The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
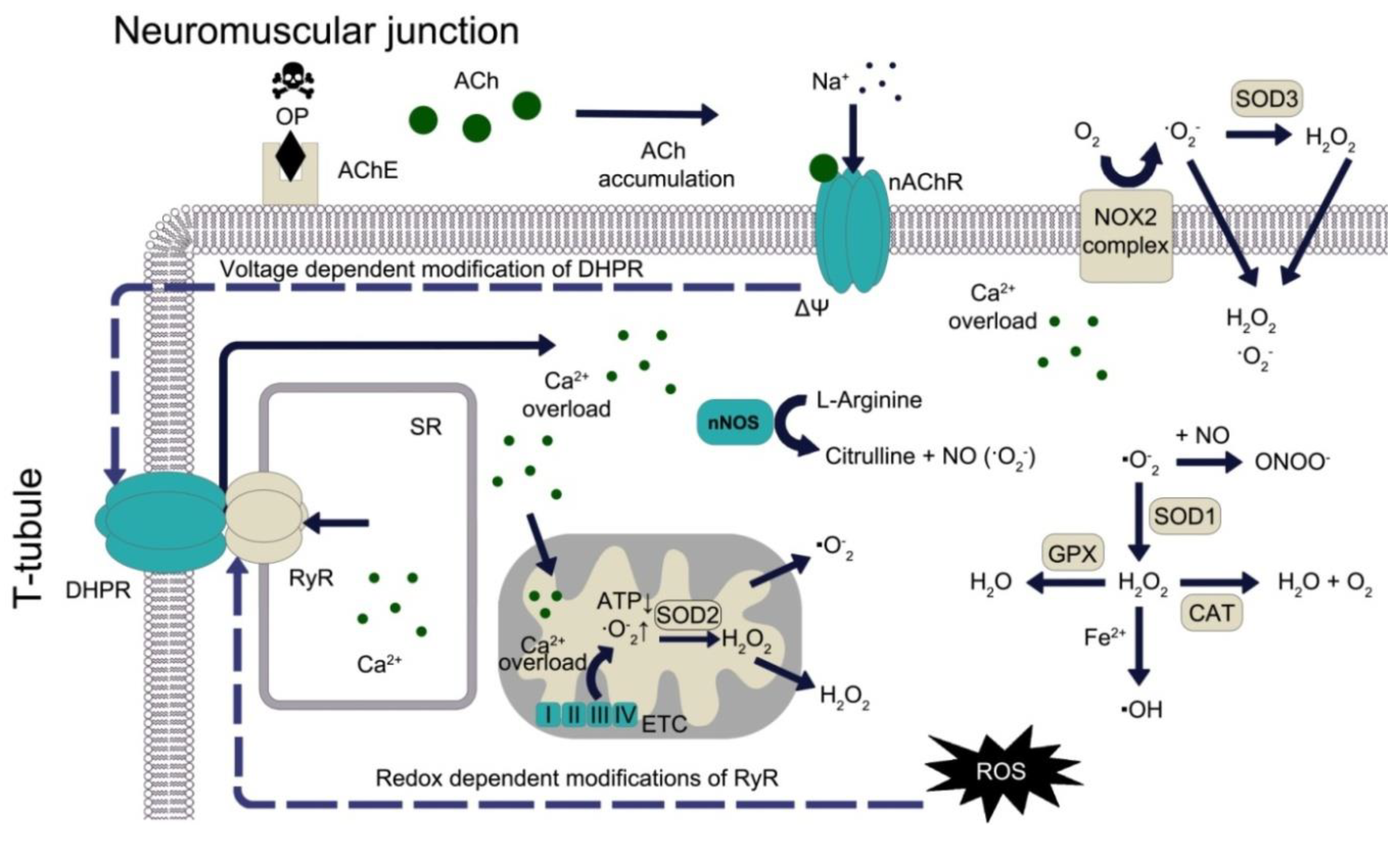
:1. Redox System in Health and Disease: Brief Overview
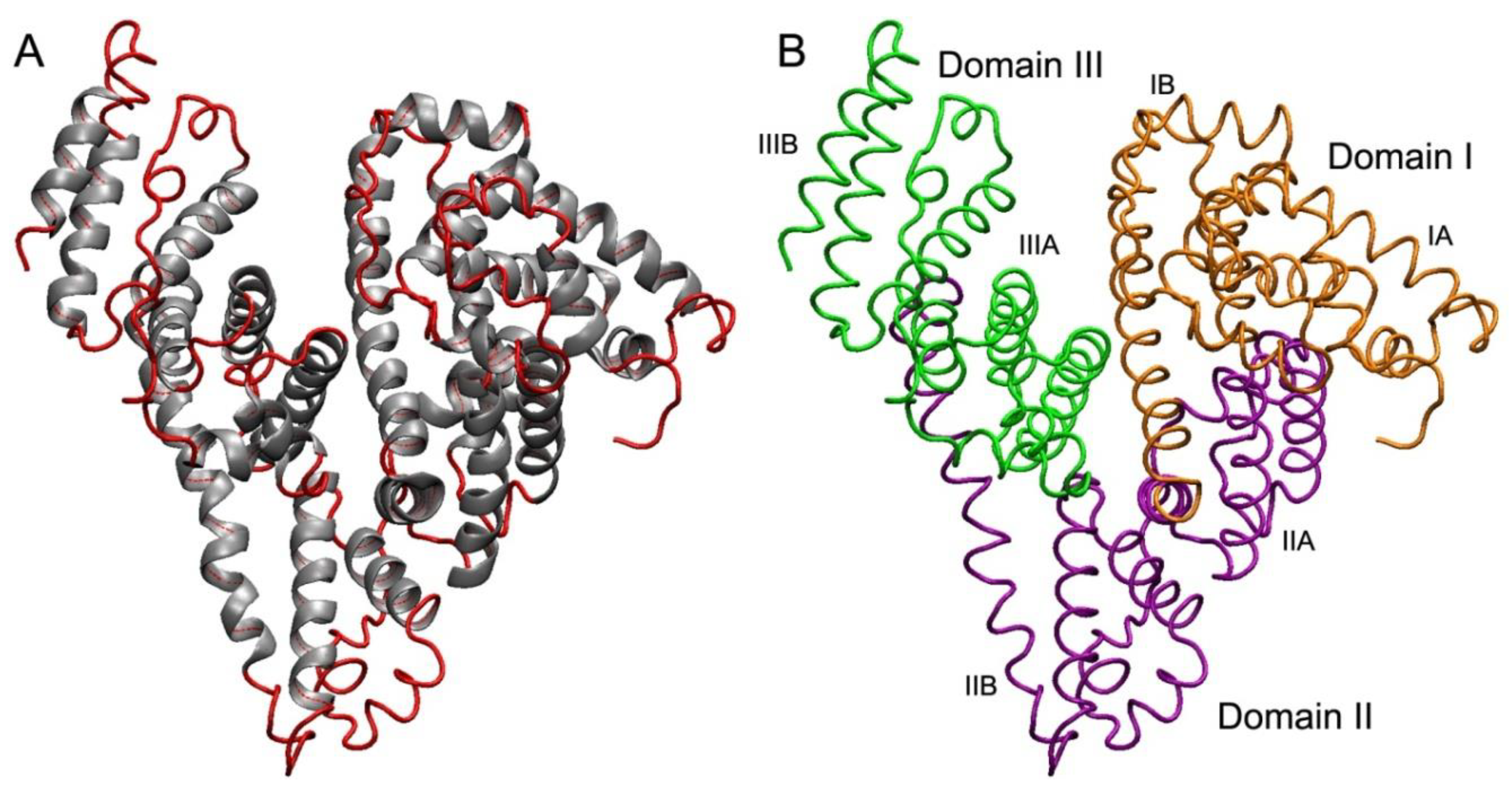
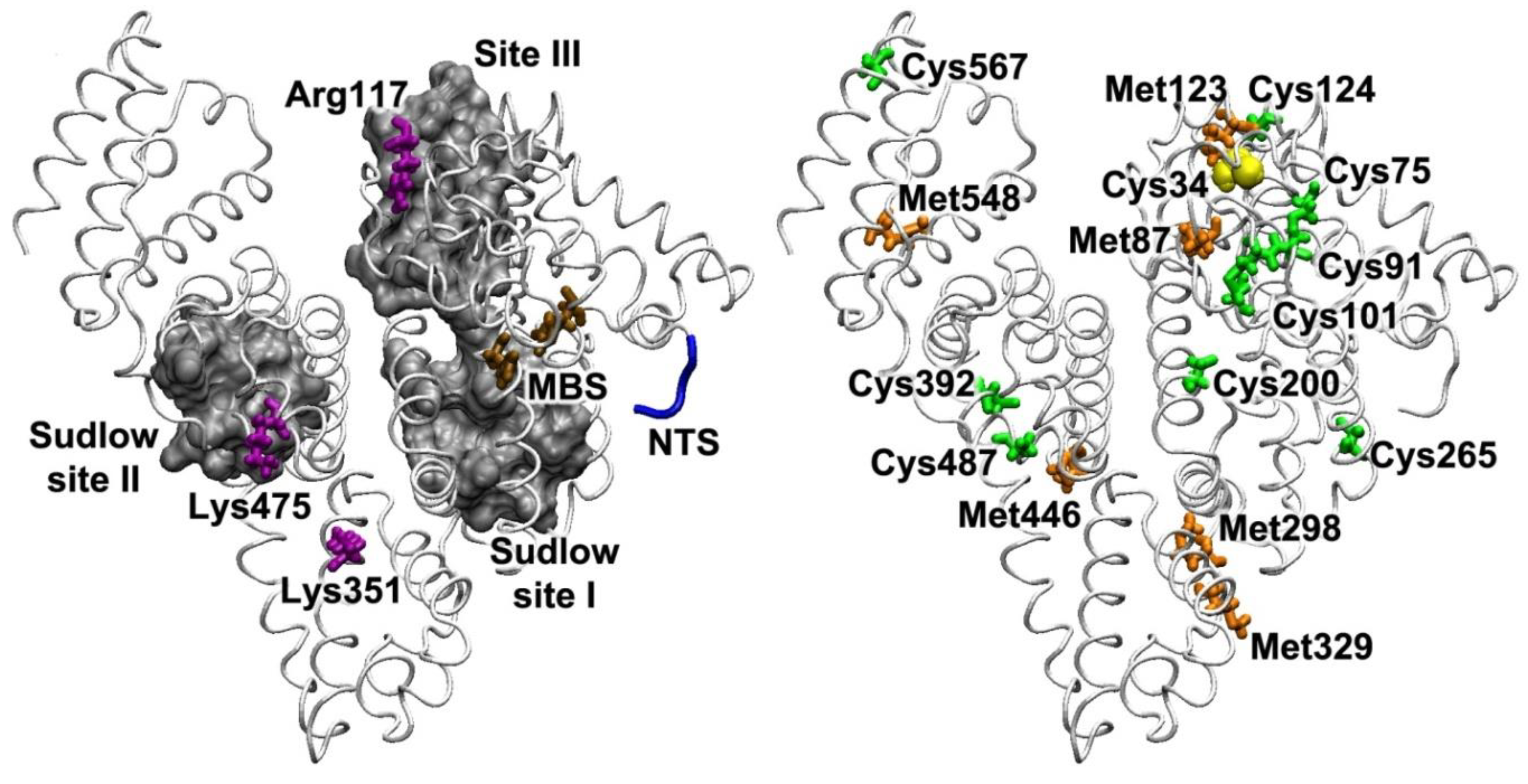
2. Structural Characteristics of Albumin and Their Interspecies Features
3. Albumin Participates in the Redox Modulation of Blood Plasma and Interstitial Fluid
3.1. Binding of Polyvalent Metal Ions
3.2. ROS Neutralisation
3.3. Interaction of Albumin with Low-Molecular-Weight Thiols
3.4. Enzymatic Antioxidant Activity of Albumin
3.5. Indirect Mechanisms of Albumin Participation in the Antioxidant Defence of the Body
4. Interplay of Binding, Enzymatic and Antioxidant Properties of Albumin
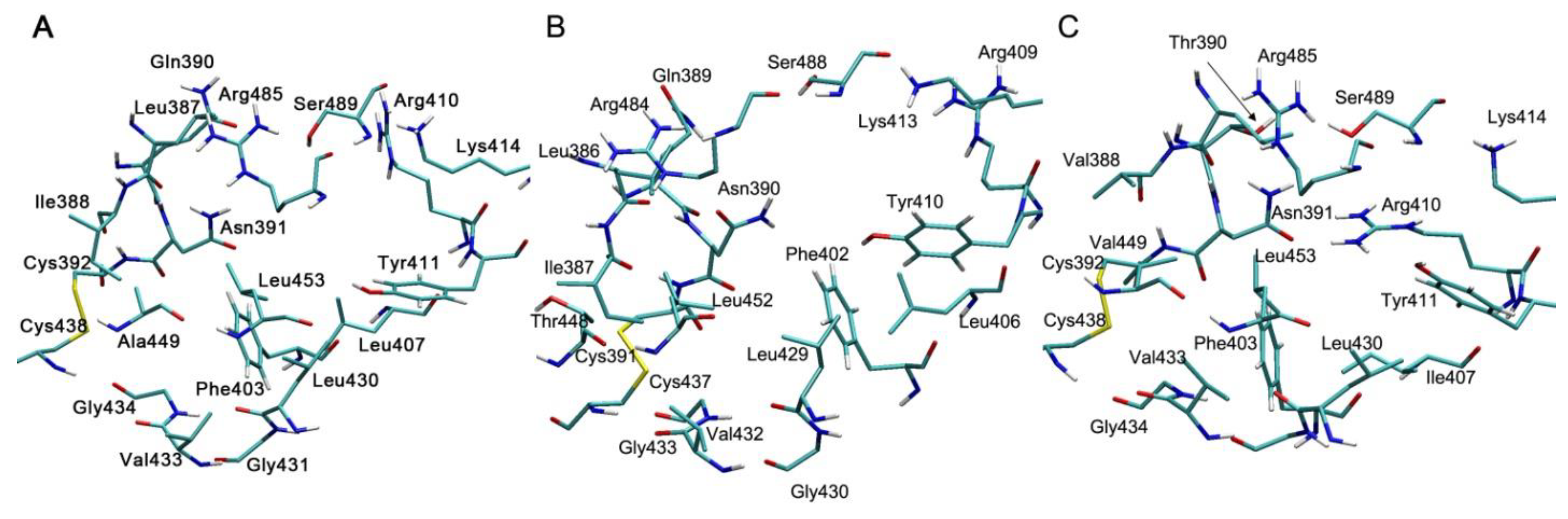
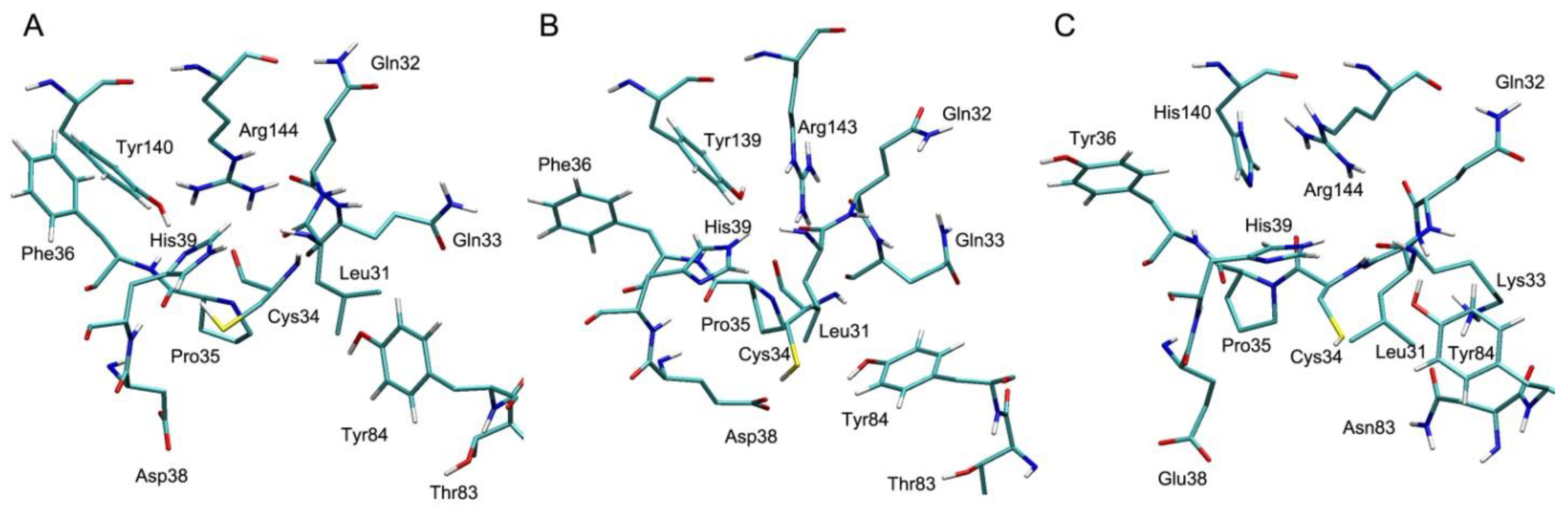
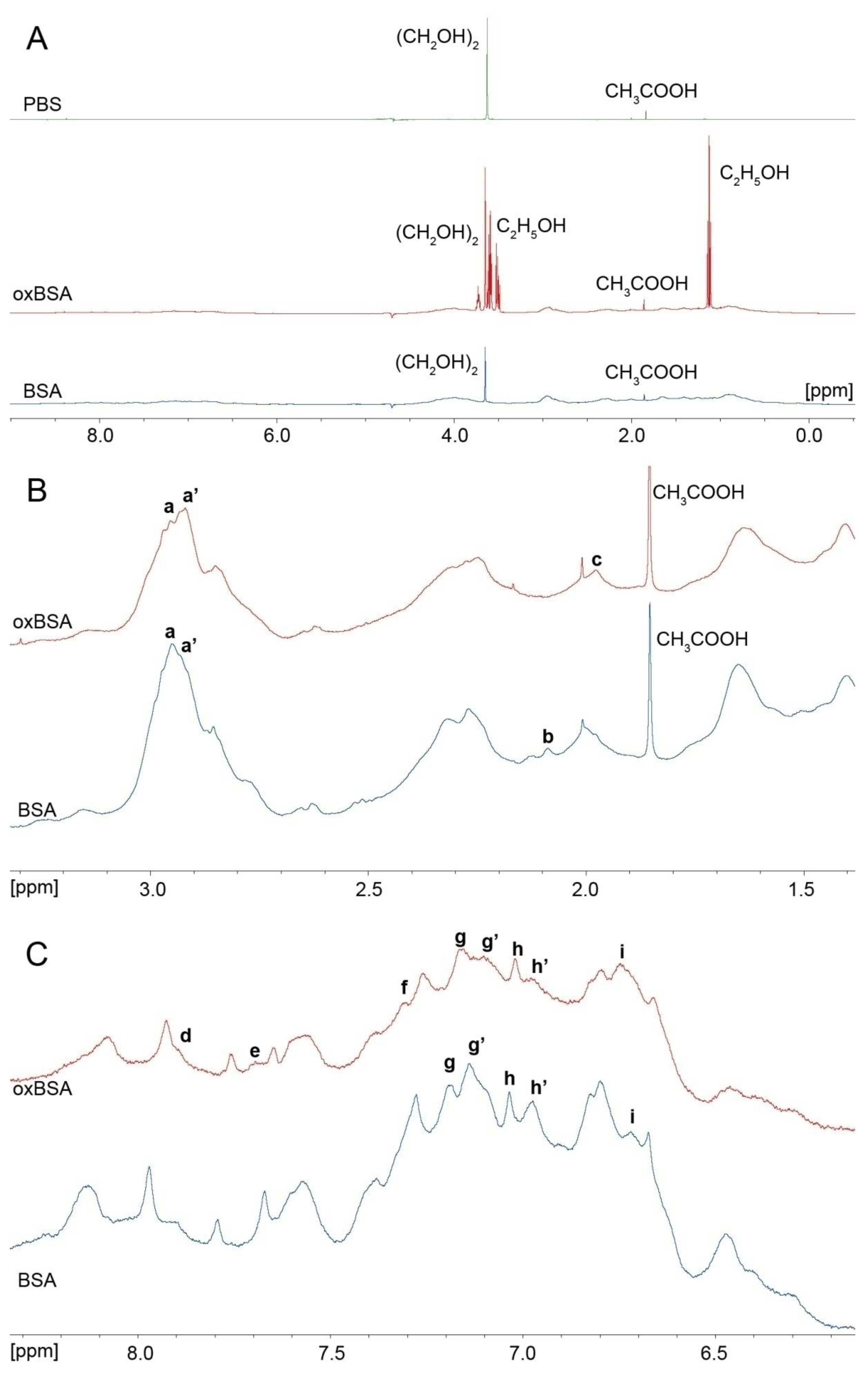
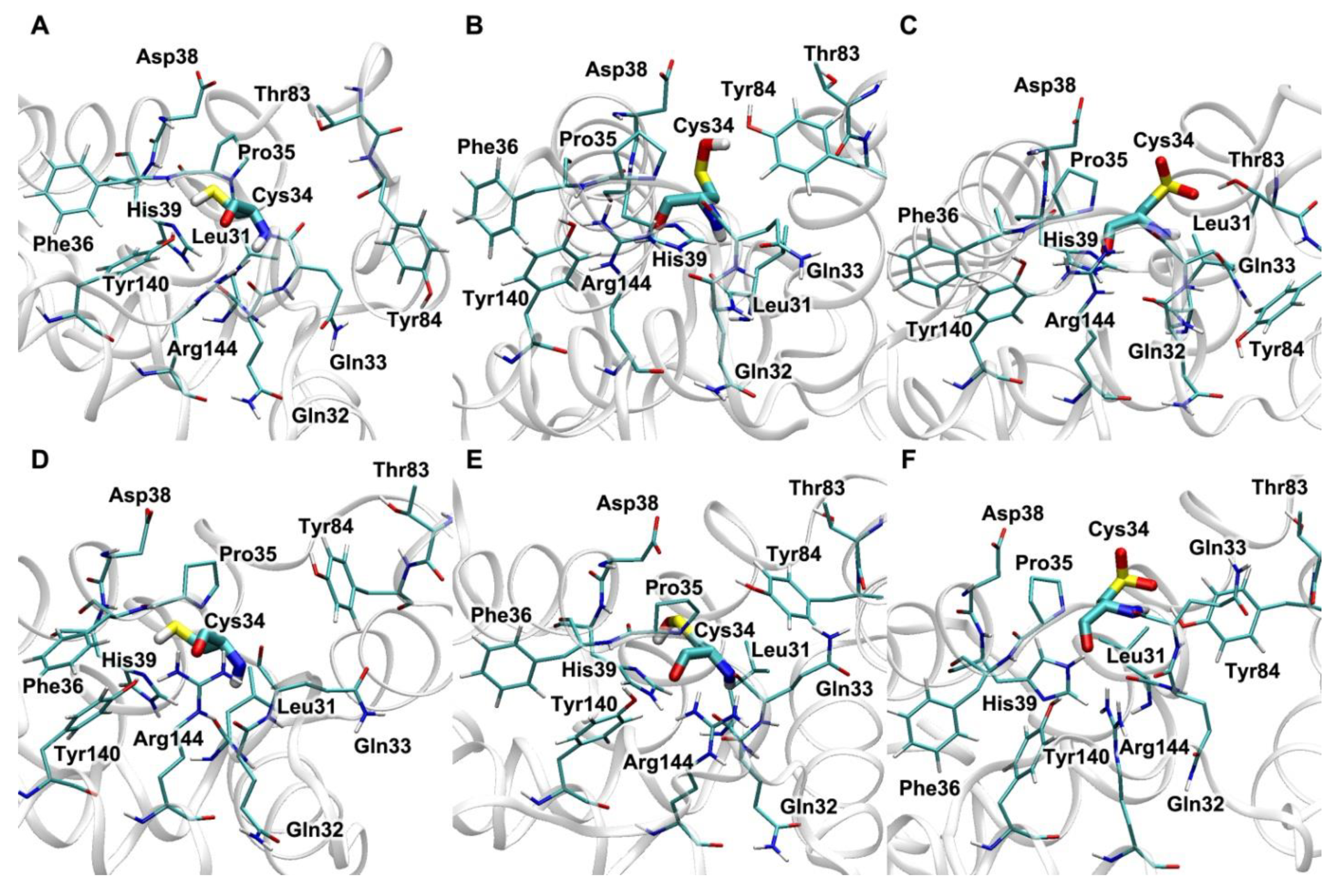
4.1. Effect of Cys34 Oxidation on the Functional Properties of Albumin
4.2. Effect of Cys34 Oxidation on the Structural Properties of Albumin
4.3. Effect of Endogenous and Exogenous Compounds on the Reactivity of the Thiol Group of Cys34
5. Antioxidant Properties of Albumin: Practical Application
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 1H NMR | Proton nuclear magnetic resonance |
| ACh | Acetylcholine |
| AChE | Acetylcholinesterase |
| BH4 | Tetrahydrobiopterin |
| BSA | Bovine serum albumin |
| CaM | Calmodulin |
| CAT | Catalase |
| CKD | Chronic kidney disease |
| COVID-19 | Coronavirus Disease 2019 |
| CYP450 | Cytochrome-dependent oxygenases |
| DHPR | Dihydropyridine receptors |
| DMP | Dimethylphosphoryl |
| DTNB | 5,5′-dithiobis-2-nitrobenzoic acid |
| DTT | Dithiothreitol |
| EGCG | Epigallocatechin gallate |
| eNOS | Endothelial nitric oxide synthase |
| EPR | Electron paramagnetic resonance |
| ESOET | 2-(ethylsulfinyl) ethanethiol |
| EtAc | Ethacrynic acid |
| ETC | Electron transport chain |
| EtOH | Ethanol |
| FAs | Fatty Acids |
| GPx | Glutathione peroxidase |
| GPx3 | Tetrameric glutathione peroxidase-3 |
| GR | Glutathione reductase |
| Grx | Glutaredoxins |
| GSH | Reduced glutathione |
| GSSG | Oxidised glutathione |
| GST | Glutathione-S-transferase |
| GTE | Green tea extract |
| HPLC | High-performance liquid chromatography |
| HSA | Human serum albumin |
| ICU | Intensive care unit |
| iNOS | Inducible nitric oxide synthase |
| LCAD | Long-chain fatty acid dehydrogenase |
| MBS | Metal-binding site |
| MCO | Metal–catalysed oxidation system |
| MPDS | Mean predicted disorder score |
| MER | Mercaptoethanol |
| MM-PBSA | Molecular Mechanics/Poisson-Boltzmann Surface Area |
| nNOS | Neuronal nitric oxide synthase |
| NOS | Nitric oxide synthase |
| NOX | NAD(P)H-oxidase system |
| NPA | p-nitrophenyl acetate |
| NTS | N-terminal site |
| ODM | O,O-Dimethyl S-2-(ethylsulfanyl)ethyl phosphorothioate |
| OPs | Organophosphates |
| oxBSA | Oxidised bovine serum albumin |
| PBS | Phosphate buffered saline |
| PHGPx | Phospholipid hydroperoxide glutathione peroxidase |
| PON | Paraoxonase |
| Prxs | Peroxiredoxin |
| PUFAs | Polyunsaturated fatty acids |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| RSA | Rat albumin |
| RSS | Reactive sulfur species |
| RyR | Ryanodine receptors |
| S-LME | S-lauroylmercaptoethanol |
| SOD | Superoxide dismutase |
| SR | Sarcoplasmic reticulum |
| TR | Thioredoxin reductase |
| Trx | Thioredoxin |
| VLCAD | Very long-chain fatty acid dehydrogenase |
| XO | Xanthine oxidase |
References
- Weidinger, A.; Kozlov, A.V. Biological Activities of Reactive Oxygen and Nitrogen Species: Oxidative Stress versus Signal Transduction. Biomolecules 2015, 5, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milkovic, L.; Gasparovic, A.C.; Cindric, M.; Mouthuy, P.-A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncharov, N.; Avdonin, P.; Nadeev, A.; Zharkikh, I.; Jenkins, R. Reactive Oxygen Species in Pathogenesis of Atherosclerosis. Curr. Pharm. Des. 2015, 21, 1134–1146. [Google Scholar] [CrossRef] [Green Version]
- Chocry, M.; Leloup, L. The NADPH Oxidase Family and Its Inhibitors. Antioxid. Re. Sign. 2020, 33, 332–353. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.F.; Laitano, O. Regulation of NADPH oxidases in skeletal muscle. Free Radic. Biol. Med. 2016, 98, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violi, F.; Oliva, A.; Cangemi, R.; Ceccarelli, G.; Pignatelli, P.; Carnevale, R.; Cammisotto, V.; Lichtner, M.; Alessandri, F.; De Angelis, M.; et al. Nox2 activation in Covid-19. Redox Biol. 2020, 36, 101655. [Google Scholar] [CrossRef]
