Genome-Wide Identification, Classification, Characterization, and Expression Analysis of the Wall-Associated Kinase Family during Fruit Development and under Wound Stress in Tomato (Solanum lycopersicum L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Wounding Treatment
2.2. SlWAK-RLKs Identification
2.3. SlWAK-RLKs Sequence Analysis
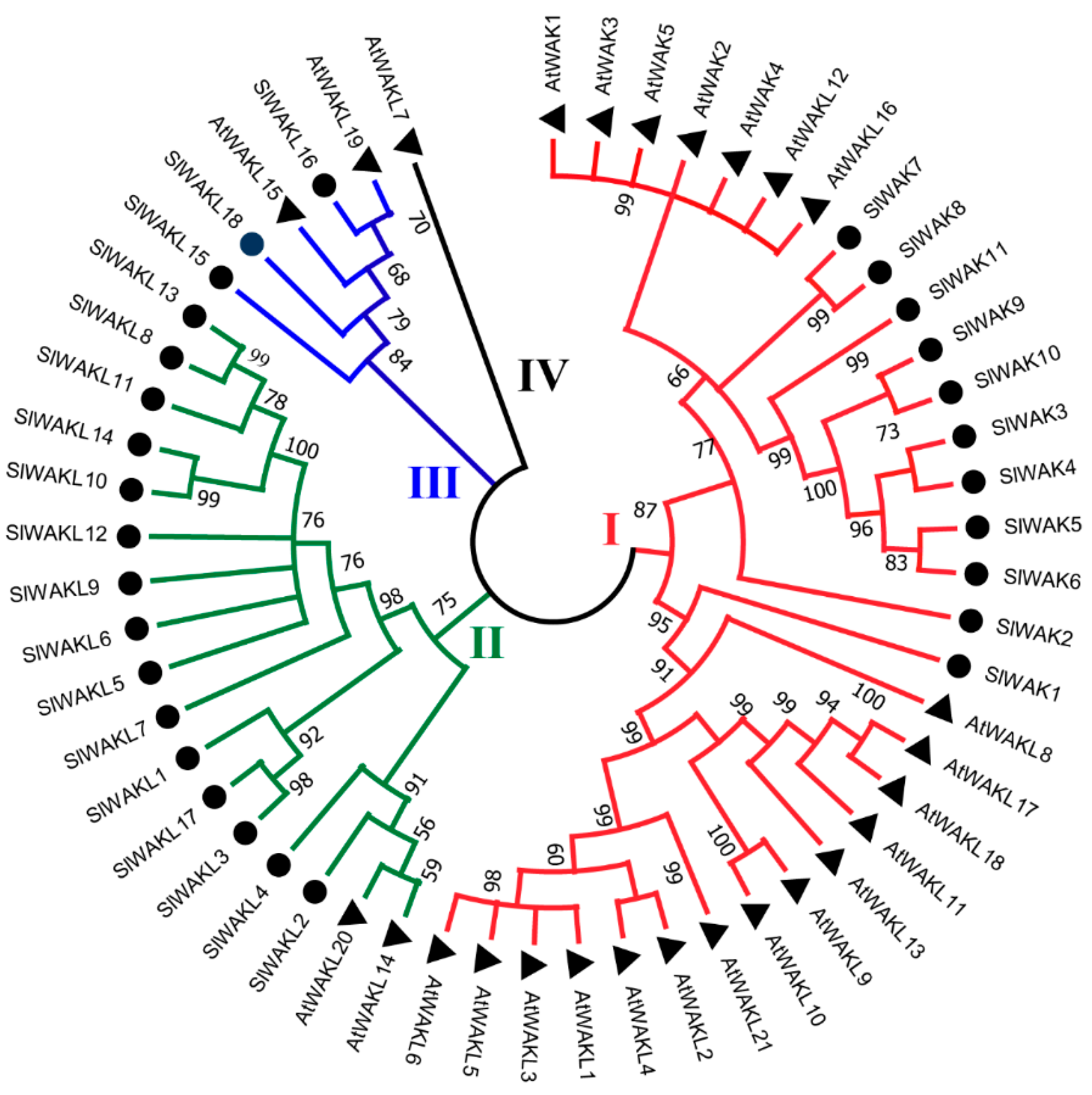
2.4. Phylogenetic Analysis
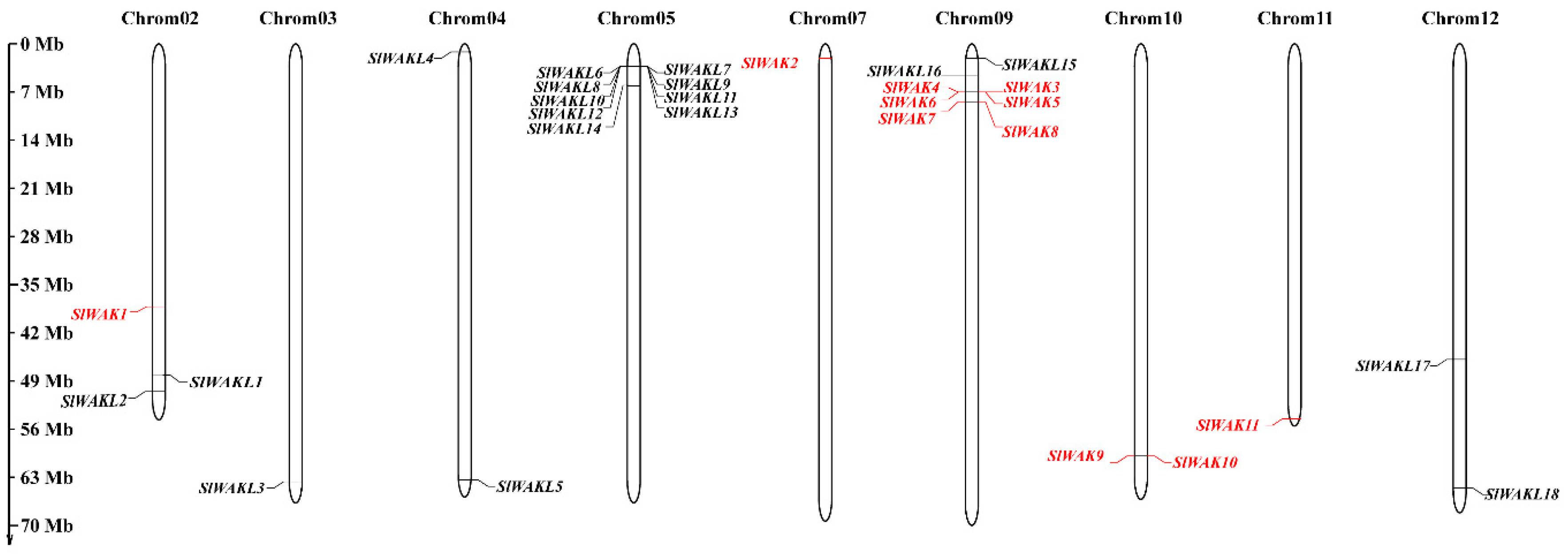
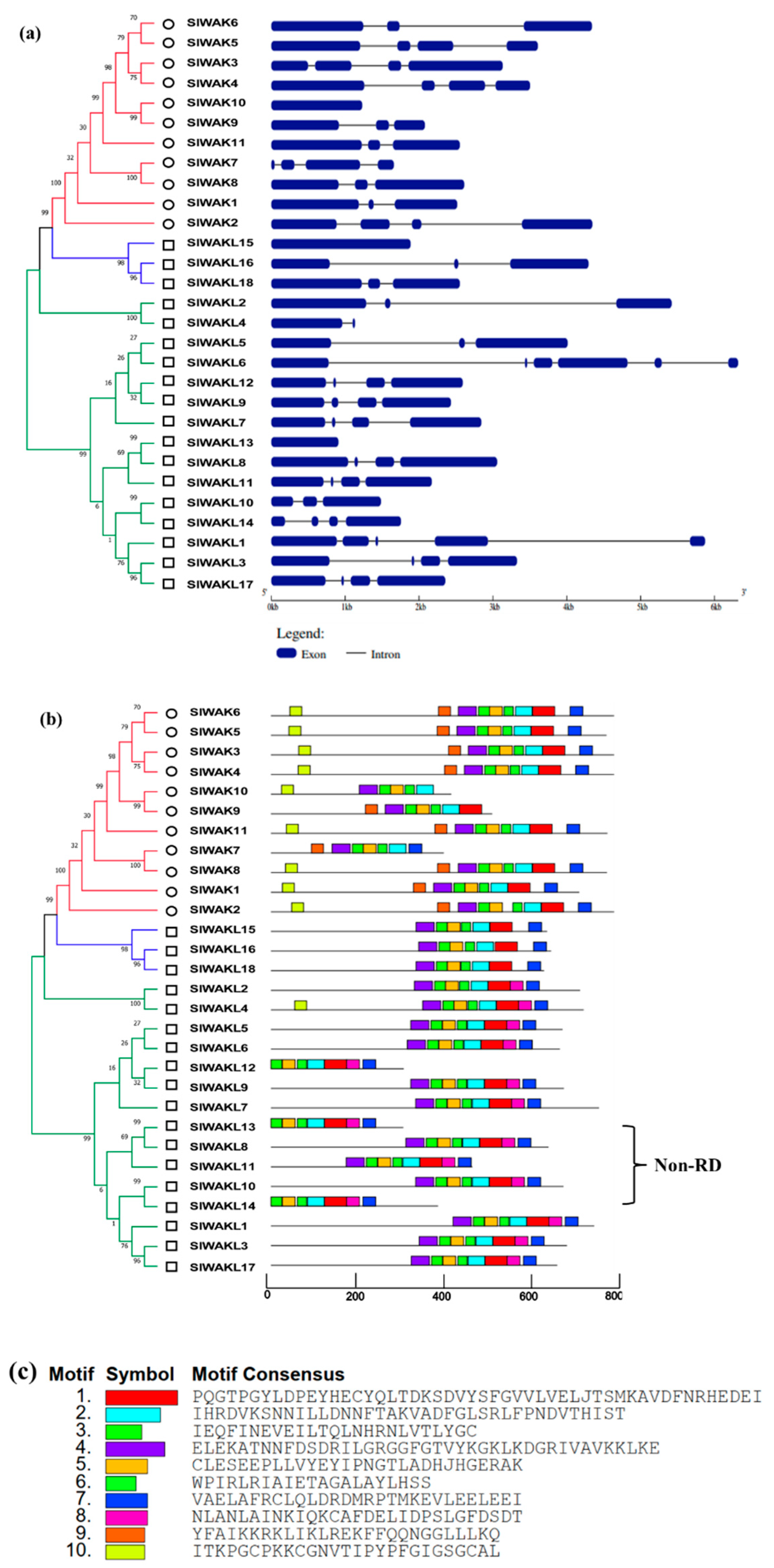
2.5. Gene Structure, Conserved Motif and Chromosome Location Analysis
2.6. Cis-Element Prediction for SlWAK-RLK Gene Promoter
2.7. Heat Map Analysis
2.8. RNA Isolation and qRT-PCR Analysis
2.9. RNA Sequencing and Bioinformatics Analysis
3. Results
3.1. Genome-Wide Identification of SlWAK-RLK Genes in Tomato
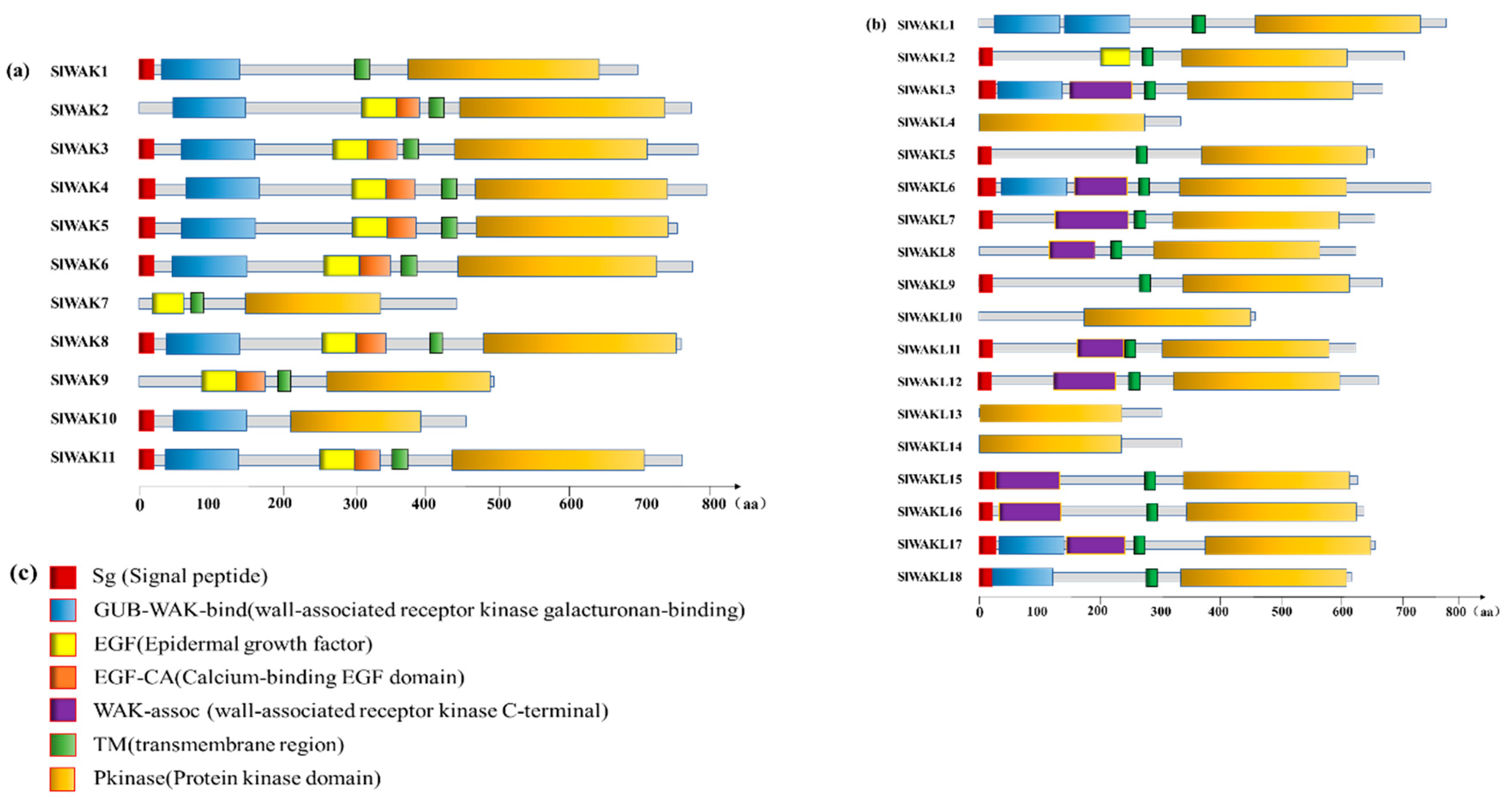
3.2. Conserved Motif and Evolutionary Analysis of SlWAK-RLK Proteins
3.3. Chromosome Location, Gene Structure, and Motif Patterns of SlWAK-RLKs
3.4. Putative Cis-Regulatory Elements in the Promoter Regions of SlWAK-RLKs
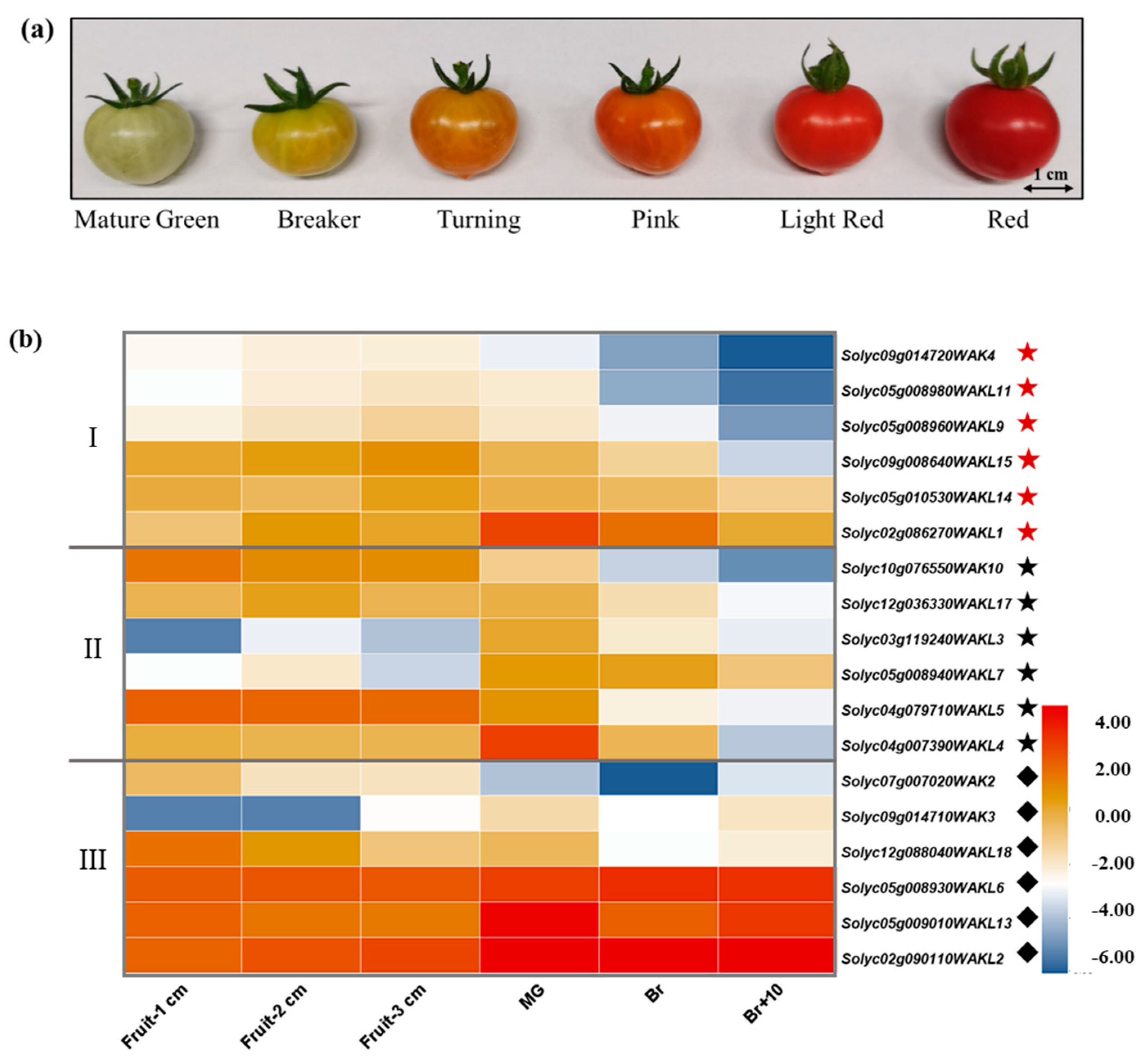
3.5. Expression Patterns of SlWAK-RLKs during Tomato Fruit Development and Ripening
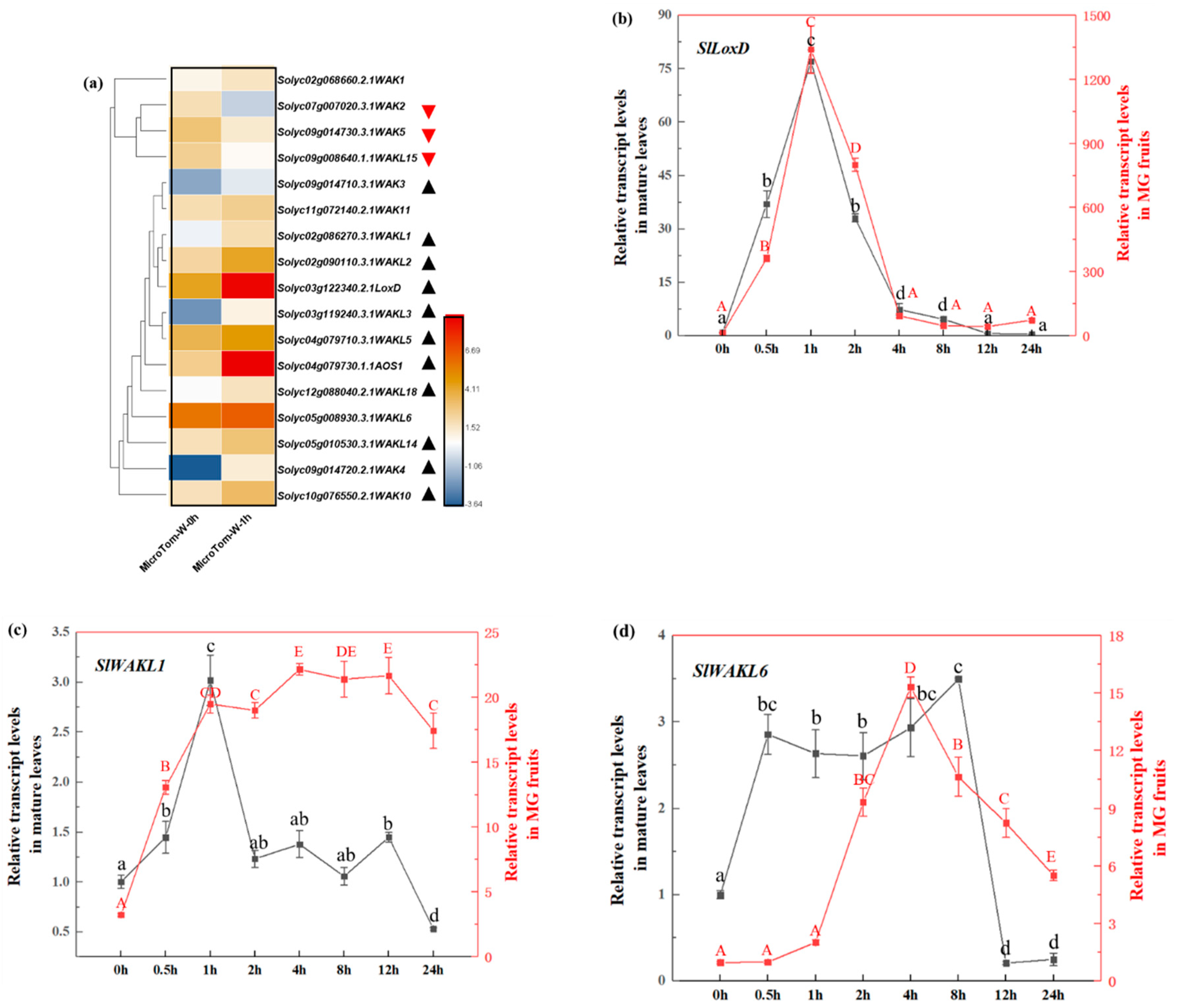
3.6. Screen and Identified Mechanical Wound Induced SlWAK-RLK Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hasanuzzaman, M.; Hossain, M.A.; Silva, J.A.T.D.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar]
- Shiu, S.H.; Bleecker, A.B. Expansion of the receptor-like kinase/Pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol. 2003, 132, 530–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.H. Plant receptor-like kinase gene family: Diversity, function, and signaling. Sci. STKE 2001, 2001, re22. [Google Scholar] [CrossRef] [PubMed]
- Uzman, A. Molecular Cell Biology (4th edition): Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David Baltimore and James Darnell; Freeman & Co., New York, NY, 2000, 1084 pp., list price $102.25, ISBN 0-7167-3136-3. Biochem. Mol. Biol. Educ. 2001, 29, 126–128. [Google Scholar]
- Lehti-Shiu, M.D.; Zou, C.; Shiu, S.-H. Origin, Diversity, Expansion History, and Functional Evolution of the Plant Receptor-Like Kinase/Pelle Family; ResearchGate: Berlin/Heidelberg, Germany, 2012; pp. 1–22. [Google Scholar]
