Yogurt Produced by Novel Natural Starter Cultures Improves Gut Epithelial Barrier In Vitro

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Strains Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 and Growth Conditions
2.2. Physiological, Biochemical, and Technological Characterization of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains
2.3. Safety Assessment
2.3.1. Hemolytic and Gelatinase Activity Assays
2.3.2. Antibiotic Susceptibility Testing
2.4. Antimicrobial Activity
2.5. Proteolytic Activity
2.6. Yogurt Manufacturing
2.7. Survival in Simulated Gastrointestinal Tract Conditions
2.8. Cell Culture and Treatments
2.9. Cytotoxicity Assay
2.10. RNA Isolation and qPCR
2.11. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains
3.2. Safety Assessment of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains
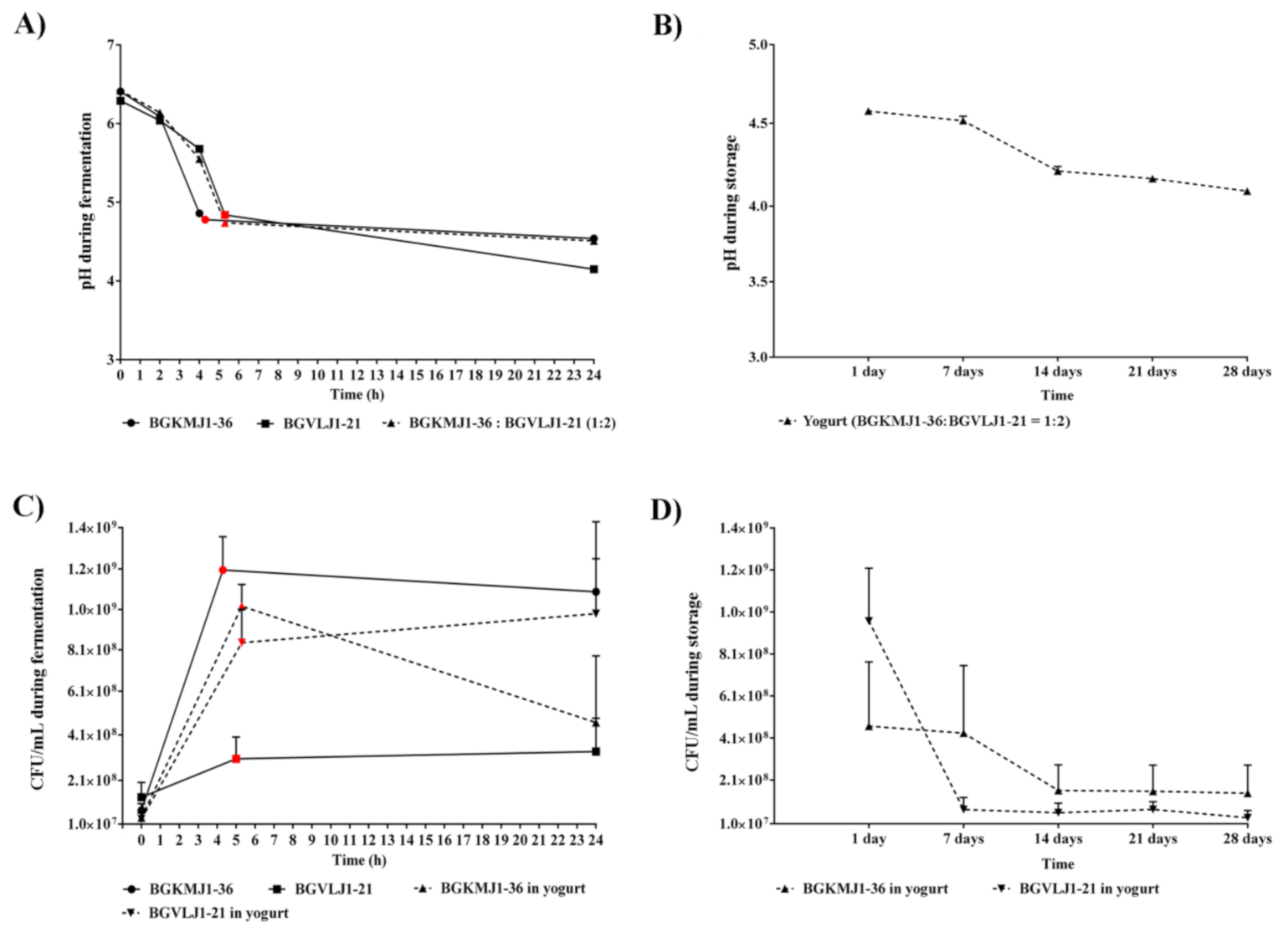
3.3. Acidifying Kinetics of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains During Milk Fermentation and Yogurt Storage
3.4. Growth Kinetics of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains During Milk Fermentation and Storage
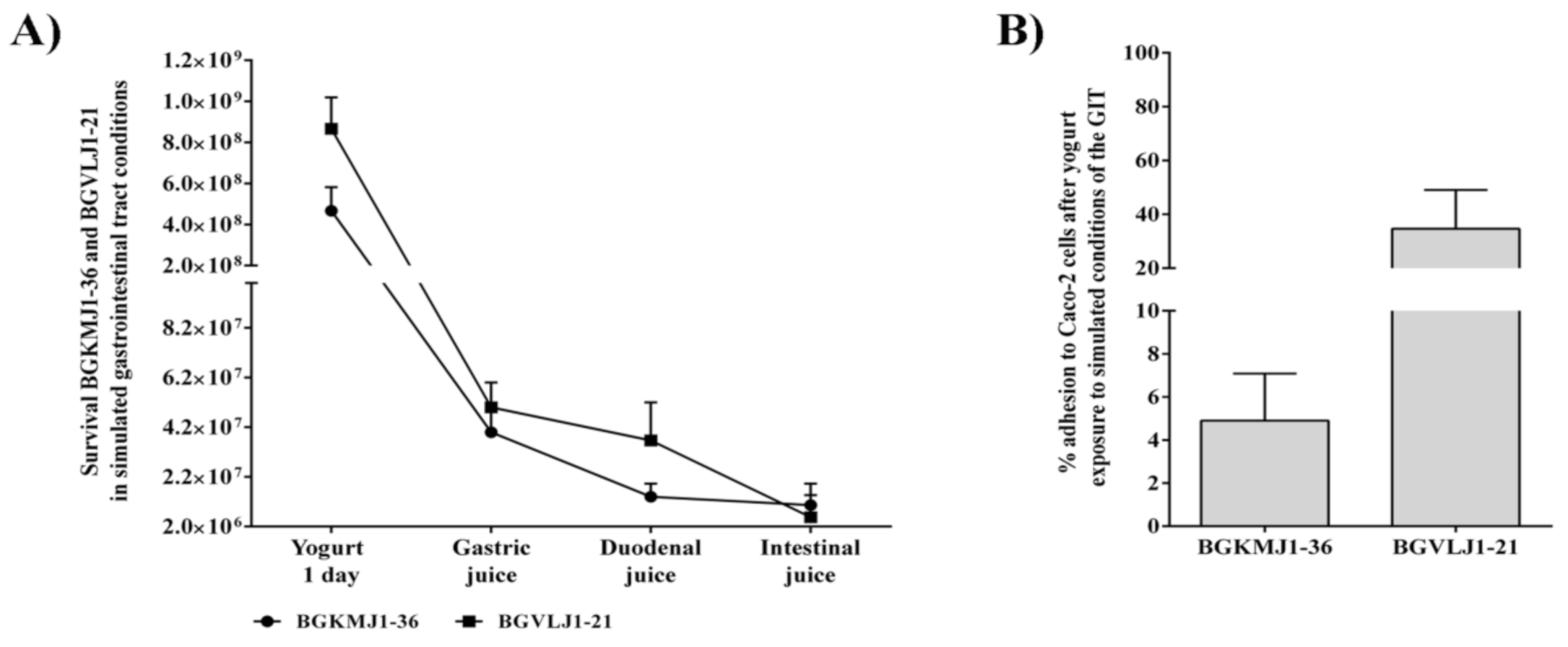
3.5. Effects of Simulated Gastrointestinal Tract Conditions on the Viability of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains in Yogurt
3.6. Adhesion of Streptococcus thermophilus BGKMJ1-36 and Lactobacillus bulgaricus BGVLJ1-21 Strains to Caco-2 Cells After Simulated Gastrointestinal Tract Conditions
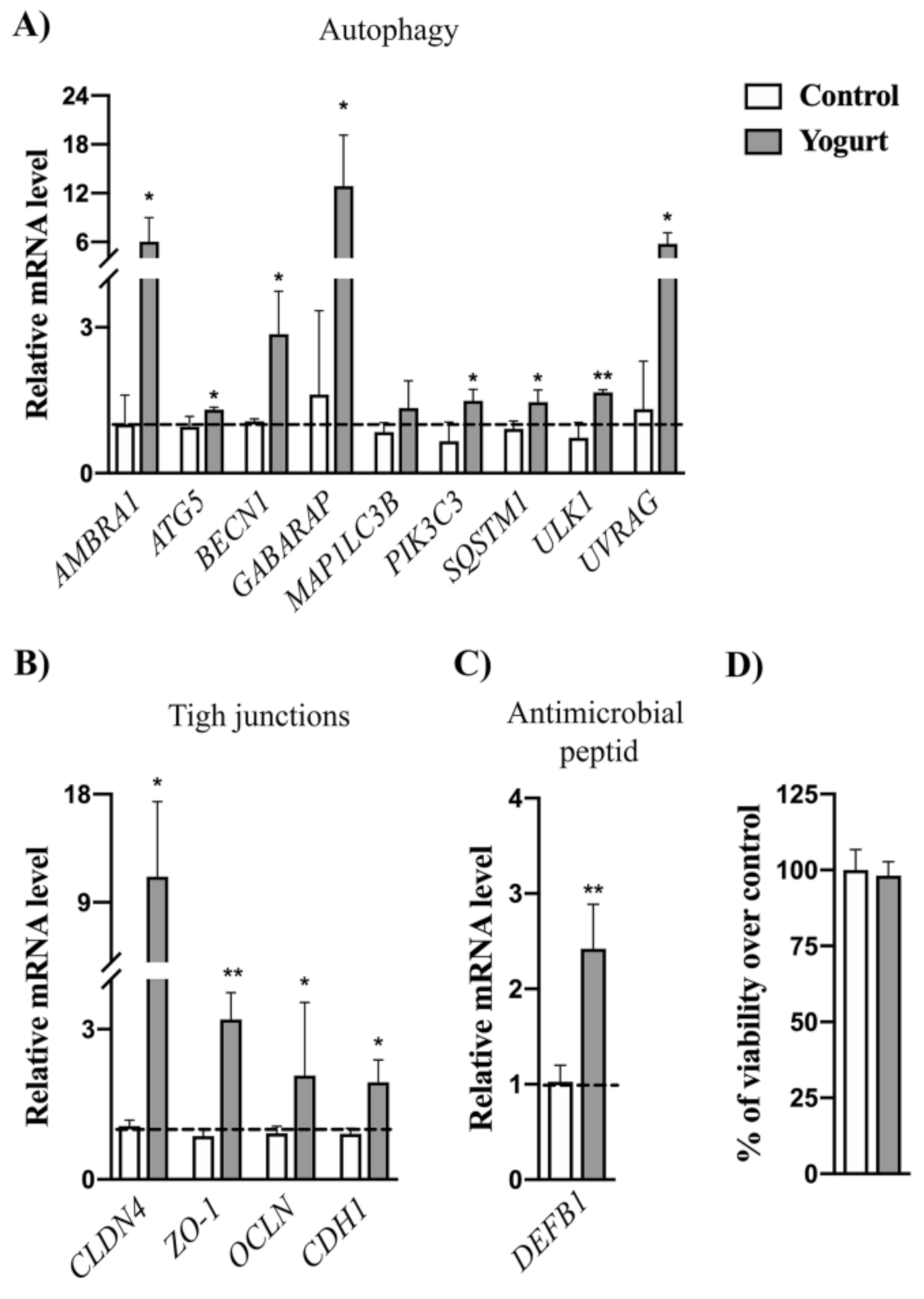
3.7. Gene Expression Analysis Revealed the Upregulation of Autophagy and Epithelial Barrier Defense Markers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shiby, V.K.; Mishra, H.N. Fermented milks and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L. Yogurt, living cultures, and gut health. Am. J. Clin. Nutr. 2014, 99, 1248S–1250S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zhao, S.; Hao, G.; Yu, H.; Tian, H.; Zhao, G. Role of lactic acid bacteria on the yogurt flavour: A review. Int. J. Food Prop. 2017, 20, S316–S330. [Google Scholar] [CrossRef] [Green Version]
- Ali, K.; Mehmood, M.H.; Iqbal, M.A.; Masud, T.; Qazalbash, M.; Saleem, S.; Ahmed, S.; Tariq, M.R.; Safdar, W.; Nasir, M.A.; et al. Isolation and characterization of exopolysaccharide-producing strains of Lactobacillus bulgaricus from curd. Food Sci. Nutr. 2019, 7, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- El-Abbadi, N.H.; Dao, M.C.; Meydani, S.N. Yogurt: Role in healthy and active aging. Am. J. Clin. Nutr. 2014, 99, 1263S–1270S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryana, K.J.; Olson, D.W. A 100-Year Review: Yogurt and other cultured dairy products. J. Dairy Sci. 2017, 100, 9987–10013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef] [PubMed]
