Noisy Galvanic Vestibular Stimulation (Stochastic Resonance) Changes Electroencephalography Activities and Postural Control in Patients with Bilateral Vestibular Hypofunction

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
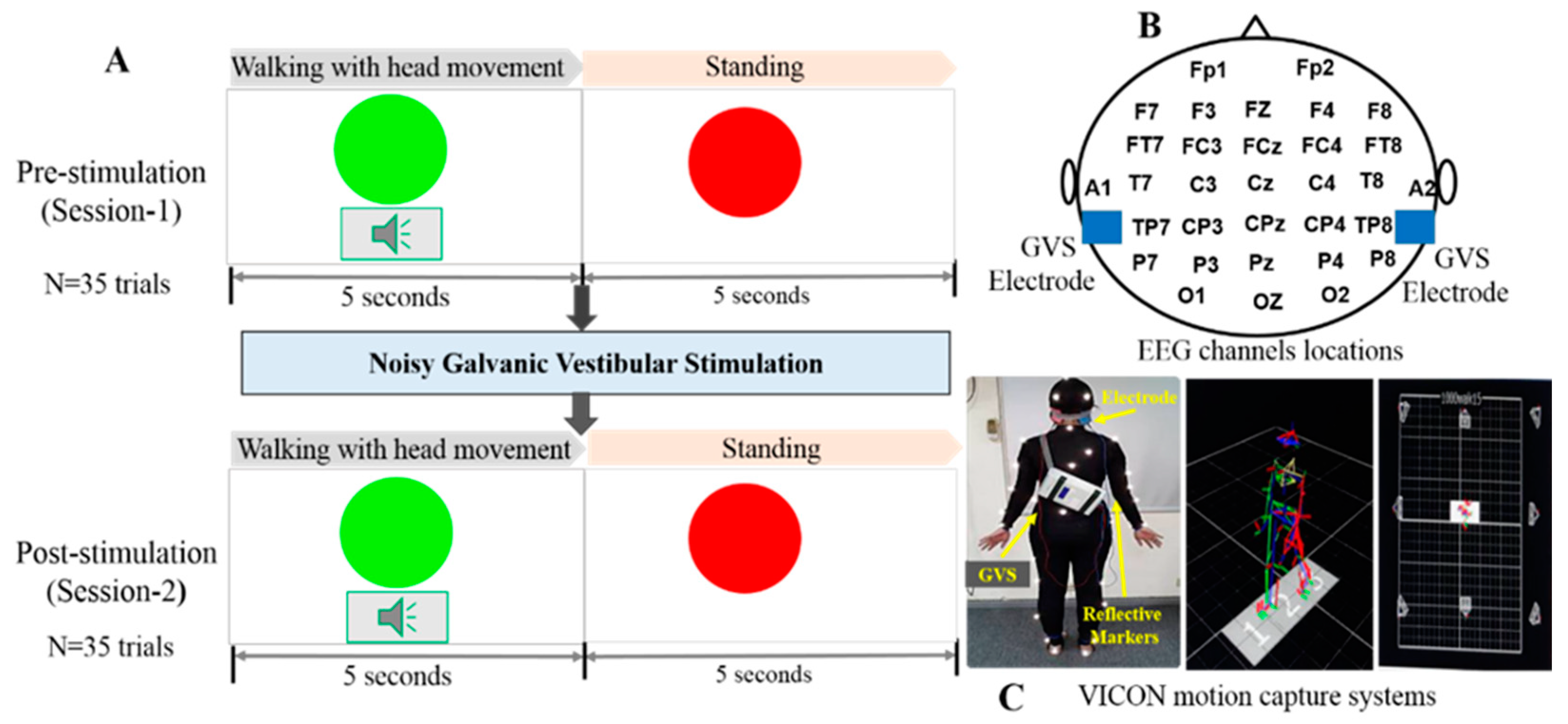
2.2. Experimental Design
2.3. Noisy Galvanic Vestibular Stimulation Process
2.4. EEG Acquisition and Analysis
2.5. Statistical Analysis
3. Results
3.1. Demographic Data
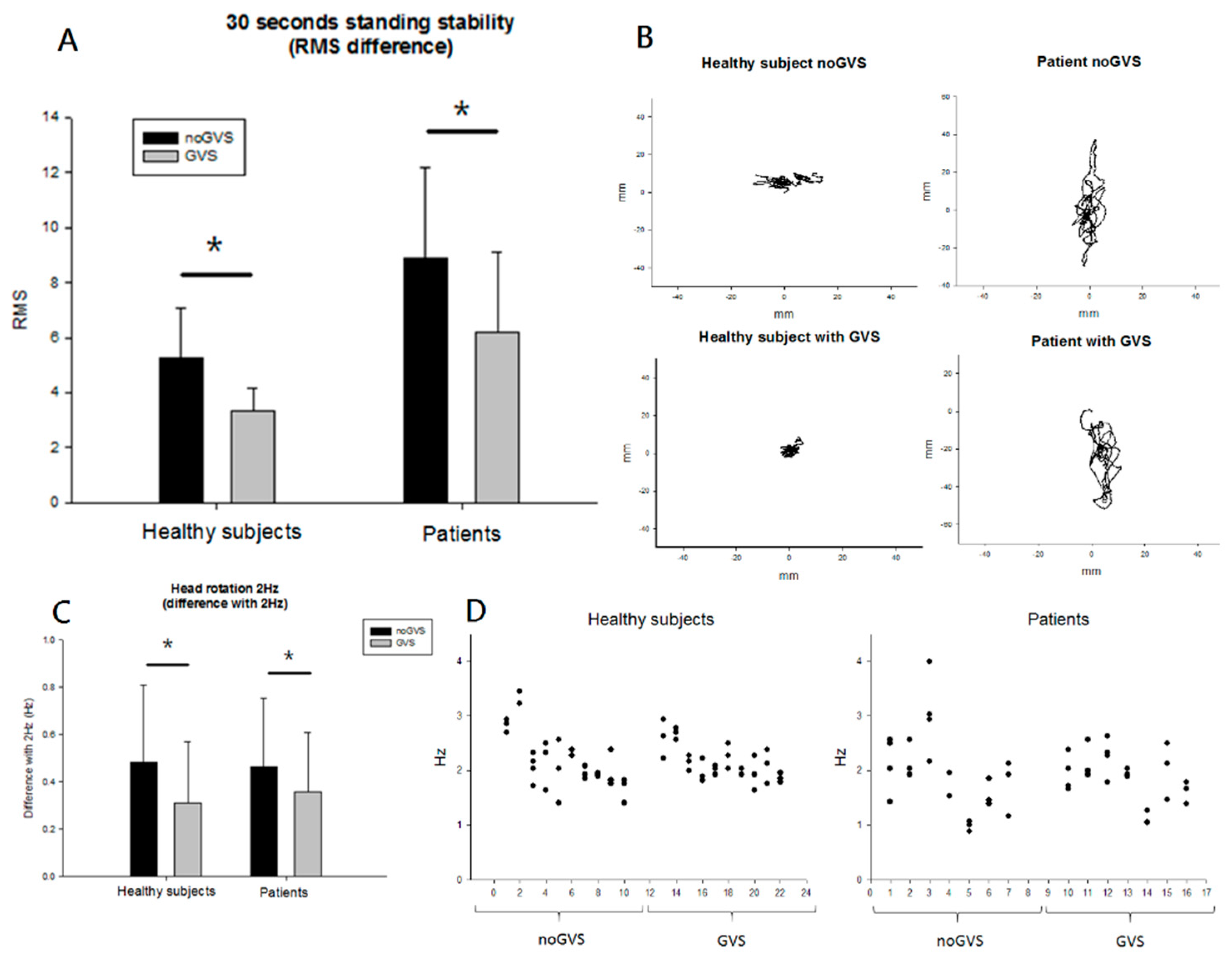
3.2. Behavioral Results
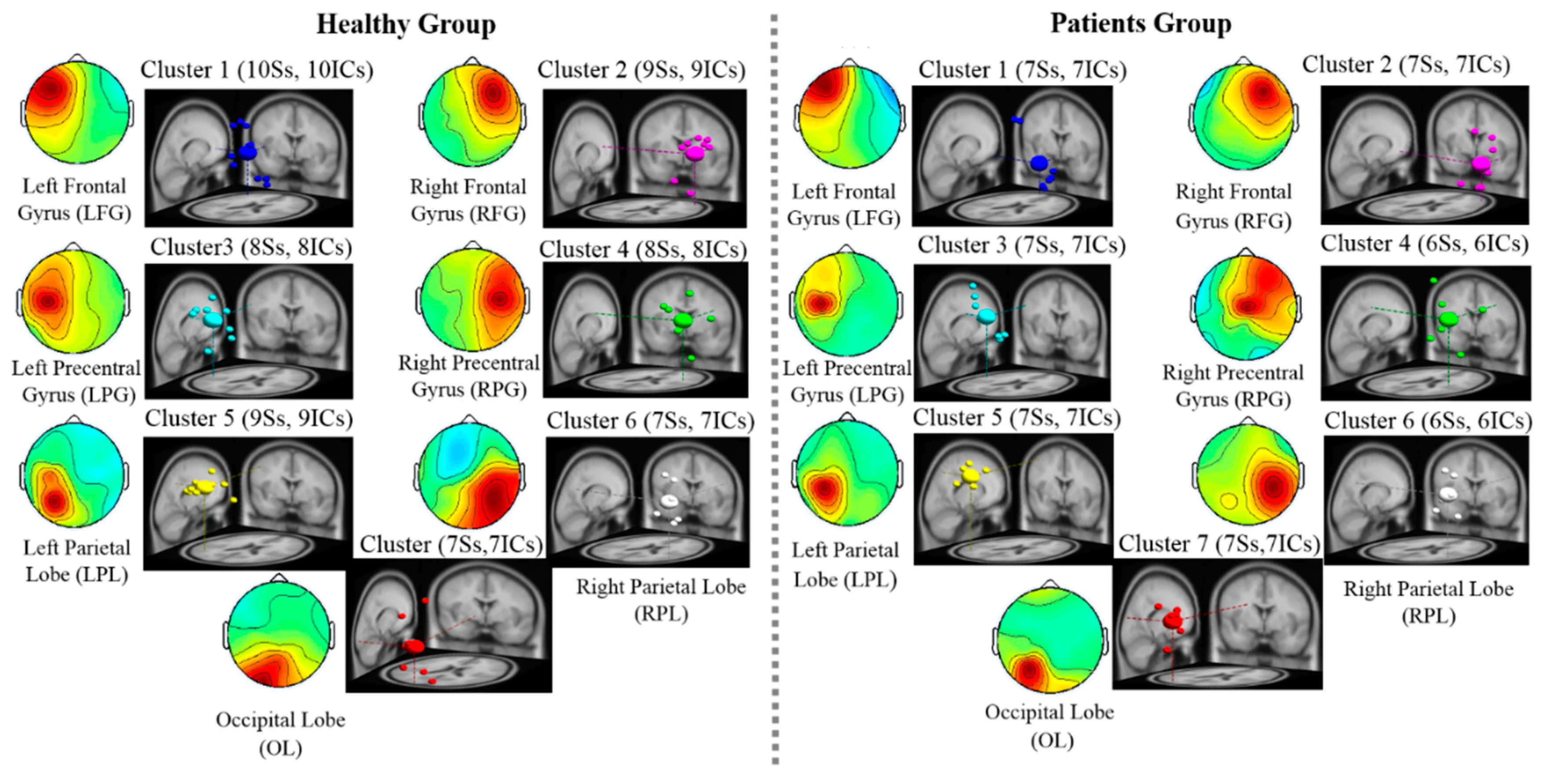
3.3. EEG Results: EEG Scalp Map and Dipole Source Locations
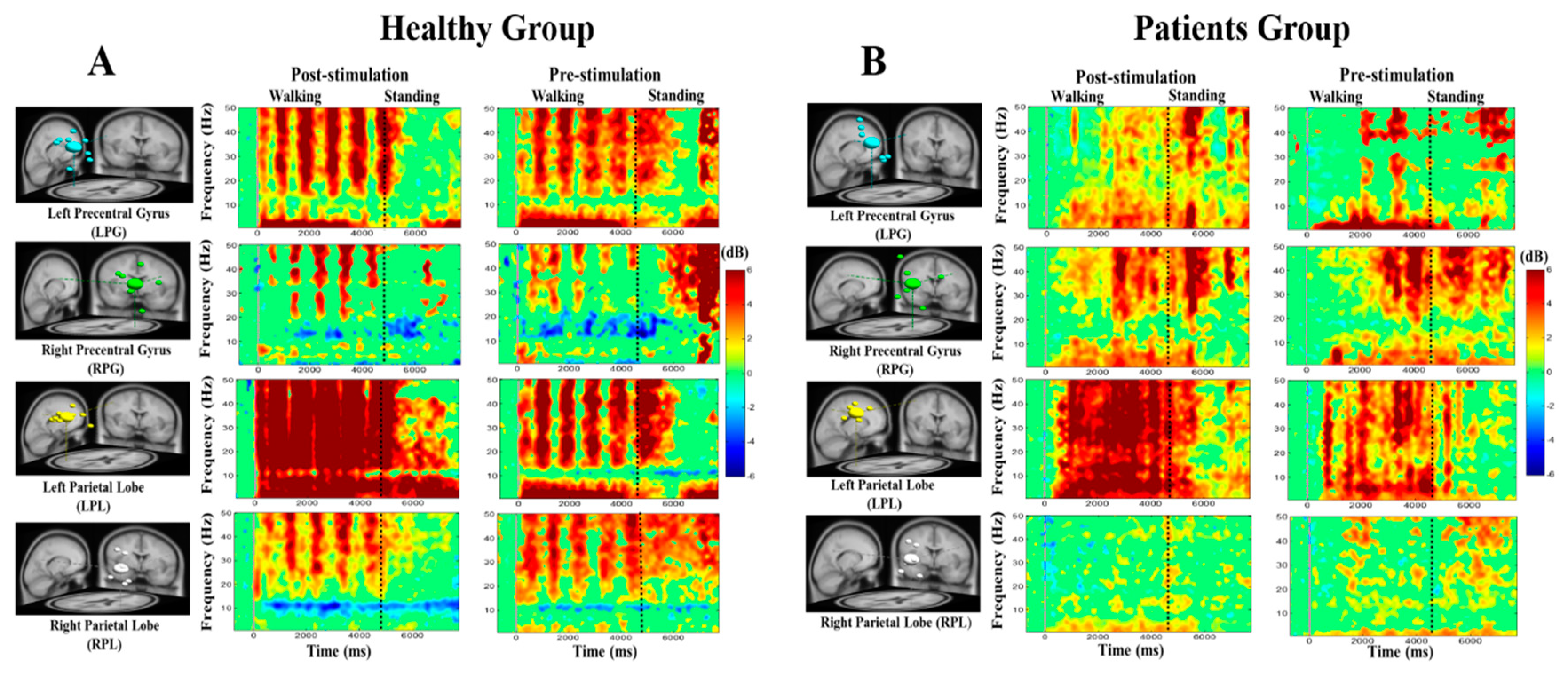
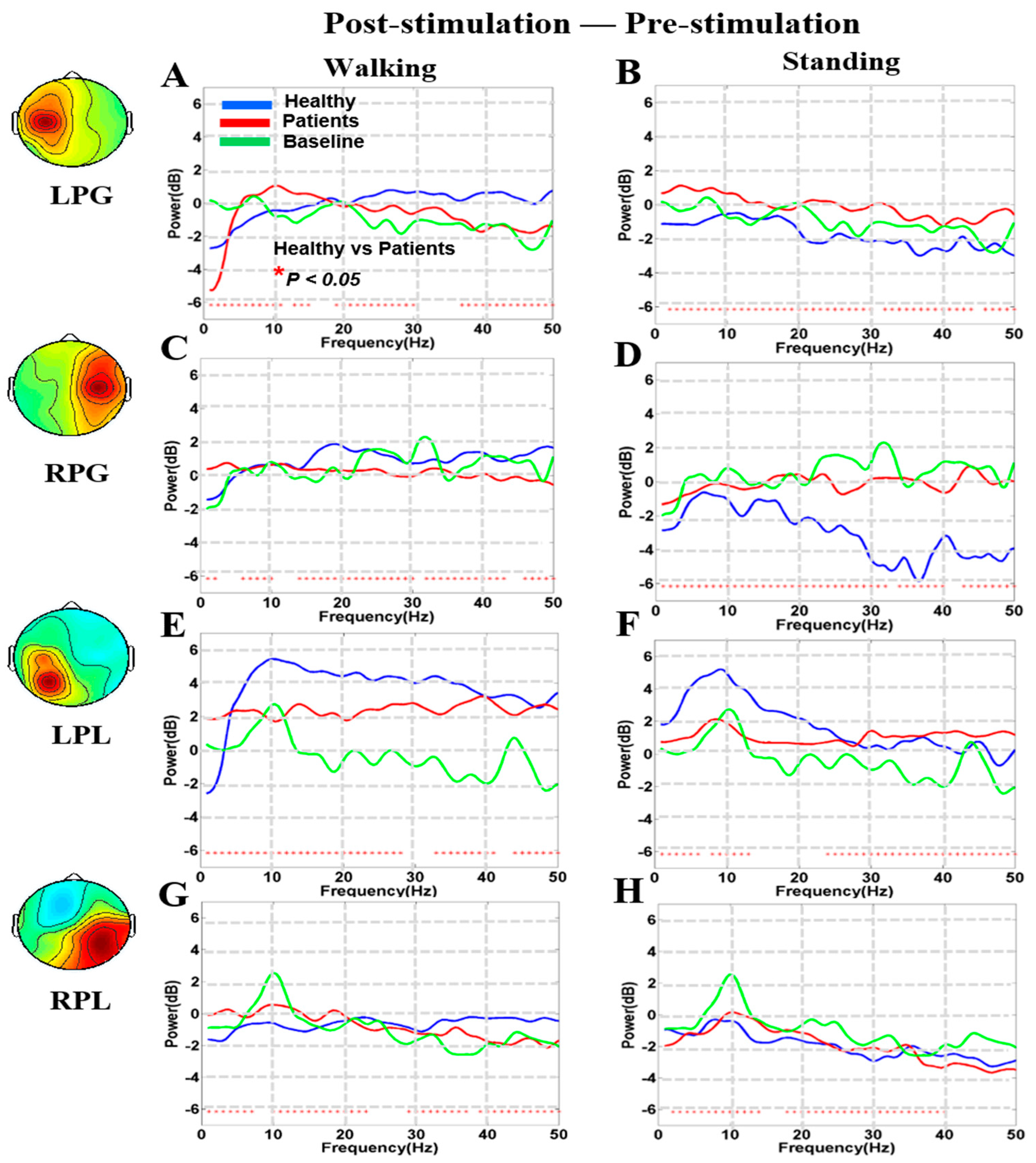
3.4. Noisy GVS Increases EEG Activities in Patients with BVH and Healthy Subjects
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GVS | galvanic vestibular stimulation |
| EEG | electroencephalography |
| SR | stochastic resonance |
| RMS | root mean square |
| COP | center of pressure |
| ICA | independent component analysis |
| BBS | blind source separation |
| BVH | bilateral vestibular hypofunction |
| ERSP | event-related spectral perturbation |
| LFG | left frontal gyrus |
| RFG | right frontal gyrus |
| LPG | left precentral gyrus |
| RPG | right precentral gyrus |
| LPL | left parietal lobe |
| RP | right parietal lobe |
| OL | occipital lobe |
References
- Wuehr, M.; Decker, J.; Schniepp, R. Noisy galvanic vestibular stimulation: An emerging treatment option for bilateral vestibulopathy. J. Neurol. 