Circadian Rhythm in Adipose Tissue: Novel Antioxidant Target for Metabolic and Cardiovascular Diseases


Department of Pharmacology, Johannes Gutenberg University Medical Center, Langenbeckstr, 1, 55131 Mainz, Germany
*
Author to whom correspondence should be addressed.
Antioxidants 2020, 9(10), 968; https://doi.org/10.3390/antiox9100968
Submission received: 30 August 2020
/
Revised: 29 September 2020
/
Accepted: 3 October 2020
/
Published: 9 October 2020
(This article belongs to the Special Issue Antioxidants and Chronic Inflammation)
Abstract
:Obesity is a major risk factor for most metabolic and cardiovascular disorders. Adipose tissue is an important endocrine organ that modulates metabolic and cardiovascular health by secreting signaling molecules. Oxidative stress is a common mechanism associated with metabolic and cardiovascular complications including obesity, type 2 diabetes, and hypertension. Oxidative stress can cause adipose tissue dysfunction. Accumulating data from both humans and experimental animal models suggest that adipose tissue function and oxidative stress have an innate connection with the intrinsic biological clock. Circadian clock orchestrates biological processes in adjusting to daily environmental changes according to internal or external cues. Recent studies have identified the genes and molecular pathways exhibiting circadian expression patterns in adipose tissue. Disruption of the circadian rhythmicity has been suggested to augment oxidative stress and aberrate adipose tissue function and metabolism. Therefore, circadian machinery in the adipose tissue may be a novel therapeutic target for the prevention and treatment of metabolic and cardiovascular diseases. In this review, we summarize recent findings on circadian rhythm and oxidative stress in adipose tissue, dissect the key components that play a role in regulating the clock rhythm, oxidative stress and adipose tissue function, and discuss the potential use of antioxidant treatment on metabolic and cardiovascular diseases by targeting the adipose clock.
1. Introduction
Obesity is a major public health concern worldwide, and due to its growing frequency, has become a serious financial burden to many countries [1]. Obesity contributes to the pathogenesis of most metabolic and cardiovascular diseases, including diabetes and hypertension [2]. Obesity is characterized by the excess accumulation of fat in adipose tissue, which can result from unhealthy lifestyle such as irregular sleep/wake cycle and high snacking frequency [3]. Adipose tissue is a critical modulator of metabolic and cardiovascular health. There is an increasing consensus that recognize adipose tissue as an important endocrine organ that can alter the metabolism of other distal organs. This is accomplished by the production and secretion of adipocyte-derived cytokines (also known as adipokines). Adipokines have a wide range of signaling and metabolic effects, such as regulating insulin sensitivity, inflammation, and blood pressure [4]. Indeed, adipose function shows strong fluctuation throughout the day. For example, the transportation, storage and metabolism of triglycerides (TGs) or glucose are carried out in the different phases of the day [5]. Adipokine levels show robust oscillations throughout the day [6,7]. These rhythmic oscillations are not mere responses to external stimuli, but are also facilitated by the local clock system that coordinates physiology and external cues. Dysfunction of the adipose tissue could exert deleterious effects on the metabolic and cardiovascular systems.
Oxidative stress is a common pathophysiological event in metabolic and cardiovascular diseases, including obesity, diabetes, and hypertension. Oxidative stress occurs when there is a redox imbalance [8]. Under normal conditions, homeostatic reactive oxygen species (ROS) play important roles as secondary messengers in various intracellular signaling pathways including programmed cell death or necrosis, gene expression regulation, cell signaling cascades activation [9], as well as in both innate and adaptive immune responses [10], and body weight control by modulating satiety and hunger behavior [11]. Oxidative stress can cause adipose tissue dysfunction by stimulating preadipocyte proliferation, adipogenesis and chronic inflammation, which leads to obesity [12]. In obesity, systematic oxidative stress can also be induced by oxidative phosphorylation in mitochondria, superoxide generation from nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX), uncoupled endothelial nitric oxide synthase (eNOS) [13] and reduced antioxidant defense [8,14]. Interestingly, the production of antioxidants and expression of anti-oxidative enzymes have been reported to be regulated by the circadian rhythm [15]. Therefore, oxidative stress seems to have an innate connection with the circadian clock.
Many organisms have developed an intrinsic mechanism responding to the environmental light-dark cycle. This core timing system ha’s an intimate relationship with many important physiologies and pathways. Circadian (Latin “circa” meaning around and “diem” meaning a day) clock orchestrates biological processes in adjusting to daily environmental changes. Circadian rhythm generally oscillates in a daily cycle around 24 hours light and dark period in response to abiotic and biotic factors [16]. The circadian clock is composed of molecular clock networks in the core and peripheral tissues. Core circadian clock is located in the hypothalamic suprachiasmatic nuclei (SCN), while peripheral clocks are found in other tissues including the kidney, liver, blood vessels and adipose tissues [17]. The intrinsic circadian clock is autonomous and self-sustaining by the negative feedback loop of molecular clock control. In addition, internal or external factors, which are known as “zeitgeber” (German for time giver), may cue the endogenous circadian cycle and modulate the clock from the molecular to behavioral level [18,19].
Desynchronization of the clock or the misalignment between circadian rhythm and metabolism contributes to the pathogenesis of chronic metabolic and cardiovascular diseases [20]. In experimental models, knockout and mutation in clock genes that lead to the disruption of circadian rhythmicity have revealed the tight link between the circadian clock, adipose tissue function and metabolism [21]. It is estimated that nearly half of all murine genes are expressed in circadian rhythm in at least one tissue of the body [22]. Recently, a circadian transcriptome study in human adipose tissue has shown that 837 transcripts exhibit circadian expression profiles with clear separation of transcripts peaking in the morning (258 probes) and evening (579 probes) [23], suggesting that circadian rhythm in adipose tissue could be a novel therapeutic target for metabolic and cardiovascular diseases.
In this review, we summarize current knowledge on circadian rhythm, adipose tissue function, ROS production and oxidative stress in metabolic and cardiovascular diseases; dissect the key components that play a dual role in regulating the clock rhythm, as well as adipose tissue metabolism, and link adipose circadian rhythms with oxidative stress; and discuss the potential antioxidant treatment of metabolic and cardiovascular diseases by targeting circadian clock in adipose tissue.
2. Role of Adipose Tissue in Metabolic and Cardiovascular Health
Adipose tissue has been regarded as a reservoir for energy storage, while currently known as an endocrine organ that regulates energy metabolism and influences systemic metabolic homeostasis through adipokines [21]. In mammals, at least two types of adipose tissue have been described, the white adipose tissue (WAT) and the brown adipose tissue (BAT). These two adipose tissues are originated from different stem cells, and they have distinct localizations, gene expression profiles and functions [24]. In addition, beige adipose tissue is recently described and is located within WAT but shares similar features with BAT [24]. The balance between WAT, BAT, and beige adipocytes is important for maintaining overall energy and lipid metabolism.
2.1. White Adipose Tissue
WAT is the most abundant type of adipose tissue and the main energy reservoir in mammals. White adipocytes are characterized by containing a single large unilocular lipid droplet and only a few mitochondria. The primary function of WAT is to synthesize and store excess dietary energy as triacylglycerols (TAGs), and hydrolyze TAG to generate fatty acids during energy deprivation [25]. WAT is also responsible for the secretion of a huge number of hormones and adipokines that regulate energy intake, metabolism and insulin resistance [26,27]. WAT converts excess dietary fats and carbohydrates from the circulation to TAGs by lipogenesis [28]. On the other hand, TAGs are broken down into glycerol and free fatty acids (FFAs) by lipolysis to release and supply the needs of distal organs [28].
2.2. Brown Adipose Tissue
Brown adipocytes are characterized by containing multilocular lipid droplets and high mitochondrial density which facilitates thermogenesis [29]. BAT is developmentally different from WAT. BAT originates from early paired box protein 7 (Pax7)+ and myogenic factor 5 (Myf5)+ embryonic progenitor cells or postnatal myoblasts, while late Pax7 expression is restricted to the skeletal muscle lineage [30,31]. In addition, PR domain containing 16 (Prdm16) is also a known lineage determining factor and transcription cofactor for the development of brown adipocytes [31]. The major activator of BAT is the sympathetic nervous system, signaled by norepinephrine [32]. Upon stimulation, such as cold exposure, BAT is activated to promote non-shivering thermogenesis via uncoupling protein 1 (UCP1), a thermogenic mitochondrial protein that uncouples electron transport from adenosine triphosphate (ATP) production [29]. In addition to its primary role in maintaining body temperature, BAT is also important for energy metabolism, insulin sensitivity, and lipid metabolism due to its high metabolic rate [33,34].
