Intensified Use of Reproductive Technologies and Reduced Dimensions of Breeding Schemes Put Genetic Diversity at Risk in Dairy Cattle Breeds

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
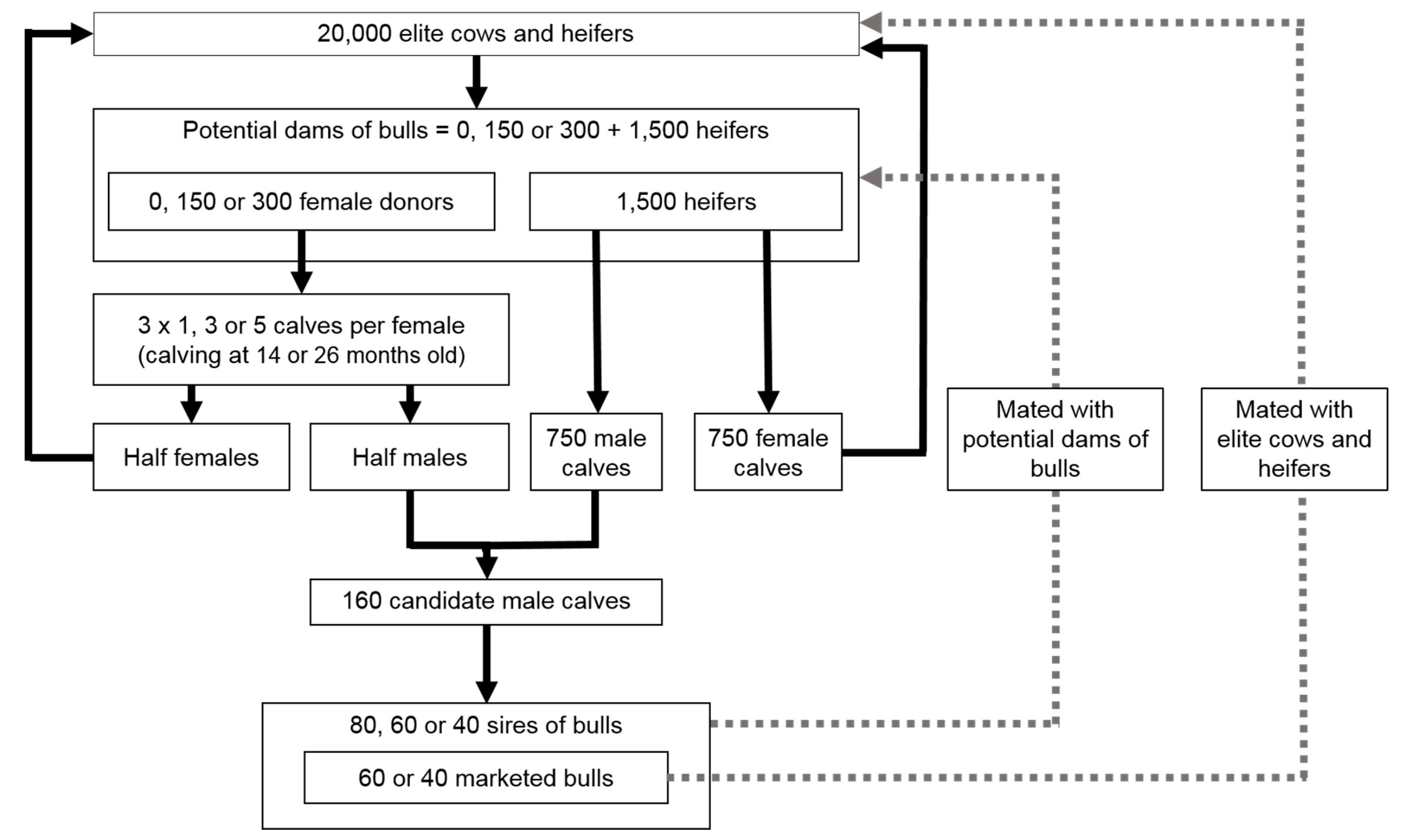
2.1. Simulated Population and Scenarios
2.2. Evaluation of Genetic Gain and Genetic Diversity Outcomes
3. Results
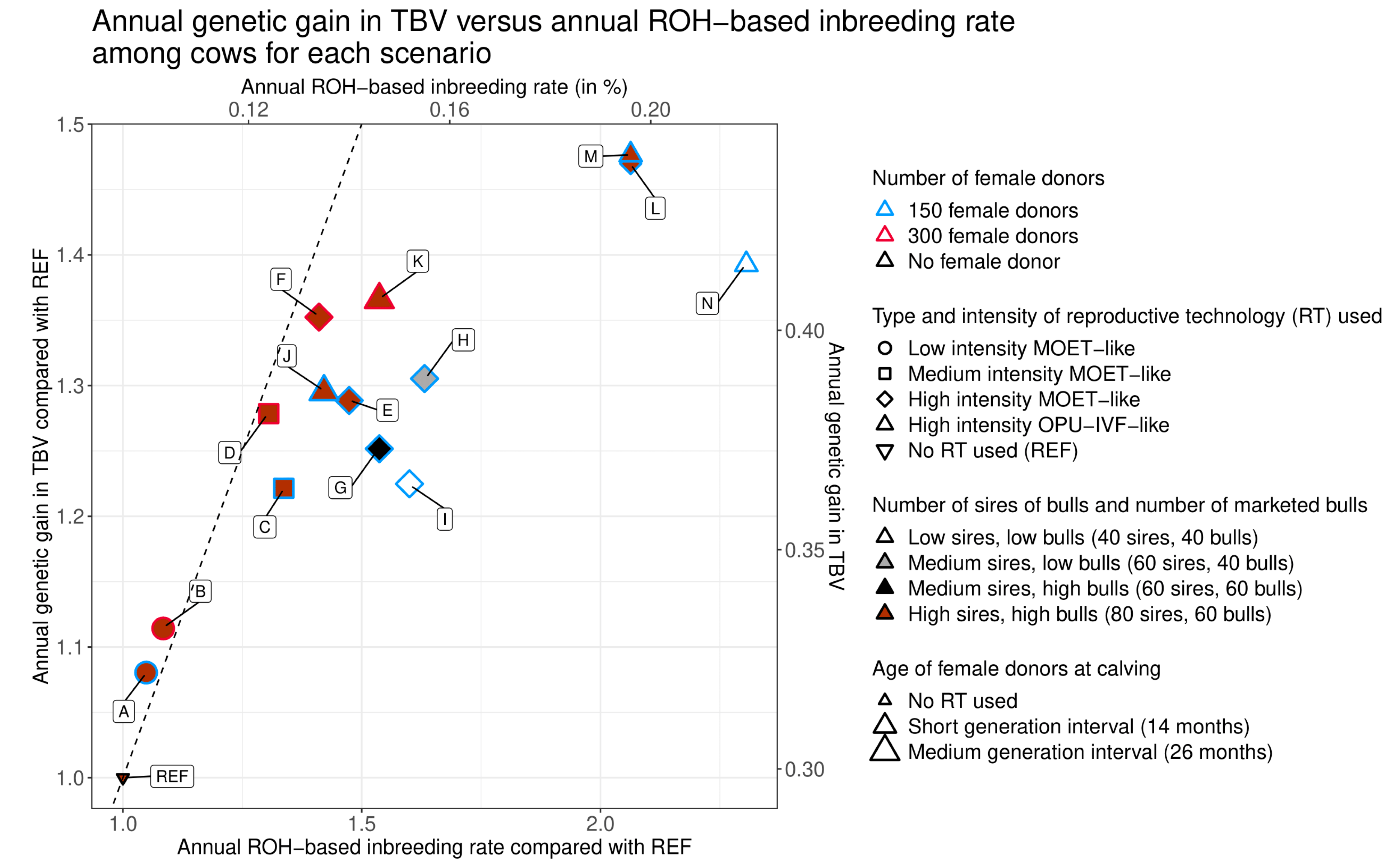
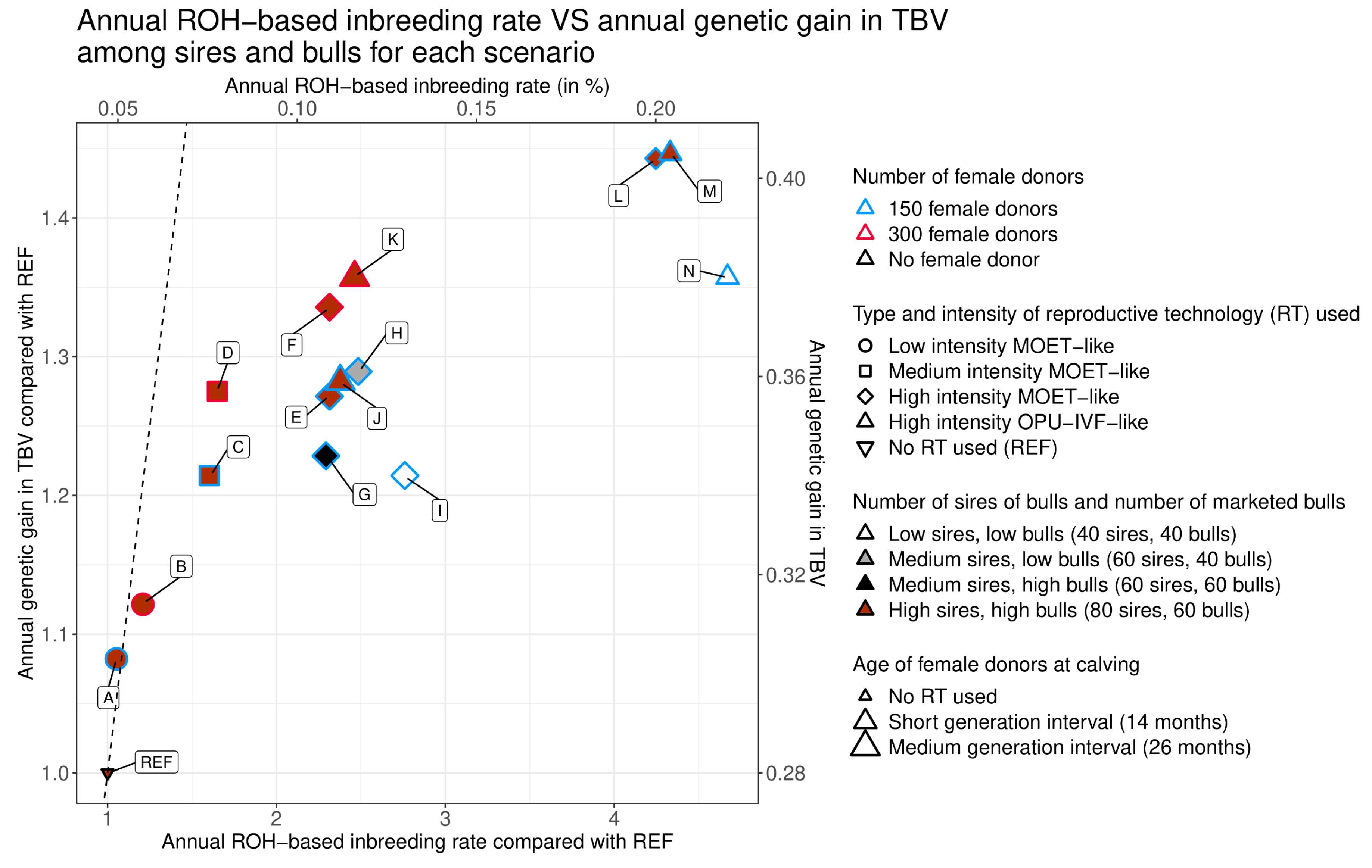
3.1. General Observations
3.2. Intensity of Reproductive Technologies (RT) Use and Number of Female Donors
3.3. Generation Interval between Female Donors and Type of RT Used
3.3.1. Generation Interval between Female Donors
3.3.2. Type of RT Used
3.4. Number of Sires of Bulls and of Marketed Bulls
3.4.1. Number of Sires of Bulls
3.4.2. Number of Marketed Bulls
3.4.3. Effect of RT Type on Reductions in the Number of Sires of Bulls and Marketed Bulls
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stachowicz, K.; Sargolzaei, M.; Miglior, F.; Schenkel, F.S. Rates of inbreeding and genetic diversity in Canadian Holstein and Jersey cattle. J. Dairy Sci. 2011, 94, 5160–5175. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, G.E.; Hazel, L.N. Effectiveness of selection on progeny performance as a supplement to earlier culling in livestock. J. Agric. Res. 1944, 69, 459–476. [Google Scholar]
- Notter, D.R. The importance of genetic diversity in livestock populations of the future. J. Anim. Sci. 1999, 77, 61. [Google Scholar] [CrossRef]
- Leroy, G. Inbreeding depression in livestock species: Review and meta-analysis. Anim. Genet. 2014, 45, 618–628. [Google Scholar] [CrossRef]