2017, 264, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, S.; Yamamoto, Y.; Togo, F.; Kinoshita, M.; Yoshifuji, Y.; Fujimoto, C.; Yamasoba, T. Noisy vestibular stimulation improves body balance in bilateral vestibulopathy. Neurology 2014, 82, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Wuehr, M.; Nusser, E.; Decker, J.; Krafczyk, S.; Straube, A.; Brandt, T.; Jahn, K.; Schniepp, R. Noisy vestibular stimulation improves dynamic walking stability in bilateral vestibulopathy. Neurology 2016, 86, 2196–2202. [Google Scholar] [CrossRef] [PubMed]
- Gittis, A.H.; du Lac, S. Intrinsic and synaptic plasticity in the vestibular system. Curr. Opin. Neurobiol. 2006, 16, 385–390. [Google Scholar] [CrossRef]
- Lopez, C.; Blanke, O.; Mast, F. The human vestibular cortex revealed by coordinate-based activation likelihood estimation meta-analysis. Neuroscience 2012, 212, 159–179. [Google Scholar] [CrossRef]
- Zu Eulenburg, P.; Caspers, S.; Roski, C.; Eickhoff, S.B. Meta-analytical definition and functional connectivity of the human vestibular cortex. Neuroimage 2012, 60, 162–169. [Google Scholar] [CrossRef]
- Goldberg, J.; Smith, C.E.; Fernandez, C. Relation between discharge regularity and responses to externally applied galvanic currents in vestibular nerve afferents of the squirrel monkey. J. Neurophysiol. 1984, 51, 1236–1256. [Google Scholar] [CrossRef]
- Minor, L.B.; Goldberg, J.M. Vestibular-nerve inputs to the vestibulo-ocular reflex: A functional-ablation study in the squirrel monkey. J. Neurosci. 1991, 11, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Struzik, Z.R.; Soma, R.; Ohashi, K.; Kwak, S. Noisy vestibular stimulation improves autonomic and motor responsiveness in central neurodegenerative disorders. Ann. Neurol. 2005, 58, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Soma, R.; Kwak, S.; Yamamoto, Y. Improvement of motor functions by noisy vestibular stimulation in central neurodegenerative disorders. J. Neurol. 