Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants


Integrative Medicine Cluster, Advanced Medical and Dental Institute, Universiti Sains Malaysia, Bertam, Kepala Batas 13200, Malaysia
*
Authors to whom correspondence should be addressed.
Plants 2020, 9(10), 1345; https://doi.org/10.3390/plants9101345
Submission received: 8 September 2020
/
Revised: 7 October 2020
/
Accepted: 7 October 2020
/
Published: 12 October 2020
(This article belongs to the Special Issue Structural and Functional Analysis of Extracts in Plants)
Abstract
:The zebrafish (Danio rerio) is used as an embryonic and larval model to perform in vitro experiments and developmental toxicity studies. Zebrafish may be used to determine the toxicity of samples in early screening assays, often in a high-throughput manner. The zebrafish embryotoxicity model is at the leading edge of toxicology research due to the short time required for analyses, transparency of embryos, short life cycle, high fertility, and genetic data similarity. Zebrafish toxicity studies range from assessing the toxicity of bioactive compounds or crude extracts from plants to determining the optimal process. Most of the studied extracts were polar, such as ethanol, methanol, and aqueous solutions, which were used to detect the toxicity and bioactivity. This review examines the latest research using zebrafish as a study model and highlights its power as a tool for detecting toxicity of medicinal plants and its effectiveness at enhancing the understanding of new drug generation. The goal of this review was to develop a link to ethnopharmacological zebrafish studies that can be used by other researchers to conduct future research.
1. Introduction
Herbal plants have pharmacological and therapeutic characteristics due to the natural chemical compounds they contain [1]. Hence, they are widely utilized every day for culinary purposes and nutritional supplements to promote health [2]. This product can be toxic, so its toxicity must be measured to ensure adequate safety for human health [3]. Although many people view most medicinal plants as safe, poisoning can occur in some cases. Consumers can also be exposed to potential health risks caused by specific components or contaminants of botanical products; thus, their risk needs to be evaluated [4,5]. Plant materials and their extracts contain various toxic substances synthesized by plants as a defense against disease, insects, and other organisms [6]. Botanical toxicity studies are complicated due to expense, time, use of animals, and the complex mix of components [4,7]. The most commonly used extraction solvents, from polar to non-polar, are water, ethanol, methanol, acetone, ethyl acetate, chloroform, dichloromethane and hexane, and numerous researchers have studied the effectiveness of these solvents [8]. Various extracts may display distinctive cytotoxicity traits and a wide variety of pharmacological consequences at different concentrations [9]. Additionally, numerous factors affect compounds, such as the type and volume of extraction solvents used and varying storage environments [8]. Bioactive compounds in medicinal plants can have toxic impacts on cardiac glycosides, phorbol esters, alkaloids, cyanogenic glycosides, and lectins [10]. The statistical evaluation carried out by the National Pharmaceutical Control Bureau and Health ministry’s Malaysian Adverse Drug Reaction Advisory Committee in 2013 confirmed that 11,437 cases of harmful drug reaction had been reported, and 0.2% were due to herbal remedies [1]. Thus, the evaluation of the toxicity of potential drug compounds has been enhanced in recent years [11]. Allopathy drugs and complementary and alternative medicines require a toxicology assay to identify any harmful effects that are not well-known until signs and symptoms appear after high consumption [12].
However, the main objective of toxicity research is to predict human toxicity via fast and accurate testing of many substances [13] based on model systems [14], and different routes of drug delivery systems such as using nanoparticles in the gastrointestinal tract [15] or chemical fertilizers and water retention as an example [16]. Previously, classical toxicity screening (including rodents, dogs, and rabbits) involved the compilation of data from a given laboratory for one substance at a time [17]; however, these tests are frequently expensive, time-consuming, and tedious [18]. This classic method focused on studying chemical compounds on phenotypic cell or animal results [19]. Thus, an attractive alternative approach involves applying the 3R concept of human-animal studies (i.e., reduction, replacement, and refinement), but it is not consistent with the use of rodent animal models [20]. Replacement: Zebrafish assays can be used to substitute such animal-toxicity studies using larval zebrafish, and it can be shown that larval zebrafish represent a critical system model; Reduction: zebrafish larvae can be used as a first-level model for toxicity to classify toxic drug candidates so that more stable drugs can be evaluated in mammalian models; Refinement: The model of embryonic and larval zebrafish provides a refined design to research organisms since embryos are partially fertilized and translucent in their early life [21]. It is recommended that the number of animals is limited, the test methods minimize pain and suffering of experimental animals, and approved substituted animal tests are used as much as possible [22].
The zebrafish (Danio rerio) is a suitable model for screening drugs for potential use to treat human diseases [23] based on phylogenetic analysis of fish and human genomes, which shows similar morphology and physiology of the nervous, cardiovascular, and digestive systems [18]. The zebrafish genome has been sequenced in full [24]. Zebrafish are a fast model for the study of genetic and de novo mutations [25]. The genes can inactivate in vivo, simulate human phenotypes, and obtain information on human diseases with genetic background through genomic editing approaches, such as CRISPR/Cas9 or artificial site-specific nucleases, zinc-finger nucleases, and transcription activator-like nucleases [25]. Zebrafish also provide a significant data file for vertebrate animals that enables researchers to anchor biochemical, genetic, and cellular hypotheses to high-performance observations at structural, functional, and behavioral levels [26]. The zebrafish embryotoxicity test, or fish embryotoxicity test (FET), is gaining popularity because it provides a total and well-defined developmental duration for a vertebrate embryo and allows the study of its early life stages [27]. As experimental models, both adults and zebrafish embryos are used [22]. For example, various compound tests (e.g., measurement of drug cardiotoxicity) have been conducted to assess drug effects in zebrafish efficiently [11]. Additionally, for the last two decades, zebrafish have been used to study angiogenesis, metastasis, anticancer drug screening, and an assessment of drug toxicity [28]. In summary, the cell structural and biochemical similarities between humans and zebrafish enable rapid forecasting of the possible impacts of chemical and other substances on human communities. Because zebrafish are becoming increasingly important as a test model, husbandry requirements to enhance the reproducibility and efficiency of this type of model in research environments are needed [29].
Toxicity studies generally begin in in vitro studies, with many different cell lines at different sample concentrations. The substance is then tested in several animal models, especially in mice and rats, before using on patients [30]. As mentioned in Table 1, there are several explanations why zebrafish as an animal model could be extremely useful for intermediate toxicity tests.
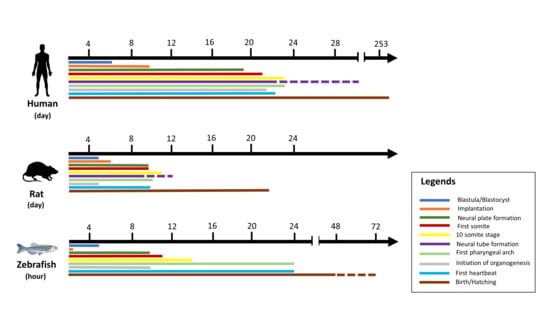
In vitro toxicity studies are inexpensive, fast, and easy, as shown in Table 1, but cultured cells are poorly associated with in vivo processes and therefore have limited translation benefit, while accurate data are obtained from laboratory rodent studies to extrapolate toxicants to humans [32]. Hence, the gold standard for predictive analysis of chemical risks to humans remains vertebrate toxicity studies; however, these studies are unsuitable for initial toxicity screening due to the reasons above and often involve significant amounts of a valuable test compound [26]. There has been a trend in the last 30 years towards the restricted use of higher animals in herbal toxicology studies in particular [22]. These animals have a series of restrictions. For example, rodents may be immune to the cardiotoxicity, especially when the endpoint is left ventricular contractile function. This may be due to rodents’ ability to substitute for myocyte failure by employing alternative mechanisms [25]. The zebrafish model is especially useful for the bioassay-guided identification of bioactive secondary metabolites [33]. The National Health Institute (NIH), USA, recently promoted a zebrafish model organism to study various genetically engineered diseases [34]. The Food and Drug Administration has recognized zebrafish tests for toxicity and safety evaluations for investigative newly developing drugs [26]. Large volume screening platforms exist in the form of a multi-wave plate format for testing chemical impacts on embryo development by assessing deformities, mortality, and structural characteristics across a range of concentrations [26]. Small molecules may be introduced directly into the water in multi-well racks, where fish take them up via diffusion. The researched drugs can also be injected into the yolk sac [35]. Thus, the zebrafish is a great model for studying the premature embryonic diet on toxicants [36]. Although researchers hope that both adults and embryos will prove useful for toxicology studies, embryos are ideal for toxicity examinations because of the transparency of the egg, which makes it easy to detect developmental phases and evaluate endpoints during the toxicity test [22,37]. This enables technical and economic benefits over rodent models [26,38] to reduce the number of substances and reduce the cost of animals in the development of drugs [39]. Up to 200–300 embryos can be developed per pair of adult zebrafish, while a typical pair of rodents only produce 5–10 descendants per matching occurrence [24]. Five days after post fertilization (dpf), the heart, liver, brain, pancreas, and other organs are created [40]. The three-lobe liver of zebrafish reflects humans’ biological function, including the absorption and production of lipids, vitamins, proteins, and carbohydrates [25]. One of the significant benefits of using zebrafish is that compounds can be easily supplied by adding water, similar to chemicals in the cell culture medium, which generally requires an overall amount of only 100 μL during growth [41]. Their optical clarity makes it easy to produce and recognize phenotypic properties during mutagenesis screening, and determine toxicity endpoints during toxicological analysis [32]. In addition, zebrafish embryos have been incredibly helpful in studying heart development and the functional effects of toxicants [14]. Figure 1 illustrates the temporal distinction between humans, rats, and zebrafish in early developmental life [26,41]. During development, there are still several limitations in the chemical identification of zebrafish. The fish is ectothermic and lacks heart septa, limbs, synovial joints, cancellous bones, lungs, and other organs [42]. Age is one of the most important limitations. Although chemical tests can be undertaken in adult fish, a high-performance scan using appropriate quantities of small molecules requires the animals to fit into a multi-well plate [19]. The analysis of gene expression throughout the larvae is feasible due to the optical transparency of the zebrafish tissues, which enables good penetration for light microscopy [32].
An alternative preliminary toxicity test is called the brine shrimp lethality test, which involves testing different toxicant concentrations. However, this technique does not reveal abnormalities or causes of death [43]. In contrast, using zebrafish as a screening model animal has some rational benefits, including large embryos, large numbers of embryos for testing, and simple visualization of organogenesis using fluorescence and transgenic strains [36]. Additionally, the ability to view biological processes provides the researcher with visual links to the organ system dynamics and, possibly, interorgan systems [44].
Today, many herbal products claim to provide pharmaceutical health benefits but do not provide any toxicological data [1]; thus, the safety of these drugs is questionable. This review aimed to compare the results of studies of the toxicity of medicinal plants and assess the toxicity tests used to analyze some of these commercial products from plants. This paper is the first review to assess the toxic and teratogenic effects of plant extracts on the zebrafish model to the best of our knowledge.
2. Zebrafish as a Suitable Alternative Animal Model for Toxicity Tests
One of the main goals of medicinal plant toxicology research is to identify many bioactive compounds with specific toxicity within a short period [13]. For many years, higher species have been used as models to measure medicinal products based on their toxicity [11]. For instance, recent studies of acute toxicity have mainly used mice as the animal model. Nevertheless, it is difficult to achieve a thorough and immediate toxicity check with this organism because of its strong state of breeding, high expenses, complicated procedures, and ethical restrictions [45]. Instead of using rodents, fish are the best candidate for this purpose. Fish traditionally have been used in toxicity testing of individual substances and effluents [46], and today zebrafish are commonly utilized to test for developmental toxicity, general toxicity and to carry out medication screening as a credible vertebrate model [11,26]. Hence, the first large-scale studies for bioactive molecules using zebrafish embryos were published, using the merits of zebrafish as phenotypic testing models to evaluate the effects of biomolecules and explore different bioactive compounds [47]. The zebrafish is widely used in numerous subfields of biology, and without a doubt, it is one of the leading species in various research areas, including developmental biology, ecotoxicology, and genetics [48]. Zebrafish embryos can also quickly consume tiny molecular compounds, thus providing a valuable model for drug testing and evaluating teratogenic effects [49,50] of exposure to toxic compounds [38]. Zebrafish embryos and larvae are outstanding models for testing the toxicity of substances, especially if those substances are present in low quantities [51]. Zebrafish larvae are very useful in imaging studies, and they likely will prove useful for non-imaging endpoints. Scientists are discovering new innovative paths for evaluating biochemical processes [19]. Within 5–6 days dpf, zebrafish development reflects the full developmental period of a vertebrate embryo before it becomes self-sustainable and, therefore, this substitute organism is not currently recognized as a genuine in vivo form by European law [52].
Teratology is the study of unusual growth, and a teratogen is any substance that triggers the production of a congenital anomaly or enhances the occurrence of a specific hereditary deficiency [53]. Screening for teratogenicity includes introducing zebrafish embryos to the required concentration of the compound of interest, and it has become a popular model for analyzing the teratogenic effects of medicines [35]. Teratological and embryo-toxic effects are easy to detect due to the transparency of zebrafish embryos during their growth outside of their parent [54]. Teratogenic effects include tail malformation, pericardial edema, malformation of the notochord, scoliosis, yolk edema, and growth delays [22]. Therefore, it is essential to evaluate the embryotoxic and teratogenic toxicity of medicinal plants on embryo growth. Embryo malformations may be caused by activation of the Caspase-3 enzyme, which is the leading cause of apoptosis [55], or by other factors such as reactive oxygen species-induced oxidative stress [6].
The effectiveness of the zebrafish model system was evident in the 1960s and 1970s, as numerous studies used zebrafish as bioassays of chemicals affecting normal functioning and reproductive success. During the 1980s and 1990s, many researchers verified that zebrafish are a convenient and reliable option for comprehensive toxicological screening and early life and lifetime exposure tests [56]. In 2000, the first chemical screening procedure was reported that allowed researchers to test a very slight concentration of compounds on live zebrafish in 96-well plates [19]. To date, there are ten types of tests used to study toxicity in zebrafish: (1) the zebrafish embryo toxicity test (FET); carcinogenicity; (2) developmental toxicity and teratogenicity assessments; (3) reproductive toxicity assessment; (4) behavioral toxicity assessment; (5) endocrine disorders; (6) acute toxicity; (7) neurotoxicity; (8) optical (ocular toxicity); (9) cardiotoxicity; and (10) vascular toxicity [31]. Among the available tests, FET is the most useful for assessing chemical and substance toxicity in zebrafish [48]. Numerous toxicity studies of specific chemicals have shown a significant correlation between results for zebrafish embryos and acute toxicity of fish [57]. The FET test aims to determine the acute toxicity of the embryonic phases and establish variables for zebrafish [52]. It is based on mortality and teratogenesis of the zebrafish embryos [46]. FET is a valuable option for replacing the use of adult animals to evaluate toxicity and enhance existing toxicity assays [48]. Advantages of this test include:
- A wide range of chemicals may be relevant;
- Embryos are short-lived, susceptible, cost-effective, and have low variability;
- The regulatory and scientific communities consider them to be standardized;
- The tolerance of various species or organisms can be compared [58].
The possibility of tracking many toxicity endpoints, including alteration of molecular processes and malformations, is one of FET’s main advantages with Danio rerio [59]. With the FET test, acute toxicity is assessed based on positive results, and embryo formation is examined every day for any unusual developmental phenotypes, especially morphological defects, including (i) coagulation of fertilized eggs; (ii) lack of somite formation; (iii) lack of detachment of the tail-bud from the yolk sac; and (iv) lack of a heartbeat [52]. Nagel (2002) outlined these toxicity indicators as measured using an inverse light microscope based on “yes” or “no” responses to the presence of the four factors [60]. On the other hand, Hermsen et al. [61] introduced a new model of embryotoxicity (ZET) using the general morphology score (GMS). A separate scoring list was established for teratogenic impact, which helped track slow progress, developmental delay, and teratogenicity. However, the ZET, along with GMS, operates as an essential and useful test method for screening the chemicals’ embryotoxic properties in the compound groups studied [27]. Furthermore, the MolDarT was created with zebrafish eggs/larvae to develop a molecular mechanism test method [61]. It is based in theory on the DarT (Danio rerio teratogenicity test), established by Nagel et al. [60], which reveals and tracks the developmental impact and deformities of freshly fertilized zebrafish eggs within 48 h [62].
