Astrogliosis in an Experimental Model of Hypovitaminosis B12: A Cellular Basis of Neurological Disorders due to Cobalamin Deficiency

 , , , , , and
, , , , , and 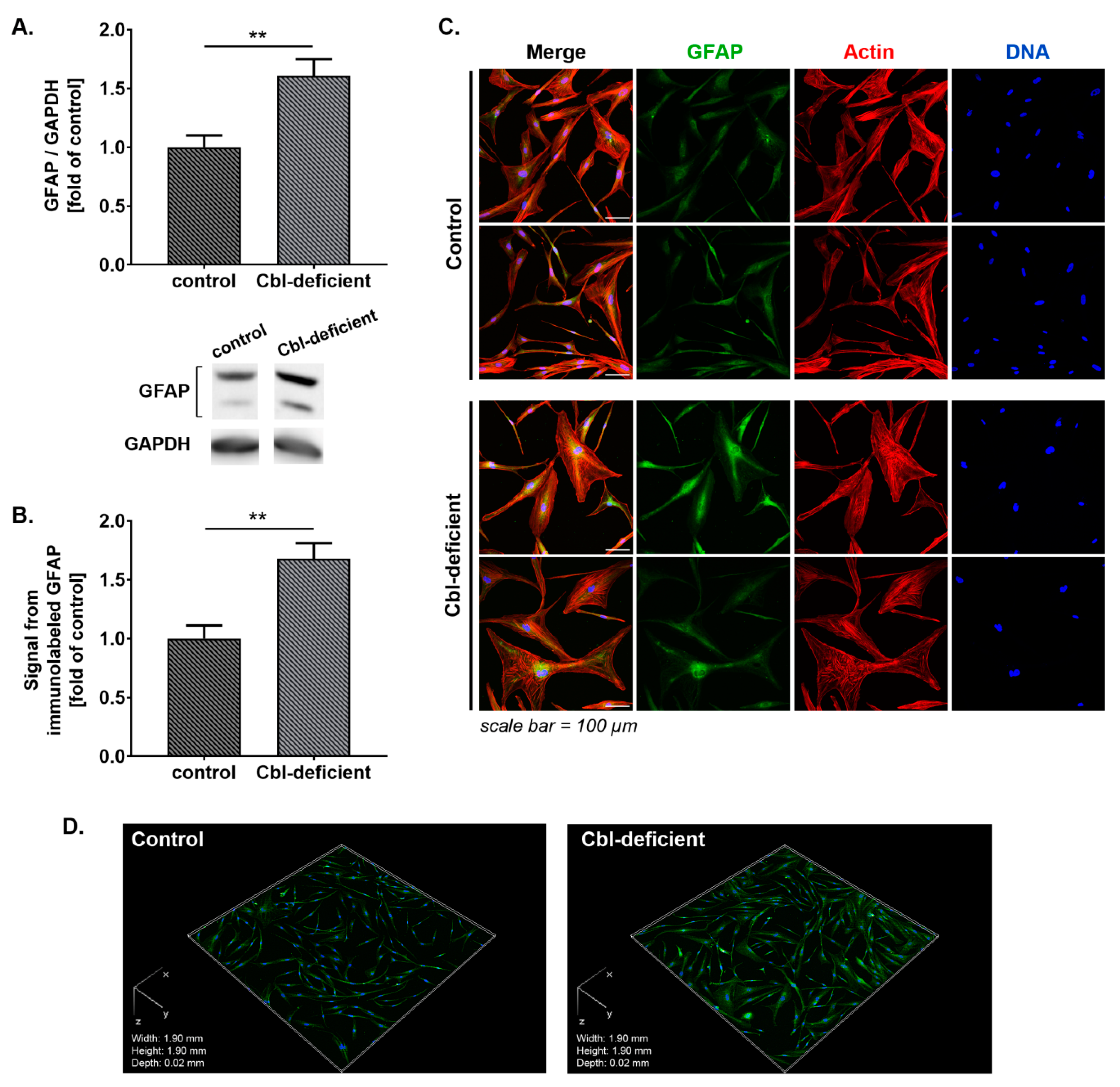{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Experimental Model
2.4. Western Blot Analysis
2.5. Immunocytochemistry and Fluorescence Imaging
2.6. Zebrafish Toxicity Evaluation
2.7. Statistical Analysis
3. Results
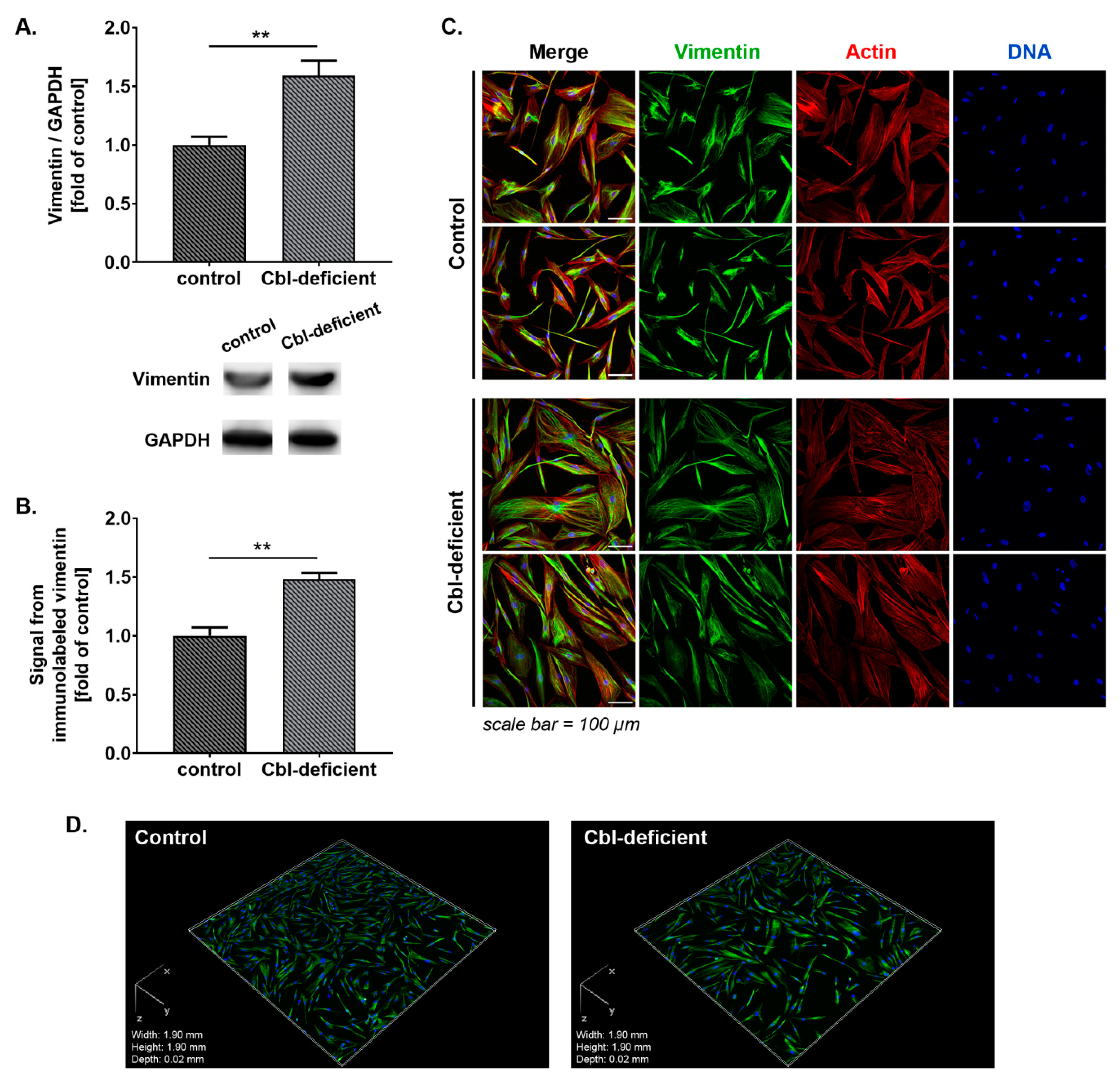
3.1. GFAP and Vimentin Was Overexpressed in Astrocytes Treated with c-lactam OH-Cbl
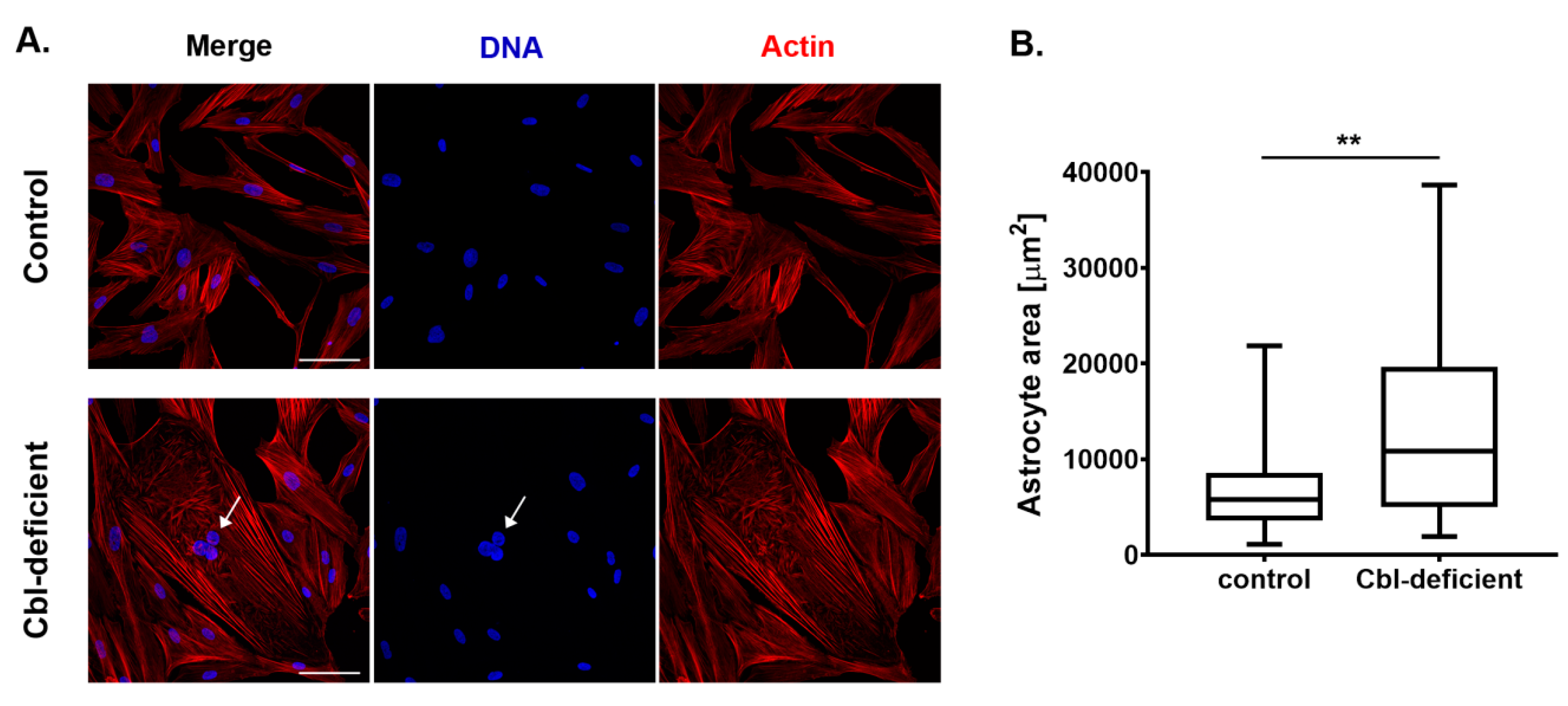
3.2. Astrocytes Upon Cobalamin Deficiency Developed Hypertrophic Morphology
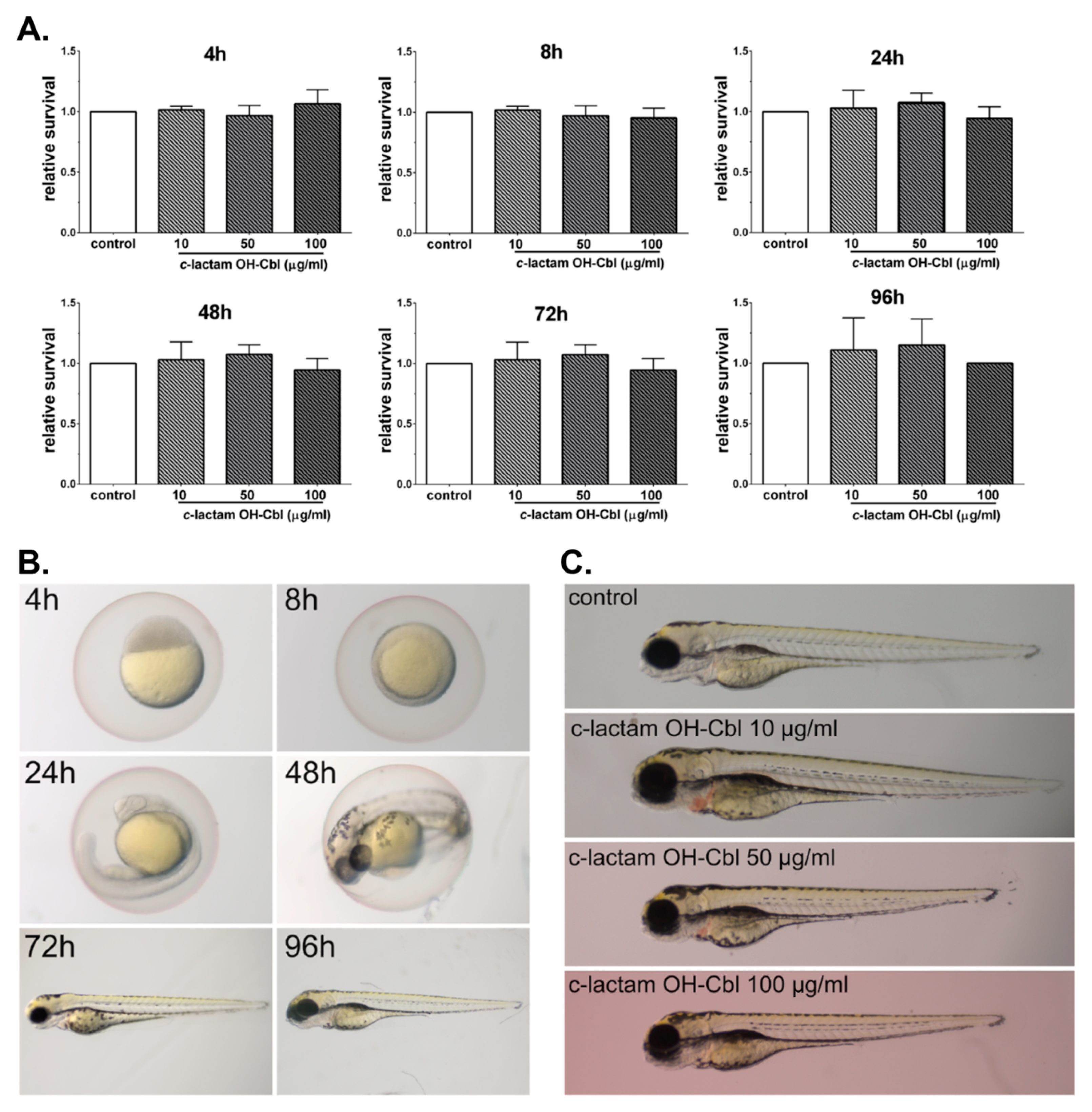
3.3. In Vivo Non-Toxocity of c-lactam OH-Cbl
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Green, R.; Allen, L.H.; Bjørke-Monsen, A.L.; Brito, A.; Guéant, J.L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.H.; et al. Vitamin B12 deficiency. Nat. Rev. Dis. Primers 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.; Davis, B. Iron, vitamin B12 and folate. Medicine 2017, 45, 198–203. [Google Scholar] [CrossRef]
- Wolffenbuttel, B.H.R.; Wouters, H.J.C.M.; Heiner-Fokkema, M.R.; van der Klauw, M.M. The many faces of cobalamin (vitamin B12) deficiency. Mayo Clin. Proc. Innov. Qual. Outcomes 2019, 3, 200–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brass, E.P.; Allen, R.H.; Ruff, L.J.; Stabler, S.P. Effect of hydroxycobalamin[c-lactam] on propionate and carnitine metabolism in the rat. Biochem. J. 1990, 266, 809–815. [Google Scholar]
- Stabler, S.P.; Brass, E.P.; Marcell, P.D.; Allen, R.H. Inhibition of cobalamin-dependent enzymes by cobalamin analogues in rats. J. Clin. Investig. 1991, 87, 1422–1430. [Google Scholar] [CrossRef]