2008, 255, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Soma, R.; Kwak, S.; Yamamoto, Y. Functional stochastic resonance in human baroreflex induced by 1/f-type noisy galvanic vestibular stimulation. In: Fluctuations and Noise in Biological, Biophysical, and Biomedical Systems. Int. Soc. Opt. Photonics 2003, 69–76. [Google Scholar]
- Fujimoto, C.; Yamamoto, Y.; Kamogashira, T.; Kinoshita, M.; Egami, N.; Uemura, Y.; Togo, F.; Yamasoba, T.; Iwasaki, S. Noisy galvanic vestibular stimulation induces a sustained improvement in body balance in elderly adults. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, C.; Egami, N.; Kawahara, T.; Uemura, Y.; Yamamoto, Y.; Yamasoba, T.; Iwasaki, S. Noisy galvanic vestibular stimulation sustainably improves posture in bilateral vestibulopathy. Front. Neurol. 2018, 9, 900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, S.; Karino, S.; Kamogashira, T.; Togo, F.; Fujimoto, C.; Yamamoto, Y.; Yamasoba, T. Effect of noisy galvanic vestibular stimulation on ocular vestibular-evoked myogenic potentials to bone-conducted vibration. Front. Neurol. 2017, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeber, M.; Scherer, R.; Wagner, J.; Solis-Escalante, T.; Müller-Putz, G.R. EEG beta suppression and low gamma modulation are different elements of human upright walking. Front. Hum. Neurosci. 2014, 8, 485. [Google Scholar] [CrossRef] [Green Version]
- Bruijn, S.M.; Van Dieën, J.H.; Daffertshofer, A. Beta activity in the premotor cortex is increased during stabilized as compared to normal walking. Front. Hum. Neurosci. 2015, 9, 593. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Durnford, S.J.; Crowley, J.S.; Rupert, A.H. Visual vestibular interaction in the dynamic visual acuity test during voluntary head rotation. Aviat. Space Environ. Med. 1997, 68, 111–117. [Google Scholar]