- Pryce, J.E.; Haile-Mariam, M.; Goddard, M.E.; Hayes, B.J. Identification of genomic regions associated with inbreeding depression in Holstein and Jersey dairy cattle. Genet. Sel. Evol. 2014, 46, 71. [Google Scholar] [CrossRef] [Green Version]
- Eynard, S.E.; Windig, J.J.; Hiemstra, S.J.; Calus, M.P.L. Whole-genome sequence data uncover loss of genetic diversity due to selection. Genet. Sel. Evol. 2016, 48. [Google Scholar] [CrossRef] [Green Version]
- Doublet, A.-C.; Croiseau, P.; Fritz, S.; Michenet, A.; Hozé, C.; Danchin-Burge, C.; Laloë, D.; Restoux, G. The impact of genomic selection on genetic diversity and genetic gain in three French dairy cattle breeds. Genet. Sel. Evol. 2019, 51, 52. [Google Scholar] [CrossRef] [Green Version]
- Weigel, K.A. Controlling Inbreeding in Modern Breeding Programs. J. Dairy Sci. 2001, 84, E177–E184. [Google Scholar] [CrossRef]
- Bouquet, A.; Sørensen, A.C.; Juga, J. Genomic selection strategies to optimize the use of multiple ovulation and embryo transfer schemes in dairy cattle breeding programs. Livest. Sci. 2015, 174, 18–25. [Google Scholar] [CrossRef]
- Thomasen, J.R.; Willam, A.; Egger-Danner, C.; Sørensen, A.C. Reproductive technologies combine well with genomic selection in dairy breeding programs. J. Dairy Sci. 2016, 99, 1331–1340. [Google Scholar] [CrossRef]
- Maignel, L.; Boichard, D.; Verrier, E. Genetic variability of French dairy breeds estimated from pedigree information. Interbull Bull. 1996, 49. [Google Scholar]
- Le Mézec, P.; Danchin-Burge, C.; Moureaux, S. Davantage de diversité avec la génomique? plutôt non... Available online: http://idele.fr/contact/publication/idelesolr/recommends/les-programmes-de-selection-et-de-diffusion-de-taureaux-dia-a-lere-de-la-genomique-et-leurs-effets.html (accessed on 21 January 2019).