Experiments to test toxicity include acute, sub-chronic, and chronic studies of compounds’ toxicity to specific organ pathways and hypothesis-driven research [31]. Zebrafish were found to be a useful tool for the comprehension not only of neurotoxicants’ structural and chemical effects but also for the assessment of behavioral dysfunction correlated with such toxicity [38]. Several behavioral endpoints are used in neurotoxicity studies to determine the therapeutic influences of medications and their neurotoxins on neuron development in zebrafish [63]. There is an incomparable system to identify endocrine activity lacking significant morphological abnormalities [13]. For the reasons described above, zebrafish toxicity screening provides many significant experimental advantages, including embryo and larvae visibility, high-efficiency short test time, low number of necessary compounds, ease of handling, and direct delivery compounds [54]. The zebrafish embryo develops quickly outside the mother’s body and enters maturity in a couple of months, among other benefits [38]. Additionally, more than 70% of the disease-associated genes are similar to those found in human diseases [50]. Furthermore, the physical, biochemical, genetic, and molecular makeup of zebrafish, including organs and tissues, has been demonstrated to be identical to their mammalian relatives. Metabolites, signaling mechanisms, and neurological and cognitive structures are similar to those in mammals [54].
A compound’s toxicity is measured using two indicators: the median effective concentration (EC50) and the median lethal concentration (LC50). The concentration-response curve is generated using 24 h of data to obtain the EC50 (teratogenic effects) and LC50 (embryotoxic effects). The ratio of LC50 to EC50 is the corresponding therapeutic index (TI). In pharmaceutical administration, a TI value < 1 is optimal [50,64]. The strength of the association between zebrafish embryo LC50 values and rodent LD50 values for 60 different compounds was demonstrated in one particular laboratory study by Ali and colleagues [42]. Test protocols for the use of zebrafish embryos (FET) in the Organization for Economic Cooperation and Development (OECD) test guideline 236 were tested and applied [65]. As determined by OECD guidelines, the value of LC50 is calculated based on lethality, coagulation, lack of somite formation, heartbeat failure, and a lack of detachment of the tail. In contrast, teratogenic effects are used to compute the EC50 values [22]. Parng et al. (2002) showed that the toxicity of the drug tested (ethanol) was similar for zebrafish and mammals with log LC50 values of 4.0 and 3.8 mg/mL, respectively [66]. Throughout the screening steps, zebrafish bioassays have been used to classify the active fractions based on endpoints developed and tested in other species, which led to identification of the actual effect of extracts on different life stages in zebrafish (Table 2).
Additionally, indicators such as delayed growth, restricted movement, irregular head-trunk angle, scoliosis/flexure, and yolk sac edema have been analyzed in developing zebrafish [73]. However, recent findings indicate that herbal products’ metabolism using zebrafish can represent actual outcomes of mammalian methods, while single in vitro methods cannot do so [39]. Table 3 lists some toxicity screening studies of medicinal plants that used zebrafish as the model organism.
3. The Most Relevant Behavioral Effects and Basic Methodology Based on Toxicological Assessment Observed in Zebrafish Larvae
Previously, zebrafish were used for toxicity evaluations of agrochemicals, but recently they have been used for toxicity assessments for therapeutic compounds [11]. Of note, the zebrafish response is also a sensitive predictor of irregular toxicity changes [34]. In particular, specific tasks have been created or changed to test the behavior of zebrafish compared to rodent models. Quite basic swimming steps and the ability to capture/consume can often be helpful [38]. The creation of behavioral tests that can employ repeated tests in a short window (i.e., within 24 h) may prove to be the perfect compromise, and some researchers have begun designing tasks that use multiple tests to measure their acquisition during a single session [38]. If one particular mode of action is to be detected, various experimental parameters can contribute to incoherent actions (hypo-or hyperactivity) responses of zebrafish when determining toxicity [122]. As a consequence, several behavioral evaluation methods have been developed to address different behavioral endpoints, including spontaneous tail coiling, photomotor response (PMR), locomotor response (LMR), and the alternating light/dark-induced locomotor response (LMR-L/D) [122], which we will not discuss further in this review.
Zebrafish are easy to control in toxicology screening tests using different platforms (exposure format) for detecting parameters. The type of exposure format is classified into ten categories, six of which are useful. Zebrafish body length at 5 dpf is about 4 mm, so a multiple-well plate can be used to incubate zebrafish larvae [17]. However, embryos cover a wide range of sizes based on the performing test, so 6-well to 96-well transparent plates can be used. Analytical experiments that use microplates, such as cell-based evaluation, can be conducted and applied to high-performance primary drug screening [123]. Furthermore, zebrafish larvae fit onto 96 or 386-well microplates [24]. Because the zebrafish embryo is tiny, can be handled easily [31], and requires only a small number of compounds per test, they can be screened in a 96-well microplate format [123]. However, OECD TG236 suggests using a 24-well plate (with a medium of 2 mL per well), as it overcomes the problem of high concentration developing in conjunction with evaporation [17]. Nevertheless, TG236 specifically checks for lethality and only considers developmental defects indirectly [17]. Table 4 lists several studies that analyzed embryotoxicity and apoptotic induction.
Apoptotic functions are identical in zebrafish and humans [66], and can be easily detected using fluorescent labeling methods [123]. The zebrafish model was used to discover the toxicity of 69 plants in this review, including 88 crude plant extracts, eight polyherbal formulates/commercial products, and two phytocompounds. In terms of exposure format, out of 58 studies described in Table 4, 25 used 24-well plates, and 10 used 96-well plates.
4. Using Zebrafish Embryos to Detect Developmental Toxicity
It seems that the number of zebrafish screening tests to evaluate the toxicity of compounds will keep increasing each year [19]. The use of fish embryos was proposed as an alternative screening method to assess fish’s acute toxicity [21,57]. In transparent species, the effects of compounds on different organs, such as the brain, heart, cartilage, liver, intestine, and kidney, were identified without arduous screening [66]. This specific trait also makes it feasible to quickly measure toxicity endpoints of multiple substances [6,124], highlighting the efficacy of toxicity models using zebrafish embryos [66]. For example, after 2005, regular sewage surveillance monitoring of fish embryotoxicity has been made compulsory, substituting conventional fish tests no longer approved for standard whole effluent assessment [62].
From the egg phase, zebrafish embryos can survive by ingesting yolk and can visibly be tested for malformation for a few days in a single well of a microplate [32]. When the embryo is not clumped, has a cardiac beat, has fully shaped body sections, and the tail is detached from the yolk, the standard endpoints of embryogenesis have been said to have occurred [28]. The main characteristics of embryogenesis and the genetic basis of growth in zebrafish have been widely researched [125]. Another significant indicator for assessing toxicity is the hatching rate, and zebrafish embryos start to hatch at about 48 h post-fertilization (hpf) under ordinary conditions [8]. Delayed growth of zebrafish embryos can result in a low hatchability and can thus be among the critical aspects of the sub-lethal effects of plant extracts [53].
The essential cell structure and biochemical similarities between humans and animals enable researchers to use the zebrafish model to forecast the possible impacts of chemicals and other human communities [10]. Furthermore, it is easier to study the biochemical mechanisms of significant zebrafish organs, whereas histological analysis required in the mouse model is more difficult [66]. Testing parameters commonly used for zebrafish include embryo survival rate, lethality, behavior, and organ deformity; researchers have found that zebrafish exhibit a strong dose reaction to toxicity, making it a useful animal model for toxicity screening [123]. Previously, we mentioned FET; most studies use OECD guidelines to measure toxicity of crude extract/fractions, and those guidelines for FET set levels as follows: dangerous (10 mg/L < LC50 < 100 mg/L), toxic (1 mg/L < LC50 < 10 mg/L), and carcinogenic (LC50 < 1 mg/L) [126].
Zebrafish developmental toxicity testing is a type of FET (FET can be used for any fish species), and it is primarily aimed at supplementing developmental toxicity screening in mammals [17]. When detecting developmental toxicity in zebrafish embryos, endpoint parameters are categorized in two ways compared to a standard (negative control). First, lethality is assessed based on coagulation, malformation of somites, absence of heartbeat, and lack of tail detachment. Second, teratogenicity is evaluated based on abnormal eye development, lack of spontaneous movement, unusual heart rate, lack of pigmentation, and edema. The standard shows normal development [60,127].
5. Defects in Zebrafish Organs Found in Developmental Toxicity Studies
In developmental toxicity studies, the mortality of zebrafish embryos occurs before 24 hpf or just after hatching, and cardiac deformities are sometimes detected, such as pericardial edema, abnormal heart form due to edema or aplasia, and irregular heartbeat [4]. The motility of the zebrafish embryo can also be affected by toxins, as exhibited by coagulation, heart failure, and non-development of the yolk embryo’s tail [112]. Other factors, such as reactive oxygen species-induced oxidative stress, are also assumed to be linked to abnormal development during embryogenesis [128]. A teratogenicity test is carried out to assess developmental toxicity (i.e., if the compound has teratogenic effects on the embryos). After treatment, the development stages of embryos are observed under an inverted microscope to look for malformations based on the numerical system designed from 5 to 0.5, where 1 indicates severe malformation, 4 indicates slight malformation and 5 is totally normal [50]. The teratogenic score is calculated as follows:
In such analyses, it is essential to specify the observational setup and endpoint evaluation (i.e., which particular behavioral change(s) will be analyzed). Below we discuss the defects to vital organs of zebrafish that can occur during the toxicity assay.
5.1. Heart (Cardiotoxicity)
The zebrafish is an excellent model organism for studying organogenesis and the cardiovascular system [86,129]; thus, it was developed quickly as a cardiovascular research model organism [35]. The first active organ that develops in zebrafish is the heart [73,130,131]. The heartbeat is a primary and relevant sub-lethal endpoint in the embryonic fish toxicology assay [73,130]. It is regularly assessed as an indicator of toxicity in zebrafish embryos [73] and estimates fish metabolic function as a biological factor [6,132]. In zebrafish, the normal embryonic heartbeat of 120–180 beats per minute is close to that of the human heartbeat. At the embryo pharyngula level, the heartbeat pulse can be tracked because the tail is visibly pigmented [102,118]. Ismail et al. reported that the most vulnerable stage for external stimulation (e.g., chemicals, toxicants, and physical pressure) is the early growth stage of embryos [2]. In toxic environments, pericardial edema can be caused by the general response. Pericardial edema in zebrafish embryos can be caused by many different toxicants [133], and changes in heartbeat rate may be a common reaction to toxicant exposure [130]. The toxicity mechanism of a compound is due to activation of Na+ and K+ inhibitors, and these Na+ channels likely trigger the heart rate. An elevated heart rate can overwhelm the myocardium and cause operational and biological damage to the heart of zebrafish [134]. For example, studies of the effects of medicinal plants such as Andrographis paniculata, Cinnamon zeylanicum, Curcuma xanthorrhiza, Eugenia polyantha, Orthosiphon stamineus [2], Tinospora cordifolia [118], Carthamus tinctorius [83], and Euodia retaecara [92] on zebrafish embryos reported a decreased heartbeat. In contrast, embryos treated with curcumin showed a heartbeat increase [1].
The heart is anteroventral in zebrafish and lies between the operculum and pectoral girdle in the thoracic cavity [35,135]. It is divided into the atrium, sinus venous, ventricle, and bulbous arteriosus, it starts beating at about 22 to 26 hpf [17,136], and produces a full set of ion channels and metabolic regulation [123]. In later stages, the normal function of the heart plays a leading role in the growth of the embryo. An abnormally formed cardiovascular system can result in the unusual overall growth of the animal, severe malformations, and a malfunctioning body [8]. Additionally, when the heart is swollen due to the active compounds in the pericardial sac, cardiac cells become irritated [43]. The following method is used to measure a normalized cardiac rate [120]:
Zebrafish embryos are also being used in cardiotoxicity (reduction of heartbeat) studies to model various human diseases [11]. The morbidity and mortality of people with cancer are associated with cardiotoxicity [35]. Most common drugs have cardiotoxic effects and affect the heart in zebrafish model systems [86]. A very accurate zebrafish cardiotoxicity assay assessing potential drug toxicity to the human cardiovascular system has been documented [11]. Thus, the effects of cardiotoxicity, such as minimal or no blood flow, may be explained by the fact that a large percentage of apoptotic heart cells lead to underdevelopment of the heart and pericardium, which in turn can cause an unusual heartbeat and a delay in body development (growth retardation) [137,138]. Gao et al. reported that the expression of two heart markers (amhc and vmhc) reflected the seriousness of heart failure in zebrafish [133].
At day 3 of the formation of zebrafish embryos, the primary and sprout vessels become active, and the blood vessel structure is quite like that in the human body [91]. A single endothelial cell layer surrounded by supporting cells forms the vessels that constitute the circulatory system. Endothelial precursor cells result from vasculogenesis and angiogenesis within the lateral mesoderm of mammal embryos. Vasculogenesis explains the de novo establishment of blood vessels by angioblast coalescence. In comparison, angiogenesis includes the creation of new vessels by earlier developed vessels and is typically described by the sprouting of an endothelial cell [139]. Kinna et al. reported the potential role of zebrafish crim1 in regulating vascular and somatic development [140].
5.2. Gills
Gills are essential to fish because they are the leading site for gas exchange [141,142,143], and they engage in osmoregulation [90], ionic control, acid-base balance, and excretion of nitrogen waste in fish [144]. Several studies on the histological organization and physiology of gills in many fishes have been carried out [145]. Enhanced green fluorescent protein (EFGP) is a successful marker for toxic chemicals in transgenic zebrafish. One of the benefits of this zebrafish transgenic model system is that spatio-temporal patterns of EGFP expression can be assessed during initial stages after exposure to a particular toxicant [146]. A study by Seok and colleagues found that the most effective and earliest expression of EGFP was seen in zebrafish gills, indicating that the gill is the most sensitive tissue and is impacted at relatively low metal concentrations [146]. The studies conducted by Rajini et al. [147] showed that visible damage such as inflammatory cell infiltration in gills, minimum aggregation in primary lamellas, secondary lamellae fusion, diffuse epithelial hyperplasia, and multifocal cell mucosal hyperplasia occurred in zebrafish exposed to sublethal concentrations of a combination of pesticides [147]. Generally, in zebrafish, gills exposed to toxicants from 48 to 72 h show fusion, and clubbing of distal lamellar regions [148]. However, for herbal toxicity, this organ has not fully been considered.
5.3. Tail
One of the endpoints of embryonic growth in zebrafish is when the tail is detached entirely from the yolk [28], which can be affected by toxicants. The tail emerges during the larval stages from the primordium in the ventral fin fold, which coincides with a gap in the melanophore line immediately before the back tip of the notochord [149]. Tail kink and tail bending are malformations caused by toxicity. However, tail malformations and spinal axis disabilities can also be affected by predation and a drastic decrease in the food supply [6].
5.4. Yolk Sac
Like human embryos, zebrafish embryos possess a protruding yolk sac, but in zebrafish, it serves as a nutritional reservoir for the embryo [36,43]. Yolk sac edema is a common pathology in toxicity tests of zebrafish [32,150], and it is a sign of reduced nutrient adsorption by the embryo [43]. It may be affected by overhydration of osmoregulation and toxin accumulation in the yolk sac [150]. Malformation in other organ systems that help allocate nutrients may be the trigger for this deficiency. This deficit could also contribute to nutritional absorption, undernourished embryos, and embryo mortality [43]. After maternal exposure, toxicants can be placed in the yolk sac as well [36]. As they are deposited in the yolk, the embryo becomes directly exposed to these toxicants, which allows for definitive exposure timing. Hence, a finite quantity of maternally stored embryo yolk is used in the zebrafish embryo for early feeding, and it provides an understanding of embryo nutrition processes and disruptions that occur due to toxicity exposure [36].