- Sponne, I.E.; Gaire, D.; Stabler, S.P.; Droesch, S.; Barbé, F.M.; Allen, R.H.; Lambert, D.A.; Nicolas, J.P. Inhibition of vitamin B12 metabolism by OH-cobalamin c-lactam in rat oligodendrocytes in culture: A model for studying neuropathy due to vitamin B12 deficiency. Neurosci. Lett. 2000, 288, 191–194. [Google Scholar] [CrossRef]
- Sauer, S.W.; Opp, S.; Haarmann, A.; Okun, J.G.; Kölker, S.; Morath, M.A. Long-term exposure of human proximal tubule cells to hydroxycobalamin[c-lactam] as a possible model to study renal disease in methylmalonic acidurias. J. Inherit. Metab. Dis. 2009, 32, 720–727. [Google Scholar] [CrossRef]
- Haegler, P.; Grünig, D.; Berger, B.; Krähenbühl, S.; Bouitbir, J. Impaired mitochondrial function in HepG2 cells treated with hydroxy-cobalamin[c-lactam]: A cell model for idiosyncratic toxicity. Toxicology 2015, 336, 48–58. [Google Scholar] [CrossRef]
- Haegler, P.; Grünig, D.; Berger, B.; Terracciano, L.; Krähenbühl, S.; Bouitbir, J. Hepatic effects of pharmacological doses of hydroxy-cobalamin[c-lactam] in mice. PLoS ONE 2017, 12, e0171026. [Google Scholar] [CrossRef]
- Zelder, F.; Sonnay, M.; Prieto, L. Antivitamins for medicinal applications. Chembiochem 2015, 16, 1264–1278. [Google Scholar] [CrossRef]
- Rzepka, Z.; Respondek, M.; Pawlik, J.; Beberok, A.; Gryko, D.; Wrześniok, D. Cobalamin deficiency: Effect on homeostasis of cultured human astrocytes. Cells 2019, 8, 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyenet, S.J.; Nguyen, H.T.; Hwang, B.H.; Schwartz, M.W.; Baskin, D.G.; Thaler, J.P. High-fat diet feeding causes rapid, non-apoptotic cleavage of caspase-3 in astrocytes. Brain Res. 2013, 