2.3. Beige Cells and Browning
Recent studies have identified beige adipocytes, which are mitochondria rich and express UCP1, within WAT [35,36]. Beige adipocyte can be considered phenotypically as a fat cell that possesses characteristics between those of the WAT and BAT [37], which indicate that they are energy accumulators that possess thermogenic capacity to dissipate heat through UCP1 [38].
The biogenesis of beige adipocytes or the white-to-brown transition of adipocytes is called browning/beiging, which is a temporary adaptive response to external environmental signals. Browning can be stimulated by cold exposure, adrenergic stimulation and treatments with peroxisome proliferation-activated receptor gamma (PPARγ) agonists [39,40], fibroblast growth factor 21 (FGF21) [41], atrial natriuretic peptide (ANP) [42], and bone morphogenetic proteins (BMP) [43]. Browning of adipose tissue has been shown to be beneficial in preventing metabolic and cardiovascular diseases [44]. In addition, cold exposure can stimulate glucose uptake and TG clearance in adipose tissue, which may contribute to the modulation of oxidative stress in adipose tissue [45,46]. Interestingly, FGF21 has recently been shown to have both antihypertrophic and cardioprotective actions [47,48], and has been considered as a promising new therapy target obesity and associated diseases, by activating BAT and by acting on WAT and the liver [49]. Remarkably, FGF21 can regulate circadian behavior and metabolism by acting on the nervous system [50].
2.4. Perivascular Adipose Tissue
Apart from the three types of adipose tissue mentioned above, perivascular adipose tissue (PVAT) is also a critical modulator of cardiovascular health [51,52]. PVAT is the ectopic fat depot surrounding most vasculature including large arteries and veins, small and resistance vessels, and skeletal muscle microvessels [52]. This intimate connection between the vascular system and PVAT highlights the importance of PVAT. PVAT controls vascular tone on vessels through releasing various PVAT-derived relaxing factors (PVRFs) [53] and PVAT-derived contracting factors (PVCFs) [54,55]. Moreover, these PVAT-derived factors closely and directly affect vascular homeostatic processes, including vascular inflammation and oxidative stress, vascular tone, smooth muscle proliferation and migration and vascular remodeling [51,56,57].
To date, PVAT has been reported to contain both WAT and BAT, while the white-to-brown ratio is different across the vascular bed. For example, PVATs surrounding larger blood vessels are more BAT-like, whereas those surrounding smaller blood vessels are WAT-like [56,58,59,60]. It is suggested that the adipocytes in PVAT may share a common smooth muscle protein 22 alpha (SM22α+) precursors with vascular smooth muscle cells (VSMCs) [61]. During obesity and aging, the gradual shift into WAT-like characteristics of PVAT is associated with the alteration of secretome profile, which may lead to vascular dysfunction, arterial remodeling and increase in blood pressure [62,63]. Apart from cold exposure, exercise training has been shown to induce a BAT-like shift and thermogenic response, which is associated with enhanced eNOS expression and reduced oxidative stress in PVAT [64].
2.5. Adipokines
Adipose tissue produces and secretes a variety of bioactive molecules, adipokines, which signals to nearby or remote target organs. Adipose tissue accumulation and altered adipokine profile are linked to chronic inflammation and metabolic and cardiovascular disorders [65]. The primary adipokines that play a role in inflammation, metabolic and cardiovascular diseases include adiponectin, leptin, resistin, visfatin, tumor necrosis factor alpha (TNF-α), interleukin-6(IL-6), plasminogen activator inhibitor-1 (PAI-1), and vascular endothelial growth factor (VEGF) [66,67]. In general, adipokine levels are positively correlated with fat mass, and the functions of adipokines are also related to their site of production. For example, BAT-derived IL-6, an adipokine that are upregulated by proinflammatory stimulation, has been shown to mediate glucose metabolism and energy balance [34]. PVAT-derived adiponectin is an important contributor to vascular function, partly by enhancing eNOS phosphorylation in the endothelium [68]. Notably, most hormones, including adipokines, display daily fluctuation of secretion during the day-night cycle. In addition to the altered adipokine profile, the disruption of this rhythmic secretion of adipokines is also an aggravating factor for the development of metabolic and cardiovascular diseases [69].
3. Circadian Rhythm in Adipose Tissue
The intrinsic biological clock of around 24-h cycle that orchestrates the body physiology is controlled by a multi-oscillatory network composed by circadian clocks in most cells and tissues, including the adipose tissue [70]. Clinical studies have reported the circadian pattern and diurnal variation of physiological and pathological cardiovascular events such as heart rate, blood pressure and endothelial function [71,72,73,74,75].
The central circadian is located in the SCN at the anterior hypothalamus, while peripheral clocks are found in the liver, intestine, heart, blood vessel wall and adipose tissue [17,76,77]. In the SCN, photic information transmitted from the retina synchronizes neurons to coordinate circadian outputs [78]. The central clock regulates peripheral clock directly by neuronal or hormonal signaling or indirectly by driving appetite, blood pressure, and body temperature, in order to coordinate circadian gene expression [79,80].
The local circadian clocks present in adipocytes regulate many essential adipose tissue processes including lipolysis, adipogenesis, inflammation, browning, thermogenesis, as well as the expression and secretion of adipokines [81]. Therefore, circadian disruption may alter the physiology of adipose tissue and thereby affect whole body energy homeostasis. The rhythmic expression of circadian genes in different white adipose depots in rodents [5,82,83] and humans [84,85,86] have been revealed by both in vitro and in vivo studies. In murine adipose tissue, approximately 10% of genes are expressed in circadian rhythm, which underlines the circadian control over the adipose tissue function [83].
3.1. The Molecular Clock Mechanism in Adipose Tissue
At the molecular level, circadian clock in the adipose tissue is based on interlocked transcription–translation feedback loops of clock genes and proteins [87]. These circadian proteins include important transcription factors brain and muscle aryl hydrocarbon receptor nuclear translocator-like protein 1 (BMAL1 a.k.a. ARNTL) and circadian locomotor output cycles kaput (CLOCK), and transcriptional modulator families of Period 1/2/3 (PER1/2/3) and Cryptochrome ½ (CRY1/2) [87,88]. The molecular clockwork in adipose tissue relies on the dimerization of BMAL1 and CLOCK proteins, whereas the heterodimer of CLOCK:BMAL1 binds to E-box sequence (5′-CACGTG-3′) in the promoters and activates the transcription of other circadian genes including Per1–3 and Cry1/2 [17]. When PERs and CRYs are expressed in certain amount in the cytoplasm, these proteins dimerize (PERs:CRYs) and translocate into the nucleus, which leads to the inhibition of CLOCK:BMAL1-mediated transcription [17,89].
There are also reinforcing loops that are composed by the circadian nuclear receptors, reverse ERB (REV-ERB α/β) and retinoic acid receptor-related orphan receptors (RORα/β/γ) that control the rhythmic transcription of Bmal1 and Clock. REV-ERBα negatively regulates Bmal1 and Clock expression [90], whereas RORα and RORγ positively regulate Bmal1 and Clock expression via ROR response elements at the promotor regions [91,92]. All the expression patterns of these clock genes exhibit a 24-hour oscillation in adipose tissue [93]. In human adipose tissue, the expression of BMAL1 and CLOCK peaks in the late evening, while the expression of PERs, CRY2, and REV-ERBα peak around late morning [85]. In addition, post-translational modifications also contribute to the regulation of the circadian clock [70,94].
Apart from the interlocked transcription–translation feedback loop of the clock genes, the oscillation of the circadian clock also leads to rhythmic expression of clock-controlled genes (CCGs) through activation of circadian promoter elements including E-boxes, D-boxes, and ROR response elements [85]. While the same clock machineries are found in most cells, the rhythmic expression of these CCGs are tissue-specific or even cell-type-specific [70] (Figure 1).
3.2. Clock Genes in Adipose Function
Apart from controlling the circadian machinery, circadian clock genes also have important functions in the metabolic and cardiovascular system. In different animal models, circadian clock disruptions generally lead to reduced lifespan, accelerated aging and dramatic effects on body weight gain, adiposity and TG metabolism [95,96,97].