- Saborido, R.; Leis, R. [Yogurt and dietary recommendations for lactose intolerance]. Nutr. Hosp. 2018, 35, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Araya, M.; Gopal, P.; Lindgren, S.E.; Lodi, R.; Oliver, G.; Saxelin, M.L.; Severin, A.L.; Stanton, C. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation, Food and Agriculture Organization of the United Nations; World Health Organization: Rome, Italy, 2006; pp. 2–3. ISBN 978-92-5-105513-7. [Google Scholar]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to live yoghurt cultures and improved lactose digestion (ID 1143, 2976) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1763. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Iravani, S.; Korbekandi, H.; Mirmohammadi, S.V. Technology and potential applications of probiotic encapsulation in fermented milk products. J. Food Sci. Technol. 2015, 52, 4679–4696. [Google Scholar] [CrossRef] [PubMed]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [Green Version]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: Reiterating What They Are and What They Are Not. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, K. An overview of lactic acid bacteria. Int. J. Biosci. (IJB) 2011, 1, 1–13. [Google Scholar]
- Guarino, A.; Guandalini, S.; Lo Vecchio, A. Probiotics for Prevention and Treatment of Diarrhea. J. Clin. Gastroenterol. 2015, 49 (Suppl. 1), S37–S45. [Google Scholar] [CrossRef]
- Lisko, D.J.; Johnston, G.P.; Johnston, C.G. Effects of Dietary Yogurt on the Healthy Human Gastrointestinal (GI) Microbiome. Microorganisms 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, T.; Sequoia, J. Probiotics for Gastrointestinal Conditions: A Summary of the Evidence. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar]
- Yoon, H.; Park, Y.S.; Lee, D.H.; Seo, J.-G.; Shin, C.M.; Kim, N. Effect of administering a multi-species probiotic mixture on the changes in fecal microbiota and symptoms of irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. J. Clin. Biochem. Nutr. 2015, 57, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Sokovic Bajic, S.; Djokic, J.; Dinic, M.; Veljovic, K.; Golic, N.; Mihajlovic, S.; Tolinacki, M. GABA-Producing Natural Dairy Isolate From Artisanal Zlatar Cheese Attenuates Gut Inflammation and Strengthens Gut Epithelial Barrier in vitro. Front. Microbiol. 2019, 10, 527. [Google Scholar] [CrossRef]
- Bajić, S.S.; Đokić, J.; Dinić, M.; Tomić, S.; Popović, N.; Brdarić, E.; Golić, N.; Tolinački, M. GABA potentiate the immunoregulatory effects of Lactobacillus brevis BGZLS10-17 via ATG5-dependent autophagy in vitro. Sci. Rep. 2020, 10, 1347. [Google Scholar] [CrossRef] [PubMed]
- Dinić, M.; Lukić, J.; Djokić, J.; Milenković, M.; Strahinić, I.; Golić, N.; Begović, J. Lactobacillus fermentum Postbiotic-induced Autophagy as Potential Approach for Treatment of Acetaminophen Hepatotoxicity. Front. Microbiol. 2017, 8, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bluma, A.; Ciprovica, I.; Sabovics, M. The influence of non-starter lactic acid bacteria on swiss-type cheese quality. Agric. Food 2017, 5, 34–41. [Google Scholar]
- Hansen, E.B. Commercial bacterial starter cultures for fermented foods of the future. Int. J. Food Microbiol. 2002, 78, 119–131. [Google Scholar] [CrossRef]
- Johansen, E. Use of Natural Selection and Evolution to Develop New Starter Cultures for Fermented Foods. Annu. Rev. Food Sci. Technol. 2018, 9, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Terzić-Vidojević, A.; Veljović, K.; Tolinački, M.; Živković, M.; Lukić, J.; Lozo, J.; Fira, Đ.; Jovčić, B.; Strahinić, I.; Begović, J.; et al. Diversity of non-starter lactic acid bacteria in autochthonous dairy products from Western Balkan Countries — Technological and probiotic properties. Food Res. Int. 2020, 136, 109494. [Google Scholar] [CrossRef] [PubMed]