- Kim, D.J.; Yogendrakumar, V.; Joyce Chiang, E.T.; Wang, Z.J.; McKeown, M.J. Noisy galvanic vestibular stimulation modulates the amplitude of EEG synchrony patterns. PLoS ONE 2013, 8, e69055. [Google Scholar] [CrossRef] [Green Version]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Makeig, S.; Debener, S.; Onton, J.; Delorme, A. Mining event-related brain dynamics. Trends Cogn. Sci. 2004, 8, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.P.; Makeig, S.; Westerfield, M.; Townsend, J.; Courchesne, E.; Sejnowski, T.J. Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects. Clin. Neurophysiol. 2000, 111, 1745–1758. [Google Scholar] [CrossRef]
- Makeig, S.; Inlow, M. Lapses in alertness: Coherence of fluctuations in performance and EEG spectrum. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 23–35. [Google Scholar] [CrossRef]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ko, L.W.; Shih, Y.C.; Chikara, R.K.; Chuang, Y.T.; Chang, E.C. Neural mechanisms of inhibitory response in a battlefield scenario: A simultaneous fMRI-EEG study. Front. Hum. Neurosci. 2016, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Chikara, R.K.; Chang, E.C.; Lu, Y.C.; Lin, D.S.; Lin, C.T.; Ko, L.W. Monetary reward and punishment to response inhibition modulate activation and synchronization within the inhibitory brain network. Front. Hum. Neurosci. 2018, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Guldin, W.; Grüsser, O. Is there a vestibular cortex? Trends Neurosci. 1998, 21, 254–259. [Google Scholar] [CrossRef]
- Collins, J.; Chow, C.C.; Imhoff, T.T. Stochastic resonance without tuning. Nature 1995, 376, 236–238. [Google Scholar] [CrossRef]