1512, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aras, R.; Barron, A.M.; Pike, C.J. Caspase activation contributes to astrogliosis. Brain Res. 2012, 1450, 102–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villapol, S.; Acarin, L.; Faiz, M.; Castellano, B.; Gonzalez, B. Survivin and heat shock protein 25/27 colocalize with cleaved caspase-3 in surviving reactive astrocytes following excitotoxicity to the immature brain. Neuroscience 2008, 153, 108–119. [Google Scholar] [CrossRef]
- Osborn, L.M.; Kamphuis, W.; Wadman, W.J.; Hol, E.M. Astrogliosis: An integral player in the pathogenesis of Alzheimer’s disease. Prog. Neurobiol. 2016, 144, 121–141. [Google Scholar] [CrossRef]
- Oksanen, M.; Lehtonen, S.; Jaronen, M.; Goldsteins, G.; Hämäläinen, R.H.; Koistinaho, J. Astrocyte alterations in neurodegenerative pathologies and their modeling in human induced pluripotent stem cell platforms. Cell. Mol. Life Sci. 2019, 76, 2739–2760. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Hu, X.; Qian, L.; O’Callaghan, J.P.; Hong, J.S. Astrogliosis in CNS pathologies: Is there a role for microglia? Mol. Neurobiol. 2010, 41, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.J.; Olabarria, M.; Chvatal, A.; Verkhratsky, A. Astroglia in dementia and Alzheimer’s disease. Cell Death Differ. 2009, 16, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Rzepka, Z.; Respondek, M.; Beberok, A.; Ó Proinsias, K.; Gryko, D.; Wrześniok, D. Vitamin B12 deficiency induces imbalance in melanocytes homeostasis—A cellular basis of hypocobalaminemia pigmentary manifestations. Int. J. Mol. Sci. 2018, 19, 2845. [Google Scholar] [CrossRef] [Green Version]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.; Carr, G.J.; Cenijn, P.; Fochtman, P.; Gourmelon, A.; Hübler, N.; et al. OECD validation study to assess intra- and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD Publishing: Paris, France, 2013. [Google Scholar] [CrossRef]