ClockΔ19 and Bmal1−/− circadian mutant mice display low and nonrhythmic blood FFA and glycerol levels together with decreased lipolysis rates and increased sensitivity to fasting [5]. Circadian disruption promotes the accumulation of TGs in WAT which leads to increased adiposity and adipocyte hypertrophy [5]. CLOCK and BMAL are also critical in maintaining glucose homeostasis while Clock-mutant mice display phenotype of cardiovascular and metabolic diseases such as obesity and hypertension [7,98]. Deficiency of Bmal1 leads to impaired adaptive vascular remodeling and increased collagen deposition in the medial layer of blood vessels in mice [99]. Moreover, transcriptome analysis of tissue-specific ClockΔ19 mutant mice shows altered regulation of at least 660 genes due to impaired WAT clocks, of which 26% are repressed, while the remaining 74% are induced [95]. A part of these differentially expressed genes is involved in lipogenesis, including those encodes for fatty acid transporters, fatty acid binding protein 9 (FABP9) and glycerol kinase 2 (GK2) [95]. Overexpression of BMAL1 in adipocytes has been shown to increase lipid synthesis activity and promote several factors associated with lipogenesis in vitro [100]. BMAL1 modulates adipogenesis via wingless-type MMTV integration site family (Wnt) signaling in WAT [101], while it exerts transcriptional control on the transforming growth factor-β (TGF-β) pathway in BAT. Either global ablation or adipocyte-selective inactivation of BMAL1 increases the mass of BAT and cold tolerance [102]. BMAL1 inhibits brown fat formation through direct transcriptional control of TGF-β and bone morphogenetic protein (BMP), which are known to suppress [103] and drive [104] brown adipocyte differentiation, respectively. Activation of TGF-β/Smad3 or inhibition of BMP pathways normalizes the enhanced differentiation in Bmal1-deficient brown adipocytes [102]. Interesting, antioxidant N-acetyl-L-cysteine can ameliorate symptoms of premature aging in Bmal1-deficient mice [105].
REV-ERB and ROR families are important component that link the core clock mechanism with lipid metabolism. REV-ERBs and RORs have been shown to be responsible for adipocyte differentiation [106], lipogenesis, and lipid storage [107,108]. Rev-erbα-deficient mice show increased adiposity on both normal chow and high-fat diet (HFD) as a result of increased fat uptake by adipose tissue [109]. REV-ERB agonist treatment has been shown to reduce fat mass and alleviate dyslipidemia and hyperglycemia in obese mice [110]. REV-ERB agonist can suppress orexinergic gene expression, while Rev-erbβ-deficient mice have increased orexinergic transcripts, suggesting the involvement of REV-ERB in regulating appetite [111]. On the other hand, REV-ERBα appears to have an opposite role on adiposity in BAT. Deletion of Rev-erbα results in an improved thermogenic response to cold challenge, while downregulation of Rev-erbα leads to the induction of UCP1 expression [112]. As opposed to BMAL1, REV-ERBα inhibits key components of the TGF-β signaling pathway [113], thereby promotes BAT development and adipogenesis.
Per2−/− mice displays altered lipid metabolism, with drastic reduction of total TAGs and non-esterified fatty acids [114]. PER2 represses PPARγ-mediated transcriptions, thereby altering the expression of genes involved in adipogenesis, insulin sensitivity and inflammatory response [114]. Moreover, lack of PER2 promotes adipocyte differentiation in cultured fibroblasts [114]. PER3 has been reported to modulate adipogenesis by regulating Krüppel-like factor 15 (KLF15) expression, while deletion of Per3 promotes adipogenesis in vivo [115]. Cry1−/−Cry2−/− mice have reduced body weight and WAT due to higher energy expenditure and heat production compared with wild-type controls under normal chow [116]. Cry1−/−Cry2−/− mice also have an increased sensitivity to HFD-induced obesity, which is associated with increased insulin secretion, elevated lipogenesis and TG accumulation in WAT [117] and augmented proinflammatory cytokine levels [118]. In addition, meta-analysis indicates that genetic variants at Cry1 are associated with insulin resistance and diabetes risk in human [119].
To date, only a few studies have reported the phenotypes and effects on adipose tissue function and oxidative stress in adipose tissue-specific clock gene knockout mice (Table 1). Adipocyte-specific deletion of Bmal1 results in obesity in mice with a shift in diurnal rhythm of food intake and loss of circadian variation in plasma TG and glucose levels [120]. Deletion of the adipocyte Bmal1 is associated with a reduced number of polyunsaturated fatty acids in adipocyte triglycerides, and loss of rhythmic expression of clock and clock-output genes in both BAT and WAT [120]. In addition, adipocyte-specific ClockΔ19 mutation increases body weight and fat mass in mice [121]. The young mortality rates of these mice are increased and the rates of glucose tolerance are reduced compared to wild type mice [121]. Moreover, brown adipocyte-selective Bmal1-deficient mice have reduced blood pressure during resting phase, associated with reduced PVAT-induced vasoconstriction [122]. Indeed, angiotensinogen mRNA and angiotensin II levels in PVAT of brown adipocyte-selective Bmal1-deficient mice are significantly reduced [122]. These studies suggest that clock aberration specifically in adipose tissue can result in significant alterations of the cardiovascular and metabolic health. Circadian clock genes in adipose tissues are important in maintaining the robust relationship between circadian rhythm and metabolic and cardiovascular health. However, most of the observations currently made are based on animal models with global ablation or mutants of circadian clock genes, the contribution of these circadian clock genes in adipose tissue function warrants further studies by using adipose tissue-specific knockout/mutant mice.
4. Oxidative Stress and Circadian Rhythm
Generation of ROS is a normal physiological process in all aerobic organisms. Under normal conditions, deleterious ROS are mostly removed by the cellular antioxidant systems. Oxidative stress occurs when the production of oxidant molecules, such as superoxide (•O−2) and hydrogen peroxide (H2O2), exceeds the capacity of antioxidants, including catalase, glutathione peroxidase (GPx), superoxide dismutase (SOD), NAPDH and peroxisomes to defend against these insults. Excessive ROS are known to cause lipid peroxidation and oxidative modifications of proteins and nucleic acids that cause metabolic and cardiovascular complications [123,124,125,126].
In obese animals and patients, adipocytes become hypertrophic leading to hypoxia [52]. The expression of the key mediator of hypoxia, hypoxia-inducible factor alpha (HIF-1α) is augmented in the adipose tissue of obese subjects [127]. HIF-1α stimulates the production of inflammatory mediator, such as TNF-α and IL-6, and suppresses the expression of adiponectin [128]. HIF-1α is interconnected to the circadian clock and has been shown to regulate clock gene expression by direct promotor binding, while CRY1/2 stabilize HIF-1α in response to hypoxia [129,130]. This suggests a bidirectional interaction between hypoxia signaling pathway and the circadian clock. Obesity-induced adipocyte hypertrophy is associated with reduced insulin sensitivity, increased oxidative stress, and abnormal adipokine secretory profile, including an elevated array of other pro-inflammatory factors [IL-1β, IL-8, resistin and monocyte chemoattractant protein 1 (MCP1)] and a reduced array of anti-inflammatory factors (IL-10, adiponectin and FGF21) [131,132]. These inflamed WATs become severely dysfunctional and fail to store surplus energy, which lead to ectopic fat deposition in other metabolic tissues such as liver and skeletal muscle [133]. Therefore, obesity engenders nutrient stress that alters mitochondrial function, while the elevated mitochondrial substrate load augments electron transport chain activity and ROS production [134]. The increased oxidative stress in adipose tissue alters the secretion of adipokines and leads to metabolic and cardiovascular complications [51,134]. As mentioned, secretion of adipokines is in circadian rhythm [135]. Increased secretion of IL-6 is associated with excessive sleeping at day cycle and fatigue, while a reduction of IL-6 secretion is associated with sufficient sleeping at night cycle, whereas both TNF-α and IL-6 secretions have been reported to increase in sleep apnea independently of obesity [136]. Indeed, the susceptibility of wild-type mice to HFD-induced obesity depends on the time of feeding [137]. Mice fed with HFD late in their activity period are more susceptible to induce obesity while mice fed with HFD early in the activity period are resistant to obesity [137]. This suggests that the pathogenesis of metabolic and cardiovascular diseases may not solely be due to excessive nutrient-induced obesity, while circadian dysregulation of metabolism and oxidative stress may be the key components that drives obesity.
The cellular redox state in plants and animals has been known to oscillate over circadian time [138]. The expression of major antioxidant enzymes is rhythmic, but there are high divergency of amplitude, phasing, and presumable protective value among different tissues [15]. Bmal1−/− mice are reported to have increased ROS levels in different tissues and show symptoms of premature aging [97]. Apart from obesity, people with sleeping disorder or night shift workers also show augmented levels of oxidative stress damages due to a reduced antioxidant defenses [139,140], which may contribute to the increased risk of metabolic and cardiovascular diseases. These results suggest an intrinsic relationship between oxidative stress, circadian rhythm, and adipose tissue function.
