Staphylococcus aureus and Methicillin Resistant S. aureus in Nepalese Primates: Resistance to Antimicrobials, Virulence, and Genetic Lineages


 ,
,  ,
,
Abstract
:
1. Introduction
2. Results
2.1. CC for MSSA Typing and CC and SCCmec Typing for MRSA Ancestral Lineage
2.2. Antibiotic Resistance Genes
2.3. Accessory and Virulence Factors
3. Discussion
4. Materials and Methods
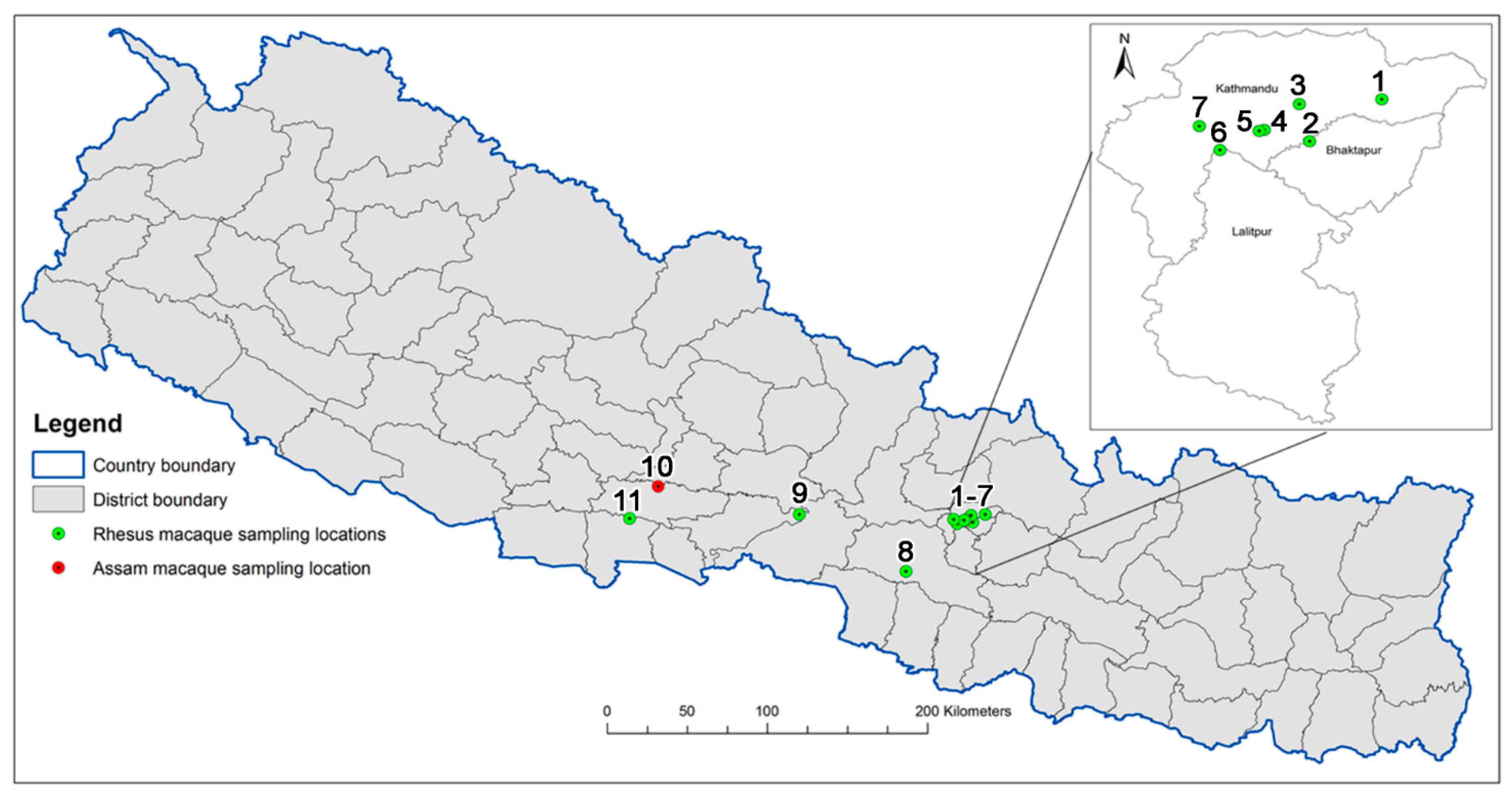
4.1. Primate Sampling
4.2. Ethical Statement
4.3. Identification of S. aureus from Primates
4.4. Antibiotic Susceptibility Testing
4.5. DNA Microarray Analysis
4.6. Multilocus Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus epidemiology in wildlife: A systematic review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soge, O.O.; No, D.; Michael, K.; Dakoff, J.; Lane, J.; Vogel, K.; Smedley, J.; Roberts, M.C. Transmission of MDR MRSA between primates personnel and environment at a United States primate center. J. Antimicrob. Chemother. 2016, 71, 2798–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.C.; Feßler, A.T.; Monecke, S.; Ehricht, R.; No, D.; Schwarz, S. Molecular analysis of two different MRSA clones ST188 and ST2368 from primates (Macaca spp.) in a United States primate center. Front. Microbiol. Antimicrob. Resist. Chemother. 2018, 9, 2199. [Google Scholar] [CrossRef]
- Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Muller, E.; Gawlik, D.; Paudel, S.; Acharya, M.; Bhattarai, S.; Pokharel, S.; et al. MRSA strains among Nepalese Rhesus macaques (Macaca mulatta) and their environment. Front. Microbiol. Antimicrob. Resist. Chemother. 2019, 10, 2505. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C.; Joshi, P.R.; Greninger, A.L.; Melendez, D.; Paudel, S.; Acharya, M.; Bimali, N.K.; Koju, N.P.; No, D.; Chalise, M.; et al. The human clone ST22 SCCmec IV methicillin-resistant Staphylococcus aureus isolated from swine herds and wild primates in Nepal: Is man the common source? FEMS Microbiol. Ecol. 2018, 1, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senok, A.; Somily, A.; Raji, A.; Gawlik, D.; Al-Shahrani, F.; Baqi, S.; Boswihi, S.; Skakni, L.; Udo, E.E.; Weber, S.; et al. Diversity of methicillin-resistant Staphylococcus aureus CC22-MRSA-IV from Saudi Arabia and the Gulf region. Int. J. Infect. Dis. 2016, 51, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokhrel, R.H.; Aung, M.S.; Thapa, B.; Chaudhary, R.; Mishra, S.K.; Kawaguchiya, M.; Urushibara, N.; Kobayashi, N. Detection of ST772 Panton-Valentine leukocidin-positive methicillin-resistant Staphylococcus aureus (Bengal Bay clone) and ST22 S. aureus isolates with a genetic variant of elastin binding protein in Nepal. New Microbes New Infect. 2016, 11, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.R.; Acharya, M.; Aryal, R.; Thapa, K.; Kakshapati, T.; Seng, R.; Singh, A.; Sitthisak, S. Emergence of staphylococcal cassette chromosome mec type I with high-level mupirocin resistance among methicillin-resistant Staphylococcus aureus. Asian Pac. J. Trop. Biomed. 2017, 7, 193–197. [Google Scholar] [CrossRef]