- Tykhomyrov, A.A.; Pavlova, A.S.; Nedzvetsky, V.S. Glial fibrillary acidic protein (GFAP): On the 45th anniversary of its discovery. Neurophysiology 2016, 48, 54–71. [Google Scholar] [CrossRef]
- Yu, P.; Wang, H.; Katagiri, Y.; Geller, H.M. An in vitro model of reactive astrogliosis and its effect on neuronal growth. Methods Mol Biol. 2012, 814, 327–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielsson, F.; Peterson, M.K.; Caldeira Araújo, H.; Lautenschläger, F.; Gad, A.K. Vimentin diversity in health and disease. Cells 2018, 7, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Rodriguez, A.B.; Acaz-Fonseca, E.; Viveros, M.P.; Garcia-Segura, L.M. Changes in cannabinoid receptors, aquaporin 4 and vimentin expression after traumatic brain injury in adolescent male mice. Association with edema and neurological deficit. PLoS ONE 2015, 10, e0128782. [Google Scholar] [CrossRef] [Green Version]
- Pekny, M.; Pekna, M. Astrocyte reactivity and reactive astrogliosis: Costs and benefits. Physiol. Rev. 2014, 94, 1077–1098. [Google Scholar] [CrossRef]
- Ekmark-Lewén, S.; Lewén, A.; Israelsson, C.; Li, G.L.; Farooque, M.; Olsson, Y.; Ebendal, T.; Hillered, L. Vimentin and GFAP responses in astrocytes after contusion trauma to the murine brain. Restor. Neurol. Neurosci. 2010, 28, 311–321. [Google Scholar] [CrossRef]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [Green Version]