- Kalogeris, T.; Bao, Y.; Korthuis, R.J. Mitochondrial reactive oxygen species: A double edged sword in ischemia/reperfusion vs preconditioning. Re. Biol. 2014, 2, 702–714. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J. An Update on Mitochondrial Reactive Oxygen Species Production. Antioxidants 2020, 9, 472. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Hear. J. 2011, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Santolini, J. What does “NO-Synthase” stand for? Front. Biosci. 2019, 24, 133–171. [Google Scholar] [CrossRef]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Battelli, M.G.; Polito, L.; Bortolotti, M.; Bolognesi, A. Xanthine Oxidoreductase-Derived Reactive Species: Physiological and Pathological Effects. Oxid. Med. Cell. Longev. 2015, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Li, C.; Mozziconacci, O.; Zhu, R.; Xu, Y.; Tang, Y.; Chen, R.; Huang, Y.; Holzbeierlein, J.M.; Schöneich, C.; et al. Xanthine oxidase-mediated oxidative stress promotes cancer cell-specific apoptosis. Free Radic. Biol. Med. 2019, 139, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Nagata, N.; Shimakami, T.; Shirakura, T.; Matsui, C.; Ni, Y.; Zhuge, F.; Xu, L.; Chen, G.; Nagashimada, M.; et al. Xanthine oxidase inhibition attenuates insulin resistance and diet-induced steatohepatitis in mice. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Fatunde, O.A.; Brown, S.-A. The Role of CYP450 Drug Metabolism in Precision Cardio-Oncology. Int. J. Mol. Sci. 2020, 21, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, C.; Wang, S.; Zhou, Q.; Dai, D.; Shi, J.; Xu, X.; Luo, Q. Cytochrome P450-Based Drug-Drug Interactions of Vonoprazan In Vitro and In Vivo. Front. Pharmacol. 2020, 11, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ghiaty, M.A.; Shoieb, S.M.; El-Kadi, A.O. Cytochrome P450-mediated drug interactions in COVID-19 patients: Current findings and possible mechanisms. Med. Hypoth. 2020, 144, 110033. [Google Scholar] [CrossRef]
- Speed, N.; Blair, I.A. Cyclooxygenase- and lipoxygenase-mediated DNA damage. Can. Metast. Rev. 2011, 30, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Del Río, L.A. Role of Peroxisomes as a Source of Reactive Oxygen Species (ROS) Signaling Molecules. Subcell. Biochem. 2013, 69, 231–255. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, A.J.; Pinto, J.R.; Prokić, M.; Arsa, G.; Vasconsuelo, A. Oxidative Stress in Muscle Diseases: Current and Future Therapy 2019. Oxid. Med. Cell. Longev. 2020, 2020, 1–4. [Google Scholar] [CrossRef]
- Kowalska, M.; Wize, K.; Prendecki, M.; Lianeri, M.; Kozubski, W.; Dorszewska, J. Genetic Variants and Oxidative Stress in Alzheimer’s Disease. Curr. Alzh. Res. 2020, 17, 208–223. [Google Scholar] [CrossRef]
- Pearson, J.N.; Patel, M. The role of oxidative stress in organophosphate and nerve agent toxicity. Ann. N. Y Acad. Sci. 2016, 1378, 17–24. [Google Scholar] [CrossRef]
- Zhang, X.; Lee, M.D.; Wilson, C.; McCarron, J.G. Hydrogen peroxide depolarizes mitochondria and inhibits IP3-evoked Ca2+ release in the endothelium of intact arteries. Cell Calc. 2019, 84, 102108. [Google Scholar] [CrossRef]
- Folz, R.J.; Crapo, J.D. Extracellular Superoxide Dismutase (SOD3): Tissue-Specific Expression, Genomic Characterization, and Computer-Assisted Sequence Analysis of the Human EC SOD Gene. Genomics 1994, 22, 162–171. [Google Scholar] [CrossRef]
- Fisher, A.B. Redox Signaling Across Cell Membranes. Antiox. Redox Signal. 2009, 11, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Ind. J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.L.; Cox, M.M. Principles of Biochemistry, 7th ed.; W.H. Freeman & Co.: New York, NY, USA, 2017. [Google Scholar]
- Lloyd, D.R.; Phillips, D.H. Oxidative DNA damage mediated by copper (II), iron (II) and nickel (II) Fenton reactions: Evidence for site-specific mechanisms in the formation of double-strand breaks, 8-hydroxydeoxyguanosine and putative intrastrand cross-links. Mutat. Res. Mol. Mech. Mutagen. 1999, 424, 23–36. [Google Scholar] [CrossRef]
- Kushnir, A.; Marks, A.R. The Ryanodine Receptor in Cardiac Physiology and Disease. Adv. Pharmacol. 2010, 59, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncharov, N.V.; Nadeev, A.D.; Jenkins, R.O.; Avdonin, P.V. Markers and Biomarkers of Endothelium: When Something Is Rotten in the State. Oxid. Med. Cell. Longev. 2017, 2017, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P.; Rahman, H.S. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, X.G.; Zhu, J.-H.; Cheng, W.-H.; Bao, Y.; Ho, Y.-S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S.J. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [Green Version]
- Islam, O.; Bacchetti, T.; Ferretti, G. Alterations of Antioxidant Enzymes and Biomarkers of Nitro-oxidative Stress in Tissues of Bladder Cancer. Oxid. Med. Cell. Longev. 2019, 2019, 2730896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, S.S.; Singh, S.P.; Singhal, P.; Horne, D.; Singhal, J.; Awasthi, S. Antioxidant role of glutathione S-transferases: 4-Hydroxynonenal, a key molecule in stress-mediated signaling. Toxicol. Appl. Pharmacol. 2015, 289, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, Glutaredoxins, and Peroxiredoxins—Molecular Mechanisms and Health Significance: From Cofactors to Antioxidants to Redox Signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef]
- Rajković, M.G.; Rumora, L.; Barišić, K. The paraoxonase 1, 2 and 3 in humans. Biochem. Med. 2011, 21, 122–130. [Google Scholar] [CrossRef]