Recently, considerable evidence has been shown to support the hypothesis that oxidative stress and circadian rhythm are interrelated, as the circadian clock regulates the rhythmic oscillation of cellular redox function, while changes in redox state in the cell can influence the clock machinery [141] (Figure 2). Indeed, oxidative stress can drive the expression of REV-ERBα, which in turn regulates the expression of antioxidant transcription factor forkhead box protein O-1 (FOXO1) as well as stimulates autophagy and mitochondrial biogenesis [142]. Circadian oscillation of H2O2 level is observed in mammalian cells and mouse liver, and can directly regulate CLOCK circadian function via cysteine oxidation [143]. Moreover, nicotinamide adenine dinucleotide (NAD) has been recently recognized as a “metabolic oscillator” that displays circadian oscillations and is the missing-link between energy metabolism and circadian control [144,145,146]. Inhibition of catalase results in a lengthened period of the Per1 gene oscillation and hypoxia leads to a higher amplitude oscillation of NAD+/NADH ratio and a highly oxidized cellular environment during the night [147]. The DNA binding activity of CLOCK:BMAL1 heterodimer is dependent on the cellular redox status (NADH/NAD and NADPH/NADP ratios) [148], while NADPH/NADP ratio oscillates in a circadian manner [147]. Pentose phosphate pathway (PPP) can regulate the circadian clock via NADPH metabolism [149]. Inhibition of PPP has been shown to increase oxidative stress, lengthen circadian period, activate nuclear factor erythroid 2-related factor 2 (NRF2) pathway, increase CLOCK:BMAL1 DNA binding and alter CCGs expression in vitro [149]. In addition, NRF2 pathway appears to regulate clock function, as Nrf−/− cells have flattened clock gene expression and blunted Per2 circadian expression [150].
Recently, thermogenic effect of ROS has been reported. Increased adipocyte ROS levels and oxidative stress can promote thermogenesis in adipocyte tissue [151,152]. Indeed, activation of thermogenesis in murine BAT by either cold exposure or β-adrenergic stimulation is associated with an elevated levels of mitochondrial superoxide, H2O2, and lipid hydroperoxides [152]. The thermogenic action of mitochondrial ROS is mediated through protein cysteine modification [152]. Moreover, the substantial and selective accumulation of succinate, a mitochondrial metabolite, may be a potent source for thermogenic ROS in brown and beige fat [151].
Another evidence that links circadian rhythm and oxidative stress is the experimental results from the use of antioxidants to entrain the clock. Melatonin is a well-known hormone involved in circadian rhythm regulation and contributed to protection against oxidative stress [153]. Indeed, melatonin is a direct free radical scavenger, which has anti-inflammatory and antioxidant effects, due to its electron-rich aromatic indole ring [154,155]. Recently, melatonin treatment has been shown to stimulate thermogenesis of BAT in response to cold exposure via activating citrate synthase and respiratory chain complexes I and IV in diabetic and obese rats [156]. Moreover, melatonin can promote the differentiation of white adipocytes into the beige adipocytes and improve adipose tissue function [157]. These experiments suggest that potential use of antioxidants to entrain the circadian clock.
4.1. Effects of Time-Restricted Feeding and Intermittent Fasting on Circadian Rhythm and Metabolism
Feeding and fasting patterns have been shown to drive daily rhythms in the activities of key energy regulators including adenosine 5′-monophosphate–activated protein kinase (AMPK), cAMP-response element binding protein (CREB) and protein kinase B (AKT) [158,159], which suggest a clear connection between circadian rhythm and metabolism. Constant light causes disruptions in the rhythmic oscillation of plasma levels of glucose and TG in mice and eliminations of the circadian expression of genes involved in lipid metabolism in WAT, which can be restored by time-restricted feeding to various degrees [160]. This suggests that food intake is an important zeitgeber, which may be stronger than light-dark cycle, that regulates lipid metabolism in WAT. Feeding and restrictive meal timing have been shown to modulate circadian rhythm and generate pronounced phase shifts in circadian gene expression in different peripheral tissues [161,162]. Temporally restricted feeding in mice alters the circadian expression profile in BAT, inguinal WAT and epididymal WAT [83]. In rat, either wrong time restricted feeding or high-fat-high-sugar diet disturbs the peripheral clock rhythmicity, and alters Ucp1 and PPARγ coactivator 1 alpha (Pgc-1α) expression in BAT [163]. In mice, time restricted feeding attenuates obesity, hyperinsulinemia and inflammation, associated with improvements in circadian clock oscillation, and mTOR and AMPK pathway function [158]. In a recent study, time restricted feeding has been shown to prevent obesity and improve metabolic function in whole-body Cry1;Cry2 knockout and in liver-specific Bmal1 and Rev-erba/b knockout mice [164].
Intermittent fasting is an dietary pattern in which minimal calories are consumed 1 to 4 days per week, followed by ad libitum diet on the remaining days [165], and has been shown to reduce the risk for metabolic diseases in human [166,167,168]. Intermittent fasting has been shown to decrease energy intake, body weight, adiposity, and improve glucose tolerance in HFD-fed mice [169]. HFD-induced adipose tissue inflammation and fibrosis can also be ameliorated by intermittent fasting [169]. Diurnal intermittent fasting during Ramadan can reduce inflammation and oxidative stress [170], and affect circadian rhythms of energy expenditure and body temperature in healthy people [171]. In addition, Ramadan intermittent fasting ameliorates the expression of antioxidants (Sod2, and Nrf2), and metabolic regulatory genes (Sirt1 and Sirt3) in obese and overweight subjects [172]. A recent randomized clinical study has suggested the intermittent fasting-medicated alteration of peripheral clock genes expression in subcutaneous adipose tissue in obese women, despite intermittent fasting does not uniformly impact the expression of clock genes [173]. Nevertheless, studies on time-restricted feeding and intermittent fasting have provided strong evidence that connects circadian clock and energy metabolism.
4.2. Clock-Controlled Pathways in Adipose Tissue
As mentioned, the molecular clock machinery drives the circadian transcription of CCGs in peripheral tissues. These CCGs encode for many important regulatory proteins and enzymes that involve in different vital cellular processes. The circadian regulation of CCGs, however, is highly different among tissues, which complicates the research field of circadian rhythm. To date, there are only few studies focused on the circadian regulation in adipose tissue. Identification of important CGGs and pathways involved in adipose tissue function is important to improve the understanding of clock involvement in the pathophysiology of metabolic diseases. Here, we propose some important targets and summarize important findings that link circadian rhythm and oxidative stress in adipose tissue.
4.2.1. Peroxisome Proliferator–Activated Receptors
PPARs are ligand-activated transcription factors of nuclear receptor superfamily consisting of three subtypes (PPARα, PPARβ/δ, and PPARγ), which are differentially expressed among tissues in mammals [174]. PPAR family plays a major role in regulating energy homeostasis and metabolic function, via controlling the transcription of genes involved in lipid and glucose metabolism [175,176]. PPARα is abundant in BAT, whereas PPARγ is expressed in both BAT and WAT [177]. Pparα and Pparγ mRNA expression are maximal in WAT at the end of the inactive phase, which prepare the metabolic machinery to metabolize fat [178]. PPARβ/δ promotes lipolysis by activating genes involved in fatty acid oxidation in adipocytes [179]. Expression of PPARβ/δ has been shown to oscillate in-phase with that of UCP1 [178], suggesting that PPARβ/δ could regulate the circadian control of energy dissipation in BAT. PPARγ and PPARα synergize to induce browning of WAT in vivo, via PPARγ activation and PPARα-mediated FGF21 expression [180].
In clinical practice, PPARα agonists are proven to reduce low density lipoproteins (LDLs) and TGs, whereas PPARγ ligands are used to improve glycemic control via insulin sensitization in patients with type 2 diabetes [181]. In obese rats, PPARγ agonists also increase insulin sensitivity, reduce fasting insulin levels and TG concentration, and increase hydrogen sulphide (H2S) production in PVAT, which improves in the anticontractile effect of PVAT on aortic rings [182]. H2S activates PPARγ by sulphhydrating cysteine 139 residue in PPARγ leading to enhancement of glucose uptake and lipid storage [183].
The rhythmic expression of PPARs is mediated by the CLOCK:BMAL heterodimer [184]. On the other hand, PPARα activates Bmal1 expression by binding to the peroxisome-proliferator response element (PPRE) at its promotor region [185,186]. Obesity attenuates expression of BMAL1and REV-ERB [187], which are correlated with increased expression of PPARα [16]. Pparγ-deficient mice display dampened behavioral and cellular circadian rhythms [188], while PPARα agonists have been shown to promote locomotor activity and feeding daily rhythms in mice [189]. In addition, PPARα agonist, bezafibrate, has been shown to normalize Per2, Bmal1, and Rev-erbα circadian expression in adipose tissue of mice with circadian disruption [190]. Indeed, many PPARγ target genes, including Adiponectin and Leptin, show rhythmic expression in WAT [178]. Moreover, PPARγ pathway also regulates important antioxidant enzymes including heme oxygenase-1 (HO-1) and NRF2 [191], suggesting the possible link between circadian rhythm and oxidative stress.