In zebrafish developmental toxicology research, there are numerous classes of yolk phenotypes. Yolk sac edema is quite like yolk edema. Yolk retention refers to decreased mobility/use or malabsorption of the yolk. If the area of the yolk sac is significantly greater in a test fish than in the control fish, it suggests that yolk absorption has been affected. On the other hand, the rapid use of yolk leads to a smaller yolk area and indicates enhanced movement or use of the yolk [36].
5.5. Hatchability
A zebrafish embryo is assumed to have been hatched once it is entirely out of the chorion. Certain drugs and extracts can affect hatchability. Lack of hatchability may imply a growth lag or slow development [102]. For example, zebrafish embryos exposed to Millettia pachycarpa extract exhibited a considerable dose-dependent reduction in embryo hatching [6]. Zebrafish embryos start to hatch at 48 hpf in ordinary conditions [8], but percent hatchability after 72 h post-treatment exposure (hpte) is calculated using the following formula [73]:
According to the literature, this model to determine the toxicity of crude extracts or bioactive compounds, and toxicological effects can be compared.
6. Chemical Studies of Embryotoxicity
Changes in fish behavior are sensitive indicators of unintended chemical pollution [151]. The zebrafish model is vital in evaluating the effects on embryonic/larval activity of developmental exposure to harmful compounds [38]. Comprehension and monitoring of these considerations and future revision/harmonization of procedures will reduce the uncertainty of the outcomes for chemical hazard assessments [122].
6.1. Salinity
Several early life researches have been carried on the effects of salinity on zebrafish [152]. When larval zebrafish are subjected to mild environmental stressors, severe salinity shifts lead to a quick locomotive reaction [153]. Lee and colleagues showed that larvae (5 dpf) treated with NaCl exhibit dramatically increased locomotive activity and concentration-dependent reactions. A linear relationship between whole-body cortisol and sodium chloride in the concentration range of 5.84 ppt was shown in zebrafish larvae [154].
6.2. Phytochemical Compounds
Yumnamcha et al. [6] reported the presence of saponins, alkaloids, phenolic compounds, and triterpenoids in the aqueous extract of Millettia pachycarpa. Potent hemolytic activity was found to lead to acute cell membrane death in zebrafish. The toxic function of saponin in the zebrafish embryo is uncertain. However, prior studies have shown that it can be caused by physicochemical characteristics (surfactant) and/or membranolytic impacts on the chorion, a semipermeable membrane that covers the embryo until it hatches [6,155]. Moreover, according to Liu et al. [156] and Yumnamcha et al. [10], some bioactive compounds such as saponin mixtures and alkaloids can cause damage to DNA. Artemisia capillaris was evaluated for its toxicity based on embryotoxicity, and isofraxidine 7-O-(6′-O-p-coumaroyl)-β-glucopyranoside was isolated from this plant. The toxicity, death, and cardiac rates of handled zebrafish embryos are used to diagnose safe and effective concentrations [157].
In contrast to flavone, artificial flavonoids such as kaempherol, 7-hydroxyflavone, 6-methoxyflavon, and 7-methoxyflanone induced considerable toxicity in zebrafish larvae [158]. Additionally, the lack of flavonoids can potentially trigger oxidative stress to cellular molecules, which leads to lower survival rates of zebrafish embryos [12]. Bugel et al. [63] reported that 15 of 24 flavonoids affected at least one or more behavioral and developmental endpoints in zebrafish. However, they focused on two endpoints in their experiment: abnormal spastic behaviors (72 hpf) and alternations in the larval photomotor responses assay (120 hpf). Li et al. [99] exposed zebrafish to Xiaoping at high doses, and many larvae opted to swim at a medium to a low rate, which led to decreased locomotion ability. Changes in larval behavior have been confirmed to be associated with damage to the central nervous system. The function of neural population numbers in animals with a broader central nervous system is being measured using fluorescence and new devices developed over the last decade [159]. In one study, zebrafish embryos subjected to different caffeine dosages demonstrated a decrease in susceptibility to touch-induced movement. Such abnormalities were linked to changes in muscle fibers and axon projections of both primary and secondary motor neurons through immunohistochemistry [160]. Furthermore, toxicological testing may also impact other environmental variables (temperature, water, pH, total hardness or dissolved oxygen) [161].
7. Conclusions
The data collected in this study suggest that zebrafish embryotoxicity tests are able to evaluate drug toxicity and that the zebrafish model offers a suitable replacement for laboratory animals such as rats, mice, and rabbits. As a toxicology model, zebrafish can expose developmental toxicity mechanisms because they are close to mammals. Zebrafish embryos and larvae showed significantly higher susceptibility to toxins than did adult zebrafish. In this review, most of the extracts were polar, such as ethanol, methanol and aqueous extracts, which were used to detect the toxicity and bioactivity. However, the use of the zebrafish model will provide insight into the mechanisms of toxicity of medicinal plants and will help identify and discover new medications for the treatment of human diseases. The zebrafish model is planned as a replacement for models based on higher vertebrate animals to study medicinal plants’ toxicity.
Author Contributions
Project administration, V.L. and H.A.; writing-original draft preparation, A.M.C.; writing-review and editing, A.M.C., V.L.; funding acquisition, V.L. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to thank Ministry of Higher Education, Malaysia for funding support from Fundamental Research Grant Scheme, FRGS (203.CIPPT.6711684).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alafiatayo, A.A.; Lai, K.S.; Syahida, A.; Mahmood, M.; Shaharuddin, N.A. Phytochemical evaluation, embryotoxicity, and teratogenic effects of Curcuma longa extract on zebrafish (Danio rerio). Evid. Based Complement. Altern. Med. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ismail, H.F.; Hashim, Z.; Soon, W.T.; Ab Rahman, N.S.; Zainudin, A.N.; Abdul Majid, F.A. Comparative study of herbal plants on the phenolic and flavonoid content, antioxidant activities and toxicity on cells and zebrafish embryo. J. Tradit. Complement. Med. 2017, 7, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Falcao, M.A.P.; de Souza, L.S.; Dolabella, S.S.; Guimaraes, A.G.; Walker, C.I.B. Zebrafish as an alternative method for determining the embryo toxicity of plant products: A systematic review. Environ. Sci. Pollut. Res. Int. 2018, 25, 35015–35026. [Google Scholar] [CrossRef]
- Gebrelibanos Hiben, M.; Kamelia, L.; de Haan, L.; Spenkelink, B.; Wesseling, S.; Vervoort, J.; Rietjens, I.M.C.M. Hazard assessment of Maerua subcordata (Gilg) DeWolf. for selected endpoints using a battery of in vitro tests. J. Ethnopharmacol. 2019, 241, 111978. [Google Scholar]
- Fürst, R.; Zündorf, I. Evidence-based phytotherapy in Europe: Where do we stand? Planta Med. 2015, 81, 962–967. [Google Scholar] [CrossRef]
- Yumnamcha, T.; Roy, D.; Devi, M.D.; Nongthoma, U. Evaluation of developmental toxicity and apoptotic induction of the aqueous extract of Millettia pachycarpa using zebrafish as model organism. Toxicol. Environ. Chem. 2015, 97, 1363–1381. [Google Scholar] [CrossRef]
- Little, J.G.; Marsman, D.S.; Baker, T.R.; Mahony, C. In silico approach to safety of botanical dietary supplement ingredients utilizing constituent-level characterization. Food Chem. Toxicol. 2017, 107, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, M.; Gong, Z.; Zonyane, S.; Xu, S.; Makunga, N.P. Comparative cardio and developmental toxicity induced by the popular medicinal extract of Sutherlandia frutescens (L.) R.Br. detected using a zebrafish Tuebingen embryo model. BMC Complement. Altern. Med. 2018, 18, 273. [Google Scholar]
- Mclaughlin, J.L.; Chang, C.J.; Smith, D.L. Bench Top Bioassays for the Discovery of Bioactive Natural Products: An Update. In Studies in Natural Products Chemistry; Atta-Ur-Rahaman, Ed.; Elsevier: Amsterdam, The Netherlands, 1991; Volume 9, pp. 383–409. [Google Scholar]
- Yumnamcha, T.; Nongthomba, U.; Devi, M.D. Phytochemical screening and evaluation of genotoxicity and acute toxicity of aqueous extract of Croton tiglium L. Int. J. Sci. Res. Publ. 2014, 4, 1–5. [Google Scholar]
- Caballero, M.V.; Candiracci, M. Zebrafish as Toxicological model for screening and recapitulate human diseases. J. Unexplor. Med. Data 2018, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Thiagarajan, S.K.; Krishnan, K.R.; Ei, T.; Shafie, N.H.; Arapoc, D.J.; Bahari, H. Evaluation of the effect of aqueous Momordica charantia Linn. extract on zebrafish embryo model through acute toxicity assay assessment. Evid. Based Complement. Alternat. Med. 2019, 9152757. [Google Scholar] [CrossRef] [Green Version]
- Bambino, K.; Chu, J. Zebrafish in toxicology and environmental health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar]
- Heideman, W.; Antkiewicz, D.S.; Carney, S.A.; Peterson, R.E. Zebrafish and cardiac toxicology. Cardiovasc. Toxicol. 2005, 5, 203–214. [Google Scholar] [CrossRef]
- Ayub, A.D.; Hock, I.C.; Mat Yusuf, S.N.A.; Abd Kadir, E.; Ngalim, S.H.; Lim, V. Biocompatible disulphide cross-linked sodium alginate derivative nanoparticles for oral colon-targeted drug delivery. Artif. Cells Nanomed. Biotechnol. 2019, 47, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Abuchenari, A.; Hardani, K.; Abazari, S.; Naghdi, F.; Ahmady Keleshteri, M.; Jamavari, A.; Modarresi Chahardehi, A. Clay-reinforced nanocomposites for the slow release of chemical fertilizers and water retention. J. Compos. Comp. 2020, 2, 85–91. [Google Scholar] [CrossRef]
- Nishimura, Y.; Inoue, A.; Sasagawa, S.; Koiwa, J.; Kawaguchi, K.; Kawase, R.; Maruyama, T.; Kim, S.; Tanaka, T. Using zebrafish in systems toxicology for developmental toxicity testing. Congenit. Anom. Kyoto 2016, 56, 18–27. [Google Scholar]
- Han, H.S.; Jang, G.H.; Jun, I.; Seo, H.; Park, J.; Glyn-Jones, S.; Seok, H.K.; Lee, K.H.; Montovani, D.; Kim, Y.C.; et al. Transgenic zebrafish model for quantification and visualization of tissue toxicity caused by alloying elements in newly developed biodegradable metal. Sci. Rep. 2018, 8, 13818. [Google Scholar] [CrossRef]
- Rennekamp, A.J.; Peterson, R.T. 15 years of zebrafish chemical screening. Curr. Opin. Chem. Biol. 2015, 24, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Van der Laan, J.W.; Chapin, R.E.; Haenen, B.; Jacobs, A.C.; Piersma, A. Testing strategies for embryo-fetal toxicity of human pharmaceuticals. Animal models vs. in vitro approaches: A workshop report. Regul. Toxicol. Pharmacol. 2012, 63, 115–123. [Google Scholar] [CrossRef]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of zebrafish in drug discovery toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Jayasinghe, C.D.; Jayawardena, U.A. Toxicity Assessment of Herbal Medicine Using Zebrafish Embryos: A Systematic Review. Evid. Based Complement. Altern. Med. 2019, 7272808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizgirev, I.V.; Revskoy, S. A new zebrafish model for experimental leukemia therapy. Cancer Biol. Ther. 2010, 9, 895–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, D.H.; Roo, B.D.; Nguyen, X.B.; Vervaele, M.; Kecskes, A.; Ny, A.; Copmans, D.; Vriens, H.; Locquet, J.P.; Hoet, P.; et al. Use of zebrafish larvae as a multi-endpoint platform to characterize the toxicity profile of Silica Nanoparticles. Sci. Rep. 2016, 6, 37145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, M.V.; Candiracci, M. Zebrafish as screening model for detecting toxicity and drugs efficacy. J. Unexplor. Med. Data 2018, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.R.; Noyes, P.D.; Tanguay, R.L. Advancements in zebrafish applications for 21st century toxicology. Pharmacol. Ther. 2016, 161, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermsen, S.A.B.; van den Brandhof, E.J.; van der Ven, L.T.M.; Piersma, A.H. Relative embryotoxicity of two classes of chemicals in a modified zebrafish embryotoxicity test and comparison with their in vivo potencies. Toxicol. In Vitro 2011, 25, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Fazry, S.; Mohd Noordin, M.A.; Sanusi, S.; Mat Noor, M.; Aizat, W.M.; Mat Lazim, A.; Dyari, H.R.E.; Jamar, N.H.; Remali, J.; Othman, B.A.; et al. Cytotoxicity and toxicity evaluation of Xanthone crude extract on hypoxic human hepatocellular carcinoma and zebrafish (Danio rerio) embryos. Toxics 2018, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R. Zebrafish Early Life-stages and Adults as A Tool for Ecotoxicity Assessment. Master’s Thesis, Universidade de Aveiro, Aveiro, Portugal, 2009. [Google Scholar]
- Rizzo, L.Y.; Golombek, S.K.; Mertens, M.E.; Pan, Y.; Laaf, D.; Broda, J.; Jayapaul, J.; Mockel, D.; Subr, V.; Hennink, W.E.; et al. In vivo nanotoxicity testing using the zebrafish embryo assay. J. Mater. Chem. B. 2013, 1, 3918–3925. [Google Scholar] [CrossRef]
- D’Amora, M.; Giordani, S. The Utility of Zebrafish as a Model for Screening Developmental Neurotoxicity. Front. Neurosci. 2018, 12, 976. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Koziol, E.; Deniz, F.S.S.; Orhan, I.E.; Marcourt, L.; Budzynska, B.; Wolfender, J.L.; Crawford, A.D.; Skalicka-Wozniak, K. High-performance counter-current chromatography isolation and initial neuroactivity characterization of furanocoumarin derivatives from Peucedanum alsaticum L (Apiaceae). Phytomed 2019, 54, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Zebrafish: A complete animal model to enumerate the nanoparticle toxicity. J. Nanobiotechnol. 2016, 14, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaria, Z.Z.; Benslimane, F.M.; Nasrallah, G.K.; Shurbaji, S.; Younes, N.N.; Mraiche, F.; Da’as, S.I.; Yalcin, H.C. Using zebrafish for investigating the molecular mechanisms of drug-induced cardiotoxicity. BioMed Res. Int. 2018, 2018, 1642684. [Google Scholar] [CrossRef] [PubMed]
