scholar] [CrossRef] [Green Version]
- Tomaso, H.; Hotzel, H.; Otto, P.; Myrtennäs, K.; Forsman, M. Antibiotic susceptibility in vitro of Francisella tularensis subsp. holarctica isolates from Germany. J. Antimicrob. Chemother. 2017, 72, 2539–2543. [Google Scholar] [CrossRef] [Green Version]
- Appelt, S.; Köppen, K.; Radonić, A.; Drechsel, O.; Jacob, D.; Grunow, R.; Heuner, K. Genetic Diversity and Spatial Segregation of. Front. Cell. Infect. Microbiol. 2019, 9, 376. [Google Scholar] [CrossRef] [Green Version]
- Tärnvik, A.; Priebe, H.S.; Grunow, R. Tularaemia in Europe: An epidemiological overview. Scand. J. Infect. Dis. 2004, 36, 350–355. [Google Scholar] [CrossRef]
Figure 1.
Cumulative cases of human tularemia in Austria divided by provinces from 1990–2019. Abbreviations: V = Vienna, B = Burgenland, LA = Lower Austria, UA = Upper Austria, C = Carinthia, ST = Styria, S = Salzburg, T = Tyrol, VO = Vorarlberg [82].
Figure 1.
Cumulative cases of human tularemia in Austria divided by provinces from 1990–2019. Abbreviations: V = Vienna, B = Burgenland, LA = Lower Austria, UA = Upper Austria, C = Carinthia, ST = Styria, S = Salzburg, T = Tyrol, VO = Vorarlberg [82].

Figure 2.
Summative geographic distribution of human tularemia cases in the various Austrian provinces from 1990 to 2019. The size of the dots corresponds to the number of humans infected with F. tularensis during a five year period [82].
Figure 2.
Summative geographic distribution of human tularemia cases in the various Austrian provinces from 1990 to 2019. The size of the dots corresponds to the number of humans infected with F. tularensis during a five year period [82].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Seiwald, S.; Simeon, A.; Hofer, E.; Weiss, G.; Bellmann-Weiler, R. Tularemia Goes West: Epidemiology of an Emerging Infection in Austria. Microorganisms 2020, 8, 1597. https://doi.org/10.3390/microorganisms8101597
AMA Style
Seiwald S, Simeon A, Hofer E, Weiss G, Bellmann-Weiler R. Tularemia Goes West: Epidemiology of an Emerging Infection in Austria. Microorganisms. 2020; 8(10):1597. https://doi.org/10.3390/microorganisms8101597
Chicago/Turabian StyleSeiwald, Stefanie, Anja Simeon, Erwin Hofer, Günter Weiss, and Rosa Bellmann-Weiler. 2020. "Tularemia Goes West: Epidemiology of an Emerging Infection in Austria" Microorganisms 8, no. 10: 1597. https://doi.org/10.3390/microorganisms8101597
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.