- Schaumburg, F.; Mugisha, L.; Kappeler, P.; Fichtel, C.; Kock, R.; Kondgen, S.; Becker, K.; Leendertz, F.H.; Peters, G. Evaluation of non-invasive biological samples to monitor Staphylococcus aureus colonization in great apes and lemurs. PLoS ONE 2013, 8, e78046. [Google Scholar] [CrossRef]
- Monecke, S.; Baier, V.; Coombs, G.W.; Slickers, P.; Ziegler, A.; Ehricht, R. Genome sequencing and molecular characterisation of Staphylococcus aureus ST772-MRSA-V, “Bengal Bay Clone”. BMC Res. Notes 2013, 6, 548. [Google Scholar] [CrossRef] [Green Version]
- Steinig, E.J.; Duchene, S.; Robinson, D.A.; Monecke, S.; Yokoyama, M.; Slickers, P.; Andersson, P.; Williamson, D.; Kearns, A.; Goering, R.V.; et al. Evolution and Global Transmission of a Multidrug-Resistant, Community-Associated Methicillin-Resistant Staphylococcus aureus Lineage from the Indian Subcontinent. mBiotechnology 2019, 10, e01105-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakara, S.; Khedkar, S.; Loganathan, R.M.; Chandana, S.; Malali, G.; Arakere, G.; Seshasayee, A.S.N. Draft genome sequence of Staphylococcus aureus 118 (ST772), a major disease clone from India. J. Bacteriol. 2012, 194, 3727–3728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Luedicke, C.; Slickers, P.; Ehricht, R. Molecular epidemiology of Staphylococcus aureus in asymptomatic carriers. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Labor Report. Labor Migration in Nepal. Available online: https://www.ilo.org/kathmandu/areasofwork/labour-migration/lang--en/index.htm (accessed on 1 September 2020).
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef]
- Azis, N.M.; Pung, H.P.; Bachman, A.R.A.; Nordin, S.A.; Sarchio, S.N.E.; Suhaili, Z.; Desa, M.N.M. A persistant antimicrobial resistance pattern and limited methicillin-resistance-associated genotype in a short-term Staphylococcus aureus carriage isolated from a student population. J. Infect. Publ. Health 2017, 10, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Penades, M.; Viana, D.; Garcia-Quiros, A.; Munoz-Siverstre, A.; Moreno-Grua, E.; Perez-Fuentes, S.; Pascual, J.; Corpa, J.M.; Selva, L. Differences in virulence between the two more prevalent Staphylococcus aureus clonal complexes in rabbitries (CC121 and CC96) using an experimental model of mammary gland infection. Vet. Res. 2020, 51, 11. [Google Scholar] [CrossRef] [Green Version]
- Harrison, E.M.; Paterson, G.K.; Holden, M.T.G.; Larsen, J.; Stegger, M.; Larsen, A.R.; Petersen, A.; Skov, R.L.; Christensen, J.M.; Zeuthen, A.B.; et al. Whole genome sequencing identifies zoonotic transmission of MRSA isolates with the novel mecA homologue mecC. EMBO Mol. Med. 2013, 5, 509–515. [Google Scholar] [CrossRef]