- Anderson, C.M.; Wagner, T.A.; Perret, M.; He, Z.H.; He, D.; Kohorn, B.D. WAKs: Cell wall-associated kinases linking the cytoplasm to the extracellular matrix. Plant Mol. Biol. 2001, 47, 197–206. [Google Scholar] [CrossRef]
- Kohorn, B.D. WAKs: Cell wall associated kinases. Curr. Opin. Cell Biol. 2001, 13, 529–533. [Google Scholar] [CrossRef]
- Stahl, Y.; Simon, R. Receptor-Like Kinases in Plants; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Lally, D.; Ingmire, P.; Tong, H.Y.; He, Z.H. Antisense expression of a cell wall-associated protein kinase, WAK4, inhibits cell elongation and alters morphology. Plant Cell 2001, 13, 1317–1331. [Google Scholar]
- Wagner, T.A.; Kohorn, B.D. Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell 2001, 13, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Kohorn, B.D.; Kobayashi, M.; Johansen, S.; Riese, J.; Huang, L.F.; Koch, K.; Fu, S.; Dotson, A.; Byers, N. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 2006, 46, 307–316. [Google Scholar] [CrossRef]
- He, Z.H.; He, D.Z.; Kohorn, B.D. Requirement for the induced expression of a cell wall associated receptor kinase for survival during the pathogen response. Plant J. 1998, 14, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Sivaguru, M.; Ezaki, B.; He, Z.H.; Tong, H.Y.; Osawa, H.; Baluska, F.; Volkmann, D.; Matsumoto, H. Aluminum-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003, 132, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Zuo, W.; Chao, Q.; Zhang, N.; Ye, J.; Tan, G.; Li, B.; Xing, Y.; Zhang, B.; Liu, H.; Fengler, K.A.; et al. A maize wall-associated kinase confers quantitative resistance to head smut. Nat. Genet. 2015, 47, 151–157. [Google Scholar] [CrossRef]
- Hurni, S.; Scheuermann, D.; Krattinger, S.G.; Kessel, B.; Wicker, T.; Herren, G.; Fitze, M.N.; Breen, J.; Presterl, T.; Ouzunova, M.; et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. USA 2015, 112, 8780–8785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Praz, C.; Li, B.; Singla, J.; Robert, C.A.M.; Kessel, B.; Scheuermann, D.; Luethi, L.; Ouzunova, M.; Erb, M.; et al. Fungal resistance mediated by maize wall-associated kinase ZmWAK-RLK1 correlates with reduced benzoxazinoid content. New Phytol. 2019, 221, 976–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhou, S.-Y.; Zhao, W.-S.; Su, S.-C.; Peng, Y.-L. A novel wall-associated receptor-like protein kinase gene, OsWAK1, plays important roles in rice blast disease resistance. Plant Mol. Biol. 2009, 69, 337–346. [Google Scholar] [CrossRef]