- Danchin-Burge, C.; Danvy, S.; Laloë, D.; Verrier, E. Création d’un observatoire de la VARiabilité génétique des RUMinants et des Equidés (VARUME). Innov. Agron. 2017, 55, 235–245. [Google Scholar]
- Eynard, S.E.; Windig, J.J.; Leroy, G.; van Binsbergen, R.; Calus, M. The effect of rare alleles on estimated genomic relationships from whole genome sequence data. BMC Genet. 2015, 16, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomasen, J.R.; Liu, H.; Sørensen, A.C. Genotyping more cows increases genetic gain and reduces rate of true inbreeding in a dairy cattle breeding scheme using female reproductive technologies. J. Dairy Sci. 2020, 103, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kardos, M.; Luikart, G.; Allendorf, F.W. Measuring individual inbreeding in the age of genomics: Marker-based measures are better than pedigrees. Heredity 2015, 115, 63–72. [Google Scholar] [CrossRef]
- Doekes, H.P.; Veerkamp, R.F.; Bijma, P.; Hiemstra, S.J.; Windig, J.J. Trends in genome-wide and region-specific genetic diversity in the Dutch-Flemish Holstein–Friesian breeding program from 1986 to 2015. Genet. Sel. Evol. 2018, 50, 15. [Google Scholar] [CrossRef] [Green Version]
- Forutan, M.; Ansari Mahyari, S.; Baes, C.; Melzer, N.; Schenkel, F.S.; Sargolzaei, M. Inbreeding and runs of homozygosity before and after genomic selection in North American Holstein cattle. BMC Genom. 2018, 19, 98. [Google Scholar] [CrossRef]
- Granleese, T.; Clark, S.A.; Kinghorn, B.P.; Werf, J.H.J. van der Optimizing female allocation to reproductive technologies considering merit, inbreeding and cost in nucleus breeding programmes with genomic selection. J. Anim. Breed. Genet. 2019, 136, 79–90. [Google Scholar] [CrossRef]