- Monecke, S.; Gavier-Widen, D.; Mattsson, R.; Rangstrup-Christensen, L.; Lazaris, A.; Coleman, D.C.; Shore, A.C.; Ehricht, R. Detection of mecC-positive Staphylococcus aureus (CC130-MRSA-XI) in diseased european hedgehogs (Erinaceus europaeus) in Sweden. PLoS ONE 2013, 8, e66166. [Google Scholar] [CrossRef] [Green Version]
- Loffler, B.; Hussain, M.; Grundmeier, M.; Bruck, M.; Holzinger, D.; Varga, G.; Roth, J.; Kah, B.C.; Proctor, R.A.; Peters, G. Staphylococcus aureus panton-valentine leukocidin is a very potent cytotoxic factor for human neutrophils. PLoS Pathog. 2010, 6, e1000715. [Google Scholar] [CrossRef]
- Kyes, R.C.; Jones-Engel, L.; Chalise, M.K.; Engel, G.; Heidrich, J.; Grant, R.; Bajimaya, S.S.; McDonough, J.; Smith, D.G.; Ferguson, B. Genetic characterization of rhesus macaques (Macaca mulatta) in Nepal. Am. J. Primatol. 2006, 68, 445–455. [Google Scholar] [CrossRef]
- Khanal, L.; Chalise, M.K.; He, K.; Acharya, B.K.; Kawamoto, Y.; Jiang, X. Mitochondrial DNA analyses and ecological niche modeling reveal post-LGM expansion of the Assam macaque (Macaca assamensis) in the foothills of Nepal Himalaya. Am. J. Primatol. 2018, 80, e22748. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, K.P.; Khanal, L.; Chalise, M.K. Status and effects of food provisioning on ecology of Assamese monkey (Macaca assamensis) in Ramdi area of Palpa, Nepal. J. Inst. Sci. Technol. 2018, 22, 183–190. [Google Scholar] [CrossRef]
- Evans, T.S.; Barry, P.A.; Gilardi, K.V.; Goldstein, T.; Deere, J.D.; Fike, J.; Yee, J.-A.; Ssebide, B.J.; Karmacharya, D.; Cranfield, M.R.; et al. Optimization of a novel non-invasive oral sampling technique for zoonotic pathogen surveillance in nonhuman primates. PLoS Negl. Trop. Dis. 2015, 9, e0003813. [Google Scholar] [CrossRef]
- Roberts, M.C.; Soge, O.O.; Horst, J.A.; Ly, K.A.; Milgrom, P. Methicillin-resistant Staphylococcus aureus from dental school clinic surfaces and students. Am. J. Infect. Control. 2011, 39, 628–632. [Google Scholar] [CrossRef]
- ISO 20776-1 (2006). Clinical Laboratory Testing and In Vitro Diagnostic Test. Systems—Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test. Devices—Part. 1: Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases. 2006. Available online: http://www.eucast.org/ast_of_bacteria/mic_determination/ (accessed on 1 September 2020).
- Kinneyvey, P.M.; Shore, A.C.; Brennan, G.I.; Sullivan, D.J.; Ehricht, R.; Monecke, S.; Slickers, P.; Coleman, D.C. Emergence of sequence type 779 methicillin-resistant Staphylococcus aureus harboring a novel pseudo staphylococcal cassette chromosome mec (SCCmec)-SCC-SCCCRISPR composite element in Irish hospitals. Antimicrob. Agents Chemother. 2012, 57, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Jatzwauk, L.; Muller, E.; Nitschke, H.; Pfohl, K.; Slickers, P.; Reissig, A.; Ruppelt-Lorz, A.; Ehricht, R. Diversity of SCCmec elements in Staphylococcus aureus as observed in South-Eastern Germany. PLoS ONE 2016, 11, e0162654. [Google Scholar] [CrossRef]