- Sant, K.E.; Timme-Laragy, A.R. Zebrafish as a model for toxicological perturbation of yolk and nutrition in the early embryo. Curr. Environ. Health Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Lammer, E.; Wendler, K.; Rawling, J.M.; Belanger, S.E.; Braunbeck, T. Is the fish embryo toxicity test (FET) with the zebrafish (Danio rerio) a potential alternative for the fish acute toxicity test? Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2009, 149, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.; Oliveri, A.; Levin, E.D. Zebrafish model systems for developmental neurobehavioral toxicology. Birth Defects Res. Part C Embryo Today Rev. 2013, 99, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Li, C.; Wang, C.; Peng, Y.; Shu, L.; Jia, X.; Ma, W.; Wang, B. Metabolism of tanshinone IIA, cryptotanshinone and tanshinone I from Radix Salvia miltiorrhiza in zebrafish. Molecules 2012, 17, 8617–8632. [Google Scholar] [CrossRef] [Green Version]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ulimann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Carlson, B.M. Human Embryology and Developmental biology E-book; Elsevier Health Sciences: St. Louis, MI, USA, 2018. [Google Scholar]
- Ali, S.; van Mil, H.G.; Richardson, M.K. Large-scale assessment of the zebrafish embryo as a possible predictive model in toxicity testing. PLoS ONE 2011, 6, e21076. [Google Scholar] [CrossRef] [Green Version]
- Syahbirin, G.; Mumuh, N.; Mohamad, K. Curcuminoid and toxicity levels of ethanol extract of Javanese ginger (Curcuma xanthorriza) on brine shrimp (Artemia salina) larvae and zebrafish (Danio rerio) embryos. Asian J. Pharm. Clin. Res. 2017, 10, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Littleton, R.M.; Hove, J.R. Zebrafish: A nontraditional model of traditional medicine. J. Ethnopharmacol. 2013, 145, 677–685. [Google Scholar]
- Wang, T.; Wang, C.; Wu, Q.; Zheng, K.; Chen, J.; Lan, Y.; Qin, Y.; Mei, W.; Wang, B. Evaluation of tanshinone IIA developmental toxicity in zebrafish embryos. Molecules 2017, 22, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunbeck, T.; Boettcher, M.; Hollert, H.; Kosmehl, T.; Leist, E.; Rudolf, M.; Seitz, N. Towards an alternative for the acute fish LC50 test in chemical assessment: The fish embryo toxicity test goes multi-species -- an update. ALTEX 2005, 22, 87–102. [Google Scholar] [PubMed]
- Yang, L.; Ho, N.Y.; Alshut, R.; Legradi, J.; Weiss, C.; Reischl, M.; Mikut, R.; Liebel, U.; Muller, F.; Strahle, U. Zebrafish embryos as models for embryotoxic and teratological effects of chemicals. Reprod. Toxicol. 2009, 28, 245–253. [Google Scholar] [CrossRef]
- Di Paolo, C.; Seiler, T.B.; Keiter, S.; Hu, M.; Muz, M.; Brack, W.; Hollert, H. The value of zebrafish as an integrative model in effect-directed analysis—A review. Environ. Sci. Eur. 2015, 27, 8. [Google Scholar] [CrossRef] [Green Version]
- Stern, H.M.; Zon, L.I. Cancer genetics and drug discovery in the zebrafish. Nat. Rev. Cancer 2003, 3, 533–539. [Google Scholar] [CrossRef]
- R, E.B.; Jesubatham, P.D.; Berlin Grace, V.M.; Viswanathan, S.; Srividya, S. Non-toxic and non teratogenic extract of Thuja orientalis L. inhibited angiogenesis in zebra fish and suppressed the growth of human lung cancer cell line. Biomed. Pharmacother. 2018, 106, 699–706. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Gallagher, E.; O’Connor, P.; Prieto, J.; Mora-soler, L.; Grealy, M.; Hayes, M. Development of a seaweed derived platelet activating factor acetylhydrolase (PAF-AH) inhibitory hydrolysate, synthesis of inhibitory peptides and assessment of their toxicity using the zebrafish larvae assay. Peptides 2013, 50, 119–124. [Google Scholar] [CrossRef]
- Bertelli, P.R.; Biegelmeyer, R.; Rico, E.P.; Klein-Junior, L.C.; Toson, N.S.B.; Minetto, L.; Bordignon, S.A.L.; Gasper, A.L.; Moura, S.; de Oliveira, D.L.; et al. Toxicological profile and acetylcholinesterase inhibitory potential of Palicourea deflexa, a source of beta-carboline alkaloids. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 201, 44–50. [Google Scholar] [CrossRef]
- Tolentino, J.; Undan, J.R. Embryo-toxicity and teratogenicity of Derris elliptica leaf extract on zebraish (Danio rerio) embryos. Int. J. Pure Appl. Biosci. 2016, 4, 16–20. [Google Scholar] [CrossRef]
- He, J.H.; Gao, J.M.; Huang, C.J.; Li, C.Q. Zebrafish models for assessing developmental and reproductive toxicity. Neurotoxicol. Teratol. 2014, 42, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yu, L.; Liu, C.; Yu, K.; Shi, X.; Yeung, L.W.Y.; Lam, P.K.S.; Wu, R.S.S.; Zhou, B. Hexabromocyclododecane-induced developmental toxicity and apoptosis in zebrafish embryos. Aquat. Toxicol. 2009, 93, 29–36. [Google Scholar] [CrossRef]
- Carwan, M.J., III; Heiden, T.K.; Tomasiewicz, H.C. The utility of zebrafish as a model for toxicological research. In Biochemistry and Molecular Biology of Fishes; Mommsen, T.P., Moon, T.W., Eds.; Chapter 1; Elsevier: New York, NY, USA, 2005; Volume 6, pp. 3–41. [Google Scholar]
- Knobel, M.; Busser, F.J.M.; Rico-Rico, A.; Kramer, N.I.; Hermens, J.L.M.; Hafner, C.; Tanneberger, K.; Schirmer, K.; Scholz, S. Predicting adult fish acute lethality with the zebrafish embryo: Relevance of test duration, endpoints, compound properties, and exposure concentration analysis. Environ. Sci. Technol. 2012, 46, 9690–9700. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, A.; Lewis, M.A.; Menzer, R.E. Methods in environmental toxicology. In Principles and Methods of Toxicology, 4th ed.; Hayes, A.W., Ed.; Chapter 42; CRC Press, Taylor & Francis Group: London, UK, 2001; pp. 1759–1801. [Google Scholar]
- Massei, R.; Hollert, H.; Krauss, M.; von Tumpling, W.; Weidauer, C.; Haglund, P.; Kuster, E.; Gallampois, C.; Tysklind, M.; Brack, W. Toxicity and neurotoxicity profiling of contaminated sediments from Gulf of Bothnia (Sweden): A multi-endpoint assay with Zebrafish embryos. Environ. Sci. Eur. 2019, 31, 8. [Google Scholar] [CrossRef] [Green Version]
- Nagel, R. DarT: The embryo test with the zebrafish Danio rerio--a general model in ecotoxicology and toxicology. ALTEX 2002, 19, 38–48. [Google Scholar] [PubMed]
- Liedtke, A.; Muncke, J.; Rufenacht, K.; Eggen, R.I.L. Molecular multi-effect screening of environmental pollutants using the MolDarT. Environ. Toxicol. 2008, 23, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Njiwa, J.R.K.; Suter, M.J.-F.; Eggen, R.I. Zebrafish Embryo Toxicity Assay, Combining Molecular and Integrative Endpoints at Various Developmental Stages. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 4481–4489. [Google Scholar]
- Bugel, S.M.; Bonventre, J.A.; Tanguay, R.L. Comparative developmenal toxicity of flavonoids using an integrative zebrafish system. Toxicol. Sci. 2016, 154, 55–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haldi, M.; HArden, M.; D’Amico, L.; DeLise, A.; Seng, W.L. Developmental toxicity assessment in zebrafish. In Zebrafish: Methods for Assessing Drug Safety and Toxicity; McGrath, P., Ed.; Phylonix: Cambridge, MA, USA, 2011. [Google Scholar]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belnger, S.E.; Braunbeck, T.; Carr, G.J.; Cenijn, P.; Fochtman, P.; Gourmelon, A.; Hubler, N.; et al. OECD validation study to assess intra and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef]
- OECD. Fish Embryo Toxicity Test; Organization for Economic Cooperation and Development: Paris, France, 2006. [Google Scholar]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawia, J.S.; Craddock, C.; Kyzar, E.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Chueh, T.C.; Hsu, L.S.; Kao, C.M.; Hsu, T.W.; Liao, H.Y.; Wang, K.Y.; Chen, S.C. Transcriptome analysis of zebrafish embryos exposed to deltamethrin. Environ. Toxicol. 2017, 32, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Han, Y.; Liu, Y.; Zhang, J.; Hu, C. Relationship between fluoroquinolone structure and neurotoxicity revealed by zebrafish neurobehavior. Chem. Res. Toxicol. 2018, 31, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.Y.; Cowden, J.; Simmons, S.O.; Padilla, S.; Ramabhadran, R. Gene expression changes in developing zebrafish as potential markers for rapid developmental neurotoxicity screening. Neurotoxicol. Teratol. 2009, 32, 91–98. [Google Scholar] [CrossRef] [PubMed]
- OECD. Fish Acute Toxicity Test; Organization for Economic Cooperation and Development: Medmenham, UK, 1992. [Google Scholar]
- Shaikh, A.; Kohale, K.; Ibrahim, M.; Khan, M. Teratogenic effects of aqueous extract of Ficus glomerata leaf during embryonic development in zebrafish (Danio rerio). J. Appl. Pharm. Sci. 2019, 9, 107–111. [Google Scholar]
- Paramanik, V.; Basumatary, M.J.; Nagesh, G.; Kushwaha, J.K.; Kurrey, K. Ethanolic extracts of candidate Indian Traditional Medicines Acorus calamus, Terminalia chebula and Achyranthes aspera are neuroprotective in Zebrafish. IBRO Rep. 2019, 6, S247. [Google Scholar] [CrossRef]
- Yan, C.; Zhang, S.; Wang, C.; Zhang, Q. A fructooligosaccharide from Achyranthes bidentata inhibits osteoporosis by stimulating bone formation. Carbohydr. Polym. 2019, 210, 110–118. [Google Scholar] [CrossRef]
- Aleksandar, P.; Dragana, M.C.; Nebojsa, J.; Biljana, N.; Natasa, S.; Branka, V.; Jelena, K.V. Wild edible onions—Allium flavum and Allium carinatum—Successfully prevent adverse effects of chemotherapeutic drug doxorubicin. Biomed. Pharmacother. 2019, 109, 2482–2491. [Google Scholar] [CrossRef]
- Ponrasu, T.; Ganeshkumar, M.; Suguna, L. Developmental toxicity evaluation of ethanolic extract of Annona squamosa in zebrafish (Danio rerio) embryo. J. Pharm. Res. 2012, 5, 277–279. [Google Scholar]
- Grisola, M.A.C.; Fuentes, R.G. Phenotype-based screening of selected mangrove methanolic crude extracts with anti-melanogenic activity using zebrafish (Danio rerio) as a model. ScienceAsia 2017, 43, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Batista, F.L.A.; Lima, L.M.G.; Abrante, I.A.; de Araujo, J.I.F.; Batista, F.L.A.; Abrante, I.A.; Lima, M.C.L.; do Prado, B.S.; Moura, L.F.W.G.; Guedes, M.I.F.; et al. Antinociceptive activity of ethanolic extract of Azadirachta indica A. Juss (Neem, Meliaceae) fruit through opioid, glutamatergic and acid-sensitive ion pathways in adult zebrafish (Danio rerio). Biomed. Pharmacother. 2018, 108, 408–416. [Google Scholar]
- Teh, L.E.; Kue, C.S.; Ng, C.H.; Lau, B.F. Toxicity effect of Bougainvillea glabra (paper flower) water extracts on zebrafish embryo. INNOSC Ther. Pharmacol. Sci. 2019, 2, 23–26. [Google Scholar] [CrossRef] [Green Version]
- De Vera, J.S.; De Castro, M.E.G.; Dulay, R.M.R. Phytochemical constituents and teratogenic effect of lyophilized extracts of Bixa orellana L. (Achuete) and Piper betle L. (Ikmo) leaves in Danio rerio embryos. Der Pharma Chem. 2016, 8, 432–437. [Google Scholar]
- Vargas-Mendez, L.Y.; Rosado-Solano, D.N.; Sanabria-Florez, P.L.; Puerto-Galvis, C.E.; Kouznetsov, V. In vitro antioxidant and anticholinesterase activities and in vivo toxicological assessment (Zebrafish embryo model) of ethanolic extracts of Capsicum chinense Jacq. J. Med. Plants Res. 2016, 10, 59–66. [Google Scholar]
- Xia, Q.; Ma, Z.; Luo, J.; Wang, Y.; Li, T.; Feng, Y.; Ni, Y.; Zou, Q.; Lin, R. Assay for the developmental toxicity of safflower (Carthamus tinctorius L.) to zebrafish embryos/larvae. J. Tradit. Chin. Med. Sci. 2017, 4, 71–81. [Google Scholar]
- Murugesu, S.; Khatib, A.; Ahmad, Q.U.; Ibrahim, Z.; Uzir, B.F.; Benchoula, K.; Yusoff, N.I.N.; Perumal, V.; Aljami, M.F.; Salamah, S.; et al. Toxicity study on Clinacanthus nutans leaf hexane fraction using Danio rerio embryos. Toxicol. Rep. 2019, 6, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Paucar, A.M.; Serruys, A.S.K.; Afrikanova, T.; Maes, J.; De Borggraeve, W.; Alen, J.; Leon-Tamariz, F.; Wilches-Arizabala, I.M.; Crawford, A.M.; de Witte, P.A.M.; et al. Anticonvulsant activity of bisabolene sesquiterpenoids of Curcuma longa. Epilepsy Behav. 2012, 24, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.R.; Vincent, S.G. Cynodon dactylon and Sida acuta extracts impact on the function of the cardiovascular system in zebrafish embryos. J. Biomed. Res. 2012, 26, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Memita, A.Y.B.; Macalinao, S.D.; Reyes, L.E.G.; Damian, E.E.; Dulay, R.M.R. Toxicity and teratogenicity of Dieffenbachia amoena leaf extract. Int. J. Biol. Pharm. Allied Sci. 2018, 7, 1591–1600. [Google Scholar]
- Flaviano, J.F.; Kenniker, M.A.A.; Ganda, F.J.C.; Sacedor, J.; Lee, M.Y.S.; Cabuhat, K.S.P. Assessment of the toxic and teratogenic effects of Kamagong (Diospyros discolor Willd) leaves extracts to the developing embryo of zebrafish (Danio rerio). Egypt. Acad. J. Biol. Sci. B. Zool. 2018, 10, 53–60. [Google Scholar] [CrossRef]
- Ponpornpisit, A.; Pirarat, N.; Suthikrai, W.; Binwihok, A. Toxicity test of Kameng (Ecliptra prostrate Linn.) and Kradhuawean (Spilanthes acmella (Linn.) Murr.) to early life stage of zebrafish (Danio rerio). Thai. J. Vet. Med. 2011, 41, 523–527. [Google Scholar]
- Xavier, J.; Kripasana, K. Acute Toxicity of Leaf Extracts of Enydra fluctuans Lour in Zebrafish (Danio rerio Hamilton). Sci. Cairo 2020, 2020, 3965376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, B.; Feng, Y.; Liu, J.; Ma, Z.; Zheng, J.; Xia, Q.; Ni, Y.; Li, F.; Lin, R. Anti-angiogenic activity of water extract from Euphorbia pekinensis Rupr. J. Ethnopharmacol. 2017, 206, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Ma, L.; Li, S.; Cui, K.; Lei, L.; Ye, Z. Evaluation of the cardiotoxicity of evodiamine in vitro and in vivo. Molecules 2017, 22, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.L.; Li, K.; Lin, Q.H.; Ren, J.; He, Z.H.; Li, H.; Shen, N.; Wei, P.; Feng, F.; He, M.F. Gambogic acid causes fin developmental defect in zebrafish embryo partially via retinoic acid signaling. Reprod. Toxicol. 2016, 63, 161–168. [Google Scholar] [CrossRef]
- Sajkowska-Kozielewicz, J.J.; Kozielewicz, P.; Barnes, N.M.; Wawer, I.; Paradowska, K. Antioxidant, cytotoxic, and antiproliferative activities and total polyphenol contents of the extracts of Geissospermum reticulatum bark. Oxid. Med. Cell. Longev. 2016, 2016, 2573580. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, I.V.F.; de Souza, G.C.; Santana, G.R.; Duarte, J.L.; Fernandes, C.P.; Keia, H.; Velazquez-Moyado, J.A.; Navarrete, A.; Ferreira, I.M.; Carvalho, H.O. Histopathology in zebrafish (Danio rerio) to evaluate the toxicity of medicine: An anti-inflammatory phytomedicine with Janaguba milk (Himatanthus drasticus Plumel). Histopathol. Update 2018, 39–64. [Google Scholar]
- Lira, S.M.; Dionisio, A.P.; Holanda, M.O.; Marques, C.G.; da Silva, G.S.; Correa, L.C.; Santos, G.B.M.; de Abreu, F.A.P.; Magalhaes, F.E.A.; Reboucas, E.D.; et al. Metabolic profile of pitaya (Hylocereus polyrhizus (FA.C. Weber) Britton & Rose) by UPLC-QTOF-MSE and assessment of its toxicity and anxiolytic-like effect in adult zebrafish. Food Res. Intl. 2020, 127, 108701. [Google Scholar]
- He, Y.L.; Shi, J.Y.; Peng, C.; Hu, L.J.; Liu, J.; Zhou, Q.M.; Guo, L.; Xiong, L. Angiogenic effect of motherwort (Leonurus japonicus) alkaloids and toxicity of motherwort essential oil on zebrafish embryos. Fitoterapia 2018, 128, 36–42. [Google Scholar] [CrossRef]