- Pedersen, L.D.; Kargo, M.; Berg, P.; Voergaard, J.; Buch, L.H.; Sørensen, A.C. Genomic selection strategies in dairy cattle breeding programmes: Sexed semen cannot replace multiple ovulation and embryo transfer as superior reproductive technology. J. Anim. Breed. Genet. 2012, 129, 152–163. [Google Scholar] [CrossRef]
- Pryce, J.E.; Goddard, M.E.; Raadsma, H.W.; Hayes, B.J. Deterministic models of breeding scheme designs that incorporate genomic selection. J. Dairy Sci. 2010, 93, 5455–5466. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, A.C.; Sørensen, M.K. Inbreeding Rates in Breeding Programs with Different Strategies for Using Genomic Selection. In Proceedings of the 2009 Interbull Meeting, Barcelona, Spain, 21–24 August 2009; p. 4. [Google Scholar]
- Thomasen, J.R.; Egger-Danner, C.; Willam, A.; Guldbrandtsen, B.; Lund, M.S.; Sørensen, A.C. Genomic selection strategies in a small dairy cattle population evaluated for genetic gain and profit. J. Dairy Sci. 2014, 97, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Land, R.B.; Hill, W.G. The possible use of superovulation and embryo transfer in cattle to increase response to selection. Anim. Sci. 1975, 21, 1–12. [Google Scholar] [CrossRef]
- Galli, C.; Crotti, G.; Notari, C.; Turini, P.; Duchi, R.; Lazzari, G. Embryo production by ovum pick up from live donors. Theriogenology 2001, 55, 1341–1357. [Google Scholar] [CrossRef]
- Pook, T. MoBPS: Simulation of Breeding Programs, version 1.0.2; R Package: Göttigen, Germany, 2018. [Google Scholar]
- Pook, T.; Schlather, M.; Simianer, H. MoBPS-Modular Breeding Program Simulator. G3 Genes Genomes Genet. 2020. [Google Scholar] [CrossRef] [Green Version]