- Hu, W.; Lv, Y.; Lei, W.; Li, X.; Chen, Y.; Zheng, L.; Xia, Y.; Shen, Z. Cloning and characterization of the Oryza sativa wall-associated kinase gene OsWAK11 and its transcriptional response to abiotic stresses. Plant Soil 2014, 384, 335–346. [Google Scholar] [CrossRef]
- Zhang, N.; Pombo, M.A.; Rosli, H.G.; Martin, G.B. Tomato wall-associated kinase SlWak1 depends on Fls2/Fls3 to promote apoplastic immune responses to Pseudomonas syringae. Plant Physiol. 2020, 183, 1869–1882. [Google Scholar] [CrossRef]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar]
- Zuo, C.; Liu, Y.; Guo, Z.; Mao, J.; Chu, M.; Chen, B. Genome-wide annotation and expression responses to biotic stresses of the WALL-ASSOCIATED KINASE-RECEPTOR-LIKE KINASE (WAK-RLK) gene family in Apple (Malus domestica). Eur. J. Plant Pathol. 2019, 153, 771–785. [Google Scholar] [CrossRef]
- Valter de Oliveira, L.F.; Christoff, A.P.; de Lima, J.C.; Feijo de Ross, B.C.; Sachetto-Martins, G.; Margis-Pinheiro, M.; Margis, R. The Wall-associated Kinase gene family in rice genomes. Plant Sci. 2014, 229, 181–192. [Google Scholar] [CrossRef]
- Yan, L.; Zhai, Q.; Wei, J.; Li, S.; Wang, B.; Huang, T.; Du, M.; Sun, J.; Kang, L.; Li, C.-B.; et al. Role of Tomato Lipoxygenase D in Wound-Induced Jasmonate Biosynthesis and Plant Immunity to Insect Herbivores. PLoS Genet. 2013, 9, e1003964. [Google Scholar] [CrossRef] [Green Version]
- Fragkostefanakis, S.; Dandachi, F.; Kalaitzis, P. Expression of arabinogalactan proteins during tomato fruit ripening and in response to mechanical wounding, hypoxia and anoxia. Plant Physiol. Biochem. 2012, 52, 112–118. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lehti-Shiu, M.D.; Shiu, S.-H. Diversity, classification and function of the plant protein kinase superfamily. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2619–2639. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Wang, H.; Li, Y.; Zhu, B.; Zang, Y.; He, Y.; Cao, J.; Zhu, Z.; Yu, Y. Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum. Int. J. Mol. Sci. 2018, 19, 2290. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soeding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Verica, J.A.; He, Z.H. The cell wall-associated kinase (WAK) and WAK-like kinase gene family. Plant Physiol. 2002, 129, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.K.; Mattoo, A.K. Genome-Wide Identification of Tomato (Solanum lycopersicum L.) Lipoxygenases Coupled with Expression Profiles During Plant Development and in Response to Methyl-Jasmonate and Wounding. J. Plant Physiol. 2018, 231, 318–328. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, J.; Zhou, X.; Luan, Y.; Luan, F. Genome-wide identification, characterization and expression analysis of the TLP gene family in melon (Cucumis melo L.). Genomics 2020, 112, 2499–2509. [Google Scholar] [CrossRef]
- Chao, J.; Kong, Y.; Wang, Q.; Sun, Y.; Gong, D.; Lv, J.; Liu, G. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Yichuan 2015, 37, 91–97. [Google Scholar]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Anders, S. Analysing RNA-Seq Data with the DESeq Package; European Molecular Biology Laboratory: Heidelberg, Germany, 2011; pp. 1–22. [Google Scholar]
- Verma, A.; Warner, S.L.; Vankayalapati, H.; Bearss, D.J.; Sharma, S. Targeting Axl and Mer Kinases in Cancer. Mol. Cancer Ther. 2011, 10, 1763–1773. [Google Scholar] [CrossRef] [Green Version]
- Dardick, C.; Schwessinger, B.; Ronald, P. Non-arginine-aspartate (non-RD) kinases are associated with innate immune receptors that recognize conserved microbial signatures. Curr. Opin. Plant Biol. 2012, 15, 358–366. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Yang, L.; Liu, N.; Yang, J.; Zhou, X.-K.; Xia, Y.-C.; He, Y.; He, Y.-Q.; Gong, H.-J.; Ma, D.-F.; et al. Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber. BMC Plant Biol. 2019, 19, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lahiri, S.; Orr, D. Biological Control in Tomato Production Systems: Theory and Practice; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Cheng, X.; Li, M.; Li, D.; Zhang, J.; Jin, Q.; Sheng, L.; Cai, Y.; Lin, Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2017, 6, 1602–1613. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, V.; Gupta, A.K. Wall associated kinases from plants—An overview. Physiol. Mol. Biol. Plants 2008, 14, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Kohorn, B.D. Cell wall-associated kinases and pectin perception. J. Exp. Bot. 2016, 67, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, M.; Deng, L.; Shen, J.; Fang, M.; Chen, Q.; Lu, Y.; Wang, Q.; Li, C.; Zhai, Q. MYC2 Regulates the Termination of Jasmonate Signaling via an Autoregulatory Negative Feedback Loop. Plant Cell 2019, 31, 106–127. [Google Scholar] [CrossRef] [Green Version]
- Howe, G.A.; Ryan, C.A. Suppressors of systemin signaling identify genes in the tomato wound response pathway. Genetics 1999, 153, 1411–1421. [Google Scholar]
- Verica, J.A.; Chae, L.; Tong, H.Y.; Ingmire, P.; He, Z.H. Tissue-specific and developmentally regulated expression of a cluster of tandemly arrayed cell wall-associated kinase-like kinase genes in arabidopsis. Plant Physiol. 2003, 133, 1732–1746. [Google Scholar] [CrossRef] [Green Version]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [Green Version]
- Harkenrider, M.; Sharma, R.; De Vleesschauwer, D.; Tsao, L.; Zhang, X.; Chern, M.; Canlas, P.; Zuo, S.; Ronald, P.C. Overexpression of Rice Wall-Associated Kinase 25 (OsWAK25) Alters Resistance to Bacterial and Fungal Pathogens. PLoS ONE 2016, 11, e0147310. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Ruzvidzo, O.; Morse, M.; Donaldson, L.; Kwezi, L.; Gehring, C. The Arabidopsis Wall Associated Kinase-Like 10 Gene Encodes a Functional Guanylyl Cyclase and Is Co-Expressed with Pathogen Defense Related Genes. PLoS ONE 2010, 5, e8904. [Google Scholar] [CrossRef] [Green Version]
- Delteil, A.; Gobbato, E.; Cayrol, B.; Estevan, J.; Michel-Romiti, C.; Dievart, A.; Kroj, T.; Morel, J.B. Several wall-associated kinases participate positively and negatively in basal defense against rice blast fungus. BMC Plant Biol. 2016, 16, 17. [Google Scholar] [CrossRef]
- Dmochowska-Boguta, M.; Kloc, Y.; Zielezinski, A.; Werecki, P.; Nadolska-Orczyk, A.; Karlowski, W.M.; Orczyk, W. TaWAK6 encoding wall-associated kinase is involved in wheat resistance to leaf rust similar to adult plant resistance. PLoS ONE 2020, 15, e0227713. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; He, S.Y.; Zhou, J.-M.; Xin, X.-F. Pattern-recognition receptors are required for NLR-mediated plant immunity. bioRxiv 2020, 4, 1–37. [Google Scholar]