- de Pablo, Y.; Nilsson, M.; Pekna, M.; Pekny, M. Intermediate filaments are important for astrocyte response to oxidative stress induced by oxygen-glucose deprivation and reperfusion. Histochem. Cell Biol. 2013, 140, 81–91. [Google Scholar] [CrossRef]
- Wilhelmsson, U.; Li, L.; Pekna, M.; Berthold, C.H.; Blom, S.; Eliasson, C.; Renner, O.; Bushong, E.; Ellisman, M.; Morgan, T.E.; et al. Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration. J. Neurosci. 2004, 24, 5016–5021. [Google Scholar] [CrossRef] [Green Version]
- Pablo, Y.D.; Marasek, P.; Pozo-Rodrigálvarez, A.; Wilhelmsson, U.; Inagaki, M.; Pekna, M.; Pekny, M. Vimentin phosphorylation is required for normal cell division of immature astrocytes. Cells 2019, 8, 1016. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wu, Y.; Xie, F.; Zhong, Y.; Wang, Y.; Xu, M.; Feng, J.; Charish, J.; Monnier, P.P.; Qin, X. RGMa mediates reactive astrogliosis and glial scar formation through TGFβ1/Smad2/3 signaling after stroke. Cell Death Differ. 2018, 25, 1503–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eide, P.K.; Hansson, H.A. Astrogliosis and impaired aquaporin-4 and dystrophin systems in idiopathic normal pressure hydrocephalus. Neuropathol. Appl. Neurobiol. 2018, 44, 474–490. [Google Scholar] [CrossRef] [PubMed]
- Wallis, N.; Lau, C.L.; Farg, M.A.; Atkin, J.D.; Beart, P.M.; O’Shea, R.D. SOD1 mutations causing familial amyotrophic lateral sclerosis induce toxicity in astrocytes: Evidence for bystander effects in a continuum of astrogliosis. Neurochem. Res. 2018, 43, 166–179. [Google Scholar] [CrossRef] [PubMed]