- Arias, J.A.; Keehan, M.; Fisher, P.; Coppieters, W.; Spelman, R. A high density linkage map of the bovine genome. BMC Genet. 2009, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Hayes, B.; Goddard, M. The distribution of the effects of genes affecting quantitative traits in livestock. Genet. Sel. Evol. 2001, 33, 209–229. [Google Scholar] [CrossRef] [Green Version]
- Eynard, S.E.; Croiseau, P.; Laloë, D.; Fritz, S.; Calus, M.P.L.; Restoux, G. Which Individuals to Choose to Update the Reference Population? Minimizing the Loss of Genetic Diversity in Animal Genomic Selection Programs. G3 Bethesda Md 2018, 8, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Laloë, D.; Phocas, F. A proposal of criteria of robustness analysis in genetic evaluation. Livest. Prod. Sci. 2003, 80, 241–256. [Google Scholar] [CrossRef]
- Mardia, K.V.; Kent, J.T.; Bibby, J.M. Multivariate Analysis; Probability and Mathematical Statistics; Academic Press: London, UK, 1979; Volume 15. [Google Scholar]
- McQuillan, R.; Leutenegger, A.-L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of Homozygosity in European Populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [Green Version]
- de Cara, M.Á.R.; Villanueva, B.; Toro, M.Á.; Fernández, J. Using genomic tools to maintain diversity and fitness in conservation programmes. Mol. Ecol. 2013, 22, 6091–6099. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Purcell, S.; Chang, C. PLINK 1.9. Available online: www.cog-genomics.org/plink/1.9/ (accessed on 17 October 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: Estimated Marginal Means, aka Least-Squares Means, version 1.5.1; R package, 2020; Available online: https://CRAN.R-project.org/package=emmeans (accessed on 16 October 2020).