- Baldassarre, V.; Cabassi, G.; Spadafora, N.D.; Aprile, A.; Mueller, C.T.; Rogers, H.J.; Ferrante, A. Wounding tomato fruit elicits ripening-stage specific changes in gene expression and production of volatile compounds. J. Exp. Bot. 2015, 66, 1511–1526. [Google Scholar] [CrossRef]
- Yu, W.; Zhao, R.; Sheng, J.; Shen, L. SIERF2 Is Associated with Methyl Jasmonate-Mediated Defense Response against Botrytis cinerea in Tomato Fruit. J. Agric. Food Chem. 2018, 66, 9923–9932. [Google Scholar] [CrossRef]
- Campos-Vargas, R.; Saltveit, M.E. Involvement of putative chemical wound signals in the induction of phenolic metabolism in wounded lettuce. Physiol. Plant. 2002, 114, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, V.; Gupta, A.K. RNAi mediated silencing of a wall associated kinase, OsiWAK1 in Oryza sativa results in impaired root development and sterility due to anther indehiscence. Physiol. Mol. Biol. Plants 2011, 17, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Huang, H.-J.; Ren, S.-T.; Li, J.-J.; Sun, Y.; Sun, D.-Y.; Zhang, S.-Q. The Rice Wall-Associated Receptor-Like Kinase Gene OsDEES1 Plays a Role in Female Gametophyte Development. Plant Physiol. 2012, 160, 696–707. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chr | AA | MW (kDa) | pI |
|---|---|---|---|---|---|
| SlWAK 1 | Solyc02g068660 | 2 | 701 | 77.7 | 7.38 |
| SlWAK 2 | Solyc07g007020 | 7 | 781 | 87.4 | 7.84 |
| SlWAK 3 | Solyc09g014710 | 9 | 787 | 87 | 8.39 |
| SlWAK 4 | Solyc09g014720 | 9 | 799 | 88.3 | 6.13 |
| SlWAK 5 | Solyc09g014730 | 9 | 757 | 84.2 | 6.36 |
| SlWAK 6 | Solyc09g014740 | 9 | 780 | 86.7 | 6.22 |
| SlWAK 7 | Solyc09g015230 | 9 | 393 | 44.1 | 5.74 |
| SlWAK 8 | Solyc09g015240 | 9 | 764 | 85.5 | 6.19 |
| SlWAK 9 | Solyc10g076530 | 10 | 503 | 56.4 | 6.11 |
| SlWAK 10 | Solyc10g076550 | 10 | 410 | 45.4 | 7.06 |
| SlWAK 11 | Solyc11g072140 | 11 | 765 | 84.3 | 5.91 |
| SlWAKL 1 | Solyc02g086270 | 2 | 775 | 86 | 5.65 |
| SlWAKL 2 | Solyc02g090110 | 2 | 703 | 77.8 | 7.73 |
| SlWAKL 3 | Solyc03g119240 | 3 | 673 | 75.3 | 5.55 |
| SlWAKL 4 | Solyc04g007390 | 4 | 335 | 33.7 | 5.39 |
| SlWAKL 5 | Solyc04g079710 | 4 | 679 | 75.3 | 6.82 |
| SlWAKL 6 | Solyc05g008930 | 5 | 746 | 84.1 | 6.77 |
| SlWAKL 7 | Solyc05g008940 | 5 | 657 | 74.1 | 6.65 |
| SlWAKL 8 | Solyc05g008950 | 5 | 626 | 70.9 | 8.38 |