- Collins, J.J.; Imhoff, T.T.; Grigg, P. Noise-enhanced information transmission in rat SA1 cutaneous mechanoreceptors via aperiodic stochastic resonance. J. Neurophysiol. 1996, 76, 642–645. [Google Scholar] [CrossRef]
- Fallon, J.B.; Carr, R.W.; Morgan, D.L. Stochastic resonance in muscle receptors. J. Neurophysiol. 2004, 91, 2429–2436. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F.G.; Fu, Q.J.; Morse, R. Human hearing enhanced by noise. Brain Res. 2000, 869, 251–255. [Google Scholar] [CrossRef]
- Priplata, A.A.; Niemi, J.B.; Harry, J.D.; Lipsitz, L.A.; Collins, J.J. Vibrating insoles and balance control in elderly people. Lancet 2003, 362, 1123–1124. [Google Scholar] [CrossRef]
- Reeves, N.P.; Cholewicki, J.; Lee, A.S.; Mysliwiec, L.W. The effects of stochastic resonance stimulation on spine proprioception and postural control in chronic low back pain patients. Spine 2009, 34, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Rosengren, S.M.; Colebatch, J.G. Stochastic galvanic vestibular stimulation produces a small reduction in sway in Parkinson’s disease. J. Vestib. Res. 2009, 19, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Guyot, J.P.; Gay, A.; Izabel, K.M.; Pelizzone, M. Ethical, anatomical and physiological issues in developing vestibular implants for human use. J. Vestib. Res. 2012, 22, 3–9. [Google Scholar] [CrossRef]
- Jasper, H.; Penfield, W. Electrocorticograms in man: Effect of voluntary movement upon the electrical activity of the precentral gyrus. Arch. Psychiatr. Nervenkrankh. 1949, 183, 163–174. [Google Scholar] [CrossRef]
- Crone, N.E.; Miglioretti, D.L.; Gordon, B.; Sieracki, J.M.; Wilson, M.T.; Uematsu, S.; Lesser, R.P. Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. I. Alpha and beta event-related desynchronization. Brain 1998, 121, 2271–2299. [Google Scholar] [CrossRef]