- de Amorim, V.C.; Júnior, M.S.; da Silva, A.B.; David, J.M.; David, J.P.; Costa, M.D.; Butt, A.M.; da Silva, V.D.; Costa, S.L. Agathisflavone modulates astrocytic responses and increases the population of neurons in an in vitro model of traumatic brain injury. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Joshi, N. Astrocytes: Inexplicable cells in neurodegeneration. Int. J. Neurosci. 2017, 127, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Sosunov, A.; Wu, X.; McGovern, R.; Mikell, C.; McKhann, G.M.; Goldman, J.E. Abnormal mitosis in reactive astrocytes. Acta Neuropathol. Commun. 2020, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Bito, T.; Yasui, M.; Iwaki, T.; Yabuta, Y.; Ichiyanagi, T.; Yamaji, R.; Nakano, Y.; Inui, H.; Watanabe, F. Dodecylamine derivative of hydroxocobalamin acts as a potent inhibitor of cobalamin-dependent methionine synthase in mammalian cultured COS-7 cells. Food Nutr. Sci. 2014, 5, 1318–1325. [Google Scholar] [CrossRef] [Green Version]
- Wasel, O.; Freeman, J.L. Chemical and genetic zebrafish models to define mechanisms of and treatments for dopaminergic neurodegeneration. Int. J. Mol. Sci. 2020, 21, 5981. [Google Scholar] [CrossRef]
- Oosterhof, N.; Holtman, I.R.; Kuil, L.E.; van der Linde, H.C.; Boddeke, E.W.; Eggen, B.J.; van Ham, T.J. Identification of a conserved and acute neurodegeneration-specific microglial transcriptome in the zebrafish. Glia 2017, 65, 138–149. [Google Scholar] [CrossRef]
- Koiwa, J.; Shiromizu, T.; Adachi, Y.; Ikejiri, M.; Nakatani, K.; Tanaka, T.; Nishimura, Y. Generation of a triple-transgenic zebrafish line for assessment of developmental neurotoxicity during neuronal differentiation. Pharmaceuticals (Basel) 2019, 12, 145. [Google Scholar] [CrossRef] [Green Version]