- Wang, W.; Feng, S.; Xiong, Z. Protective effect of polysaccharide from Ligusticum chuanxiong hort against H2O2-induced toxicity in zebrafish embryo. Carbohydr. Polym. 2019, 221, 73–83. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Liu, K.; He, Q.; Sun, C.; Han, J.; Han, L.; Tian, Q. Xiaoaiping induces developmental toxicity in zebrafish embryos through activation of ER stress, apoptosis and the Wnt pathway. Front Pharmacol. 2018, 9, 1250. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.F.; Abutaha, N.; Nasr, F.A.; Alqahtani, A.S.; Noman, O.M.; Wadaan, M.A.M. Bitter gourd (Momordica charantia) possess developmental toxicity as revealed by screening the seeds and fruit extracts in zebrafish embryos. BMC Complement. Altern. Med. 2019, 19, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.M.R.; Agpaoa, A.R.; Sayson, A.D.; De Castro, M.E.G.; Dulay, R.M.R. Toxic and teratogenic effects of water leaf extract of Momordica charantia in zebrafish (Danio rerio) embryos. Der Pharma Chem. 2017, 9, 119–122. [Google Scholar]
- David, C.R.S.; Angeles, A.; Angoluan, R.C.; Santos, J.P.E.; David, E.S.; Dulay, R.M.R. Moringa oleifera (Malunggay) water extracts exhibit embryo-toxic and teratogenic activity in zebrafish (Danio rerio) embryo model. Der Pharm. Lett. 2016, 8, 163–168. [Google Scholar]
- Elsayed, E.A.; Farooq, M.; Sharaf-Eldin, M.A.; El-Enshasy, H.A.; Wadaan, M. Evaluation of developmental toxicity and anti-angiogenic potential of essential oils from Moringa oleifera and Moringa peregrina seeds in zebrafish (Danio rerio) model. S. Afr. J. Bot. 2019, 129, 229–237. [Google Scholar] [CrossRef]
- De Santana Silva, L.L.; Alves, R.N.; de Paulo, D.V.; da Silva, J.D.F. Ecotoxicity of water-soluble lectin from Moringa oleifera seeds to zebrafish (Danio rerio) embryos and larvae. Chemosphere 2017, 185, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Babic, S.; Malev, O.; Pflieger, M.; Lebedev, A.T.; Mazur, D.M.; Kuzic, A.; Coz-Rakovac, R.; Trebse, P. Toxicity evaluation of olive oil mill wastewater and its polar fraction using multiple whole-organism bioassays. Sci. Total Environ. 2019, 686, 903–914. [Google Scholar] [CrossRef]
- Safavi, F.; Farimani, M.M.; Golalipour, M.; Leung, P.-C.; Lau, K.-M.; Kwok, H.-F.; Wong, C.-W.; Bayat, H.; Lau, C.B.-S. Investigations on the wound healing properties of Onosma dichroantha Boiss root extracts. S. Afr. J. Bot. 2019, 125, 344–352. [Google Scholar] [CrossRef]
- Santhoshkumar, J.; Sowmya, B.; Kumar, S.V.; Rajeshkumar, S. Toxicology evaluation and antidermatophytic activity of silver nanoparticles synthesized using leaf extract of Passiflora caerulea. S. Afr. J. Chem. Eng. 2019, 29, 17–23. [Google Scholar] [CrossRef]
- Lamban, I.Q.; Balbeuna, E.S.; Lee, M.Y.S.; Sacdalan, M.; Carpio JR, A.P.; Cabuhat, K.S.P. Toxic and teratogenic effects of Sampa-Sampalukan (Phyllanthus niruri) leaves extract using Danio rerio embryo assay. Egypt. Acad. J. Biol. Sci. 2019, 11, 109–115. [Google Scholar] [CrossRef]
- Thanh, D.T.H.; Thanh, N.L.; Thang, N.D. Toxicological and melanin synthesis effects of Polygonum multiflorum root extracts on zebrafish embryos and human melanocytes. Biomed. Res. Ther. 2016, 3, 808–818. [Google Scholar] [CrossRef]
- Yang, J.B.; Li, W.F.; Liu, Y.; Wang, Q.; Cheng, X.L.; Wei, F.; Wang, A.G.; Jin, H.T.; Ma, S.C. Acute toxicity screening of different extractions, components and constituents of Polygonum multiflorum Thunb. on zebrafish (Danio rerio) embryos in vivo. Biomed. Pharmacother. 2018, 99, 205–213. [Google Scholar] [PubMed]
- Xia, Q.; Wei, L.; Zhang, Y.; Kong, H.; Shi, Y.; Wang, X.; Chen, X.; Han, L.; Liu, K. Psoralen induces developmental toxicity in zebrafish embryos/larvae through oxidative stress, apoptosis, and energy metabolism disorder. Front. Pharmacol. 2018, 9, 1457. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, I.; Permadi, K.; Hartati, R.; Damayanti, S. Ethanolic extract of pomegranate (Punica granatum L) peel: Acute toxicity tests on zebrafish (Danio rerio) embryos and its toxicity prediction by in silico. J. Appl. Pharm. Sci. 2018, 8, 082–086. [Google Scholar]
- Huang, Y.; Bai, Y.; Wang, Y.; Kong, H. Solidago canadensis L. extracts to control algal (Microcystis) blooms in ponds. Ecol. Eng. 2014, 70, 263–267. [Google Scholar] [CrossRef]
- Dos Santos Sampaio, T.I.; de Melo, N.C.; de Freitas Paiva, B.T.; da Silva Aleluia, G.A.; da Silva Neto, F.L.P.; da Silva, H.R.; Keita, H.; Cruz, R.A.S.; Sanchez-Ortiz, B.L.; Pineda-Pena, E.A.; et al. Leaves of Spondias mombin L. a traditional anxiolytic and antidepressant: Pharmacological evaluation on zebrafish (Danio rerio). J. Ethnopharmacol. 2018, 224, 563–578. [Google Scholar]
- Kumar, P.B.S.; Kar, B.; Dolai, N.; Haldar, P.K. Study on developmental toxicity and behavioral safety of Streblus asper Lour. bark on zebrafish embryos. Indian J. Nat. Prod. Res. 2013, 4, 255–259. [Google Scholar]
- Zonyane, S.; Chen, L.; Xu, M.J.; Gong, Z.N.; Xu, S.; Makung, N.P. Geographic-based metabolomic variation and toxicity analysis of Sutherlandia frutescens L. R.Br.—An emerging medicinal crop in South Africa. Ind. Crops Prod. 2019, 133, 414–423. [Google Scholar]
- Russo, H.M.; Queiroz, E.F.; Marcourt, L.; Crawford, A.D.; Bolzani, V.; Wolfender, J.L. Chemical profile and toxicity study of Tetrapterys multiglandulosa secondary metabolites using zebrafish toxicity model. In Brazilian Conference on Natural Products and Annual Meeting on Micromolecular Evolution, Systematics and Ecology; GALOÁ: Campinas, Brazil, 2018. [Google Scholar]
- Romagosa, C.M.R.; David, E.S.; Dulay, R.M.R. Embryo-toxic and teratogenic effects of Tinospora cordifolia leaves and bark extracts in Zebrafish (Danio rerio) embryos. Asian J. Plant Sci. Res. 2016, 6, 37–41. [Google Scholar]
- Aleksic, I.; Ristivojevic, P.; Pavic, A.; Radojevic, I.; Comic, L.R.; Vasiljevic, B.; Opsenica, D.; Milojkovic-Opsenica, D.; Senerovic, L. Anti-quorum sensing activity, toxicity in zebrafish (Danio rerio) embryos and phytochemical characterization of Trapa natans leaf extracts. J. Ethnopharmacol. 2018, 222, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.P.; Feng, F.; Zhang, X.Q.; Liu, X.X.; Wang, Y.B.; She, J.X.; He, Z.H.; He, M.F. Toxicity assessment of 7 anticancer compounds in zebrafish. Int. J. Toxicol. 2014, 33, 98–105. [Google Scholar] [CrossRef]
- Puerto Galvis, C.E.; Kouznetsov, V.V. Synthesis of zanthoxylamide protoalkaloids and their in silico ADME-Tox screening and in vivo toxicity assessment in zebrafish embryos. Eur. J. Pharm. Sci. 2019, 127, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Ogungbemi, A.; Leuthold, D.; Scholz, S.; Kuster, E. Hypo- or hyperactivity of zebrafish embryos provoked by neuroactive substances: A review on how experimental parameters impact the predictability of behavior changes. Environ. Sci. Eur. 2019, 31, 88. [Google Scholar] [CrossRef]
- Ma, C.; Parng, C.; Wen, L.S.; Zhang, C.; Willett, C.; McGrath, P. Zebrafish-an in vivo model for drug screening. Innov. Pharm. Technol. 2003, 3, 38–45. [Google Scholar]
- Truong, L.; Harper, S.L.; Tanguay, R.L. Evaluation of embryotoxicity using the zebrafish model. Methods Mol. Biol. 2017, 691, 271–279. [Google Scholar]
- Driever, W.; Stemple, D.; Schier, A.; Solnica-Krezel, L. Zebrafish: Genetic tools for studying vertebrate development. Trends Genet. 1994, 10, 152–159. [Google Scholar] [CrossRef]
- OECD. Guidelines for Testing of Chemical-Fish Embryo Acute Toxicity (FET) Test; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Calienni, M.N.; Feas, D.A.; Igartua, D.E.; Chiaramoni, N.S.; Alonso, S.V.; Prieto, M.J. Nanotoxicological and teratogenic effects: A linkage between dendrimer surface charge and zebrafish developmental stages. Toxicol. Appl. Pharmacol. 2017, 337, 1–11. [Google Scholar] [CrossRef]
- Yamashita, M. Apoptosis in zebrafish development. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 731–742. [Google Scholar] [CrossRef]
- Haffter, P.; Granato, M.; Brand, M.; Mullins, M.C.; Hammerschmidt, M.; Kane, D.A.; Odenthal, J.; van Eeden, F.J.; Jiang, Y.J.; Heisenberg, C.P.; et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 1996, 123, 1–36. [Google Scholar]
- Li, H.; Yu, S.; Cao, F.; Wang, C.; Zheng, M.; Li, X.; Qiu, L. Developmental toxicity and potential mechanisms of pyraoxystrobin to zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2018, 151, 1–9. [Google Scholar] [CrossRef]
- Bakkers, J. Zebrafish as a model to study cardiac development and human cardiac disease. Cardiovasc. Res. 2011, 91, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Thorarensen, H.; Galiugher, P.E.; Farrel, A.P. The limitations of heart rate as a predictor of metabolic rate in fish. J. Fish Biol. 1996, 49, 226–236. [Google Scholar] [CrossRef]
- Gao, X.Y.; Li, K.; Jiang, L.L.; He, M.F.; Pu, C.H.; Kang, D.; Xie, J. Developmental toxicity of auranofin in zebrafish embryos. J. Appl. Toxicol. 2017, 37, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Liu, H.; Fang, C.; Liu, Y.; Liu, X.; Liu, J. Cardiotoxicity evaluation and comparison of diterpene alkaloids on zebrafish. Drug Chem. Toxicol. 2019, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.P.; Pieperhoff, S. Design and physiology of the heart/cardiac anatomy in fishes. Encycl. Fish Physiol. 2011, 2, 998–1005. [Google Scholar]
- Hu, N.; Sedmera, D.; Yost, H.J.; Clark, E.B. Structure and function of the developing zebrafish heart. Anat. Rec. 2000, 260, 148–157. [Google Scholar] [CrossRef]
- Yamauchi, H.; Hotta, Y.; Konishi, M.; Miyake, A.; Kawahara, A.; Itoh, N. Fgf21 is essential for haematopoiesis in zebrafish. EMBO Rep. 2006, 7, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, X.; He, Q.; Chen, X. Toxic effects of celastrol on embryonic development of zebrafish (Danio rerio). Drug Chem. Toxicol. 2011, 34, 61–65. [Google Scholar] [CrossRef]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Kinna, G.; Kolle, G.; Carter, A.; Lieschke, G.J.; Perkins, A.; Little, M.H. Knockdown of zebrafish crim1 results in a bent tail phenotype with defects in somite and vascular development. Mech. Dev. 2006, 123, 277–287. [Google Scholar] [CrossRef]
- Cinar, K.; Aksoy, A.; Emre, Y.; Asti, R.N. The histology and histochemical aspects of gills of the flower fish, Pseudophoxinus antalyae. Vet. Res. Commun. 2009, 33, 453. [Google Scholar] [CrossRef]
- Mauceri, A.; Fossi, M.C.; Leonzio, C.; Ancora, S.; Minniti, F.; Maisano, M.; Ferrando, S.; Fasulo, S. Stress factors in the gills of Liza aurata (Perciformes, Mugilidae) living in polluted environments. Ital. J. Zool. 2005, 72, 285–292. [Google Scholar] [CrossRef]
- Hughes, G. Introduction to the study of gills. Semin. Ser.-Soc. Exp. Biol 1982, 16, 1–24.ill. [Google Scholar]
- Evans, D.H. The fish gill: Site of action and model for toxic effects of environmental pollutants. Environ. Health Perspect. 1987, 71, 47–58. [Google Scholar] [CrossRef]
- Srivastava, N.; Kumari, U.; Rai, A.K.; Mittal, S.; Mittal, A.K. Alterations in the gill filaments and secondary lamellae of Cirrhinus mrigala exposed to “nuvan,” an organophosphorus insecticide. J. Histol. 2014, 190139. [Google Scholar] [CrossRef] [Green Version]
- Seok, S.-H.; Baek, M.-W.; Lee, H.-Y.; Kim, D.-J.; Na, Y.-R.; Noh, K.-J.; Park, S.-H.; Lee, H.-K.; Lee, B.-H.; Ryu, D.-Y.; et al. Quantitative GFP fluorescence as an indicator of arsenite developmental toxicity in mosaic heat shock protein 70 transgenic zebrafish. Toxicol. Appl. Pharmacol. 2007, 225, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Rajini, A.; Revathy, K.; Selvam, G. Histopathological changes in tissues of Danio rerio exposed to sub lethal concentration of combination pesticide. Indian J. Sci. Technol. 2015, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vutukuru, S.S. Assessment of the Histological Changes in the Gill Architecture of Zebrafish (Danio rerio) Exposed to Nicotine and Cotinine Extracts from Cigarette Tar. EC Pharmacol. Toxicol. 2019, 7, 888–895. [Google Scholar]
- Ertzer, R.; Müller, F.; Hadzhiev, Y.; Rathnam, S.; Fischer, N.; Rastegar, S.; Strähle, U. Cooperation of sonic hedgehog enhancers in midline expression. Dev. Biol. 2007, 301, 578–589. [Google Scholar] [CrossRef]
- Park, H.; Lee, J.-Y.; Park, S.; Song, G.; Lim, W. Developmental toxicity and angiogenic defects of etoxazole exposed zebrafish (Danio rerio) larvae. Aquat. Toxicol. 2019, 217, 105324. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Han, X.; Huang, T. The use of zebrafish (Danio rerio) behavioral responses in identifying sublethal exposures to deltamethrin. Intl. J. Environ. Res. Public Health 2014, 11, 3650–3660. [Google Scholar] [CrossRef] [Green Version]
- Dabrowski, K.; Miller, M. Contested Paradigm in Raising Zebrafish (Danio rerio). Zebrafish 2018, 15, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.B.; Schwab, T.L.; Sigafoos, A.N.; Gauerke, J.L.; Krug, R.G., 2nd; Serres, M.R.; Jacobs, D.C.; Cotter, R.P.; Das, B.; Peterson, M.O.; et al. Novel zebrafish behavioral assay to identify modifiers of the rapid, nongenomic stress response. Genes Brain Behav. 2019, 18, e12549. [Google Scholar] [CrossRef] [PubMed]
- De Marco, R.J.; Groneberg, A.H.; Yeh, C.-M.; Treviñ, M.; Ryu, S. The behavior of larval zebrafish reveals stressor-mediated anorexia during early vertebrate development. Front. Behav. Neurosci. 2014, 8, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, K.R.; Johnson, I.T.; Fenwick, G.R. The chemistry and biological significance of saponins in foods and feedingstuffs. Crit. Rev. Food Sci. Nutr. 1987, 26, 27–135. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Di iorgio, C.; Lamidi, M.; Elias, R.; Ollivier, E.; De Meo, M.P. Genotoxic and clastogenic activity of saponins extracted from Nauclea bark as assessed by the micronucleus and the comet assays in Chinese hamster ovary cells. J. Ethnopharmacol. 2011, 137, 176–183. [Google Scholar] [CrossRef]
- Yim, S.-H.; Tabassum, N.; Kim, W.H.; Cho, H.; Lee, J.H.; Batkhuu, G.J.; Kim, H.J.; Oh, W.K.; Jung, D.W.; Williams, D.R. Isolation and characterization of isofraxidin 7-O-(6′-Op-Coumaroyl)-β-glucopyranoside from Artemisia capillaris Thunberg: A novel, nontoxic hyperpigmentation agent that is effective in vivo. Evid. Based Complement. Altern. Med. 2017, 1401279. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Yang, Z.S.; Wen, C.C.; Chang, Y.S.; Wang, B.C.; Hsiao, C.A.; Shih, T.L. Evaluation of the structure-activity relationship of flavonoids as antioxidants and toxicants of zebrafish larvae. Food Chem. 2012, 134, 717–724. [Google Scholar] [CrossRef]
- Yoon, Y.; Park, J.; Taniguchi, A.; Kohsaka, H.; Nakae, K.; Nonaka, S.; Ishii, S.; Nose, A. System level analysis of motor-related neural activities in larval Drosophila. J. Neurogenet. 2019, 33, 179–189. [Google Scholar] [CrossRef]
- Chen, Y.H.; Huang, Y.H.; Wen, C.C.; Wang, Y.H.; Chen, W.L.; Chen, L.C.; Tsay, H.J. Movement disorder and neuromuscular change in zebrafish embryos after exposure to caffeine. Neurotoxicol. Teratol. 2008, 30, 440–447. [Google Scholar] [CrossRef]
- Mendis, J.C.; Tennakoon, T.K.; Jayasinghe, C.D. Zebrafish embryo toxicity of a binary mixture of pyrethroid insecticides: D-tetramethrin and cyphenothrin. J. Toxicol. 2018, 2018, 4182694. [Google Scholar] [CrossRef]
Figure 1.