- Miller, K.J.; Leuthardt, E.C.; Schalk, G.; Rao, R.P.; Anderson, N.R.; Moran, D.W.; Miller, J.W.; Ojemann, J.G. Spectral changes in cortical surface potentials during motor movement. J. Neurosci. 2007, 27, 2424–2432. [Google Scholar] [CrossRef]
- Severens, M.; Nienhuis, B.; Desain, P.; Duysens, J. Feasibility of measuring event related desynchronization with electroencephalography during walking. In Proceedings of the 2012 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, San Diego, CA, USA, 28 August–1 September 2012; pp. 2764–2767. [Google Scholar]
- De Zeeuw, C.I.; Hansel, C.; Bian, F.; Koekkoek, S.K.; Van Alphen, A.M.; Linden, D.J.; Oberdick, J. Expression of a protein kinase C inhibitor in Purkinje cells blocks cerebellar LTD and adaptation of the vestibulo-ocular reflex. Neuron 1998, 20, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Grillner, S.; Hongo, T. Vestibulospinal effects on motoneurones and interneurones in the lumbosacral cord. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 1972; Volume 37, pp. 243–262. [Google Scholar]
- Grassi, S.; Pettorossi, V.E. Synaptic plasticity in the medial vestibular nuclei: Role of glutamate receptors and retrograde messengers in rat brainstem slices. Prog. Neurobiol. 2001, 64, 527–553. [Google Scholar] [CrossRef]
- Helmchen, C.; Rother, M.; Spliethoff, P.; Sprenger, A. Increased brain responsivity to galvanic vestibular stimulation in bilateral vestibular failure. Neuroimage Clin. 2019, 24, 101942. [Google Scholar] [CrossRef]