- Costa, L.G.; Giordano, G.; Cole, T.B.; Marsillach, J.; Furlong, C.E. Paraoxonase 1 (PON1) as a genetic determinant of susceptibility to organophosphate toxicity. Toxicology 2013, 307, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Penning, T.M.; Drury, J.E. Human aldo–keto reductases: Function, gene regulation, and single nucleotide polymorphisms. Arch. Biochem. Biophys. 2007, 464, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Glutathione Homeostasis and Functions: Potential Targets for Medical Interventions. J. Am. Aci. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassagnes, L.-E.; Chhour, M.; Pério, P.; Sudor, J.; Gayon, R.; Ferry, G.; Boutin, J.A.; Nepveu, F.; Reybier, K. Oxidative stress and neurodegeneration: The possible contribution of quinone reductase 2. Free Radic. Biol. Med. 2018, 120, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Brocker, C.; Koppaka, V.; Chen, Y.; Jackson, B.C.; Matsumoto, A.; Thompson, D.C.; Vasiliou, V. Aldehyde dehydrogenases in cellular responses to oxidative/electrophilicstress. Free Radic. Biol. Med. 2013, 56, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Chen, C.; Wang, H.; Rao, X.; Wang, F.; Duan, Q.; Chen, F.; Long, G.; Gong, W.; Zou, M.-H.; et al. Protective Effects of Acyl-coA Thioesterase 1 on Diabetic Heart via PPARα/PGC1α Signaling. PLoS ONE 2012, 7, e50376. [Google Scholar] [CrossRef]
- Franklin, M.P.; Sathyanarayan, A.; Mashek, D.G. Acyl-CoA Thioesterase 1 (ACOT1) Regulates PPARα to Couple Fatty Acid Flux With Oxidative Capacity During Fasting. Diabetes 2017, 66, 2112–2123. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damagein vivoand in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-Y.; Srinivasan, M.; Dasari, S.; Narvekar, P.; Samy, A.L.P.A.; Dontaraju, V.S.; Peng, L.; Anderson, G.L.; Munirathinam, G. Antioxidant Activity of Yichun Blue Honeysuckle (YBHS) Berry Counteracts CCl4-Induced Toxicity in Liver Injury Model of Mice. Antioxidants 2017, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musante, L.; Bruschi, M.; Candiano, G.; Petretto, A.; DiMasi, N.; Del Boccio, P.; Urbani, A.; Rialdi, G.; Ghiggeri, G.M. Characterization of oxidation end product of plasma albumin ‘in vivo’. Biochem. Biophys. Res. Commun. 2006, 349, 668–673. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef]
- Sozarukova, M.M.; Proskurnina, E.V.; Vladimirov, Y.A. Serum albumin as a source of and a target for free radicals in pathology. Target. ONCOTHERAPY 2016, 2016, 56–61. [Google Scholar] [CrossRef]
- Friedrichs, B. Th. Peters. Jr.: All about Albumin. Biochemistry, Genetics, and Medical Applications. XX and 432 pages, numerous figures and tables. Food/Nahrung 1997, 41, 382. [Google Scholar] [CrossRef]
- Lu, R.; Li, W.-W.; Katzir, A.; Raichlin, Y.; Yu, H.-Q.; Mizaikoff, B. Probing the secondary structure of bovine serum albumin during heat-induced denaturation using mid-infrared fiberoptic sensors. Analyst 2015, 140, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Ascenzi, P.; Bocedi, A.; Notari, S.; Fanali, G.; Fesce, R.; Fasano, M. Allosteric Modulation of Drug Binding to Human Serum Albumin. Mini-Rev. Med. Chem. 2006, 6, 483–489. [Google Scholar] [CrossRef]
- Ascenzi, P.; Fasano, M. Allostery in a monomeric protein: The case of human serum albumin. Biophys. Chem. 2010, 148, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Hein, K.L.; Kragh-Hansen, U.; Morth, J.P.; Jeppesen, M.D.; Otzen, D.; Møller, J.V.; Nissen, P. Crystallographic analysis reveals a unique lidocaine binding site on human serum albumin. J. Struct. Biol. 2010, 171, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Fasano, M.; Curry, S.; Terreno, E.; Galliano, M.; Fanali, G.; Narciso, P.; Notari, S.; Ascenzi, P. The extraordinary ligand binding properties of human serum albumin. IUBMB Life 2005, 57, 787–796. [Google Scholar] [CrossRef]
- Sudlow, G.; Birkett, D.J.; Wade, D.N. The characterization of two specific drug binding sites on human serum albumin. Mol. Pharmacol. 1975, 11, 824–832. [Google Scholar]
- He, X.M.; Carter, D.C. Atomic structure and chemistry of human serum albumin. Nat. Cell Biol. 1992, 358, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Bujacz, A. Structures of bovine, equine and leporine serum albumin. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 1278–1289. [Google Scholar] [CrossRef]
- Bujacz, A.; Talaj, J.A.; Zielinski, K.; Pietrzyk, A.J.; Neumann, P. Crystal structures of serum albumins from domesticated ruminants and their complexes with 3,5-diiodosalicylic acid. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 896–909. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Yokomaku, K.; Kureishi, M.; Akiyama, M.; Kihira, K.; Komatsu, T. Artificial Blood for Dogs. Sci. Rep. 2016, 6, 36782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokomaku, K.; Akiyama, M.; Morita, Y.; Kihira, K.; Komatsu, T. Core–shell protein clusters comprising haemoglobin and recombinant feline serum albumin as an artificial O2 carrier for cats. J. Mater. Chem. B 2018, 6, 2417–2425. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.T.; Kuhlmann, M.; Hvam, M.L.; Howard, K.A. Albumin-based drug delivery: Harnessing nature to cure disease. Mol. Cell. Ther. 2016, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Bteich, M. An overview of albumin and alpha-1-acid glycoprotein main characteristics: Highlighting the roles of amino acids in binding kinetics and molecular interactions. Heliyon 2019, 5, 02879. [Google Scholar] [CrossRef] [Green Version]
- Taborskaya, K.I.; Belinskaia, M.A.; Avdonin, P.V.; Goncharov, N.V. Building a three-dimensional model of rat albumin molecule by homology modeling. J. Evol. Biochem. Physiol. 2017, 53, 384–393. [Google Scholar] [CrossRef]