Comparative analysis of human, rat, and zebrafish based on the early stages of development.
Figure 1.
Comparative analysis of human, rat, and zebrafish based on the early stages of development.


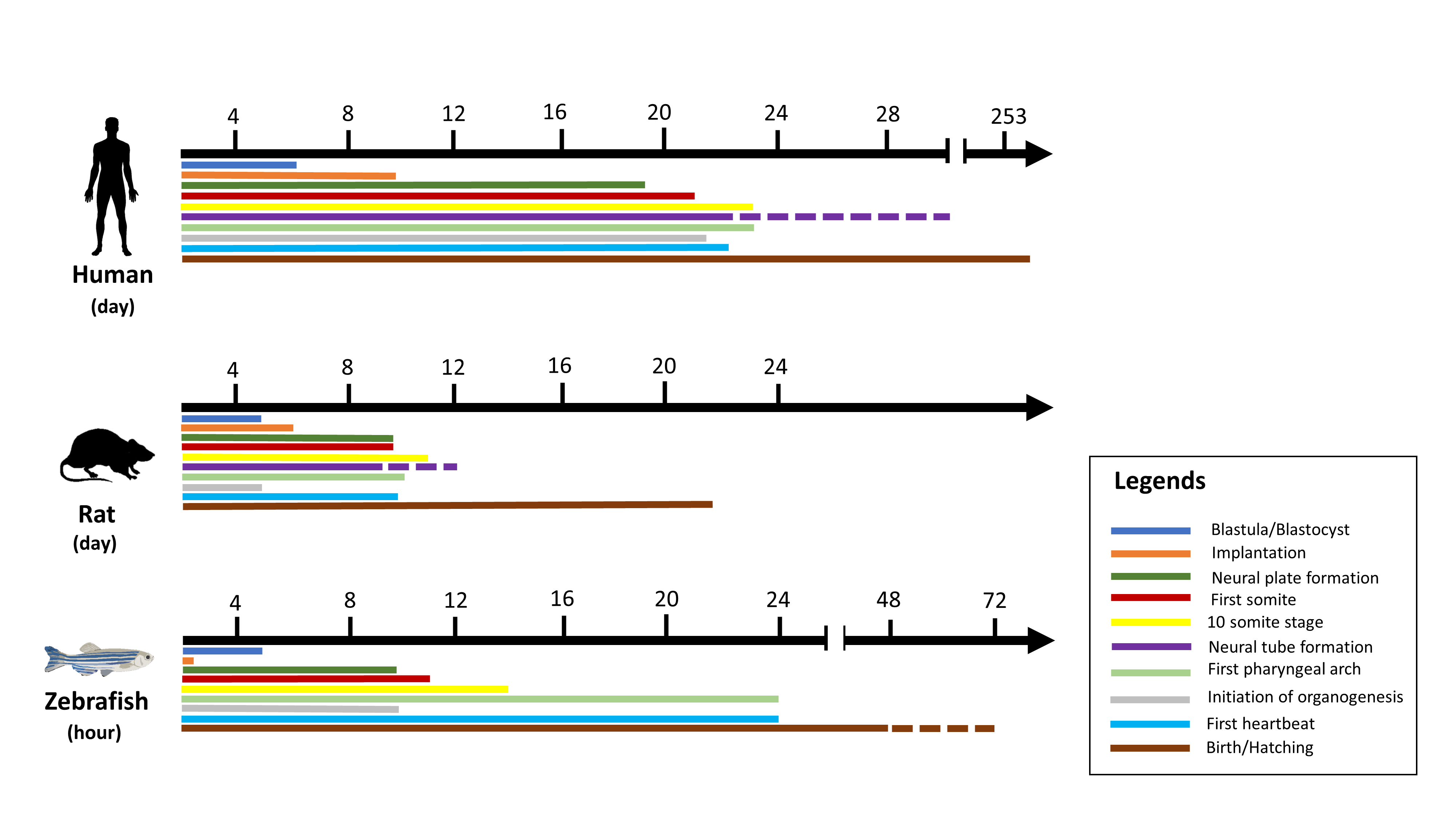{kind=link}
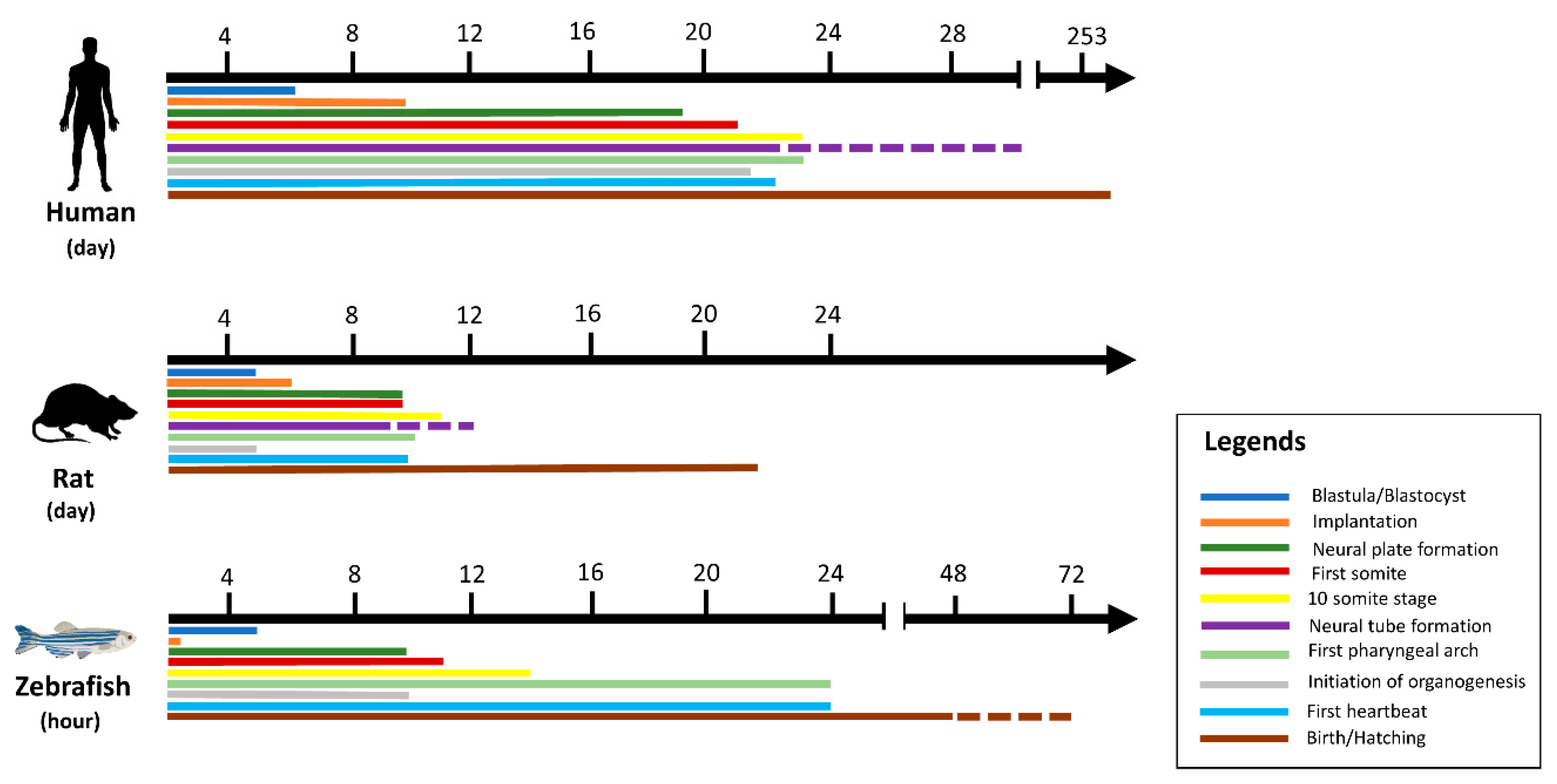{kind=link}
| Cells | Fruit Fly | Zebrafish | Rodents | Humans | |
|---|---|---|---|---|---|
| Benefits | Fast, simple, inexpensive | Short period of generation | Fast, simple, inexpensive | Complexity of body | High value translation |
| Well-standardized | Work easily with | Complexity of body | Adequate predictiveness | Credible and logical | |
| Many probable readings | Low maintenance costs | Ethical examination in vivo | Administration route | Dosage details | |
| Limitations | Modest predictivity | Genetically far from human beings | Modest flexibility | Time-consuming and costly | Time-consuming and costly |
| Simplified process | Simple anatomy | Moderate foresight | Comprehensive regulation | Comprehensive regulation | |
| Low value translation | No efficient immune system | Moderate value translation | Ethical limits | Ethical limits |
Table 2.
Zebrafish bioassays and measured endpoints.
| Life Stage | Measured Endpoints | References |
|---|---|---|
| Egg | Survival, hatching, teratology | [1,60,67] |
| Larvae | Survival, abnormalities, enzyme activities and behavior | [67,68,69,70] |
| Adults | Survival, behavior, enzyme activities and genotoxicity | [71,72] |
Table 3.
Existing studies that screened toxicity of medicinal plants using the zebrafish model.