| SlWAKL 9 | Solyc05g008960 | 5 | 665 | 75 | 6.68 |
| SlWAKL 10 | Solyc05g008970 | 5 | 458 | 52 | 6.7 |
| SlWAKL 11 | Solyc05g008980 | 5 | 631 | 71 | 6.19 |
| SlWAKL 12 | Solyc05g008990 | 5 | 663 | 74.9 | 5.81 |
| SlWAKL 13 | Solyc05g009010 | 5 | 302 | 35.3 | 6.6 |
| SlWAKL 14 | Solyc05g010530 | 5 | 380 | 42.9 | 6.31 |
| SlWAKL 15 | Solyc09g008640 | 9 | 628 | 70 | 8.23 |
| SlWAKL 16 | Solyc09g011200 | 9 | 637 | 71.3 | 8.85 |
| SlWAKL 17 | Solyc12g036330 | 12 | 651 | 72.9 | 6.17 |
| SlWAKL 18 | Solyc12g088040 | 12 | 621 | 68 | 5.76 |
| Phytohormone Responsive | Abiotic Stress-Responsive | Development-Related | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Function | Methyl Jasmonate | Abscisic Acid | Salicylic Acid | Gibberellin | Auxin | Light Response | Anaerobic | Wound | Low-Temperature | MYB Related | ||||||
| Cis-element | CGTCA-motif | TGACG-motif | ABRE | TCA-element | P-box | TATC-box | GARE-motif | TGA-element | AuxR-core | Box 4 | TCT-motif | ARE | WUN-motif | LTR | MYB | MBS |
| SlWAK1 | √ | √ | √ | √ | √ | √ | √ | |||||||||
| SlWAK2 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| SlWAK3 | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||
| SlWAK4 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAK5 | √ | √ | √ | √ | √ | |||||||||||
| SlWAK6 | √ | √ | √ | √ | √ | √ | √ | √ | √ | |||||||
| SlWAK7 | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | |||||
| SlWAK8 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| SlWAK9 | √ | √ | √ | √ | √ | |||||||||||
| SlWAK10 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAK11 | √ | √ | √ | √ | √ | |||||||||||
| SlWAKL1 | √ | √ | √ | √ | √ | √ | √ | |||||||||
| SlWAKL2 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAKL3 | √ | √ | √ | √ | √ | |||||||||||
| SlWAKL4 | √ | √ | √ | √ | ||||||||||||
| SlWAKL5 | √ | √ | √ | √ | √ | |||||||||||
| SlWAKL6 | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||
| SlWAKL7 | √ | √ | √ | √ | √ | √ | √ | |||||||||
| SlWAKL8 | √ | √ | √ | √ | ||||||||||||
| SlWAKL9 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAKL10 | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||
| SlWAKL11 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAKL12 | √ | √ | √ | √ | √ | √ | √ | |||||||||
| SlWAKL13 | √ | √ | √ | √ | √ | √ | ||||||||||