- Vyas, V.K.; Ukawala, R.D.; Chintha, C.; Ghate, M. Homology modeling a fast tool for drug discovery: Current perspectives. Indian J. Pharm. Sci. 2012, 74, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zazeri, G.; Povinelli, A.P.R.; Lima, M.D.F.; Cornélio, M.L. Experimental Approaches and Computational Modeling of Rat Serum Albumin and Its Interaction with Piperine. Int. J. Mol. Sci. 2019, 20, 2856. [Google Scholar] [CrossRef] [Green Version]
- Belinskaia, M.A.; Taborskaya, K.I.; Avdonin, P.V.; Goncharov, N.V. Modulation of the albumin–paraoxon interaction sites by fatty acids: Analysis by the molecular modeling methods. Russ. J. Bioorganic Chem. 2017, 43, 359–367. [Google Scholar] [CrossRef]
- Belinskaia, M.A.; Shmurak, V.I.; Taborskaya, K.I.; Avdonin, P.P.; Goncharov, N.V. In silico analysis of paraoxon binding by human and bovine serum albumin. J. Evol. Biochem. Physiol. 2017, 53, 191–199. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Belinskaia, M.A.; Shmurak, V.I.; Terpilowski, M.A.; Jenkins, R.O.; Avdonin, P.V. Serum Albumin Binding and Esterase Activity: Mechanistic Interactions with Organophosphates. Molecules 2017, 22, 1201. [Google Scholar] [CrossRef] [Green Version]
- Belinskaia, D.A.; Terpilovskii, M.A.; Batalova, A.A.; Goncharov, N.V. Effect of Cys34 Oxidation State of Albumin on Its Interaction with Paraoxon according to Molecular Modeling Data. Russ. J. Bioorganic Chem. 2019, 45, 535–544. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Batalova, A.A.; Goncharov, N.V. Effect of the bovine serum albumin redox state on its interaction with paraoxon as determined by molecular modeling. J. Evol. Biochem. Physiol. 2020, 56, 376–379. (In Russian) [Google Scholar] [CrossRef]
- Halliwell, B. Albumin—An important extracellular antioxidant? Biochem. Pharmacol. 1988, 37, 569–571. [Google Scholar] [CrossRef]
- Bar-Or, D.; Rael, L.T.; Lau, E.P.; Rao, N.K.; Thomas, G.W.; Winkler, J.V.; Yukl, R.L.; Kingston, R.G.; Curtis, C. An Analog of the Human Albumin N-Terminus (Asp-Ala-His-Lys) Prevents Formation of Copper-Induced Reactive Oxygen Species. Biochem. Biophys. Res. Commun. 2001, 284, 856–862. [Google Scholar] [CrossRef]
- Hureau, C.; Eury, H.; Guillot, R.; Bijani, C.; Sayen, S.; Solari, P.-L.; Guillon, E.; Faller, P.; Dorlet, P. X-ray and Solution Structures of CuIIGHK and CuIIDAHK Complexes: Influence on Their Redox Properties. Chem.–A Eur. J. 2011, 17, 10151–10160. [Google Scholar] [CrossRef] [PubMed]
- Sendzik, M.; Pushie, M.J.; Stefaniak, E.; Haas, K.L. Structure and Affinity of Cu(I) Bound to Human Serum Albumin. Inorg. Chem. 2017, 56, 15057–15065. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Hider, R.C. Influence of non-enzymatic post-translation modifications on the ability of human serum albumin to bind iron. Biochim. et Biophys. Acta (BBA)–Prot. Proteom. 2009, 1794, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Gryzunov, Y.A. Properties of Albumin Binding Centers: A Method of Their Exploration in Biological Fluids and Its Trial for Evaluation of the Organism Status. Ph.D Thesis, Federal Research and Clinical Center of Physical-Chemical Medicine, Moscow, Russia, 2003. (In Russian). [Google Scholar]
- Iwao, Y.; Ishima, Y.; Yamada, J.; Noguchi, T.; Kragh-Hansen, U.; Mera, K.; Honda, D.; Suenaga, A.; Maruyama, T.; Otagiri, M. Quantitative evaluation of the role of cysteine and methionine residues in the antioxidant activity of human serum albumin using recombinant mutants. IUBMB Life 2012, 64, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Jarabak, R.; Westley, J. Localization of the sulfur-cyanolysis site of serum albumin to subdomain 3-ab. J. Biochem. Toxicol. 1991, 6, 65–70. [Google Scholar] [CrossRef]
- Mills, B.J.; Lang, C.A. Differential distribution of free and bound glutathione and cyst(e)ine in human blood. Biochem. Pharmacol. 1996, 52, 401–406. [Google Scholar] [CrossRef]
- Turell, L.; Radi, R.; Alvarez, B. The thiol pool in human plasma: The central contribution of albumin to redox processes. Free Radic. Biol. Med. 2013, 65, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taverna, M.; Marie, A.-L.; Mira, J.-P.; Guidet, B. Specific antioxidant properties of human serum albumin. Ann. Intensiv. Care 2013, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Turell, L.; Botti, H.; Carballal, S.; Radi, R.; Alvarez, B. Sulfenic acid—A key intermediate in albumin thiol oxidation. J. Chromatogr. B 2009, 877, 3384–3392. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Li, H.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Cys34 Adducts of Reactive Oxygen Species in Human Serum Albumin. Chem. Res. Toxicol. 2012, 25, 1633–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, L.; Botti, H.; Carballal, S.; Ferrer-Sueta, G.; Souza, J.M.; Duran, R.; Freeman, B.A.; Radi, R.; Alvarez, B. Reactivity of Sulfenic Acid in Human Serum Albumin. Biochemestry 2008, 47, 358–367. [Google Scholar] [CrossRef]
- Bocedi, A.; Cattani, G.; Stella, L.; Massoud, R.; Ricci, G. Thiol disulfide exchange reactions in human serum albumin: The apparent paradox of the redox transitions of Cys 34. FEBS J. 2018, 285, 3225–3237. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, F.; Shibata, T.; Uchida, K. A unique mechanism for thiolation of serum albumins by disulphide molecules. J. Biochem. 2019, 167, 165–171. [Google Scholar] [CrossRef]