| Plant/Family | Common Name | Extraction/Part of Plant | Concentrations Range (µg/mL) | LC50 (µg/mL) | Characteristics of Effect | Reference |
|---|---|---|---|---|---|---|
| Acorus calamus/Acoraceae | Sweet flag | Ethanol/rhizome | n.a. | n.a. | Fish treated with this extract returned to normal condition and revealed a decrease level in superoxide dismutase and decreased mitochondrial volume and cell viability. | [74] |
| Achyranthes aspera/Amaranthaceae | Chaff-flower, prickly chaff flower, devil’s horsewhip, burweed | Ethanol/leaf | n.a. | n.a. | Fish treated with this extract returned to normal condition and revealed decrease in superoxide dismutase level and decreased mitochondrial and cell viability. | [74] |
| Achyranthes bidentate/Amaranthaceae | Ox knee, niu xi (Chinese) | Aqueous/root | 0–30.01 µg/mL | No mortality observed | The extract did not exhibit noticeable lethal or severe side effects on zebrafish larvae/embryos. | [75] |
| Allium carinatum/Amaryllidaceae | Keeled garlic, witch’s garlic | 70% aqueous methanol/whole plant | 1–60 µg/mL | 55.8 µg/mL | The extract reduced developmental toxicity and neutropenia. No teratogenic effects were seen. | [76] |
| Allium flavum/Amaryllidaceae | The small yellow onion, yellow-flowered garlic | 70% aqueous methanol/whole plant | 1–60 µg/mL | 50.3 µg/mL | The extract reduced developmental toxicity and neutropenia. No teratogenic effects were seen. | [76] |
| Andrographis paniculata/Acanthaceae | Green chiretta, creat | Aqueous/leaf | 0–10,000 µg/mL | 525.5 µg/mL (48 hpf) 525.6 µg/mL (96 hpf) | Morphological defects were observed at 96 hpf after exposure to teratogen concentration. | [2] |
| Annona squamosa/Annonaceae | Sugar apple, sweetsop | Ethanol/young leaf | 100–1000 µg/mL | n.a. | Late hatching process and morphological deformations occurred at higher concentrations up to 800 µg. | [77] |
| Avicennia marina/Acanthaceae | Grey mangrove or white mangrove | Methanol/bark, leaf, stem, flower and fruit | 50–100 µg/mL | n.a. | n.a. | [78] |
| Azadirachta indica/Meliaceae | Neem | Ethanol (cold)/fruit | 1000–5000 µg/mL | n.a. | Not toxic to adult zebrafish and did not alter the locomotor system. | [79] |
| Bougainvillea galbra/Nyctaginaceae | The lesser bougainvillea, paperflower | Aqueous/bracts | 1–300 µg/mL | 85.51 µg/mL | Yolk sac edema was observed at different concentrations. Hypopigmentation in the embryo was caused by the purple bract extract at 30 µg/mL. Overall, this extract generally showed moderate embryo toxicity. | [80] |
| Bixa Orellana/Bixaceae | Achiote | Hot water/leaf | 50–10,000 µg/mL | n.a. | Plant extract affected the hatchability of embryos and was linked to delayed development. Also, a coagulated embryo, and tail malformation were observed as teratogenic effects. | [81] |
| Capsicum chinense/Solanaceae | Habanero type pepper | Ethanol/fruit | 0.39–100 µg/mL | 39.7 ± 2.1 µg/mL | The embryo demonstrated late development represented by an absence of pigmentation of the tail, and irregular formation of the somites demonstrated the noticeable tail curve with a wide end. | [82] |
| Carthamus tinctorius/Asteraceae | Safflower (English), Kashefeh (Persian) | Aqueous/flower | 40–1000 µg/mL | 345,600 µg/L | Hatching inhibition, depressed heart rate, abnormal spontaneous movement, pericardial edema, yolk sac edema, unusual head-trunk tilt, suppression of melanin release, enlarged yolk, and short body length were identified. | [83] |
| Clinacanthus nutans/Acanthaceae | Sabah snake grass | Hexane/leaf | 15.63–500 µg/mL | 75.49 µg/mL | Morphological disorders (less pigmentation, tail bending, edema, abnormal yolk sac). | [84] |
| Cinnamon zeylanicum/Lauraceae | True cinnamon tree, Ceylon cinnamon | Aqueous/bark | 0–10,000 µg/mL | 985.8 µg/mL (48 hpf) 50.58 µg/mL (96 hpf) | This plant showed minimum embryotoxicity on zebrafish. In 72 hpf embryos, higher concentration of extract provoked yolk sac and pericardial edema. | [2] |
| Croton tiglium/Euphorbiaceae | Purging croton, Jamaal gota (Hindi) | Aqueous/seed | 4000–24,000 µg/mL | 11,880 µg/mL (24 h) 8160 µg/mL (48 h) | Signs of stress, enhanced respiratory rate, jerky movements, loss of equilibrium, circular swimming before losing balance. | [10] |
| Curcuma longa/Zingiberaceae | Turmeric | Methanol/rhizome | 7.8–125 µg/mL | 92.42 µg/mL (24 hpf) 79.20 µg/mL (48 hpf) 68.32 µg/mL (72 hpf) 56.68 µg/mL (96 hpf) 55.90 µg/mL (120 hpf) | Physical anatomy deformities as teratogenic effects; tail bending, yolk sac edema extension, and a curved trunk were observed at high concentration among hatched embryos. | [1] |
| Curcuma longa/Zingiberaceae | Turmeric | Methanol/rhizome | No concentration ranges were mentioned (10 µg/mL used) | n.a. | No visible symptoms of toxicity were observed: no change in heart rate, body impairment, absence or delay in reaction to tactile stimuli, or mortality. | [85] |
| Curcuma xanthorrhiza/Zingiberaceae | Javanese ginger, Temulawak | Ethanol/rhizome | 100–500 µg/mL | 180.52 µg/mL at 48 h 79.55 µg/mL at 96 h | Major malformations of the pericardial edema of embryos at a concentration of 100 µg/mL, yolk sac edema and tail malformation also were observed. | [43] |
| Javanese ginger, Temulawak | Aqueous/rhizome | 0–10,000 µg/mL | 748.6 µg/mL (48 hpf) 703.7 µg/mL (96 hpf) | Decreased survival rate, organ deformity, unusual heartbeat, and delayed hatching rates. | [2] | |
| Cynodon dactylon/Poaceae | Scutch grass | Hexane, chloroform, acetone, methanol/n.a. | 10–100 µg/mL | 32.6 µg/mL | The period of systole and diastole was decreased by the methanol extract. | [86] |
| Derris elliptica/Fabaceae | Tuba root (Indonesia), opay (Philippines) | Aqueous/leaf | 50 (0.05%), 500 (0.5%) µL/mL | n.a. | The concentration of 0.5% led to the early death of embryos: unformed head, unformed tail, coagulation, and death. A delay in growth and restricted movement compared to the control group were present at 0.05%. | [53] |
| Dieffenbachia amoena/Araceae | Spotted dumbcane | Aqueous/leaf | 0–10,000 µg/mL | 1190 µg/mL | Growth retardation, no heartbeat was noticed at 500 µg/mL and higher concentrations (cardiotoxicity). | [87] |
| Diospyros discolor/Ebanaceae | Velvet persimmon, velvet apple, mabolo tree | Aqueous/leaf | 0.05–10% | 1% | Tail malformation, delayed growth, head abnormality, yolk malformations, and abdominal swelling were observed. | [88] |
| Eclipta prostrata/Asteraceae | False daisy, yerba de tago, Karisalankanni and bhringraj | Aqueous/leaf | 0.01–40% | 1% | At the concentration of 0.01% v/v and 1% v/v, did not observe any mortality and abnormality | [89] |
| Enydra fluctuans/Asteraceae | Marsh herb, water cress | Ethanol/leaf | 12,500–400,000 µg/mL | 24 h: 204,132 µg/L 48 h: 170,513 µg/L 72 h: 139,478 µg/L 96 h: 92,956 µg/L | The death rate was found as the exposure period was extended from 24 to 96 h, the median lethal concentration decreased. A negative relationship between exposure time and LC50 was found. | [90] |
| Eugenia polyantha/Myrthaceae | Bay leaf | Aqueous/leaf | 0–10,000 µg/mL | 921.2 µg/mL (48 hpf) 60.39 µg/mL (96 hpf) | Decreased survival rate, organ deformity, unusual heartbeat, and delayed hatching rates. | [2] |
| Euphorbia pekinensis Rupr/Euphorbiaceae | The Peking spurge, daji (Chinese) | Aqueous/commercial raw herb | 100–250 µg/mL | 250–300 µg/mL (previous study) | n.a. | [91] |
| Euodia rutaecarpa (Tetradium ruticarpum)/Rutaceae | Goshuyu (Japanese), Bee tree | Evodiamine (Commercial product) | 0.05–1.6 µg/mL | 0.354 µg/mL (LC10) | Evodiamine (bioactive compound) had a 10% lethality at 354 ng/mL and caused heart defect, changes in heartbeat and blood flow, and pericardial malformation. Also, it could cause cardiovascular side effects involving oxidative stress. | [92] |
| Excoecaria agallocha/Euphorbiaceae | Back mangrove | Methanol/bark, leaf and stem | 50–100 µg/mL | n.a. | Higher concentrations induced intense impacts on embryo melanogenesis. Those concentrations also reduced the eye melanin material of the embryo. | [78] |
| Ficus glomerate/Moraceae | Cluster fig tree, Indian fig tree, goolar fig | Hot aqueous/leaf | 125–2000 µg/mL | 239.88 µg/mL | Lower hatchability, body size, heartbeat rate, and morphological growth defects of the embryo were exhibited. | [73] |
| Garcinia hanburyi/Clusiaceae | Siam gamboge, Hanbury’s Garcinia | Commercial product/dried yellow resin | 0.5–1.0 µM | 1.76 µM | Gambogic acid triggered a lack of fin developmental in the zebrafish embryo. | [93] |
| Garcinia mangostana (Xanthone crude extract)/Clusiaceae | Mangosteen | Mixture of acetone and water (80:20) | 7.81–250 µg/mL | 15.63 µg/mL | Among surviving zebrafish, no deformities were found. | [28] |
| Geissospermum reticulatum/Apocynaceae | Challua caspi (Kichwa language) | Ethanol/bark | 0.1 µg/mL | n.a. | This plant was nontoxic and caused no deformations to zebrafish, even at high concentrations. | [94] |
| Himatanthus drasticus/Apocynaceae | Janaguba milk (latex) | Commercial product | 500–1500 µg/mL | 1188.54 µg/mL | This plant can be considered to have low toxicity. No teratogenic effect was observed. | [95] |
| Hylocereus polyrhizus/Cactaceae | Pitaya | Ethanol; water solution (70:30, v/v)/peel & pulp | 100–1000 µg/mL and 20 µL) | >1000 µg/mL (96 h) | The results indicated the pulp and peel of this plant were non-toxic. | [96] |
| Leonurus japonicus/Lamiaceae | Oriental/Chinese motherwort | Essential oil/the aerial parts | 6.25–100 µg/mL | 1.67 ± 0.23 µg/mL | Motherwort essential oil was toxic to embryos. Morphological abnormalities: partial or complete absence of eye development, yolk sac oedema, curved spine, tail deformities, scattered haemorrhages in the oedematous yolk sac, incomplete heart development, and pericardial oedema. | [97] |
| Ligusticum chuanxiong/Apiaceae | Szechuan lovage | A protein-containing polysaccharide/rhizome | 0–800,000 µg/L | 965 µg/mL | No significant morphological anomalies were found. | [98] |
| Maerua subcordata (Gilg) DeWolf/Capparaceae | Methanol/fruit, leaf, root tuber, and seed | 150,000–1,500,000 µg/L | 209,000 µg/L | ≤5% sub-lethal abnormalities (signs of malformation of the heart). | [4] | |
| Marsdenia tenacissima (Xiaoaiping extract)/Asclepiadaceae | Murva (Hindi), Tong-Guan-Teng (Chinese) | Commercial product | 0–3200 µg/mL | 2660 µg/mL (24 hpf) 2310 µg/mL (48 hpf) 1920 µg/mL (72 hpf) 1910 µg/mL (96 hpf) 1790 µg/mL (120 hpf) | Severe malformation such as impairment of the swim bladder, retaining the yolk, pericardial oedema, and tail curvature. Histopathological study: Xiaoaiping induced lesion of the liver, muscle, and heart. | [99] |
| Millettia pachycarpa/Fabaceae | Bokol-bih, Holsoi, Bokoa-bih (Assamese or Hindi) | Aqueous/root | 1–7.5 µg/mL | 4.276 µg/mL | There were a few developmental anomalies, such as yolk sac oedema, curvature of the spinal cord, pericardial oedema, swim bladder swelling, reduced heart rate, and late hatching. This plant extract is also linked to reactive oxygen species and apoptosis, which induced embryonic mortality and toxicity (identified in trunk, brain, and tail). | [6] |
| Momordica charantia/Cucurbitaceae | Bitter melon, bitter gourd, bitter apple | Methanol/fruit, seed | 1–400 µg/mL | 50 µg/mL (only seed extract) | Although it affected the heartbeat, the fruit extract is considered to be harmless and no mortality was observed. The seed extract demonstrated a slight level of developmental failure and had significant cardiac toxicity. | [100] |
| Hot and cold aqueous/leaf | 15.625–1000 µg/mL | 144.54 µg/mL: Hot aqueous Chinese extract 199.53 µg/mL: Hot aqueous Indian extract 251.19 µg/mL: Cold aqueous Chinese extract | For all samples the heart beat differed at higher concentrations. Also, mild toxicity was observed, although consumption of this plant is assumed to be safe. | [12] | ||
| Aqueous/leaf | 0.05–3% | 3.0% at 24 h | Teratogenic effects such as bent back, tip of tail bent, oedema in the yolk sac, and scoliosis. Delayed development and morphological deformities of the embryo were observed. | [101] | ||
| Moringa oleifera/Moringaceae | Drumstick tree, Moringa | Hot water/leaf and bark | 0.3–6 µg/mL | 1.5 µg/mL at 36 h (leaves); 3 µg/mL at 36 h (bark) | Even low concentration was embryo-toxic and had teratogenic impacts on the developing embryos. | [102] |
| Essential oil/seed | 0.1–1000 µg/mL | 21.24 ± 0.44 µg/mL | High concentrations (≥ 50 µg/mL) caused 100% mortality of embryos. The process of angiogenic blood vessel formation in zebrafish embryos was substantially disturbed by the seed oil. | [103] | ||
| Aqueous/seed | 12.5–200 µg/mL | 190 µg/mL (48 h) 133 µg/mL (72 h) 49 µg/mL (96 h) | No toxic endpoint was seen after 24 h, hatching was prolonged, and the larval period at 96 h compared with the control was decreased. | [104] | ||
| Moringa peregrina/Moringaceae | Miracle tree | Essential oil/seed | 0.1–1000 µg/mL | 25.11 ± 0.547 µg/mL | High concentrations (≥ 50 µg/mL) caused 100% mortality of embryos. The process of angiogenic blood vessel formation in zebrafish embryos was substantially disturbed by the seed oil. | [103] |
| Olea europaea/Oleaceae | Olive | Raw and polar fraction (raw oil: commercial)/fruit | 1–100% | Raw oil: 11.98% (EC50) Polar fraction: 61.87 (EC50) | Abnormalities observed included non-hatching, pericardial oedema, and blood accumulation, and more were expressed at a concentration of 11.98%. Raw oil induced 50% of developmental abnormalities compared with the polar fraction. After 96 h, strong developmental retardations such as developmental delay and absence/prolongation of pigmentation formation were observed. | [105] |
| Onosma dichroantha/Boraginaceae | n.a. | Cyclohexane, ethyl acetate, methanol/root | 0.02–50 µg/mL | n.a. | n.a. | [106] |
| Orthosiphon stamineus/Lamiaceae | Java tea | Aqueous/whole plant | 0–10,000 µg/mL | 1685 µg/mL (48 hpf) 1685 µg/mL (96 hpf) | n.a. | [2] |
| Palicourea deflexa/Rubiaceae | n.a. | Methanol (fraction)/leaf | 1–100 µg/mL | 72.18 µg/mL | At high concentration (100 µg/mL), some possible anomalies, such as no development of somites, decreased pigmentation, and formation of oedema were observed. Lethality only appeared after 96 h. | [52] |
| Palmaria palmata/Archaeplastida | Dulse, dillisk (dilsk), red dulse, sea lettuce flakes (creathnach) | Crude protein (hydrolysate)/leaf | 1–10,000 µg/mL | n.a. | At high concentration (10 mg/mL), larvae started developing spinal curvature and a deformed yolk sac. At 5 mg/mL, larvae appeared to have a swollen yolk sac and developed a curved spine. | [51] |
| Passiflora caerulea/Passifloraceae | Bluecrown Passionflower | Aqueous/leaf | 40–120 µg/mL | 80 µg/mL (40–120 µg/mL) | Mortality increased based on concentration and delay in hatching. No apparent signs of abnormal growth or morphology were detected in embryos at 96 hpf. | [107] |
| Peucedanum alsaticum/Apiaceae | n.a. | n-heptane, ethyl acetate, methanol, water/fruit | 50–400 µg/mL | n.a. | n.a. | [33] |
| Phyllanthus niruri/Phyllanthaceae | Gale of the wind, stonebreaker | Aqueous/leaf | 0.05–10% | – | Tail malformation and coagulation were detected as the most remarkable toxic effect from the extract; dose-dependent impacts on the heartbeat and hatchability of the embryo also were detected. | [108] |