- Monecke, S.; Slickers, P.; Gawilik, D.; Muller, E.; Reissig, A.; Ruppelt-Lorz, A.; Akpaka, P.E.; Bandt, D.; Bes, M.; Boswihi, S.S.; et al. Molecular typing of ST239-MRSA-III from diverse geographic locations and the evolution its SCCmec III element during its intercontinental spread. Front. Microbiol. 2018, 9, 1436. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Jatzwauk, L.; Weber, S.; Slickers, P.; Ehricht, R. DNA microarray-based genotyping of methicillin-resistant Staphylococcus aureus strains from eastern saxony. Clin. Microbiol. Infect. 2009, 14, 534–545. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Clonal Complex | Strain | Isolate ID | Number | Sampling Location | Host Species | gapA, katA, coA, nuc1, spa, sbi, eno, clfA+B, fnbA | Staphyloxanthin Operon | agr Group | Capsule Type |
|---|---|---|---|---|---|---|---|---|---|
| CC06 | CC6-MRSA-IV | B | 1 | Nilbarahi | Rhesus | POS | not tested | I | 8 |
| CC22 | CC22-MRSA-IV (PVL+/tst+) | 93, 94, 05, 97, C | 5 | 2 Gokarna, 1 each Chitwan, Ramdi and Nilbarahi | 1 Assam, 4 Rhesus | POS | not tested | I | 5 |
| CC772 | CC772-MRSA-V (PVL+), “Bengal Bay” | 91 | 1 | Hetauda | Rhesus | POS | not tested | II | 5 |
| CC15 | CC15-MSSA | 83 | 1 | Rupandehi | Rhesus | POS | not tested | II | 8 |
| CC96 | CC96-MSSA | 89 | 1 | Ramdi | Assam | POS | not tested | III | 8 |
| CC2990 | CC2990-MSSA | H | 1 | Guheswari | Rhesus | POS | POS | II | 8 |
| UNKNOWN 01 | ST(4-13-1-105-11-5-x)-MSSA | 86, 87 | 2 | Ramdi | Assam | POS | POS | I | 5 |
| UNKNOWN 02 | ST(1-421-1-1-12-238-11)-MSSA | 52, 53 | 2 | Gokarna | Rhesus | POS | POS | I | 5 |
| UNKNOWN 03 | ST(1-3-1-15-28-x-1)-MSSA | 31, 32, 71, D | 4 | 2 Nilbarahi, 1 each Chitwan and Gokarna | Rhesus | POS | POS | I | 8 |
| UNKNOWN 04 | MSSA | 01 | 1 | Pashupati | Rhesus | POS | POS | I | 8 |
| UNKNOWN 05 | ST(1-1-1-1-28-4-11)-MSSA | 73, F | 2 | 1 Chitwan, 1 Gokarna | Rhesus | POS | POS | I | 8 |
| UNKNOWN 06 | ST(1-421-1-x-x-1-11)-MSSA | 54 | 1 | Gorkarna | Rhesus | POS | POS | I | 8 |
| UNKNOWN 07 | ST(12-x-1-66-11-x-x)-MSSA | 29, 30, 51 | 3 | 2 Swayambhu, 1 Gokarna | Rhesus | POS | POS | II | 8 |
| UNKNOWN 08 | ST(1-38-1-1-x-238-x)-MSSA | 81, 88 | 2 | 1 Ramdi, 1 Rupandehi | 1 Assam, 1 Rhesus | POS | POS | II | 8 |
| UNKNOWN 09 | ST(3-3-2-66-28-x-x)-MSSA | 10 | 1 | Pashupati | Rhesus | POS | POS | II | 8 |
| UNKNOWN 10 | ST(3-1-1-66-28-1-x)-MSSA | 82, 90 | 2 | 1 Ramdi, 1 Rupandehi | 1 Assam, 1 Rhesus | POS | POS | III | 5 |
| UNKNOWN 11 | MSSA | 50 | 1 | Bajrayogini | Rhesus | POS | POS | III | 8 |
| UNKNOWN 12 | ST(4-421-1-105-1-5-x)-MSSA | 61 | 1 | Hetauda | Rhesus | POS | POS | IV | 5 |