- Nakashima, F.; Shibata, T.; Kamiya, K.; Yoshitake, J.; Kikuchi, R.; Matsushita, T.; Ishii, I.; Bastida, J.A.G.; Schneider, C.; Uchida, K. Structural and functional insights into S-thiolation of human serum albumins. Sci. Rep. 2018, 8, 932. [Google Scholar] [CrossRef] [Green Version]
- Kurooka, S.; Yoshimura, Y. Mercaptoethanol Release upon Interaction of S-Lauroylmercaptoethanol with Serum Albumin. J. Biochem. 1973, 74, 785–795. [Google Scholar] [CrossRef]
- Cha, M.-K.; Kim, I.-H. Glutathione-Linked Thiol Peroxidase Activity of Human Serum Albumin: A Possible Antioxidant Role of Serum Albumin in Blood Plasma. Biochem. Biophys. Res. Commun. 1996, 222, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Bao, Y.; Ridley, S.; Williamson, G. Phospholipid hydroperoxide cysteine peroxidase activity of human serum albumin. Biochem. J. 1999, 338, 723–728. [Google Scholar] [CrossRef]
- Lee, H.; Cha, M.-K.; Kim, I.-H. Activation of Thiol-Dependent Antioxidant Activity of Human Serum Albumin by Alkaline pH Is Due to the B-like Conformational Change. Arch. Biochem. Biophys. 2000, 380, 309–318. [Google Scholar] [CrossRef]
- Lee, H.; Kim, I.H. Thioredoxin-linked lipid hydroperoxide peroxidase activity of human serum albumin in the presence of palmitoyl coenzyme A. Free Radic. Biol. Med. 2001, 30, 327–333. [Google Scholar] [CrossRef]
- Blanco, R.A.; Ziegler, T.R.; Carlson, B.A.; Cheng, P.-Y.; Park, Y.; Cotsonis, G.A.; Accardi, C.J.; Jones, D.P. Diurnal variation in glutathione and cysteine redox states in human plasma. Am. J. Clin. Nutr. 2007, 86, 1016–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ursini, F.; Maiorino, M.; Roveri, A. Phospholipid hydroperoxide glutathione peroxidase (PHGPx): More than an antioxidant enzyme? Biomed. Environ. Sci. 1997, 10, 327–332. [Google Scholar] [PubMed]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Yoo, M.-H.; Gu, X.; Xu, X.-M.; Kim, J.-Y.; Carlson, B.A.; Patterson, A.D.; Cai, H.; Gladyshev, V.N.; Hatfield, D.L. Delineating the Role of Glutathione Peroxidase 4 in Protecting Cells Against Lipid Hydroperoxide Damage and in Alzheimer’s Disease. Antioxid. Red. Sig. 2010, 12, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.M.; Sabet, S.; Peng, D.-F.; Nouh, M.A.; El-Shinawi, M.; El-Rifai, W. Promoter Hypermethylation and Suppression of Glutathione Peroxidase 3 Are Associated with Inflammatory Breast Carcinogenesis. Oxid. Med. Cell. Longev. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, Z.; Yingji, S.; Kim, H.; Jin, R.; Renshu, L.; Lee, D.Y.; Roh, M.R.; Yang, S. Downregulation of glutathione peroxidase 3 is associated with lymph node metastasis and prognosis in cervical cancer. Oncol. Rep. 2014, 31, 2587–2592. [Google Scholar] [CrossRef] [Green Version]
- Zunszain, P.A.; Ghuman, J.; McDonagh, A.F.; Curry, S. Crystallographic Analysis of Human Serum Albumin Complexed with 4Z,15E-Bilirubin-IXα. J. Mol. Biol. 2008, 381, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, J.; Brodersen, R. Albumin-bilirubin binding mechanism. J. Biol. Chem. 1983, 258, 6319–6326. [Google Scholar] [PubMed]
- Uchida, H.; Hanano, M. Conformational Changes of Human Serum Albumin by Binding of Small Molecules. Chem. Pharm. Bull. 1974, 22, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danesh, N.; Sedighi, Z.N.; Beigoli, S.; Sharifi-Rad, A.; Saberi, M.R.; Chamani, J. Determining the binding site and binding affinity of estradiol to human serum albumin and holo-transferrin: Fluorescence spectroscopic, isothermal titration calorimetry and molecular modeling approaches. J. Biomol. Struct. Dyn. 2017, 36, 1747–1763. [Google Scholar] [CrossRef] [PubMed]
- Barzegar, A.; Moosavi-Movahedi, A.; Sattarahmady, N.; Hosseinpour-Faizi, M.; Aminbakhsh, M.; Ahmad, F.; Saboury, A.; Ganjali, M.R.; Norouzi, P. Spectroscopic Studies of the Effects of Glycation of Human Serum Albumin on L-Trp Binding. Prot. Pept. Lett. 2007, 14, 13–18. [Google Scholar] [CrossRef]
- Fitos, I.; Kardos, J.; Visy, J. Stereoselective kinetics of warfarin binding to human serum albumin: Effect of an allosteric interaction. Chirality 2002, 14, 442–448. [Google Scholar] [CrossRef]
- Bree, F.; Urien, S.; Nguyen, P.; Tillement, J.P.; Steiner, A.; Vallat-Molliet, C.; Testa, B.; Visy, J.; Simonyi, M. Human Serum Albumin Conformational Changes as Induced by Tenoxicam and Modified by Simultaneous Diazepam Binding. J. Pharm. Pharmacol. 1993, 45, 1050–1053. [Google Scholar] [CrossRef]
- Bertucci, C.; Nanni, B.; Raffaelli, A.; Salvadori, P. Chemical modification of human albumin at cys34 by ethacrynic acid: Structural characterisation and binding properties. J. Pharm. Biomed. Anal. 1998, 18, 127–136. [Google Scholar] [CrossRef]
- Anraku, M.; Yamasaki, K.; Maruyama, T.; Kragh-Hansen, U.; Otagiri, M. Effect of oxidative stress on the structure and function of human serum albumin. Pharm. Res. 2001, 18, 632–639. [Google Scholar] [CrossRef]
- Sakurama, K.; Nishi, K.; Chuang, V.T.G.; Hashimoto, M.; Yamasaki, K.; Otagiri, M. Effects of Oxidation of Human Serum Albumin on the Binding of Aripiprazole. Biol. Pharm. Bull. 2020, 43, 1023–1026. [Google Scholar] [CrossRef]