- Kempler, G.M.; McKay, L.L. Biochemistry and Genetics of Citrate Utilization in Streptococcus lactis ssp. diacetylactis. J. Dairy Sci. 1981, 64, 1527–1539. [Google Scholar] [CrossRef]
- Zourari, A.; Roger, S.; Chabanet, C.; Desmazeaud, M. Caractérisation de bactéries lactiques thermophiles isolées de yaourts artisanaux grecs. I. Souches de Streptococcus salivarius subsp thermophilus. Le Lait 1991, 71, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Nikolic, M.; López, P.; Strahinic, I.; Suárez, A.; Kojic, M.; Fernández-García, M.; Topisirovic, L.; Golic, N.; Ruas-Madiedo, P. Characterisation of the exopolysaccharide (EPS)-producing Lactobacillus paraplantarum BGCG11 and its non-EPS producing derivative strains as potential probiotics. Int. J. Food Microbiol. 2012, 158, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, A.S.; Omar, N.B.; Abriouel, H.; López, R.L.; Veljovic, K.; Cañamero, M.M.; Topisirovic, M.K.L.; Gálvez, A. Virulence factors, antibiotic resistance, and bacteriocins in enterococci from artisan foods of animal origin. Food Control. 2009, 20, 381–385. [Google Scholar] [CrossRef]
- Lopes, M.d.F.S.; Simões, A.P.; Tenreiro, R.; Marques, J.J.F.; Crespo, M.T.B. Activity and expression of a virulence factor, gelatinase, in dairy enterococci. Int. J. Food Microbiol. 2006, 112, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Hürlimann, L.M.; Corradi, V.; Hohl, M.; Bloemberg, G.V.; Tieleman, D.P.; Seeger, M.A. The Heterodimeric ABC Transporter EfrCD Mediates Multidrug Efflux in Enterococcus faecalis. Antimicrob. Agents Chemother. 2016, 60, 5400–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golić, N.; Veljović, K.; Popović, N.; Djokić, J.; Strahinić, I.; Mrvaljević, I.; Terzić-Vidojević, A. In vitro and in vivo antagonistic activity of new probiotic culture against Clostridium difficile and Clostridium perfringens. BMC Microbiol. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Tagg, J.R.; McGiven, A.R. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef] [Green Version]
- Kojic, M.; Fira, D.; Banina, A.; Topisirovic, L. Characterization of the Cell Wall-Bound Proteinase of Lactobacillus casei HN14. Appl. Environ. Microbiol. 1991, 57, 1753–1757. [Google Scholar] [CrossRef] [Green Version]
- Dirar, H.A. The Indigenous Fermented Foods of the Sudan: A Study in African Food and Nutrition; CAB International Publishing: Wallingford, UK, 1993; ISBN 978-0-85198-858-0. [Google Scholar]
- Sánchez, B.; Fernández-García, M.; Margolles, A.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Technological and probiotic selection criteria of a bile-adapted Bifidobacterium animalis subsp. lactis strain. Int. Dairy J. 2010, 20, 800–805. [Google Scholar] [CrossRef] [Green Version]
- Živković, M.; Miljković, M.S.; Ruas-Madiedo, P.; Markelić, M.B.; Veljović, K.; Tolinački, M.; Soković, S.; Korać, A.; Golić, N. EPS-SJ Exopolisaccharide Produced by the Strain Lactobacillus paracasei subsp. paracasei BGSJ2-8 Is Involved in Adhesion to Epithelial Intestinal Cells and Decrease on E. coli Association to Caco-2 Cells. Front. Microbiol. 2016, 7, 286. [Google Scholar] [CrossRef] [Green Version]