- Galvan, G.R.; Clark, T.; Mulavara, A.; Oman, C. Exhibition of stochastic resonance in vestibular tilt motion perception. Brain Stimul. 2018, 11, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Priesol, A.J.; Valko, Y.; Merfeld, D.M.; Lewis, R.F. Motion perception in patients with idiopathic bilateral vestibular hypofunction. Otolaryngol Head Neck Surg. 2014, 150, 1040–1042. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.; Ferguson, H.J.; Worley, A. Galvanic vestibular stimulation modulates the electrophysiological response during face processing. Vis. Neurosci. 2012, 29, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, M.; Moser, M.; Boegle, R.; Conrad, J.; zu Eulenburg, P.; Dieterich, M. The cortical spatiotemporal correlate of otolith stimulation: Vestibular evoked potentials by body translations. NeuroImage 2017, 155, 50–59. [Google Scholar] [CrossRef]
- Lobel, E.; Kleine, J.F.; LEROY-WILLIG, A.; VAN DE MOORTELE, P.F.; Bihan, D.L.; GRÜSSER, O.J.; Berthoz, A. Cortical areas activated by bilateral galvanic vestibular stimulation. Ann. N. Y. Acad. Sci. 1999, 871, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Shin, Y.; Yun, K. Beta-frequency EEG activity increased during transcranial direct current stimulation. Neuroreport 2014, 25, 1433–1436. [Google Scholar] [CrossRef]
- Ray, W.J.; Cole, H.W. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef]
- Keeser, D.; Padberg, F.; Reisinger, E.; Pogarell, O.; Kirsch, V.; Palm, U.; Karch, S.; Möller, H.J.; Nitsche, M.; Mulert, C. Prefrontal direct current stimulation modulates resting EEG and event-related potentials in healthy subjects: A standardized low resolution tomography (sLORETA) study. Neuroimage 2011, 55, 644–657. [Google Scholar] [CrossRef]
- Stam, C.; Montez, T.; Jones, B.; Rombouts, S.; Van Der Made, Y.; Pijnenburg, Y.; Scheltens, P. Disturbed fluctuations of resting state EEG synchronization in Alzheimer’s disease. Clin. Neurophysiol. 2005, 116, 708–715. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Diagnosis | Onset Time | Clinical Presentations | Training | |