- Oettl, K.; Stauber, R.E. Physiological and pathological changes in the redox state of human serum albumin critically influence its binding properties. Br. J. Pharmacol. 2007, 151, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Suda, K.; Matsuyama, Y.; Era, S.; Soejima, A. Close relationship between redox state of human serum albumin and serum cysteine levels in non-diabetic CKD patients with various degrees of renal function. Clin. Nephrol. 2014, 82, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Klammt, S.; Mitzner, S.; Stange, J.; Brinkmann, B.; Drewelow, B.; Emmrich, J.; Liebe, S.; Schmidt, R. Albumin-binding function is reduced in patients with decompensated cirrhosis and correlates inversely with severity of liver disease assessed by model for end-stage liver disease. Eur. J. Gastroenterol. Hepatol. 2007, 19, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, K.; Tanaka, M.; Chuang, V.T.G.; Setoyama, H.; Watanabe, H.; Yamada, N.; Kubota, K.; Matsushita, K.; Yoshida, A.; Jinnouchi, H.; et al. Cys34-Cysteinylated Human Serum Albumin Is a Sensitive Plasma Marker in Oxidative Stress-Related Chronic Diseases. PLoS ONE 2014, 9, e85216. [Google Scholar] [CrossRef] [PubMed]
- Nolte, W.; Hartmann, H.; Ramadori, G. Glucose metabolism and liver cirrhosis. Exp. Clin. Endocrinol. Diab. 2009, 103, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Lotosh, N.Y.; Savel’Ev, S.V.; Selishcheva, A.A. Modification of albumin with different degrees of the oxidation of SH-groups in the reaction with glucose. Russ. J. Bioorganic Chem. 2016, 42, 624–630. [Google Scholar] [CrossRef]
- Van Der Vusse, G.J. Albumin as Fatty Acid Transporter. Drug Metab. Pharmacokinet. 2009, 24, 300–307. [Google Scholar] [CrossRef]
- Maciążek-Jurczyk, M.; Sułkowska, A. Spectroscopic analysis of the impact of oxidative stress on the structure of human serum albumin (HSA) in terms of its binding properties. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 136, 265–282. [Google Scholar] [CrossRef]
- Pieniazek, A.; Gwozdzinski, L.; Zbrog, Z.; Gwozdzinski, K. Alterations in conformational state of albumin in plasma in chronic hemodialyzed patients. PLoS ONE 2018, 13, e0192268. [Google Scholar] [CrossRef]
- Christodoulou, J.; Sadler, P.J.; Tucker, A. A New Structural Transition of Serum Albumin Dependent on the State of Cys34. Detection by 1H-NMR Spectroscopy. JBIC J. Biol. Inorg. Chem. 1994, 225, 363–368. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Terpilovskii, M.A.; Shmurak, V.I.; Belinskaya, D.A.; Avdonin, P.V. Comparative analysis of esterase and paraoxonase activities of different serum albumin species. J. Evol. Biochem. Physiol. 2017, 53, 271–281. [Google Scholar] [CrossRef]
- Del Coco, L.; Vergara, D.; De Matteis, S.; Mensà, E.; Sabbatinelli, J.; Prattichizzo, F.; Bonfigli, A.R.; Storci, G.; Bravaccini, S.; Pirini, F.; et al. NMR-Based Metabolomic Approach Tracks Potential Serum Biomarkers of Disease Progression in Patients with Type 2 Diabetes Mellitus. J. Clin. Med. 2019, 8, 720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beteinakis, S.; Papachristodoulou, A.; Gogou, G.; Katsikis, S.; Mikros, E.; Halabalaki, M. NMR-Based Metabolic Profiling of Edible Olives—Determination of Quality Parameters. Molecules 2020, 25, 3339. [Google Scholar] [CrossRef] [PubMed]
- Fulmer, G.R.; Miller, A.J.M.; Sherden, N.H.; Gottlieb, H.E.; Nudelman, A.; Stoltz, B.M.; Bercaw, J.E.; Goldberg, K.I. NMR Chemical Shifts of Trace Impurities: Common Laboratory Solvents, Organics, and Gases in Deuterated Solvents Relevant to the Organometallic Chemist. Organometallics 2010, 29, 2176–2179. [Google Scholar] [CrossRef] [Green Version]
- Saadat, N.; IglayReger, H.B.; Myers, M.G.; Bodary, P.; Gupta, S.V. Differences in metabolomic profiles of male db/db and s/s, leptin receptor mutant mice. Physiol. Genom. 2012, 44, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, A.; Lee, H.; You, Y.-A.; Jung, Y.; Park, S.; Kim, S.M.; Hwang, G.-S.; Kim, Y. Identification of Potential Biomarkers in the Cervicovaginal Fluid by Metabolic Profiling for Preterm Birth. Metabolites 2020, 10, 349. [Google Scholar] [CrossRef]
- Sadler, P.J.; Tucker, A. Proton NMR studies of bovine serum albumin. Assignment of spin systems. JBIC J. Biol. Inorg. Chem. 1992, 205, 631–643. [Google Scholar] [CrossRef]
- Feng, J.; Isern, N.G.; Burton, S.D.; Hu, J.Z. Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics. Metabolites 2013, 3, 1011–1035. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Jiao, Y.; Wang, Y.; Xu, L.; Wang, M.; Zhao, B.; Jia, L.; Pan, H.; Zhu, Y.; Gao, X.-M. Quantitative Profiling of Polar Metabolites in Herbal Medicine Injections for Multivariate Statistical Evaluation Based on Independence Principal Component Analysis. PLoS ONE 2014, 9, e105412. [Google Scholar] [CrossRef]
- Kostidis, S.; Addie, R.D.; Morreau, H.; Mayboroda, O.A.; Giera, M. Quantitative NMR analysis of intra- and extracellular metabolism of mammalian cells: A tutorial. Anal. Chim. Acta 2017, 980, 1–24. [Google Scholar] [CrossRef]
- Stewart, A.J.; Blindauer, C.A.; Berezenko, S.; Sleep, D.; Tooth, D.; Sadler, P.J. Role of Tyr84 in controlling the reactivity of Cys34 of human albumin. FEBS J. 2004, 272, 353–362. [Google Scholar] [CrossRef]
- Bonanata, J.; Turell, L.; Antmann, L.; Ferrer-Sueta, G.; Botasini, S.; Méndez, E.; Alvarez, B.; Coitiño, E.L. The thiol of human serum albumin: Acidity, microenvironment and mechanistic insights on its oxidation to sulfenic acid. Free Radic. Biol. Med. 2017, 108, 952–962. [Google Scholar] [CrossRef]