- Lukic, J.; Strahinic, I.; Milenkovic, M.; Golic, N.; Kojic, M.; Topisirovic, L.; Begovic, J. Interaction of Lactobacillus fermentum BGHI14 with rat colonic mucosa: Implications for colitis induction. Appl. Environ. Microbiol. 2013, 79, 5735–5744. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Shi, Y.; Guo, X.; Wang, S.; Ouyang, Y.; Hao, M.; Liu, D.; Qiao, L.; Li, N.; Zheng, J.; et al. CHOP mediates ASPP2-induced autophagic apoptosis in hepatoma cells by releasing Beclin-1 from Bcl-2 and inducing nuclear translocation of Bcl-2. Cell Death Dis. 2014, 5, e1323. [Google Scholar] [CrossRef]
- He, Z.; Liu, H.; Agostini, M.; Yousefi, S.; Perren, A.; Tschan, M.P.; Mak, T.W.; Melino, G.; Simon, H.U. p73 regulates autophagy and hepatocellular lipid metabolism through a transcriptional activation of the ATG5 gene. Cell Death Differ. 2013, 20, 1415–1424. [Google Scholar] [CrossRef] [Green Version]
- Sahani, M.H.; Itakura, E.; Mizushima, N. Expression of the autophagy substrate SQSTM1/p62 is restored during prolonged starvation depending on transcriptional upregulation and autophagy-derived amino acids. Autophagy 2014, 10, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Elamin, E.; Jonkers, D.; Juuti-Uusitalo, K.; van IJzendoorn, S.; Troost, F.; Duimel, H.; Broers, J.; Verheyen, F.; Dekker, J.; Masclee, A. Effects of Ethanol and Acetaldehyde on Tight Junction Integrity: In Vitro Study in a Three Dimensional Intestinal Epithelial Cell Culture Model. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.; McGhie, T.K.; Bentley-Hewitt, K.; Christeller, J. PPARγ as a sensor of lipase activity and a target for the lipase inhibitor orlistat. Lipids Health Dis. 2013, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Freudenberger, T.; Deenen, R.; Kretschmer, I.; Zimmermann, A.; Seiler, L.F.; Mayer, P.; Heim, H.-K.; Köhrer, K.; Fischer, J.W. Synthetic gestagens exert differential effects on arterial thrombosis and aortic gene expression in ovariectomized apolipoprotein E-deficient mice. Br. J. Pharmacol. 2014, 171, 5032–5048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laquerriere, P.; Grandjean-Laquerriere, A.; Addadi-Rebbah, S.; Jallot, E.; Laurent-Maquin, D.; Frayssinet, P.; Guenounou, M. MMP-2, MMP-9 and their inhibitors TIMP-2 and TIMP-1 production by human monocytes in vitro in the presence of different forms of hydroxyapatite particles. Biomaterials 2004, 25, 2515–2524. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wu, X.; Zhou, Q.; Yang, Y.; Tian, Y.; Huang, C.; Meng, X.; Li, J. PICK1 confers anti-inflammatory effects in acute liver injury via suppressing M1 macrophage polarization. Biochimie 2016, 127, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouser, L.; Paudyal, B.; Kaur, A.; Stenbeck, G.; Jones, L.A.; Abozaid, S.M.; Stover, C.M.; Flahaut, E.; Sim, R.B.; Kishore, U. Human Properdin Opsonizes Nanoparticles and Triggers a Potent Pro-inflammatory Response by Macrophages without Involving Complement Activation. Front. Immunol. 2018, 9, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angrisano, T.; Pero, R.; Peluso, S.; Keller, S.; Sacchetti, S.; Bruni, C.B.; Chiariotti, L.; Lembo, F. LPS-induced IL-8 activation in human intestinal epithelial cells is accompanied by specific histone H3 acetylation and methylation changes. BMC Microbiol. 2010, 10, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragicevic, S.; Milosevic, K.; Nestorovic, B.; Nikolic, A. Influence of the Polymorphism C-509T in the TGFB1 Gene Promoter on the Response to Montelukast. Pediatric Allergy Immunol. Pulmonol. 2017, 30, 239–245. [Google Scholar] [CrossRef]