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. multcomp: Simultaneous Inference in General Parametric Models, version 1.4-14; R package, 2020; Available online: https://CRAN.R-project.org/package=multcomp (accessed on 16 October 2020).
- FAO. Secondary Guidelines for the National Farm Animal Genetic Resources Management Plans: Management of Small Populations at Risk; FAO: Rome, Italy, 2000; p. 219. [Google Scholar]
- AETE. Commercial Embryo Transfer Activity in Europe 2018. 2019. Available online: https://www.aete.eu/publications/statistics/ (accessed on 17 October 2020).
- Petyim, S.; Bage, R.; Forsberg, M.; Rodriguez-Martinez, H.; Larsson, B. The Effect of Repeated Follicular Puncture on Ovarian Function in Dairy Heifers. J. Vet. Med. Ser. A 2000, 47, 627–640. [Google Scholar] [CrossRef]
- Qi, M.; Yao, Y.; Ma, H.; Wang, J.; Zhao, X.; Liu, L.; Tang, X.; Zhang, L.; Zhang, S.; Sun, F. Transvaginal Ultrasound-guided Ovum Pick-up(OPU) in Cattle. J. Biomim. Biomater. Tissue Eng. 2013, 18, 118. [Google Scholar]
- Petyim, S.; Båge, R.; Madej, A.; Larsson, B. Ovum Pick-up in Dairy Heifers: Does it Affect Animal Well-being? Reprod. Domest. Anim. 2007, 42, 623–632. [Google Scholar] [CrossRef]
- Bérodier, M.; Berg, P.; Meuwissen, T.; Brochard, M.; Ducrocq, V. Improving mating plans at herd level using genomic information. In Proceedings of the Annual Meeting of the European Association for Animal Production (EAAP), Ghent, Belgium, 26–30 August 2019; p. 210. [Google Scholar]
- Meuwissen, T.H.E. Maximizing the response of selection with a predefined rate of inbreeding. J. Anim. Sci. 1997, 75, 934–940. [Google Scholar] [CrossRef]
- Granleese, T.; Clark, S.A.; Swan, A.A.; van der Werf, J.H.J. Increased genetic gains in sheep, beef and dairy breeding programs from using female reproductive technologies combined with optimal contribution selection and genomic breeding values. Genet. Sel. Evol. 2015, 47, 70. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Scenario | Total Number of Calves Born per Female Donor | Age of Female Donors at Birth of Their Embryo-Transfer Calves | Number of Distinct Sires of Bulls Mated with Each Female Donor | Number of Female Donors (Embryos or Oocytes) | Number of Sires of Bulls | Number of Marketed Bulls | |
|---|---|---|---|---|---|---|---|
| REF | No use of RT | / | / | / | 0 | 80 | 60 |
| A | Low intensity | 3 (1 flushing) | 26 months | 1 (1 per flushing) | 150 | 80 | 60 |
| B | Low intensity 300 | 3 (1 flushing) | 26 months | 1 (1 per flushing) | 300 | 80 | 60 |
| C | Medium intensity | 9 (3 flushings) | 26 months | 3 (1 per flushing) | 150 | 80 | 60 |
| D | Medium intensity 300 | 9 (3 flushings) | 26 months | 3 (1 per flushing) | 300 | 80 | 60 |
| E | High intensity MOET-like | 15 (5 flushings) | 26 months | 5 (1 per flushing) | 150 | 80 | 60 |
| F | High intensity 300 MOET-like | 15 (5 flushings) | 26 months | 5 (1 per flushing) | 300 | 80 | 60 |
| G | High intensity MOET-like, medium sires | 15 (5 flushings) | 26 months | 5 (1 per flushing) | 150 | 60 | 60 |