| UNKNOWN 13 | ST(1-3-1-x-1-1-11)-MSSA | 62, 72 | 2 | 1 Chitwan, 1 Hetauda | Rhesus | POS | POS | IV | 5 |
| UNKNOWN 14 | ST(3-38-1-15-1-x-40)-MSSA | G | 1 | Guheswari | Rhesus | POS | POS | IV | 5 |
| UNKNOWN 15 | ST(1-421-1-1-12-1-11)-MSSA | 11, E | 2 | 1 Gokarna, 1 Thapathali | Rhesus | POS | POS | IV | 5 |
| UNKNOWN 16 | ST(3-3-1-66-4-x-x)-MSSA | 49, I | 2 | 1 Bajrayogini, 1 Guheswari | Rhesus | POS | POS | IV | 8 |
| UNKNOWN 17 | MSSA | 13 | 1 | Thapathali | Rhesus | POS | POS | IV | 8 |
| Clonal Complex | Strain | Number | PEN | OXA | FOX | GEN | TOB | CIP | LEV | MOX | ERY | CLI | LIN | TEI | VAN | TET | FOS | NIF | FUS | MUP | RIF | TSU |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC06 | CC6-MRSA-IV | 1 | R | R | R | S | S | S | S | S | R | R a | S | S | S | S | S | S | S | S | S | S |
| CC22 | CC22-MRSA-IV (PVL+/tst+) | 5 | R | R | R | R | R | R | R | R | VAR b | VAR ab | S | S | S | S | S | S | S | S | S | VAR c |
| CC772 | CC772-MRSA-V (PVL+), “Bengal Bay” | 1 | R | R | R | R | R | R | R | R | I | S | S | S | S | S | S | I | S | S | S | R |
| CC15 | CC15-MSSA | 1 | R | S | S | S | S | R | R | R | R | R a | S | S | S | S | S | S | S | S | S | S |
| CC96 | CC96-MSSA | 1 | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | S | S | S | S | S |
| CC2990 | CC2990-MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 01 | ST(4-13-1-105-11-5-x)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S |
| UNKNOWN 02 | ST(1-421-1-1-12-238-11)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 03 | ST(1-3-1-15-28-x-1)-MSSA | 4 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 04 | MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 05 | ST(1-1-1-1-28-4-11)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 06 | ST(1-421-1-x-x-1-11)-MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 07 | ST(12-x-1-66-11-x-x)-MSSA | 3 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 08 | ST(1-38-1-1-x-238-x)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S |
| UNKNOWN 09 | ST(3-3-1-66-28-x-x)-MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 10 | ST(3-1-1-66-28-1-x)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 11 | MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 12 | ST(4-421-1-105-1-5-x)-MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 13 | ST(1-3-1-x-1-1-11)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S |
| UNKNOWN 14 | ST(3-38-1-15-1-x-40)-MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 15 | ST(1-421-1-1-12-1-11)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 16 | ST(3-3-1-66-4-1-x)-MSSA | 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| UNKNOWN 17 | MSSA | 1 | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Clonal Complex | Strain | Number | mecA | Delta mecR | ccrA-2 | ccrB-2 | ccrAA | ccrC | blaZ/I/R | erm(C) | lnu(A) | msr(A) | mph(C) | aacA-aphD | aphA3+sat | dfrA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC06 | CC6-MRSA-IV | 1 | POS | POS | POS | POS | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG |
| CC22 | CC22-MRSA-IV (PVL+/tst+) | 5 | POS | POS | POS | POS | NEG | NEG | POS | VAR b | NEG | NEG | NEG | POS | NEG | POS |
| CC772 | CC772-MRSA-V(PVL+), “Bengal Bay “ | 1 | POS | NEG | NEG | NEG | POS | POS | POS | NEG | NEG | POS | POS | POS | POS | NEG |
| CC15 | CC15-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | POS | NEG | NEG | NEG | NEG | NEG | NEG |
| CC96 | CC96-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG |
| CC2990 | CC2990-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 01 | ST(4-13-1-105-11-5-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 02 | ST(1-421-1-1-12-238-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 03 | ST(1-3-1-15-28-x-1)-MSSA | 4 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 04 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 05 | ST(1-1-1-1-28-4-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 06 | ST(1-421-1-x-x-1-11)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 07 | ST(12-x-1-66-11-x-x)-MSSA | 3 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 08 | ST(1-38-1-1-x-238-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 09 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 10 | ST(3-3-1-66-28-x-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 11 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 12 | ST(4-421-1-105-1-5-x)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 13 | ST(1-3-1-x-1-1-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 14 | ST(3-38-1-15-1-x-40)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 15 | ST(1-421-1-1-12-1-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 16 | ST(3-3-1-66-4-1-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 17 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| Clonal Complex | Strain | Number | tst | sea | sec+sel | egc Genes | ORF CM14 | PVL | lukD/E b | sak | chp | scn | edinB+etD2 | cna | sasG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC06 | CC6-MRSA-IV | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | POS | NEG | POS | NEG | POS | POS |
| CC22 | CC22-MRSA-IV (PVL+/tst+) | 5 | POS | NEG | POS | POS | NEG | POS | NEG | POS | POS | POS | NEG | POS | POS |
| CC772 | CC772-MRSA-V (PVL+), “Bengal Bay “ | 1 | NEG | POS | POS | POS | POS | POS | NEG | NEG | NEG | POS | NEG | POS | POS |
| CC15 | CC15-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | POS | POS | NEG | NEG | POS |
| CC96 | CC96-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | POS |
| CC2990 | CC2990-MSSA | 1 | NEG | NEG | POS | NEG | NEG | NEG | (POS) b | POS | POS | POS | POS | POS | POS |
| UNKNOWN 01 | ST(4-13-1-105-11-5-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | POS | NEG | POS |