- Saleem, S.; Kannan, R.R. Zebrafish: An emerging real-time model system to study Alzheimer’s disease and neurospecific drug discovery. Cell. Death. Discov. 2018, 4, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Poskanzer, K.E.; Freeman, M.R.; Monk, K.R. Live-imaging of astrocyte morphogenesis and function in zebrafish neural circuits. Nat. Neurosci. 2020, 23, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, J.Y.; You, S.; Song, G.; Lim, W. Neurotoxic effects of aflatoxin B1 on human astrocytes in vitro and on glial cell development in zebrafish in vivo. J. Hazard. Mater. 2020, 386, 121639. [Google Scholar] [CrossRef] [PubMed]
- Leme, E.; Silva, E.P.; Rodrigues, P.S.; Silva, I.R.; Martins, M.F.; Bondan, E.F.; Bernardi, M.M.; Kirsten, T.B. Billings reservoir water used for human consumption presents microbiological contaminants and induces both behavior impairments and astrogliosis in zebrafish. Ecotoxicol. Environ. Saf. 2018, 161, 364–373. [Google Scholar] [CrossRef]
- Yin, G.; Du, M.; Li, R.; Li, K.; Huang, X.; Duan, D.; Ai, X.; Yao, F.; Zhang, L.; Hu, Z.; et al. Glia maturation factor beta is required for reactive gliosis after traumatic brain injury in zebrafish. Exp. Neurol. 2018, 305, 129–138. [Google Scholar] [CrossRef]
- Greibe, E.; Fedosov, S.; Nexo, E. The cobalamin-binding protein in zebrafish is an intermediate between the three cobalamin-binding proteins in human. PLoS ONE 2012, 7, e35660. [Google Scholar] [CrossRef]
- Benoit, C.R.; Stanton, A.E.; Tartanian, A.C.; Motzer, A.R.; McGaughey, D.M.; Bond, S.R.; Brody, L.C. Functional and phylogenetic characterization of noncanonical vitamin B12-binding proteins in zebrafish suggests involvement in cobalamin transport. J. Biol. Chem. 2018, 293, 17606–17621. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Ando, I.; Kukita, A. Generalized hyperpigmentation of the skin due to vitamin B12 deficiency. J. Dermatol. 2001, 28, 282–285. [Google Scholar] [CrossRef]