|---|---|---|---|---|---|
| P1 | F | idiopathic BVH | 2.5 m | Unsteady gait, dizziness | NA |
| P2 | F | idiopathic BVH | 1.5 m | Unsteady gait, dizziness | NA |
| P3 | F | idiopathic BVH | 2 m | oscillopsia | 1 m |
| P4 | F | idiopathic BVH | 2 m | Hearing loss, vertigo, unsteady gait | 1 m |
| P5 | F | idiopathic BVH | 1 m | Unsteady gait, dizziness | 1 m |
| P6 | F | idiopathic BVH | 2.5 m | Dizziness, hearing loss, unsteady gait | 0.5 m |
| P7 | F | idiopathic BVH | 1.5 m | Vertigo, unsteady gait | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, L.-W.; Chikara, R.K.; Chen, P.-Y.; Jheng, Y.-C.; Wang, C.-C.; Yang, Y.-C.; Li, L.P.-H.; Liao, K.-K.; Chou, L.-W.; Kao, C.-L. Noisy Galvanic Vestibular Stimulation (Stochastic Resonance) Changes Electroencephalography Activities and Postural Control in Patients with Bilateral Vestibular Hypofunction. Brain Sci. 2020, 10, 740. https://doi.org/10.3390/brainsci10100740
Ko L-W, Chikara RK, Chen P-Y, Jheng Y-C, Wang C-C, Yang Y-C, Li LP-H, Liao K-K, Chou L-W, Kao C-L. Noisy Galvanic Vestibular Stimulation (Stochastic Resonance) Changes Electroencephalography Activities and Postural Control in Patients with Bilateral Vestibular Hypofunction. Brain Sciences. 2020; 10(10):740. https://doi.org/10.3390/brainsci10100740
Chicago/Turabian StyleKo, Li-Wei, Rupesh Kumar Chikara, Po-Yin Chen, Ying-Chun Jheng, Chien-Chih Wang, Yi-Chiang Yang, Lieber Po-Hung Li, Kwong-Kum Liao, Li-Wei Chou, and Chung-Lan Kao. 2020. "Noisy Galvanic Vestibular Stimulation (Stochastic Resonance) Changes Electroencephalography Activities and Postural Control in Patients with Bilateral Vestibular Hypofunction" Brain Sciences 10, no. 10: 740. https://doi.org/10.3390/brainsci10100740