4.2.2. PPARγ Coactivator 1
The PGC1 is a family of transcriptional coactivators that regulates mitochondrial biogenesis. PPARγ and PGC-1α are important molecules that regulate mitochondrial and adipose function by modulating the white-to-brown differentiation of adipocytes [192]. PGC-1α expression can be induced in BAT during cold exposure [193].
All family members of PGC1 exhibit circadian expression, while PGC1α can in turn stimulate the expression of Bmal1, Clock, Per2, and Rev-erbα through coactivation of the ROR family [194,195]. Pgc-1α null mice have been shown to have disrupted clock and metabolic genes expression, which leads to abnormal diurnal rhythms of body temperature, activity, and metabolism [194]. On the other hand, Pgc-1β null mice display altered expression of various nuclear-encoded genes that are responsible for mitochondrial and metabolic functions in BAT, while HFD-feeding induces hepatic steatosis and increases serum TG and cholesterol levels in these mice [196], suggesting that PGC-1β plays an important role in controlling mitochondrial oxidative energy metabolism.
4.2.3. SIRT1
Sirtuin 1 (SIRT1) belongs to the family of nicotinamide adenine dinucleotide (NAD)-dependent protein deacetylase [197]. SIRT1 is involved in many biological processes, including cell proliferation and survival, DNA damage repairment and energy metabolism [198,199], which is an important therapeutic target for metabolic and cardiovascular diseases [200].
SIRT1 plays a pivotal role in reducing local superoxide production, modulating browning process, and promoting adipokines production in adipose tissue [201]. In adipose tissue-specific Sirt1-deficienct mice, the obesity-induced brown-to-white transition of PVAT is exaggerated in vivo, which augments endothelial dysfunction [201]. In obese mice, treatment with SRT1720 (a SIRT1 specific activator) prolongs the lifespan and reverses obesity-induced organ damages via normalizing the acetylation of PGC-1α to promote mitochondrial biogenesis in adipose tissue [202]. Activation of SIRT1 can normalize inflammatory insult-induced [203] or oxidative stress-induced adipokines dysregulation from adipose tissue and prevent arterial remodeling [204,205]. Moreover, resveratrol, a polyphenol that can activate SIRT1, can ameliorate adipokine release from dysregulated PVAT via the SIRT1/AMPK pathway [206]. In adipocytes, SIRT1 regulates the secretion of adiponectin through the interaction with FOXO1 [206].
In addition, SIRT1 is involved in the crosstalk between the circadian clock and energy metabolism [207]. SIRT1 is required for high-magnitude circadian transcription of several core clock genes [208], and regulates the transcription of other circadian-related genes by deacetylation of histone H3 lysine 9 (H3K9) on their promoters [209]. SIRT1 deacetylates BMAL1 and PER2 to affect their activities [210,211]. In addition, a negative reciprocal relationship is reported between SIRT1 and PER2, where PER2 negatively regulates the transcription of Sirt1 through binding to the CLOCK:BMAL1 binding site at Sirt1 promoter [211]. In Sirt1-deficienct mice, disrupted circadian rhythms and altered amplitude of Per1, Per2, Cry1 and Cry2 expression are observed [211].
One of the targets of CLOCK:BMAL1 heterodimer is nicotinamide phosphoribosyltransferase (NAMPT), an rate-limiting enzyme in the biosynthesis of NAD which can regulate SIRT1 activity in circadian rhythm [212,213]. As aforementioned, NAD also plays a pivotal role in circadian epigenomic regulation and shows robust diurnal rhythms [209]. High NAD level inhibits the DNA binding activity of CLOCK:BMAL1 [148]. While NAD level affects the activity of SIRT1, SIRT1-mediated deacetylation of acetyl-CoA synthetase 1 (AceCS1) affects the circadian levels of metabolites including NAD and acetyl-CoA [214]. Indeed, the circadian level of acetyl-CoA may also lead to the circadian acetylation of histones and other non-histone proteins [215,216]. These suggest that SIRT1 is a potent regulator that links circadian, oxidative stress and cellular energy metabolism.
4.2.4. BCAA Metabolism and mTOR Signaling
Recent transcriptome studies in human adipose tissue reveal the circadian regulations of lipid and nucleic acid metabolism, as well as key metabolic pathways such as the citric acid cycle and branched chain amino acid (BCAA) degradation [23]. High levels of BCAAs have been recently identified as contributors of chronic inflammation and lead to the development of insulin resistance and diabetes [217]. BCAAs may also trigger the production of ROS through the activation of NOX1 and NOX2 [218].
Mammalian target of rapamycin (mTOR) is an important regulator of eukaryotic growth and metabolism, sensor of cellular nutrient and energy levels, and is involved in protein synthesis [219,220]. BCAA can activate mTOR, while mTOR is also particularly sensitive to BCAAs. BCAAs are required for cell proliferation in a mTOR complex 1 (mTORC1)-dependent pathway [221]. mTOR signaling links the circadian clock and cellular metabolism. In the SCN, mTOR pathway can be modulated by light-dark cycle [222]. Moreover, protein 70 S6 kinase 1 (S6K1), a key factor in the mTOR pathway, is able to phosphorylate BMAL1 in circadian rhythm, thus affects the CLOCK:BMAL1-mediated circadian translational machinery [223]. In mouse adipose tissue, mRNA expression of Rictor and mTOR peaks during the day and troughs at night, while Rictor-adipose-tissue-specific KO mice have altered circadian genes expression in adipose tissue and non-dipping hypertension development [224,225].
In addition, BCAA is a negative regulator of KLF15 expression at transcriptional level via phosphoinositide 3-kinases (P13K)/AKT signaling [226]. Adipose KLF15 has been recently identified as an essential regulator of adipocyte lipid metabolism and systemic energy balance. Mice lacking KLF15 in adipose tissue have decreased adiposity and are protected from HFD-induced obesity [227]. KLF15 regulates key genes responsible for TG synthesis and inhibits lipolytic action [227]. This suggests that BCAA metabolism and mTOR signaling in adipose tissue are critical targets in modulating circadian rhythm and cardiovascular function.
4.2.5. GLP-1 and DPP-IV
The enteroendocrine incretin hormone glucagon-like petide-1 (GLP-1) has been reported to have protective effects on the cardiovascular system [228] and to improve endothelial function in obesity [229]. It is previously reported that the secretion of GLP-1 is in a circadian-manner [230,231]. GLP-1 has been shown to activate genes related to fatty acid oxidation and insulin signaling pathways, thus enhancing antioxidant capacity through its interaction with the G protein-coupled receptor GLP-1R in adipose tissue [232]. GLP-1 receptor agonist, liraglutide, has been shown to alleviate vascular dysfunction by modulating the protein kinase A (PKA)-AMPK-PGC-1α pathway and enhancing antioxidant enzymatic system Nrf2/HO-1 in PVAT of HFD-induced obese mice [233].
Dipeptidyl peptidase IV (DPP-IV) is a serine protease responsible for cleaving and inactivating of incretin peptides and GLP-1, which is correlated to the progression of metabolic and cardiovascular disease [234]. Various DPP-IV inhibitors have been shown to exert direct antioxidant effects, increase insulin secretion and improve glycemic control in diabetic animals and humans [235,236,237]. DPP-IV inhibitor, teneligliptin, reduces atherosclerosis progression in apolipoprotein E (ApoE) knockout mice by alleviating inflammation and oxidative stress in both the vasculature and PVAT [238]. Indeed, DPP-IV inhibition seems to result in an overall increase in GLP-1 levels with preserved circadian rhythm throughout the day [239]. Recently, treatment with vildagliptin, another DPP-IV inhibitor, has been shown to restore the normal dipping pattern of blood pressure in rat model of salt-dependent hypertension [240]. These results indicate DPP-IV as a potential treatment target for circadian disruption and adipose dysfunction.
4.2.6. AMPK
AMPK is a cellular energy sensor whose phosphorylation activates cellular catabolism [241]. AMPK consists of a catalytic (α) subunit and two regulatory (β, γ) subunits. The AMPKα1 isoform is expressed in adipose tissue [242]. Adipose tissue-specific AMPKα1/α2 KO mice display impaired mitochondrial integrity and biogenesis and reduced thermogenic gene expression in adipose tissue upon cold exposure [243], suggesting a role of AMPK in regulating the browning process of WAT. On the other hand, obesity-linked PVAT dysfunction is associated with AMPK phosphorylation [244].