- Egan, K.; Field, D.; Rea, M.C.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocins: Novel Solutions to Age Old Spore-Related Problems? Front. Microbiol. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuffia, F.; Bergamini, C.V.; Wolf, I.V.; Hynes, E.R.; Perotti, M.C. Characterization of volatile compounds produced by Lactobacillus helveticus strains in a hard cheese model. Food Sci. Technol. Int. 2018, 24, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Xu, P. Acetoin metabolism in bacteria. Crit. Rev. Microbiol. 2007, 33, 127–140. [Google Scholar] [CrossRef]
- Hols, P.; Hancy, F.; Fontaine, L.; Grossiord, B.; Prozzi, D.; Leblond-Bourget, N.; Decaris, B.; Bolotin, A.; Delorme, C.; Dusko Ehrlich, S.; et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol. Rev. 2005, 29, 435–463. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, C.; Li, S.; Zhang, Y.; Yang, Z. Growth and exopolysaccharide production by Streptococcus thermophilus ST1 in skim milk. Braz. J. Microbiol. 2011, 42, 1470–1478. [Google Scholar] [CrossRef] [Green Version]
- Vaningelgem, F.; Zamfir, M.; Adriany, T.; De Vuyst, L. Fermentation conditions affecting the bacterial growth and exopolysaccharide production by Streptococcus thermophilus ST 111 in milk-based medium. J. Appl. Microbiol. 2004, 97, 1257–1273. [Google Scholar] [CrossRef]
- Nordmark, E.-L.; Yang, Z.; Huttunen, E.; Widmalm, G. Structural studies of an exopolysaccharide produced by Streptococcus thermophilus THS. Biomacromolecules 2005, 6, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Marty, E.; Bodenmann, C.; Buchs, J.; Hadorn, R.; Eugster-Meier, E.; Lacroix, C.; Meile, L. Prevalence of antibiotic resistance in coagulase-negative staphylococci from spontaneously fermented meat products and safety assessment for new starters. Int. J. Food Microbiol. 2012, 159, 74–83. [Google Scholar] [CrossRef]
- Kastner, S.; Perreten, V.; Bleuler, H.; Hugenschmidt, G.; Lacroix, C.; Meile, L. Antibiotic susceptibility patterns and resistance genes of starter cultures and probiotic bacteria used in food. Syst. Appl. Microbiol. 2006, 29, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Chamba, J.F.; Jamet, E. Contribution to the safety assessment of technological microflora found in fermented dairy products. Int. J. Food Microbiol. 2008, 126, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Delgado, S.; Mayo, B.; Ruas-Madiedo, P.; Margolles, A.; de los Reyes-Gavilán, C.G. Viability and diversity of probiotic Lactobacillus and Bifidobacterium populations included in commercial fermented milks. Food Res. Int. 2004, 37, 839–850. [Google Scholar] [CrossRef]
- Saarela, M.; Mogensen, G.; Fondén, R.; Mättö, J.; Mattila-Sandholm, T. Probiotic bacteria: Safety, functional and technological properties. J. Biotechnol. 2000, 84, 197–215. [Google Scholar] [CrossRef]