| H | High intensity MOET-like, medium sires, low bulls | 15 (5 flushings) | 26 months | 5 (1 per flushing) | 150 | 60 | 40 |
| I | High intensity MOET-like, low sires, low bulls | 15 (5 flushings) | 26 months | 5 (1 per flushing) | 150 | 40 | 40 |
| J | High intensity OPU-IVF-like | 15 | 26 months | 15 (1 per calf) | 150 | 80 | 60 |
| K | High intensity 300 OPU-IVF-like | 15 | 26 months | 15 (1 per calf) | 300 | 80 | 60 |
| L | High intensity MOET-like, short interval | 15 (5 flushings) | 14 months | 5 (1 per flushing) | 150 | 80 | 60 |
| M | High intensity OPU-IVF-like, short interval | 15 | 14 months | 15 (1 per calf) | 150 | 80 | 60 |
| N | High intensity OPU-IVF-like, short interval, low sires, low bulls | 15 | 14 months | 15 (1 per calf) | 150 | 40 | 40 |
| Scenario | Annual ROH-Based Inbreeding Rate in % [95% Confidence Interval] | Annual ROH-Based Inbreeding Rate Compared with Scenario REF | Annual Genetic Gain in TBV [95% Confidence Interval] | Annual Genetic Gain in TBV Compared with Scenario REF | |
|---|---|---|---|---|---|
| REF | No use of RT | 0.095 a [0.094;0.096] | 1.00 a | 0.298 a [0.297;0.298] | 1.00 a |
| A | Low intensity | 0.100 b [0.099;0.101] | 1.05 b | 0.323 b [0.322;0.323] | 1.08 b |
| B | Low intensity 300 | 0.103 c [0.102;0.104] | 1.08 c | 0.332 c [0.332;0.333] | 1.11 c |
| C | Medium intensity | 0.127 d [0.126;0.128] | 1.34 d | 0.364 d [0.363;0.364] | 1.22 d |
| D | Medium intensity 300 | 0.124 e [0.123;0.125] | 1.31 e | 0.381 e [0.381;0.382] | 1.28 e |
| E | High intensity MOET-like | 0.140 f [0.139;0.141] | 1.47 f | 0.384 f [0.384;0.384] | 1.29 f |
| F | High intensity 300 MOET-like | 0.134 g [0.133;0.135] | 1.41 g | 0.403 g [0.403;0.403] | 1.35 g |
| G | High intensity MOET-like, medium sires | 0.146 h [0.145;0.147] | 1.54 h | 0.373 h [0.372;0.373] | 1.25 h |
| H | High intensity MOET-like, medium sires, low bulls | 0.155 i [0.154;0.156] | 1.63 i | 0.389 i [0.389;0.389] | 1.31 i |
| I | High intensity MOET-like, low sires, low bulls | 0.152 j [0.151;0.153] | 1.60 j | 0.365 j [0.365;0.366] | 1.22 j |
| J | High intensity OPU-IVF-like | 0.135 g [0.134;0.136] | 1.42 g | 0.386 k [0.386;0.386] | 1.30 k |
| K | High intensity 300 OPU-IVF-like | 0.146 h [0.145;0.147] | 1.54 h | 0.407 l [0.407;0.407] | 1.37 l |
| L | High intensity MOET-like, short interval | 0.196 k [0.195;0.197] | 2.06 k | 0.438 m [0.438;0.439] | 1.47 m |
| M | High intensity OPU-IVF-like, short interval | 0.196 k [0.195;0.197] | 2.06 k | 0.440 n [0.440;0.440] | 1.48 n |
| N | High intensity OPU-IVF-like, short interval, low sires, low bulls | 0.219 l [0.218;0.220] | 2.31 l | 0.415 o [0.415;0.416] | 1.39 o |
| Scenario | Annual ROH-based Inbreeding Rate in % [95% Confidence Interval] | Annual ROH-based Inbreeding Rate Compared with Scenario REF | Annual Genetic Gain in TBV [95% Confidence Interval] | Annual Genetic Gain in TBV Compared with Scenario REF | |
|---|---|---|---|---|---|
| REF | No use of RT | 0.04 a [0.038;0.056] | 1.00 a | 0.280 a [0.277;0.282] | 1.00 a |
| A | Low intensity | 0.050 a [0.041;0.058] | 1.05 a | 0.303 b [0.301;0.305] | 1.08 b |
| B | Low intensity 300 | 0.057 ab [0.048;0.066] | 1.21 ab | 0.314 c [0.312;0.316] | 1.12 c |