AMPK is an important energy sensor that connects nutrient signal to circadian clocks. Circadian oscillations in the phosphorylation of the AMPK substrates acetyl-coA carboxylase (ACC) and Raptor have been observed [245,246], suggesting the circadian activity of AMPK. AMPK also phosphorylates CRY1 at Ser 71 residue, which stimulates the direct binding of the F-box and leucine-rich repeat protein 3 (FBXL3) to CRY1 for ubiquitin-mediated degradation [247]. Moreover, AMPK phosphorylates and activates casein kinase Iε, which leads to degradation of PERs [248]. Indeed, pharmacological activation of AMPK has been shown to induce a phase shift in circadian rhythm in a tissue-specific manner [249,250,251]. Ampk1-deficient mice have attenuated diurnal alteration of core body temperature, altered circadian gene expression in adipose tissue, and blunted NAMPT rhythmicity [242]. These results suggest that AMPK is an important metabolic cue to entrain peripheral clocks. However, further studies are warranted to dissect the precise role of AMPK in the entrainment of adipose tissue clocks.
4.2.7. eNOS
Endogenous NO generated from eNOS is critical in modulating endothelial function and homeostasis. eNOS is also involved in other vascular processes including leukocyte adhesion, smooth muscle cell proliferation and migration, and platelet aggregation [252]. In addition, eNOS defends against oxidative stress by producing NO which can enhance superoxide dismutation by stimulating SOD expression [253,254]. The beneficial effects of important crosstalk between SIRT1 and eNOS in the endothelial function and oxidative stress defense has been reported extensively [200,255,256].
There is evidence showing that the activities of eNOS are regulated by the peripheral circadian clock in different tissues including brain, aorta and lung [257,258,259]. Impaired eNOS signaling has been reported in mice with circadian disruption [99,260,261], although eNOS expression is not altered in these mice. It has been shown that post-translational mechanisms, especially phosphorylation of eNOS activity, are compromised, consistent with observations demonstrating that eNOS activity exhibits a circadian variation [99,260,262]. eNOS uncoupling and biopterin imbalance have been reported in Bmal1-KO mice [263]. Interestingly, leptin has been shown to control the circadian release of NO via TNF-α signaling [264]. This suggests that the circadian activity of adipose eNOS may be regulated via post-translational modification but not by transcriptional control.
The involvement of eNOS in circadian regulation is controversial. Indeed, eNOS may not involves in the photic entrainment in mice, evidenced by no significant changes in phase-shift locomotor activity [265] and circadian blood pressure [266] in eNOS−/− mice compared to wild type mice during light-dark or dark-dark cycle. On the other hand, impaired circadian rhythmicity is related to a decrease of NO production with aging [260]. NO has been shown to upregulate the circadian expression of Per genes via the cAMP response element–dependent and the E-box enhancer element–dependent pathways [260]. Yet, NO cannot improve the circadian variation in blood pressure of circadian disrupted mice [260]. These suggest that eNOS may not be an important player in modulation the central circadian clock. Nevertheless, the role of eNOS in adipose clock remains to be elucidated, but likely via NO production.
Recently, both gene and protein expression of eNOS have been detected in adipose tissues [267,268], while eNOS in PVAT contributes to vascular NO production [269,270,271]. eNOS is abundantly found in both isolated brown adipocytes and BAT [272], while eNOS-derived NO can promote adiponectin synthesis and mitochondrial biogenesis [273]. On the other hand, HFD-induced PVAT dysfunction is associated with reduction of eNOS expression and NO production [274]. Treatment with an anti-inflammatory and antioxidant drug, methotrexate, has been shown to improve PVAT dysfunction and ameliorate adipokine dysregulation via the activation of the AMPK/eNOS pathway [275]. These suggest the importance of eNOS and NO in modulating adipose tissue function, browning and thermogenesis. Thereby, dissecting the crosstalk between eNOS and the local clock in adipose tissue could be an interesting direction for future studies (Figure 3).
5. Conclusions
Obesity is characterized by excess accumulation of fat in adipose tissue, which leads to adipocyte hypertrophy and hypoxia. The effects of excess energy consumption, overloading of mitochondrial activity and generation of oxidative stress in adipose tissue are previously known to associate with obesity. However, results from time-restricted-feeding experiments have shown that circadian rhythm is a critical component that can alter adipose tissue function. Misalignment between circadian rhythm and metabolism contributes to the pathogenesis of chronic metabolic and cardiovascular diseases. Disruption of the circadian rhythmicity is associated with augmented oxidative stress and aberrated adipose tissue function and metabolism.
In experimental models, knockouts or mutations in clock genes that lead to the disruption of circadian rhythmicity have suggested a tight link between the circadian clock, adipose tissue function and metabolism. However, since peripheral clock machinery is highly tissue-specific, there is a need to generate adipose tissue-specific knockout mice to dissect the adipose clock. To date, there are only a few studies that generated adipose tissue-specific clock gene knockout mice and reported the effects on adipose tissue function and oxidative stress (Table 1). Further in vivo studies using these adipose tissue-specific knockout mice are needed to dissect the detailed crosstalk between the clock and oxidative stress in adipose tissue.
Considerable evidence supports the hypothesis that oxidative stress and circadian rhythm are interrelated, as the circadian clock regulates the rhythmic oscillation of cellular redox function, while changes in redox state in the cell can drive the clock machinery. This gives rise to the use of potential antioxidant treatment to target and entrain the adipose tissue clock. Although the detailed mechanisms and targets of CCGs in adipose tissue are not completely known, we have summarized some potential molecular targets in adipose tissue for antioxidant treatment. The uses of antioxidant drugs, for example, liraglutide (GLP-1 receptor agonist), rosiglitazone (PPARγ agonist), SRT1720 (SIRT1 activator), as well as antioxidant food supplements including polyphenols can also retrain circadian clock and adipose tissue function in human. However, these warrant further systematic in vivo studies to explore the best treatment/prevention strategies.
Author Contributions
A.W.C.M. wrote the initial draft of the manuscript. N.X., and H.L. critically reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Original works from the authors’ laboratory contributing to this review were supported by grants LI-1042/1-1, LI-1042/3-1, LI-1042/5-1, and XI 139/2-1 from the Deutsche Forschungsgemeinschaft (DFG), Bonn, Germany. HL and NX were supported by a research grant from the Boehringer Ingelheim Foundation for the collaborative research consortium “Novel and neglected cardiovascular risk factors: molecular mechanisms and therapeutic implications.”
Conflicts of Interest
The authors declare no conflict of interest.
References
- Collaborators, G.O. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox, and impact of weight loss. J. Am. Coll. Cardiol. 2009, 53, 1925–1932. [Google Scholar] [CrossRef] [Green Version]
- Bass, J.; Lazar, M.A. Circadian time signatures of fitness and disease. Science 2016, 354, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Fasshauer, M.; Bluher, M. Adipokines in health and disease. Trends Pharm. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Shostak, A.; Meyer-Kovac, J.; Oster, H. Circadian regulation of lipid mobilization in white adipose tissues. Diabetes 2013, 62, 2195–2203. [Google Scholar] [CrossRef] [Green Version]
- Kalsbeek, A.; Fliers, E.; Romijn, J.A.; La Fleur, S.E.; Wortel, J.; Bakker, O.; Endert, E.; Buijs, R.M. The suprachiasmatic nucleus generates the diurnal changes in plasma leptin levels. Endocrinology 2001, 142, 2677–2685. [Google Scholar] [CrossRef]