- Yang, Q.; Xiao, H.; Cai, J.; Xie, Z.; Wang, Z.; Jing, X. Nanoparticle mediated delivery of a GST inhibitor ethacrynic acid for sensitizing platinum based chemotherapy. RSC Adv. 2014, 4, 61124–61132. [Google Scholar] [CrossRef]
- Sugiki, T.; Kobayashi, N.; Fujiwara, T. Modern Technologies of Solution Nuclear Magnetic Resonance Spectroscopy for Three-dimensional Structure Determination of Proteins Open Avenues for Life Scientists. Comput. Struct. Biotechnol. J. 2017, 15, 328–339. [Google Scholar] [CrossRef]
- Balasubramanian, K.; Gupta, S.P. Quantum Molecular Dynamics, Topological, Group Theoretical and Graph Theoretical Studies of Protein-Protein Interactions. Curr. Top. Med. Chem. 2019, 19, 426–443. [Google Scholar] [CrossRef]
- Rondeau, P.; Bourdon, E. The glycation of albumin: Structural and functional impacts. Biochimie 2011, 93, 645–658. [Google Scholar] [CrossRef]
- Anguizola, J.; Matsuda, R.; Barnaby, O.S.; Hoy, K.; Wa, C.; DeBolt, E.; Koke, M.; Hage, D.S. Review: Glycation of human serum albumin. Clin. Chim. Acta 2013, 425, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Soboleva, A.; Mavropolo-Stolyarenko, G.; Karonova, T.; Thieme, D.; Hoehenwarter, W.; Ihling, C.; Stefanov, V.E.; Grishina, T.; Frolov, A. Multiple Glycation Sites in Blood Plasma Proteins as an Integrated Biomarker of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2019, 20, 2329. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Jin, L.; Chen, J.; Shi, M.; Shi, F.; Wang, M.; Li, D.; Xu, X.; Su, X.; Yin, X.; et al. Comprehensive Glycomic Analysis Reveals That Human Serum Albumin Glycation Specifically Affects the Pharmacokinetics and Efficacy of Different Anticoagulant Drugs in Diabetes. Diabetes 2020, 69, 760–770. [Google Scholar] [CrossRef]
- Bourdon, E.; Loreau, N.; Blache, D. Glucose and free radicals impair the antioxidant properties of serum albumin. FASEB J. 1999, 13, 233–244. [Google Scholar] [CrossRef]
- Chesne, S.; Rondeau, P.; Armenta, S.; Bourdon, E. Effects of oxidative modifications induced by the glycation of bovine serum albumin on its structure and on cultured adipose cells. Biochimie 2006, 88, 1467–1477. [Google Scholar] [CrossRef]
- Rondeau, P.; Singh, N.R.; Caillens, H.; Tallet, F.; Bourdon, E. Oxidative stresses induced by glycoxidized human or bovine serum albumin on human monocytes. Free Radic. Biol. Med. 2008, 45, 799–812. [Google Scholar] [CrossRef]
- Martínez, A.E.; Regazzoni, L.; Brioschi, M.; Gianazza, E.; Agostoni, P.; Aldini, G.; Banfi, C. Pro-oxidant and pro-inflammatory effects of glycated albumin on cardiomyocytes. Free Radic. Biol. Med. 2019, 144, 245–255. [Google Scholar] [CrossRef]
- Gryzunov, Y.; Arroyo, A.; Vigne, J.-L.; Zhao, Q.; Tyurin, V.A.; Hubel, C.A.; Gandley, R.E.; Vladimirov, Y.A.; Taylor, R.; Kagan, V.E. Binding of fatty acids facilitates oxidation of cysteine-34 and converts copper-albumin complexes from antioxidants to prooxidants. Arch. Biochem. Biophys. 2003, 413, 53–66. [Google Scholar] [CrossRef]
- Torres, M.J.; Turell, L.; Botti, H.; Antmann, L.; Carballal, S.; Ferrer-Sueta, G.; Radi, R.; Alvarez, B. Modulation of the reactivity of the thiol of human serum albumin and its sulfenic derivative by fatty acids. Arch. Biochem. Biophys. 2012, 521, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Pavićević, I.D.; Jovanović, V.B.; Takić, M.M.; Penezić, A.; Aćimović, J.M.; Mandić, L.M. Fatty acids binding to human serum albumin: Changes of reactivity and glycation level of Cysteine-34 free thiol group with methylglyoxal. Chem. Interactions 2014, 224, 42–50. [Google Scholar] [CrossRef]
- Takić, M.M.; Jovanović, V.B.; Pavićević, I.D.; Uzelac, T.; Aćimović, J.M.; Ristić-Medić, D.K.; Mandić, L.M. Binding of enterolactone and enterodiol to human serum albumin: Increase of cysteine-34 thiol group reactivity. Food Funct. 2016, 7, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Penezić, A.; Aćimović, J.M.; Pavićević, I.D.; Jovanović, V.B.; Takić, M.; Mandić, L.M. The interplay between copper(II), human serum albumin, fatty acids, and carbonylating agent interferes with Cys 34 thiol reactivity and copper binding. JBIC J. Biol. Inorg. Chem. 2018, 24, 61–70. [Google Scholar] [CrossRef]
- Litus, E.A.; Permyakov, E.A.; Uversky, V.N.; Permyakov, E.A. Intrinsically Disordered Regions in Serum Albumin: What Are They For? Cell Biophys. 2017, 76, 39–57. [Google Scholar] [CrossRef]
- Nasif, W.A.; Mukhtar, M.H.; El-Emshaty, H.M.; Alwazna, A.H. Redox State of Human Serum Albumin and Inflammatory Biomarkers in Hemodialysis Patients with Secondary Hyperparathyroidism During Oral Calcitriol Supplementation for Vitamin D. Open Med. Chem. J. 2018, 12, 98–110. [Google Scholar] [CrossRef]
- Rael, L.T.; Leonard, J.; Salottolo, K.; Bar-Or, R.; Bartt, R.E.; Wagner, J.C.; Bar-Or, D. Plasma Oxidized Albumin in Acute Ischemic Stroke Is Associated with Better Outcomes. Front. Neurol. 2019, 10, 709. [Google Scholar] [CrossRef] [PubMed]