| SlWAKL14 | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||
| SlWAKL15 | √ | √ | √ | √ | √ | |||||||||||
| SlWAKL16 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| SlWAKL17 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| SlWAKL18 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| Mature Green | Breaker | Turning | Pink | Light Red | Red | |
|---|---|---|---|---|---|---|
| SlWAK3 | 1.0027 ± 0.0746 a | 0.5404 ± 0.1003 b | 0.5522 ± 0.0262 b | 0.3319 ± 0.0090 c | 0.2381 ± 0.0240 c | 0.2345 ± 0.0186 c |
| SlWAK7 | 0.0076 ± 0.0002 a | 0.0061 ± 0.0007 ab | 0.0073 ± 0.0001 a | 0.0049 ± 0.00001 b | 0.0046 ± 0.0002 b | 0.0041 ± 0.0003 b |
| SlWAK10 | 0.3709 ± 0.0198 a | 0.0666 ± 0.0021 ad | 0.1161 ± 0.0062 c | 0.0781 ± 0.0107 b | 0.0375 ± 0.0043 d | 0.0632 ± 0.0166 bd |
| SlWAKL1 | 0.7380 ± 0.1171 a | 0.7585 ± 0.0485 a | 0.7227 ± 0.1786 a | 0.3969 ± 0.0652 ab | 0.2154 ± 0.0103 b | 0.4637 ± 0.0123 b |
| SlWAKL2 | 4.6919 ± 0.4591 a | 7.3391 ± 1.0864 b | 7.5965 ± 1.0379 b | 8.4154 ± 0.2462 b | 4.3835 ± 0.2620 a | 5.0313 ± 0.2608 a |
| SlWAKL6 | 0.9826 ± 0.1147 a | 2.3389 ± 0.1333 bc | 3.0420 ± 0.2926 c | 3.4917 ± 0.0256 c | 1.7636 ± 0.1933 ab | 3.2342 ± 0.2895 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Song, Y.; Chen, D.; Zang, Y.; Zhang, Q.; Yi, Y.; Qu, G. Genome-Wide Identification, Classification, Characterization, and Expression Analysis of the Wall-Associated Kinase Family during Fruit Development and under Wound Stress in Tomato (Solanum lycopersicum L.). Genes 2020, 11, 1186. https://doi.org/10.3390/genes11101186
Sun Z, Song Y, Chen D, Zang Y, Zhang Q, Yi Y, Qu G. Genome-Wide Identification, Classification, Characterization, and Expression Analysis of the Wall-Associated Kinase Family during Fruit Development and under Wound Stress in Tomato (Solanum lycopersicum L.). Genes. 2020; 11(10):1186. https://doi.org/10.3390/genes11101186
Chicago/Turabian StyleSun, Zongyan, Yanping Song, Di Chen, Yudi Zang, Qiaoli Zhang, Yuetong Yi, and Guiqin Qu. 2020. "Genome-Wide Identification, Classification, Characterization, and Expression Analysis of the Wall-Associated Kinase Family during Fruit Development and under Wound Stress in Tomato (Solanum lycopersicum L.)" Genes 11, no. 10: 1186. https://doi.org/10.3390/genes11101186