- Senaka Ranadheera, C.; Evans, C.A.; Adams, M.C.; Baines, S.K. Probiotic viability and physico-chemical and sensory properties of plain and stirred fruit yogurts made from goat’s milk. Food Chem. 2012, 135, 1411–1418. [Google Scholar] [CrossRef]
- Dave, R.I.; Shah, N.P. Viability of yoghurt and probiotic bacteria in yoghurts made from commercial starter cultures. Int. Dairy J. 1997, 7, 31–41. [Google Scholar] [CrossRef]
- Gilliland, S.E.; Reilly, S.S.; Kim, G.B.; Kim, H.S. Viability during storage of selected probiotic lactobacilli and bifidobacteria in a yogurt-like product. J. Food Sci. 2002, 67, 3091–3095. [Google Scholar] [CrossRef]
- Riazi, A.; Ziar, H. Growth and viability of yogurt starter organisms in honey-sweetened skimmed milk. Afr. J. Biotechnol. 2008, 7, 2055–2063. [Google Scholar]
- Soni, R.; Jain, N.K.; Shah, V.; Soni, J.; Suthar, D.N.; Gohel, P. Development of probiotic yogurt: Effect of strain combination on nutritional, rheological, organoleptic and probiotic properties. J. Food Sci. Technol. 2020, 57, 2038–2050. [Google Scholar] [CrossRef]
- Fernández, M.F.; Boris, S.; Barbés, C. Probiotic properties of human lactobacilli strains to be used in the gastrointestinal tract. J. Appl. Microbiol. 2003, 94, 449–455. [Google Scholar] [CrossRef]
- Tarrah, A.; da Silva Duarte, V.; de Castilhos, J.; Pakroo, S.; Lemos Junior, W.J.F.; Luchese, R.H.; Fioravante Guerra, A.; Rossi, R.C.; Righetto Ziegler, D.; Corich, V.; et al. Probiotic potential and biofilm inhibitory activity of Lactobacillus casei group strains isolated from infant feces. J. Funct. Foods 2019, 54, 489–497. [Google Scholar] [CrossRef]
- Sánchez, A.H.; Rejano, L.; Montaño, A.; de Castro, A. Utilization at high pH of starter cultures of lactobacilli for Spanish-style green olive fermentation. Int. J. Food Microbiol. 2001, 67, 115–122. [Google Scholar] [CrossRef]
- Aziz, K.; Haseeb Zaidi, A.; Fatima, H.N.; Tariq, M. Lactobacillus fermentum strains of dairy-product origin adhere to mucin and survive digestive juices. J. Med. Microbiol. 2019, 68, 1771–1786. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.C.R.; Aynaou, A.-E.; Albrecht, S.; Schols, H.A.; De Martinis, E.C.P.; Zoetendal, E.G.; Venema, K.; Saad, S.M.I.; Smidt, H. In vitro evaluation of gastrointestinal survival of Lactobacillus amylovorus DSM 16698 alone and combined with galactooligosaccharides, milk and/or Bifidobacterium animalis subsp. lactis Bb-12. Int. J. Food Microbiol. 2011, 149, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Elizaquível, P.; Sánchez, G.; Salvador, A.; Fiszman, S.; Dueñas, M.T.; López, P.; Fernández de Palencia, P.; Aznar, R. Evaluation of yogurt and various beverages as carriers of lactic acid bacteria producing 2-branched (1,3)-β-D-glucan. J. Dairy Sci. 2011, 94, 3271–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrov, Z.; Gotova, I.; Chorbadjiyska, E. In vitro characterization of the adhesive factors of selected probiotics to Caco-2 epithelium cell line. Biotechnol. Biotechnol. Equip. 2014, 28, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Darilmaz, D.O.; Aslım, B.; Suludere, Z.; Akca, G. Influence of gastrointestinal system conditions on adhesion of exopolysaccharide-producing Lactobacillus delbrueckii subsp. bulgaricus strains to caco-2 cells. Braz. Arch. Biol. Technol. 