| C | Medium intensity | 0.076 b [0.067;0.084] | 1.60 b | 0.340 d [0.338;0.342] | 1.21 d |
| D | Medium intensity 300 | 0.078 b [0.069;0.086] | 1.65 b | 0.357 e [0.355;0.359] | 1.28 e |
| E | High intensity MOET-like | 0.109 c [0.100;0.117] | 2.31 c | 0.356 e [0.354;0.359] | 1.27 e |
| F | High intensity 300 MOET-like | 0.109 c [0.100;0.117] | 2.31 c | 0.374 f [0.372;0.376] | 1.34 f |
| G | High intensity MOET-like, medium sires | 0.108 c [0.098;0.118] | 2.29 c | 0.344 d [0.342;0.347] | 1.23 d |
| H | High intensity MOET-like, medium sires, low bulls | 0.117 c [0.107;0.127] | 2.48 c | 0.361 e [0.359;0.364] | 1.29 e |
| I | High intensity MOET-like, low sires, low bulls | 0.130 c [0.118;0.142] | 2.76 c | 0.340 d [0.337;0.343] | 1.21 d |
| J | High intensity OPU-IVF-like | 0.112 c [0.103;0.120] | 2.38 c | 0.359 e [0.357;0.361] | 1.28 e |
| K | High intensity 300 OPU-IVF-like | 0.116 c [0.107;0.125] | 2.46 c | 0.380 g [0.378;0.382] | 1.36 g |
| L | High intensity MOET-like, short interval | 0.200 d [0.191;0.209] | 4.25 d | 0.404 h [0.402;0.406] | 1.44 h |
| M | High intensity OPU-IVF-like, short interval | 0.204 d [0.195;0.212] | 4.33 d | 0.405 h [0.403;0.407] | 1.45 h |
| N | High intensity OPU-IVF-like, short interval, low sires, low bulls | 0.220 d [0.208;0.232] | 4.67 d | 0.380 fg [0.377;0.383] | 1.36 fg |
| Number of Sires and Bulls | Annual ROH-Based Inbreeding Rate Compared with Scenario REF | Annual Genetic Gain in TBV Compared with Scenario REF | ||
| MOET, Medium Interval | OPU-IVF, Short Interval | MOET, Medium Interval | OPU-IVF, Short Interval | |
| 80 sires of bulls and 60 marketed bulls | Scenario E 1.47 for cows 2.31 for sires/bulls | Scenario M 2.06 for cows 4.33 for sires/bulls | Scenario E 1.29 for cows 1.27 for sires/bulls | Scenario M 1.48 for cows 1.45 for sires/bulls |
| 40 sires of bulls and 40 marketed bulls | Scenario I 1.60 for cows 2.76 for sires/bulls | Scenario N 2.31 for cows 4.67 for sires/bulls | Scenario I 1.22 for cows 1.21 for sires/bulls | Scenario N 1.39 for cows 1.36 for sires/bulls |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doublet, A.-C.; Restoux, G.; Fritz, S.; Balberini, L.; Fayolle, G.; Hozé, C.; Laloë, D.; Croiseau, P. Intensified Use of Reproductive Technologies and Reduced Dimensions of Breeding Schemes Put Genetic Diversity at Risk in Dairy Cattle Breeds. Animals 2020, 10, 1903. https://doi.org/10.3390/ani10101903
Doublet A-C, Restoux G, Fritz S, Balberini L, Fayolle G, Hozé C, Laloë D, Croiseau P. Intensified Use of Reproductive Technologies and Reduced Dimensions of Breeding Schemes Put Genetic Diversity at Risk in Dairy Cattle Breeds. Animals. 2020; 10(10):1903. https://doi.org/10.3390/ani10101903
Chicago/Turabian StyleDoublet, Anna-Charlotte, Gwendal Restoux, Sébastien Fritz, Laura Balberini, Guillaume Fayolle, Chris Hozé, Denis Laloë, and Pascal Croiseau. 2020. "Intensified Use of Reproductive Technologies and Reduced Dimensions of Breeding Schemes Put Genetic Diversity at Risk in Dairy Cattle Breeds" Animals 10, no. 10: 1903. https://doi.org/10.3390/ani10101903