- Rudic, R.D.; McNamara, P.; Curtis, A.M.; Boston, R.C.; Panda, S.; Hogenesch, J.B.; Fitzgerald, G.A. BMAL1 and CLOCK, two essential components of the circadian clock, are involved in glucose homeostasis. PLoS Biol. 2004, 2, e377. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.M.; Leandro, A.; Azul, L.; Seica, R.; Perry, G. Vascular Oxidative Stress: Impact and Therapeutic Approaches. Front. Physiol. 2018, 9, 1668. [Google Scholar] [CrossRef] [Green Version]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 29, 345–350. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Horvath, T.L.; Andrews, Z.B.; Diano, S. Fuel utilization by hypothalamic neurons: Roles for ROS. Trends Endocrinol. Metab. TEM 2009, 20, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Forstermann, U. Uncoupling of endothelial NO synthase in atherosclerosis and vascular disease. Curr Opin Pharm. 2013, 13, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Chrysohoou, C.; Panagiotakos, D.B.; Pitsavos, C.; Skoumas, I.; Papademetriou, L.; Economou, M.; Stefanadis, C. The implication of obesity on total antioxidant capacity in apparently healthy men and women: The ATTICA study. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 590–597. [Google Scholar] [CrossRef]
- Wilking, M.; Ndiaye, M.; Mukhtar, H.; Ahmad, N. Circadian rhythm connections to oxidative stress: Implications for human health. Antioxid. Redox Signal. 2013, 19, 192–208. [Google Scholar] [CrossRef] [Green Version]
- Bass, J.; Takahashi, J.S. Circadian integration of metabolism and energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935. [Google Scholar] [CrossRef]
- Potter, G.D.; Cade, J.E.; Grant, P.J.; Hardie, L.J. Nutrition and the circadian system. Br. J. Nutr. 2016, 116, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Monk, T.H. Enhancing circadian zeitgebers. Sleep 2010, 33, 421–422. [Google Scholar] [CrossRef] [Green Version]
- Mukherji, A.; Kobiita, A.; Damara, M.; Misra, N.; Meziane, H.; Champy, M.-F.; Chambon, P. Shifting eating to the circadian rest phase misaligns the peripheral clocks with the master SCN clock and leads to a metabolic syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, E6691–E6698. [Google Scholar] [CrossRef] [Green Version]
- Froy, O.; Garaulet, M. The Circadian Clock in White and Brown Adipose Tissue: Mechanistic, Endocrine, and Clinical Aspects. Endocr Rev. 2018, 39, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christou, S.; Wehrens, S.M.T.; Isherwood, C.; Moller-Levet, C.S.; Wu, H.; Revell, V.L.; Bucca, G.; Skene, D.J.; Laing, E.E.; Archer, S.N.; et al. Circadian regulation in human white adipose tissue revealed by transcriptome and metabolic network analysis. Sci. Rep. 2019, 9, 2641. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, R.; Pfeifer, A. Fat tissues, the brite and the dark sides. Pflug. Arch. Eur. J. Physiol. 2016, 468, 1803–1807. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.; Duncan, R.E.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Triacylglycerol metabolism in adipose tissue. Future Lipidol. 2007, 2, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Aldhahi, W.; Hamdy, O. Adipokines, inflammation, and the endothelium in diabetes. Curr. Diabetes Rep. 2003, 3, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Vitali, A.; Murano, I.; Zingaretti, M.C.; Frontini, A.; Ricquier, D.; Cinti, S. The adipose organ of obesity-prone C57BL/6J mice is composed of mixed white and brown adipocytes. J. Lipid Res. 2012, 53, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Lafontan, M. Advances in adipose tissue metabolism. Int. J. Obes. 2008, 32 (Suppl. 7), S39–S51. [Google Scholar] [CrossRef] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Lepper, C.; Fan, C.M. Inducible lineage tracing of Pax7-descendant cells reveals embryonic origin of adult satellite cells. Genesis 2010, 48, 424–436. [Google Scholar] [CrossRef]
- An, Y.; Wang, G.; Diao, Y.; Long, Y.; Fu, X.; Weng, M.; Zhou, L.; Sun, K.; Cheung, T.H.; Ip, N.Y.; et al. A Molecular Switch Regulating Cell Fate Choice between Muscle Progenitor Cells and Brown Adipocytes. Dev. Cell 2017, 41, 382–391 e385. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerback, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Kayahara, T.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Recruited brown adipose tissue as an antiobesity agent in humans. J. Clin. Investig. 2013, 123, 3404–3408. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.I.; Middelbeek, R.J.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.H.; et al. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Lizcano, F. The Beige Adipocyte as a Therapy for Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 58. [Google Scholar] [CrossRef] [Green Version]
- Dewal, R.S.; Stanford, K.I. Effects of exercise on brown and beige adipocytes. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2019, 1864, 71–78. [Google Scholar] [CrossRef]
- Min, S.Y.; Kady, J.; Nam, M.; Rojas-Rodriguez, R.; Berkenwald, A.; Kim, J.H.; Noh, H.L.; Kim, J.K.; Cooper, M.P.; Fitzgibbons, T.; et al. Human ‘brite/beige’ adipocytes develop from capillary networks, and their implantation improves metabolic homeostasis in mice. Nat. Med. 2016, 22, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Altshuler-Keylin, S.; Shinoda, K.; Hasegawa, Y.; Ikeda, K.; Hong, H.; Kang, Q.; Yang, Y.; Perera, R.M.; Debnath, J.; Kajimura, S. Beige Adipocyte Maintenance Is Regulated by Autophagy-Induced Mitochondrial Clearance. Cell Metab. 2016, 24, 402–419. [Google Scholar] [CrossRef] [Green Version]
- Roh, H.C.; Tsai, L.T.Y.; Shao, M.; Tenen, D.; Shen, Y.; Kumari, M.; Lyubetskaya, A.; Jacobs, C.; Dawes, B.; Gupta, R.K.; et al. Warming Induces Significant Reprogramming of Beige, but Not Brown, Adipocyte Cellular Identity. Cell Metab. 2018, 27, 1121–1137 e1125. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; Spiegelman, B.M. FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar]
- Bordicchia, M.; Liu, D.; Amri, E.Z.; Ailhaud, G.; Dessi-Fulgheri, P.; Zhang, C.; Takahashi, N.; Sarzani, R.; Collins, S. Cardiac natriuretic peptides act via p38 MAPK to induce the brown fat thermogenic program in mouse and human adipocytes. J. Clin. Investig. 2012, 122, 1022–1036. [Google Scholar] [CrossRef] [Green Version]
- Schulz, T.J.; Huang, P.; Huang, T.L.; Xue, R.D.; McDougall, L.E.; Townsend, K.L.; Cypess, A.M.; Mishina, Y.; Gussoni, E.; Tseng, Y.H. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 2013, 495, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Aldiss, P.; Davies, G.; Woods, R.; Budge, H.; Sacks, H.S.; Symonds, M.E. ‘Browning’ the cardiac and peri-vascular adipose tissues to modulate cardiovascular risk. Int. J. Cardiol. 2017, 228, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: Effects of cold exposure and adiposity. Diabetes 2009, 58, 1526–1531. [Google Scholar] [CrossRef] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef]
- Planavila, A.; Redondo, I.; Hondares, E.; Vinciguerra, M.; Munts, C.; Iglesias, R.; Gabrielli, L.A.; Sitges, M.; Giralt, M.; van Bilsen, M.; et al. Fibroblast growth factor 21 protects against cardiac hypertrophy in mice. Nat. Commun. 2013, 4, 2019. [Google Scholar] [CrossRef]
- Liu, S.Q.; Roberts, D.; Kharitonenkov, A.; Zhang, B.; Hanson, S.M.; Li, Y.C.; Zhang, L.Q.; Wu, Y.H. Endocrine protection of ischemic myocardium by FGF21 from the liver and adipose tissue. Sci. Rep. 2013, 3, 2767. [Google Scholar] [CrossRef] [Green Version]
- Gimeno, R.E.; Moller, D.E. FGF21-based pharmacotherapy--potential utility for metabolic disorders. Trends Endocrinol. Metab. TEM 2014, 25, 303–311. [Google Scholar] [CrossRef]
- Bookout, A.L.; de Groot, M.H.; Owen, B.M.; Lee, S.; Gautron, L.; Lawrence, H.L.; Ding, X.; Elmquist, J.K.; Takahashi, J.S.; Mangelsdorf, D.J.; et al. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nat. Med. 2013, 19, 1147–1152. [Google Scholar] [CrossRef]
- Man, A.W.C.; Zhou, Y.; Xia, N.; Li, H. Perivascular Adipose Tissue as a Target for Antioxidant Therapy for Cardiovascular Complications. Antioxidants 2020, 9, 574. [Google Scholar] [CrossRef]