2011, 54, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Fernández de Palencia, P.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic strains: Survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. Eur. Food Res. Technol. 2008, 227, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.; Grondin, J.; Banskota, S.; Khan, W.I. Autophagy: Roles in intestinal mucosal homeostasis and inflammation. J. Biomed. Sci. 2019, 26, 19. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Pascual, C.; Klionsky, D.J. Autophagy: Machinery and regulation. Microb. Cell 2016, 3, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, X.; Zhang, L.; Xu, S.; Zhang, Q.; Lu, R. A surface-layer protein from Lactobacillus acidophilus NCFM induces autophagic death in HCT116 cells requiring ROS-mediated modulation of mTOR and JNK signaling pathways. Food Funct. 2019, 10, 4102–4112. [Google Scholar] [CrossRef] [PubMed]
- Nighot, P.K.; Hu, C.-A.A.; Ma, T.Y. Autophagy enhances intestinal epithelial tight junction barrier function by targeting claudin-2 protein degradation. J. Biol. Chem. 2015, 290, 7234–7246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, C.; Zhang, D.; Zhang, J.; Wang, X.; Geng, C. The altered tight junctions: An important gateway of bacterial translocation in cachexia patients with advanced gastric cancer. J. Interferon Cytokine Res. 2014, 34, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Popović, N.; Djokić, J.; Brdarić, E.; Dinić, M.; Terzić-Vidojević, A.; Golić, N.; Veljović, K. The Influence of Heat-Killed Enterococcus faecium BGPAS1-3 on the Tight Junction Protein Expression and Immune Function in Differentiated Caco-2 Cells Infected With Listeria monocytogenes ATCC 19111. Front. Microbiol. 2019, 10, 412. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta 2009, 1788, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.L.; Reardon, C.; Wang, A.; Nazli, A.; McKay, D.M. Transforming Growth Factor-β Regulation of Epithelial Tight Junction Proteins Enhances Barrier Function and Blocks Enterohemorrhagic Escherichia coli O157:H7-Induced Increased Permeability. Am. J. Pathol. 2005, 167, 1587–1597. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Kobatake, E.; Kabuki, T. S-Layer Protein of Lactobacillus helveticus SBT2171 Promotes Human β-Defensin 2 Expression via TLR2-JNK Signaling. Front. Microbiol. 2019, 10, 2414. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number of Gene | Primer Sequence 5′–3′ | Reference |
|---|---|---|---|
| AMBRA1_F | GGTGGGAGGAGAGGGGATAG | This work | |
| AMBRA1_R | NM_017749.3 | CGAGGGGCATGTCATCATTT | |
| ULK1_F | TTTTGTTTCTCCGTTGGGGC | This work | |
| ULK1_R | NM_003565.4 | ACTCTTCCCGGGCTGCTAAT | |
| UVRAG_F | AGGAAGGAGTGCACTGCAAA | This work | |
| UVRAG_R | NM_001386671.1 | AGGCAACTTGACACCGCATA | |
| GABARAP_F | CCCTCGTCCCGCTGATTTTA | This work | |
| GABARAP_R | NM_007278.2 | ATCCCTCCAGCTTGTACCCA | |
| PIK3C3_F | GCTGTCCTGGAAGACCCAAT | This work | |
| PIK3C3_R | NM_002647.4 | TTCTCACTGGCAAGGCCAAA | |
| MAP1LC3B_F | TTCAGGTTCACAAAACCCGC | This work | |
| MAP1LC3B_R | NM_022818.5 | TCTCACACAGCCCGTTTACC | |
| BECN1_F | CTGGGACAACAAGTTTGACCAT | [44] | |
| BECN1_R | GCTCCTCAGAGTTAAACTGGGTT | ||