| UNKNOWN 02 | ST(1-421-1-1-12-238-11)-MSSA | 2 | NEG | NEG | NEG | POS | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | POS |
| UNKNOWN 03 | ST(1-3-1-15-28-x-1)-MSSA | 4 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | NEG |
| UNKNOWN 04 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | (POS) b | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 05 | ST(1-1-1-1-28-4-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | POS | POS | POS |
| UNKNOWN 06 | ST(1-421-1-x-x-1-11)-MSSA | 1 | NEG | NEG | NEG | POS | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | NEG |
| UNKNOWN 07 | ST(12-x-1-66-11-x-x)-MSSA | 3 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 08 | ST(1-38-1-1-x-238-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | NEG |
| UNKNOWN 09 | ST(3-3-1-66-28-x-x)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | (POS) b | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 10 | ST(3-1-1-66-28-1-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | POS |
| UNKNOWN 11 | MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | POS |
| UNKNOWN 12 | ST(4-421-1-105-1-5-x)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | POS |
| UNKNOWN 13 | ST(1-3-1-x-1-1-11)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | POS | NEG | VAR c |
| UNKNOWN 14 | ST(3-38-1-15-1-x-40)-MSSA | 1 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | POS | POS | POS |
| UNKNOWN 15 | ST(1-421-1-1-12-1-11)-MSSA | 2 | NEG | NEG | NEG | POS | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | POS |
| UNKNOWN 16 | ST(3-3-1-66-4-1-x)-MSSA | 2 | NEG | NEG | NEG | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG |
| UNKNOWN 17 | MSSA | 1 | NEG | NEG | NEG | POS | NEG | NEG | POS | NEG | NEG | NEG | NEG | POS | NEG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Müller, E.; Gawlik, D.; Diezel, C.; Braun, S.D.; Paudel, S.; Acharya, M.; et al. Staphylococcus aureus and Methicillin Resistant S. aureus in Nepalese Primates: Resistance to Antimicrobials, Virulence, and Genetic Lineages. Antibiotics 2020, 9, 689. https://doi.org/10.3390/antibiotics9100689
Roberts MC, Joshi PR, Monecke S, Ehricht R, Müller E, Gawlik D, Diezel C, Braun SD, Paudel S, Acharya M, et al. Staphylococcus aureus and Methicillin Resistant S. aureus in Nepalese Primates: Resistance to Antimicrobials, Virulence, and Genetic Lineages. Antibiotics. 2020; 9(10):689. https://doi.org/10.3390/antibiotics9100689
Chicago/Turabian StyleRoberts, Marilyn C., Prabhu Raj Joshi, Stefan Monecke, Ralf Ehricht, Elke Müller, Darius Gawlik, Celia Diezel, Sascha D. Braun, Saroj Paudel, Mahesh Acharya, and et al. 2020. "Staphylococcus aureus and Methicillin Resistant S. aureus in Nepalese Primates: Resistance to Antimicrobials, Virulence, and Genetic Lineages" Antibiotics 9, no. 10: 689. https://doi.org/10.3390/antibiotics9100689