| Piper betle/Piperaceae | Betel, Ikmo, Daun sirih (Malay), Paan (Hindi) | Hot water/leaf | 50–10,000 µg/mL | – | The plant extract affected the hatchability of embryos and was linked to delayed development. Also, a coagulated embryo and tail malformation were observed as a teratogenic effect. | [81] |
| Polygonum multiflorum/Polygonaceae | Tuber fleeceflower | n-hexane, ethyl acetate, water/root | 0–175,000 µg/L | TIs: 1430 (ethyl acetate), 630 µg/L | 40 and 105 mg/L concentrations (all extracts) induced yolk sac oedema (heart oedema), hemovascular defects, abnormal trunk, and necrosis. The water extract induced synthesis of melanin in zebrafish through stimulation of tyrosinase. | [109] |
| Water, ethanol, methanol, acetone/dried roots | 0–10,000 µg/mL | 129.4 ± 2.7 µg/mL (water) 58.2 ± 1.9 µg/mL (30% ethanol) 39.8 ± 0.8 µg/mL (50% ethanol) 27.9 ± 2.3 µg/mL (70% ethanol) 25.5 ± 2.0 µg/mL (95% ethanol) 48.8 ± 2.8 µg/mL (methanol) 23.6 ± 2.2 µg/mL (acetone) | Teratogenic effects included coagulated embryos, lack of development of the somite, absence of tail detachment and heartbeat, and malformation of the notochord | [110] | ||
| Psoralea corylifolia/Fabaceae | Babchi | Commercial product (Psoralen) | 0–6.70 µg/mL | 3.40 µg/mL (LC50) 2.52 µg/mL (LC10) 1.98 µg/mL (LC1) | Psoralen therapy caused hatching rate and body size to decrease and the abnormality rate of zebrafish to increase significantly. Yolk retention, pericardial oedema, swim-bladder malformation, and curved body form were observed. | [111] |
| Punica granatum/Lythraceae | pomegranate | Ethanol/peel | 100–250 µg/mL | 196,037 ± 9.2 µg/mL | No issues with the reproductive organs, heart, and androgen hormones were detected. | [112] |
| Rhizosphora apiculata/Rhizosphoraceae | Mangrove | Methanol/bark, leaf, stem and root | 50–100 µg/mL | n.a. | Higher concentrations of the extract from bark, leaf, and stem did not cause mortality. | [78] |
| Salvia miltiorrhiza/Lamiaceae | Red sage | Commercial product/Tanshinone IIA (diterpene quinone)/root | 0.44–7.06 µg/mL | 5.44 µg/mL (72 hpf) 3.77 µg/mL (96 hpf) | Tan-IIA showed potential cardiotoxicity and growth inhibition in zebrafish embryos. Scoliosis, tail deformity, and pericardium oedema were the primary signs of teratogenicity. | [45] |
| Sida acuta/Malvaceae | Wireweed | Hexane, chloroform, acetone, methanol/n.a. | 10–100 µg/mL | 20.9 µg/mL | The methanolic extract led to heartbeat rate reductions (more significant than nebivolol as a positive control). This study reported no teratogenic effects. | [86] |
| Solidago canadensis/Asteraceae | Canada goldenrod, Canadian goldenrod | Ethanol/leaf | 0–500µg/mL | 0.42 ± 0.03 µg/mL (24 h) 0.33 ± 0.04 µg/mL (48 h) 0.32 ± 0.03 µg/mL (72 h) | This paper only mentioned that low toxicity on zebrafish was observed. | [113] |
| Sonneratia alba/Lythraceae | Mangrove tree | Methanol/bark and leaf | 50–100 µg/mL | – | – | [78] |
| Spilanthes acmella/Asteraceae | Paracress, toothache plant, Sichuan buttons, buzz buttons, tingflowers, electric daisy Kradhuawean (Thai) | Aqueous/leaf | 0.01–20% | n.a. | No mortality was observed in the highest concentration test. | [89] |
| Spondias mombin/Anacardiaceae | Yellow mombin, hog plum | Hydroethanolic/leaf | 1–9 g/kg (LD50); 25,000–75,000 µg/L (LC50) | 4.515 g/kg (LD50: 48 h: immersion) 49.86 µg/mL (LC50: 48 h: oral) | There were no recorded teratogenic effects. | [114] |
| Streblus asper/Moraceae | Siamese rough bush, khoi, serut, thoothbrush tree | Methanol/bark | 50–100 µg/mL | 2,000,000 µg/mL | Slight oedema of heart muscles at 100 µg/mL. No other severe malformations were observed. | [115] |
| Sutherlandia frutescens/Fabaceae | Cancer bush, balloon pea, sutherlandia | Aqueous and 80% ethanol | 5–50 µg/mL | 30.00 µg/mL | Both extracts showed bleeding and pericardial cyst development at high concentration, but the aqueous extract was less toxic to larvae. | [8] |
| Aqueous, ethanol/aerial part | 5–300 µg/mL | 297.57 µg/mL (aqueous) 40.54 µg/mL (ethanol) | The high concentration of both extracts inhibited hatching rate and mortality. | [116] | ||
| Terminalia chebula/Combretaceae | Myrobalan | Ethanol/fruit, leaf, root | n.a. | n.a. | At 50 µg/L, fish treated with this extract came back to normal condition and showed a related decrease in superoxide dismutase and decreased mitochondria and cell viability. | [74] |
| Tetrapterys (Melanolepis) multiglandulosa/Malpighiaceae | Pakalkal (Tagalog) | Dichloromethane, hexane, ethyl acetate, methanol and water/leaf | n.a. | 200 µg/mL (methanol extract) | A significant ecdysteroid was the most toxic in the zebrafish. | [117] |
| Tinospora cordifolia/Menispermaceae | Heart-leaved moonseed, gaduchi, guduchi, giloy, Makabuhay (Filipino) | Aqueous/leaf and bark | 0.01–10% | 1% (leaves), 10% (barks) | Abnormalities of head and tail, delayed development, reduced mobility, stunted tail, and scoliosis/flexure were observed. | [118] |
| Thuja orientalis/Cupressacae | Oriental Arbor-vitae | Ethanol/leaf | 150–2400 µg/mL | 702.9 µg/mL | At 2.4 mg/mL embryos were coagulated; at 1.2 mg/mL they did not hatch and oedema was observed; 0.6 mg/mL resulted in skeletal deformities; at 0.3 mg/mL slight oedema was observed; at 0.15 mg/mL normal growth was observed. | [50] |
| Trapa natans/Lythraceae | Water chestnut, water caltrop | Acetone, methanol and ethyl acetate/leaf | 100,000–1,000,000 µg/mL | 50 µg/mL (methanol) 40 µg/mL (acetone) 30 µg/mL (ethyl acetate) | Methanol and acetone extracts did not show toxicity to zebrafish. | [119] |
| Tripterygium wilfordii/Celasteraceae | Thunder god vine, léi gōng téng (Mandarin) | Purchased commercially each compound | 1/10, 1/5, 1/2 LC50 | Taxol: 0.10, 0.21, 0.53 µg/mL; Gambogic acid 0.03, 0.05, 0.13 µg/mL; Triptolide: 0.04, 0.07, 0.18; Auranofin: 0.84, 1.68, 4.21 µg/mL; Mycophenolic acid: 0.44, 0.95, 2.22; Curcumin: 0.43, 0.87, 2.17 µg/mL; Thalidomide: 0.71, 1.43, 3.56 µg/mL | From those 7 compounds, gambogic acid followed by taxol showed high anticancer activity | [120] |
| Zanthoxylum sp./Rutaceae | n.a. | Commercial product; zanthoxylum | 0–1000 µg/mL | 81.18 µg/mL | No teratogenic effects were reported. | [121] |
Days post-fertilization: dpf; Hours post-fertilization: hpf; n.a.: not applicable.
Table 4.
Description of toxicological aspects of various studies.
| Exposure Format | Number | Exposure Stage | Exposure Time | Parameters Evaluated (Endpoint) | Compound(s) | Reference |
|---|---|---|---|---|---|---|
| 96-well plates | 1/well | 24 hpf | 96 h | Embryotoxicity | Armatamide, rubecenamide, lemairamin, rubemamine, and zanthosine | [121] |
| 1/well (10/group) | 6 hpf | 96 h | Embryotoxicity and teratogenic effects | Palmitic acid, phytol, hexadecanoic acid, 1-monopalmitin, stigmast-5-ene, pentadecanoic acid, heptadecanoic acid, 1-linolenoylglycerol, and stigmasterol | [84] | |
| 30/group (90 embryos) | n.a. | 96 h | Teratogenic effects and embryotoxicity | Alkaloids, flavonoids, phenolics, and saponins | [73] | |
| 1/well | 24 hpf | 72 h | Acute toxicity assay | Polysaccharides, peptides, protein, lipids, terpenoids, saponins, phenolics, and sterols | [12] | |
| 12/group | 2 hpf | 96 h | Embryotoxicity and teratogenic effects | α/β-thujone, thujone | [50] | |
| n.a. | 1 hpf (embryo) | 72 h | Cytotoxicity and embryonic toxicity test (survival rate) | Xanthone | [28] | |
| 4/group (per well) | 12 hpf | 48 h | Embryotoxicity (mortality, hatchability, heartbeat rate, and malformation), teratogenic activity | Ascorbic acid, flavonoids, phenolics, and carotenoids | [102] | |
| 1/well | 24 hpf | 96 h | Embryotoxicity and teratogenic effects | Capsaicin, ethyl palmitate, ethyl linoleate, dihydrocapsaicin, and docosenamide | [84] | |
| 36/group | 2.5–3.7 hpf | 72 h | Developmental toxicity and behavioural safety | Roots: glycoside, siaraside (pregnane glycoside); Stem bark: α-amyrin, acetate, lupeol, and β-sitosterol | [115] | |
| 12/group | 4 cell embryo stage | 144 h | Embryotoxicity (heart rate and hatching time) | Spilanthal, and flavonoid | [89] | |
| 24-well plates | 12 fertilized eggs/group | 6 hpf | 120 h | Embryotoxicity and teratogenic effects (hatching rate, heartbeat, otolith, blood circulation, tail detachment, and motility) | Catechin, epicatechin, and naringenin | [1] |
| 20/concentration | 6 hpf | 96 h | Embryotoxicity | [76] | ||
| 10/well | 6 hpf | 96 h | Embryotoxicity | Methanol extract: phenolic compound (p-heydroxybenzoic acid), phenolic acid, flavonoid, and flavonoid glycoside | [119] | |
| 10/group (30 per tested sample) | 64-cell (egg) | 96 h | Embryotoxicity | Phenolic compounds: tyrosol, catechol. Hydroxytyrosol, gallic acid, and ascorbic acid. | [105] | |
| 1/well | 4 hpf | 96 h | Embryotoxicity | Pyrrolizidine alkaloids such as senecionine or senecivernine; phenolic compounds, terpenoids, and iridoids, glucosinolates (glucolepidin and glucobrassicin), alkaloids or amines (stachydrine and trigonelline) | [4] | |
| 4/group | 12 hpf | 48 h | Embryotoxicity, teratogenic effects | n.a. | [108] | |
| 30/group | 4 hpf | 96 h | Embryotoxicity | Phenols, and polyphenols | [107] | |
| 10/well | 16 cell stage | 72 h | Embryotoxicity | n.a. | [82] | |
| 6/well | 24 hpf | 96 h | Embryotoxicity | Sutherlandin, and sutherlandioside | [116] | |
| 6/group (per well) (n =18) | 24 hpf | 216 h | Developmental toxicity and cardiotoxicity | No pure compound isolated | [8] | |
| 15/well | 2 hpf | 96 h | Embryotoxicity, teratogenic effects | Alkaloids (stachydrine) | [97] | |
| 4/well | – | 48 h | Embryotoxicity, teratogenic effects | n.a. | [88] | |
| n.a. | 6 hpf | 96 h | Acute toxicity in vivo (zebrafish), in silico (computational chemistry) | Brevifolin, tannin, saponin, quinone, steroid/triterpenoid, and flavonoid | [112] | |
| 15/well | 4 hpf | 96 h | Embryotoxicity, teratogenic effects | Psoralen | [111] | |
| 40/group | 16-cell stage | 96 h | Zebrafish embryo acute toxicity (ZFET) | Emodin, rhein, physcion (dihydroxyanthraquinone), chrysophanol, aloe-emodin, rhaponticin, polygonumnolide, and 2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucopyranosede, and resveratrol-2-O-β-D-glucopyranoside | [110] | |
| 30/group (10/well) | 9 hpf | 55 h | Anti-melanogenic and embryotoxicity | Lupeol (triterpenoid), hydrolysable, and 3,3′-di-O-methyl ellagic acid (bark of S. alba) | [78] | |
| 1/well (20 embryos per concentration) | 16-cell stage | 168 h | Embryotoxicity | n.a. | [104] | |
| 20 eggs/group (1 embryo per well | 1 hpf (embryo) | 96 h | Embryotoxicity, teratogenic effects (body axis, head, tail, blood circulation, eyes, heart, pigmentation, somite, and yolk sac) | Curcumin, desmethoxycurcumin, and bisdemethoxycurcumin | [43] | |
| 1/well | 32-cell stage | 96 h | Embryotoxicity | harman-3-carboxylic acid, βc alkaloids: β-carboline alkaloid | [52] | |
| 30 embryos/well | 2 hpf | 96 h | Toxicity on cell and zebrafish (coagulation of eggs, tail detachment, presence of heartbeat, and hatching rate) | Alkanes, alkenes, amino acid, amines, and amides | [2] | |
| 20/well | 6 hpf | 72 h | – | 3-O-propionyl-5, 10, 14-O-triacetyl-8-O-(20 -methyl- butanoyl)-cyclomyrsinol, and mirsinance type diterpene | [91] | |
| 20/well | 2 hpf | 70 | Embryotoxicity, teratogenic effects | Gambogic acid | [93] | |
| 5/group (per well) | 6 hpf | 120 h | Developmental toxicity and apoptosis induction | Rotenone, and saponin | [6] | |
| 10/well | 24 hpf | 96 h | Embryotoxicity | n.a. | [86] | |
| 6/well | 7 dpf | 24 h | Determination of maximum tolerated concentration (MTC) | ar-turmerone, α, β-turmerone, and α-atlantone | [85] | |
| 12-well plates | 4/group (per vial) | 1 hpf (embryo) | 48 h | Embryotoxicity, teratogenic effects | Beta-momorcharin | [101] |
| 20/group | 6 hpf | 96 h | Embryotoxicity, teratogenic effects | Quinochalcones, and flavonoids | [83] | |
| 4/well | – | 48 h | Embryotoxicity, teratogenic effects | Flavonoids, terpenoids, cardiac glycosides, saponins, and tannins | [81] | |
| 4/group (per well) | 12 hpf | 48 h | Embryotoxicity, teratogenic effects | Alkaloids, di-terpenoid lactones, glycosides, steroids, sesquiterpenoid, phenolics, and aliphatic | [118] | |
| 3/well | n.a. | 48 h | Embryotoxicity and teratogenicity | Rotenone | [53] | |
| 10/well | 48 hpf | 48 h | Embryotoxicity | Peptides | [51] | |
| 12/group | egg | 120 h | Developmental toxicity (mortality, hatching rate, oedema, and malformations) | Alkaloids, flavonoids, terpene derivatives, glycosides, novel diazepine, and squalene | [77] | |
| Glass beaker | 24/group | n.a. | 96 h | Acute toxicity test | Phyllocactins, betanins, 2′-O-apiosyl-malonyl-betanin, 6′-O-malonyl-2-descarboxybetanin, flavonoids, isorhamnetin triglycosides, quercetin-3-O-hexoside, and carbohydrates. | [96] |
| 15/group | 0 hpf | 48 h | Acute toxicity test, histopathology | Fatty acids, lupeol, and terpenes | [114] | |
| Glass Petri dish | 6/group | adult | 96 h | Fish acute toxicity | Alkaloids, phenol, and flavonoids with some chemical groups of alkanes, aromatics, hydroxyls, carbonyls, aldehyde, amines, nitriles, amides and ethers | [78] |
| 30/Petri dish | 6 hpf | 72 h | Embryotoxicity | Moringa oleifera seed oil: 2,2-Bipydidine,3,3-diol, myristoleic acid, palmitoleic acid, stearic acid, oleic acid, arachidic acid, methyl-9-octadecanoate, adrenic acid, erucic acid, behenic acid, decamethylcyclopentasiloxane, α-tocopherol, γ-tocopherol, β-sitosterol, camphenone, hexahydrofarnesylacetone, linoleic acid, gadoleic/eicosenoic acid Moringa peregrina seed oil: palmitic acid, stearic acid, oleic acid, arachidic acid, gadoleic/eicosenoic acid, tricosane, erucic acid, behenic acid, α-tocopherol or γ-tocopherol, α-sitosterol, δ-tocopherol, 2-allyl-5-t-butylhydroquinone, isamoxole, hexahydrofarnesylacetone, linoleic acid, and β-sitosterol | [103] | |
| n.a. | 8-cell stage | 5 dpf | Embryotoxicity, teratogenic effects | 1,2-cyclopentanedione, 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, 1,3;2,5-dimethylene-l-rhamnitol, elemol, (-)-selina-4.alpha.,11-diol, and beta-eudesmol | [100] | |
| 20/Petri dish | 1–4 cell stage | 72 h | Embryotoxicity | Shikonin, β,β-dimethylacrylalkannin, and β,β-dimethylacryl shikonin | [106] | |
| Rectangular glass tank | 12/group | n.a. | 48 h | Acute toxicity | Fatty acids, esters, alcohols, and the sterol sitosterol | [114] |
| REKO glass (artificial egg environment) | 20 fertilized eggs | 4 hpf | 48 h | Embryotoxicity, teratogenic effects | Phenolic compounds | [94] |
| 10/glass tank | n.a. | 48 h | Evaluation of genotoxicity and acute toxicity | Phenolic compounds, alkaloids, saponins, terpenoids, carbohydrates, and tannins | [10] | |
| Vial | 4/group (per well) | n.a. | 48–72 h | Cytotoxicity, toxicity, teratogenicity (hatchability and heartbeat rate) | n.a. | [87] |
| Vessel | 10/group | adult | 72 h | Fish acute toxicity | n.a. | [113] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Modarresi Chahardehi, A.; Arsad, H.; Lim, V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants 2020, 9, 1345. https://doi.org/10.3390/plants9101345
AMA Style
Modarresi Chahardehi A, Arsad H, Lim V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants. 2020; 9(10):1345. https://doi.org/10.3390/plants9101345
Chicago/Turabian StyleModarresi Chahardehi, Amir, Hasni Arsad, and Vuanghao Lim. 2020. "Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants" Plants 9, no. 10: 1345. https://doi.org/10.3390/plants9101345
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.