| ATG5_F | CACAAGCAACTCTGGATGGGATTG | [45] | |
| ATG5_R | GCAGCCAC GGACGAAACAG | ||
| SQSTM1_F | GCCAGAGGAACAGATGGAGT | [46] | |
| SQSTM1_R | TCCGATTCTG GCATCTGTAG | ||
| CLDN4_F | ACAGACAAGCCTTACTCC | [21] | |
| CLDN4_R | GGAAGAACAAAGCAGAG | ||
| ZO-1_F | AGGGGCAGTGGTGGTTTTCTGTTCTTTC | [47] | |
| ZO-1_R | GCAGAGGTCAAAGTTCAAGGCTCAAGAGG | ||
| OCLN_F | TCAGGGAATATCCACCTATCACTTCAG | [21] | |
| OCLN_R | CATCAGCAGCAGCCATGTACTCTTCAC | ||
| CDH1_F | AGCCTGTCGAAGCAGGATTG | This work | |
| CDH1_R | NM_004360.5 | AGAAACAGCAAGAGCAGCAGA | |
| DEFB1_F | TGTCTGAGATGGCCTCAGGT | [48] | |
| DEFB1_R | GGGCAGGCAGAATAGAGACA | ||
| GAPDH_F | GTGAAGGTCGGAGTCAACG | [49] | |
| GAPDH_R | TGAGGTCAATGAAGGGGTC | ||
| IL-1β_F | TACGAATCTCCGACCACCACTACG | [50] | |
| IL-1β_R | GTACAGGTGCATCGTGCACATAAGC | ||
| IL-6_F | CACTCACCTCTTCAGAACGA | [51] | |
| IL-6_R | CTGTTCTGGAGGTACTCTAGG | ||
| TNF-α_F | AGCCCATGTTGTAGCAAACC | [52] | |
| TNF-α_R | TGAGGTACAGGCCCTCTGAT | ||
| IL-8 F | ACACAGAGCTGCAGAAATCAGG | [53] | |
| IL-8 R | GGCACAAACTTTCAGAGACAG | ||
| TGF-β F | CCGGGTTATGCTGGTTGTACAG | [54] | |
| TGF-β R | AAGGACCTCGGCTGGAAGTGG |
| Feature Tested | BGKMJ1-36 | BGVLJ1-21 | |
|---|---|---|---|
| Shape | Cocci in short and long chains | Longer rods, single and short chains | |
| Catalase test | – | – | |
| Gram staining | + | + | |
| Growth at 15 °C in GM17 broth | – | – | |
| Growth at 37 °C in GM17 broth | + | + | |
| Growth at 45 °C in GM17 broth | + | + | |
| Growth in GM17 broth with 2% NaCl | – | – | |
| Hydrolysis of arginine | – | – | |
| Citrate utilization | – | – | |
| Black zone on bile esculin agar | – | – | |
| Production of CO2 from glucose | – | – | |
| Acetoin production (VP) | + | – | |
| Diacetyl production | – | – | |
| Curd forming after | 4 h and 45 min | 5 h | |
| Exopolysaccharides production | + | – | |
| Aggregation ability | – | – | |
| Hydrolysis of | αs1 casein | ± | + |
| β casein | + | + | |
| κ casein | ± | + | |
| Antimicrobial activity | + a | + a | |
| Antibiotic | BGKMJ1-36 | BGVLJ1-21 |
|---|---|---|
| Ampicillin | ≤ 1 (2) | ≤ 1 (2) |
| Vancomycin | ≤ 0.5 (4) | ≤ 0.5 (2) |
| Gentamicin | ≤ 16 (32) | ≤ 8 (16) |
| Kanamycin | n.r. | ≤ 4 (16) |
| Streptomycin | ≤ 32 (64) | ≤ 8 (16) |
| Erythromycin | ≤ 1 (2) | ≤ 0.5 (1) |
| Clindamycin | ≤ 0.5 (2) | ≤ 0.5 (4) |
| Tetracycline | ≤ 2 (4) | ≤ 0.5 (4) |
| Chloramphenicol | ≤ 1 (4) | ≤ 1 (4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popović, N.; Brdarić, E.; Đokić, J.; Dinić, M.; Veljović, K.; Golić, N.; Terzić-Vidojević, A. Yogurt Produced by Novel Natural Starter Cultures Improves Gut Epithelial Barrier In Vitro. Microorganisms 2020, 8, 1586. https://doi.org/10.3390/microorganisms8101586
Popović N, Brdarić E, Đokić J, Dinić M, Veljović K, Golić N, Terzić-Vidojević A. Yogurt Produced by Novel Natural Starter Cultures Improves Gut Epithelial Barrier In Vitro. Microorganisms. 2020; 8(10):1586. https://doi.org/10.3390/microorganisms8101586
Chicago/Turabian StylePopović, Nikola, Emilija Brdarić, Jelena Đokić, Miroslav Dinić, Katarina Veljović, Nataša Golić, and Amarela Terzić-Vidojević. 2020. "Yogurt Produced by Novel Natural Starter Cultures Improves Gut Epithelial Barrier In Vitro" Microorganisms 8, no. 10: 1586. https://doi.org/10.3390/microorganisms8101586