- Ueno, S.; Hatano, T.; Okuzumi, A.; Saiki, S.; Oji, Y.; Mori, A.; Koinuma, T.; Fujimaki, M.; Takeshige-Amano, H.; Kondo, A.; et al. Nonmercaptalbumin as an oxidative stress marker in Parkinson’s and PARK2 disease. Ann. Clin. Transl. Neurol. 2020, 7, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Horrillo, R.; Ortiz, A.M.; Perez, A.; Mestre, A.; Ruiz, A.; Boada, M.; Grancha, S. Increased Albumin Oxidation in Cerebrospinal Fluid and Plasma from Alzheimer’s Disease Patients. J. Alzh. Dis. 2018, 63, 1395–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grounds, M.D.; Terrill, J.R.; Al-Mshhdani, B.A.; Duong, M.N.; Radley-Crabb, H.G.; Arthur, P.G. Biomarkers for Duchenne muscular dystrophy: Myonecrosis, inflammation and oxidative stress. Dis. Model. Mech. 2020, 13, dmm043638. [Google Scholar] [CrossRef] [PubMed]
- Tarhoni, M.H.; Lister, T.; Ray, D.E.; Carter, W.G. Albumin binding as a potential biomarker of exposure to moderately low levels of organophosphorus pesticides. Biomarkers 2008, 13, 343–363. [Google Scholar] [CrossRef]
- Li, B.; Ricordel, I.; Schopfer, L.M.; Baud, F.; Megarbane, B.; Nachon, F.; Masson, P.; Lockridge, O. Detection of Adduct on Tyrosine 411 of Albumin in Humans Poisoned by Dichlorvos. Toxicol. Sci. 2010, 116, 23–31. [Google Scholar] [CrossRef]
- Chu, S.; Baker, M.R.; Leong, G.; Letcher, R.J.; Gee, S.J.; Hammock, B.D.; Li, Q.X. Exploring adduct formation between human serum albumin and eleven organophosphate ester flame retardants and plasticizers using MALDI-TOF/TOF and LC-Q/TOF. Chemosphere 2017, 180, 169–177. [Google Scholar] [CrossRef]
- Kranawetvogl, A.; Siegert, M.; Eyer, F.; Thiermann, H.; John, H. Verification of organophosphorus pesticide poisoning: Detection of phosphorylated tyrosines and a cysteine-proline disulfide-adduct from human serum albumin after intoxication with dimethoate/omethoate. Toxicol. Lett. 2018, 299, 11–20. [Google Scholar] [CrossRef]
- John, H.; Siegert, M.; Eyer, F.; Worek, F.; Thiermann, H.; Kranawetvogl, A. Novel cysteine- and albumin-adduct biomarkers to prove human poisoning with the pesticide oxydemeton-S-methyl. Toxicol. Lett. 2018, 294, 122–134. [Google Scholar] [CrossRef]
- Fujii, R.; Ueyama, J.; Aoi, A.; Ichino, N.; Osakabe, K.; Sugimoto, K.; Suzuki, K.; Hamajima, N.; Wakai, K.; Kondo, T. Oxidized human serum albumin as a possible correlation factor for atherosclerosis in a rural Japanese population: The results of the Yakumo Study. Environ. Heal. Prev. Med. 2018, 23, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Violi, F.; Cangemi, R.; Romiti, G.F.; Ceccarelli, G.; Oliva, A.; Alessandri, F.; Pirro, M.; Pignatelli, P.; Lichtner, M.; Carraro, A.; et al. Is Albumin Predictor of Mortality in COVID-19? Antioxidants Redox Signal. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Ishima, Y.; Kinoshita, R.; Chuang, V.T.; Tasaka, N.; Matsuo, N.; Watanabe, H.; Shimizu, T.; Ishida, T.; Otagiri, M.; et al. A novel S-sulfhydrated human serum albumin preparation suppresses melanin synthesis. Redox Biol. 2017, 14, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Dureau, A.-F.; Hellé, S.; Betscha, C.; Senger, B.; Cremel, G.; Boulmedais, F.; Strub, J.-M.; Corti, A.; Meyer, N.; et al. A Pilot Study on Continuous Infusion of 4% Albumin in Critically Ill Patients. Crit. Care Explor. 2019, 1, e0044. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.V.; Terpilowski, M.A.; Kudryavtsev, I.V.; Serebryakova, M.K.; Belinskaia, D.A.; Sobolev, V.; Shmurak, V.I.; Korf, E.A.; Avdonin, P.V. The Rat (Rattus norvegicus) as a Model Object for Acute Organophosphate Poisoning. 2. A System Analysis of the Efficacy of Green Tea Extract in Preventing Delayed Effects of Poisoning. J. Evol. Biochem. Physiol. 2019, 55, 208–221. [Google Scholar] [CrossRef]
- Batalova, A.A.; Belinskaia, D.A.; Goncharov, N.V. Testing of polyphenols and fatty acids as modulators of albumin esterase activity towards organophosphates [in Russian]. In Proceedings of the Scientific Conference “Modern Trends in the Development of Health Care Technologies”, Moscow, Russia, 12–13 December 2019; pp. 428–434. [Google Scholar]
- Goncharov, N.; Orekhov, A.N.; Voitenko, N.; Ukolov, A.; Jenkins, R.; Avdonin, P. Organosulfur Compounds as Nutraceuticals. In Nutraceuticals; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 555–568. [Google Scholar]
- Koutakis, P.; Ismaeel, A.; Farmer, P.; Purcell, S.; Smith, R.S.; Eidson, J.L.; Bohannon, W.T. Oxidative stress and antioxidant treatment in patients with peripheral artery disease. Physiol. Rep. 2018, 6, e13650. [Google Scholar] [CrossRef]
- Barnes, P.J. Oxidative stress-based therapeutics in COPD. Redox Biol. 2020, 33, 101544. [Google Scholar] [CrossRef]
- Penislusshiyan, S.; Chitra, L.; Ancy, I.; Kumaradhas, P.; Palvannan, T. Novel antioxidant astaxanthin-s-allyl cysteine biconjugate diminished oxidative stress and mitochondrial dysfunction to triumph diabetes in rat model. Life Sci. 2020, 245, 117367. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Vovk, M.A.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin. Antioxidants 2020, 9, 966. https://doi.org/10.3390/antiox9100966
Belinskaia DA, Voronina PA, Shmurak VI, Vovk MA, Batalova AA, Jenkins RO, Goncharov NV. The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin. Antioxidants. 2020; 9(10):966. https://doi.org/10.3390/antiox9100966
Chicago/Turabian StyleBelinskaia, Daria A., Polina A. Voronina, Vladimir I. Shmurak, Mikhail A. Vovk, Anastasia A. Batalova, Richard O. Jenkins, and Nikolay V. Goncharov. 2020. "The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin" Antioxidants 9, no. 10: 966. https://doi.org/10.3390/antiox9100966