- Niiyama, S.; Mukai, H. Reversible cutaneous hyperpigmentation and nails with white hair due to vitamin B12 deficiency. Eur. J. Dermatol. 2007, 17, 551–552. [Google Scholar] [CrossRef]
- Kannan, R.; Ng, M.J. Cutaneous lesions and vitamin B12 deficiency: An often-forgotten link. Can. Fam. Physician 2008, 54, 529–532. [Google Scholar]
- Lee, M.S.; Bonner, J.R.; Bernard, D.J.; Sanchez, E.L.; Sause, E.T.; Prentice, R.R.; Burgess, S.M.; Brody, L.C. Disruption of the folate pathway in zebrafish causes developmental defects. BMC Dev. Biol. 2012, 12, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzepka, Z.; Rok, J.; Kowalska, J.; Banach, K.; Hermanowicz, J.M.; Beberok, A.; Sieklucka, B.; Gryko, D.; Wrześniok, D. Astrogliosis in an Experimental Model of Hypovitaminosis B12: A Cellular Basis of Neurological Disorders due to Cobalamin Deficiency. Cells 2020, 9, 2261. https://doi.org/10.3390/cells9102261
Rzepka Z, Rok J, Kowalska J, Banach K, Hermanowicz JM, Beberok A, Sieklucka B, Gryko D, Wrześniok D. Astrogliosis in an Experimental Model of Hypovitaminosis B12: A Cellular Basis of Neurological Disorders due to Cobalamin Deficiency. Cells. 2020; 9(10):2261. https://doi.org/10.3390/cells9102261
Chicago/Turabian StyleRzepka, Zuzanna, Jakub Rok, Justyna Kowalska, Klaudia Banach, Justyna Magdalena Hermanowicz, Artur Beberok, Beata Sieklucka, Dorota Gryko, and Dorota Wrześniok. 2020. "Astrogliosis in an Experimental Model of Hypovitaminosis B12: A Cellular Basis of Neurological Disorders due to Cobalamin Deficiency" Cells 9, no. 10: 2261. https://doi.org/10.3390/cells9102261