- Xia, N.; Li, H.G. The role of perivascular adipose tissue in obesity-induced vascular dysfunction. Br. J. Pharmacol. 2017, 174, 3425–3442. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Milton, H.; Eitzman, D.T.; Chen, Y.E. Paradoxical roles of perivascular adipose tissue in atherosclerosis and hypertension. Circ. J. 2012, CJ-12-1393. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.J.; Zeng, Z.H.; Teoh, K.; Sharma, A.M.; Abouzahr, L.; Cybulsky, I.; Lamy, A.; Semelhago, L.; Lee, R.M. Perivascular adipose tissue modulates vascular function in the human internal thoracic artery. J. Thorac. Cardiovasc. Surg. 2005, 130, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- Greenstein, A.S.; Khavandi, K.; Withers, S.B.; Sonoyama, K.; Clancy, O.; Jeziorska, M.; Laing, I.; Yates, A.P.; Pemberton, P.W.; Malik, R.A.; et al. Local inflammation and hypoxia abolish the protective anticontractile properties of perivascular fat in obese patients. Circulation 2009, 119, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.K.; Zhou, Z.; Zhang, J.; Zeng, R.; Wu, J.; Eitzman, D.T.; Chen, Y.E.; Chang, L. Perivascular adipose tissue in vascular function and disease: A review of current research and animal models. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1621–1630. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.; Chatterjee, T.K.; Tang, Y.; Hui, D.Y.; Weintraub, N.L. Proinflammatory phenotype of perivascular adipocytes. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1631–1636. [Google Scholar] [CrossRef] [Green Version]
- Gil-Ortega, M.; Somoza, B.; Huang, Y.; Gollasch, M.; Fernandez-Alfonso, M.S. Regional differences in perivascular adipose tissue impacting vascular homeostasis. Trends Endocrinol. Metab. TEM 2015, 26, 367–375. [Google Scholar] [CrossRef]
- Drosos, I.; Chalikias, G.; Pavlaki, M.; Kareli, D.; Epitropou, G.; Bougioukas, G.; Mikroulis, D.; Konstantinou, F.; Giatromanolaki, A.; Ritis, K.; et al. Differences between perivascular adipose tissue surrounding the heart and the internal mammary artery: Possible role for the leptin-inflammation-fibrosis-hypoxia axis. Clin. Res. Cardiol. 2016, 105, 887–900. [Google Scholar] [CrossRef]
- Victorio, J.A.; Fontes, M.T.; Rossoni, L.V.; Davel, A.P. Different Anti-Contractile Function and Nitric Oxide Production of Thoracic and Abdominal Perivascular Adipose Tissues. Front. Physiol. 2016, 7, 295. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Garcia-Barrio, M.T.; Chen, Y.E. Perivascular Adipose Tissue Regulates Vascular Function by Targeting Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Downs, L.C.; Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. Aging exacerbates obesity-induced oxidative stress and inflammation in perivascular adipose tissue in mice: A paracrine mechanism contributing to vascular redox dysregulation and inflammation. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2013, 68, 780–792. [Google Scholar] [CrossRef] [Green Version]
- Gil-Ortega, M.; Condezo-Hoyos, L.; Garcia-Prieto, C.F.; Arribas, S.M.; Gonzalez, M.C.; Aranguez, I.; Ruiz-Gayo, M.; Somoza, B.; Fernandez-Alfonso, M.S. Imbalance between Pro and Anti-Oxidant Mechanisms in Perivascular Adipose Tissue Aggravates Long-Term High-Fat Diet-Derived Endothelial Dysfunction. PLoS ONE 2014, 9, e95312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldiss, P.; Lewis, J.E.; Lupini, I.; Boocock, D.J.; Miles, A.K.; Ebling, F.J.; Budge, H.; Symonds, M.E. Exercise does not induce browning of WAT at thermoneutrality and induces an oxidative, myogenic signature in BAT. bioRxiv 2019, 649061. [Google Scholar]
- Dutheil, F.; Gordon, B.A.; Naughton, G.; Crendal, E.; Courteix, D.; Chaplais, E.; Thivel, D.; Lac, G.; Benson, A.C. Cardiovascular risk of adipokines: A review. J. Int. Med Res. 2018, 46, 2082–2095. [Google Scholar] [CrossRef]
- Deng, Y.; Scherer, P.E. Adipokines as novel biomarkers and regulators of the metabolic syndrome. Ann. N. Y. Acad. Sci. 2010, 1212, E1–E19. [Google Scholar] [CrossRef]
- Lafontan, M. Historical perspectives in fat cell biology: The fat cell as a model for the investigation of hormonal and metabolic pathways. Am. J. Physiol. Cell Physiol. 2012, 302, C327–C359. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.M.; Pereira, A.; Fernandes, R.; Letra, L.; Seica, R.M. Adiponectin improves endothelial function in mesenteric arteries of rats fed a high-fat diet: Role of perivascular adipose tissue. Br. J. Pharmacol. 2017, 174, 3514–3526. [Google Scholar] [CrossRef] [Green Version]
- Challet, E. Circadian aspects of adipokine regulation in rodents. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 573–582. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Kawano, H.; Motoyama, T.; Yasue, H.; Hirai, N.; Waly, H.M.; Kugiyama, K.; Ogawa, H. Endothelial function fluctuates with diurnal variation in the frequency of ischemic episodes in patients with variant angina. J. Am. Coll. Cardiol. 2002, 40, 266–270. [Google Scholar] [PubMed] [Green Version]
- Otto, M.E.; Svatikova, A.; de Mattos Barretto, R.B.; Santos, S.; Hoffmann, M.; Khandheria, B.; Somers, V. Early morning attenuation of endothelial function in healthy humans. Circulation 2004, 109, 2507–2510. [Google Scholar] [PubMed] [Green Version]
- Walters, J.F.; Skene, D.J.; Hampton, S.M.; Ferns, G.A. Biological rhythms, endothelial health and cardiovascular disease. Med Sci. Monit. 2003, 9, RA1–RA8. [Google Scholar] [PubMed]
- Singh, R.B.; Cornélissen, G.; Weydahl, A.; Schwartzkopff, O.; Katinas, G.; Otsuka, K.; Watanabe, Y.; Yano, S.; Mori, H.; Ichimaru, Y. Circadian heart rate and blood pressure variability considered for research and patient care. Int. J. Cardiol. 2003, 87, 9–28. [Google Scholar]
- Panza, J.A.; Epstein, S.E.; Quyyumi, A.A. Circadian variation in vascular tone and its relation to α-sympathetic vasoconstrictor activity. N. Engl. J. Med. 1991, 325, 986–990. [Google Scholar]
- Lee, C.; Etchegaray, J.P.; Cagampang, F.R.; Loudon, A.S.; Reppert, S.M. Posttranslational mechanisms regulate the mammalian circadian clock. Cell 2001, 107, 855–867. [Google Scholar] [CrossRef]
- Young, M.E. The circadian clock within the heart: Potential influence on myocardial gene expression, metabolism, and function. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1–H16. [Google Scholar] [CrossRef] [Green Version]
- Reppert, S.M.; Weaver, D.R. Molecular analysis of mammalian circadian rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Kraves, S.; Weitz, C.J. A role for cardiotrophin-like cytokine in the circadian control of mammalian locomotor activity. Nat. Neurosci. 2006, 9, 212–219. [Google Scholar] [CrossRef]
- Cheng, M.Y.; Bullock, C.M.; Li, C.; Lee, A.G.; Bermak, J.C.; Belluzzi, J.; Weaver, D.R.; Leslie, F.M.; Zhou, Q.Y. Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus. Nature 2002, 417, 405–410. [Google Scholar] [CrossRef]
- Kiehn, J.-T.; Koch, C.E.; Walter, M.; Brod, A.; Oster, H. Circadian rhythms and clocks in adipose tissues: Current Insights. ChronoPhysiology Ther. 2017, 7, 7–17. [Google Scholar]
- Ando, H.; Yanagihara, H.; Hayashi, Y.; Obi, Y.; Tsuruoka, S.; Takamura, T.; Kaneko, S.; Fujimura, A. Rhythmic messenger ribonucleic acid expression of clock genes and adipocytokines in mouse visceral adipose tissue. Endocrinology 2005, 146, 5631–5636. [Google Scholar] [CrossRef] [PubMed]
- Zvonic, S.; Ptitsyn, A.A.; Conrad, S.A.; Scott, L.K.; Floyd, Z.E.; Kilroy, G.; Wu, X.; Goh, B.C.; Mynatt, R.L.; Gimble, J.M. Characterization of peripheral circadian clocks in adipose tissues. Diabetes 2006, 55, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zvonic, S.; Floyd, Z.E.; Kilroy, G.; Goh, B.C.; Hernandez, T.L.; Eckel, R.H.; Mynatt, R.L.; Gimble, J.M. Induction of circadian gene expression in human subcutaneous adipose-derived stem cells. Obesity 2007, 15, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Otway, D.T.; Mantele, S.; Bretschneider, S.; Wright, J.; Trayhurn, P.; Skene, D.J.; Robertson, M.D.; Johnston, J.D. Rhythmic diurnal gene expression in human adipose tissue from individuals who are lean, overweight, and type 2 diabetic. Diabetes 2011, 60, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Santos, C.; Gomez-Abellan, P.; Madrid, J.A.; Hernandez-Morante, J.J.; Lujan, J.A.; Ordovas, J.M.; Garaulet, M. Circadian rhythm of clock genes in human adipose explants. Obesity 2009, 17, 1481–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhr, E.D.; Takahashi, J.S. Molecular components of the Mammalian circadian clock. Handb. Exp. Pharmacol. 2013, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Schibler, U.; Ripperger, J.; Brown, S.A. Peripheral circadian oscillators in mammals: Time and food. J. Biol. Rhythm. 2003, 18, 250–260. [Google Scholar] [CrossRef]
- Froy, O.; Chang, D.C.; Reppert, S.M. Redox potential: Differential roles in dCRY and mCRY1 functions. Curr. Biol. CB 2002, 12, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Ueda, H.R.; Hayashi, S.; Chen, W.; Sano, M.; Machida, M.; Shigeyoshi, Y.; Iino, M.; Hashimoto, S. System-level identification of transcriptional circuits underlying mammalian circadian clocks. Nat. Genet. 2005, 37, 187–192. [Google Scholar] [CrossRef]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, J.C. Molecular bases for circadian clocks. Cell 1999, 96, 271–290. [Google Scholar] [CrossRef] [Green Version]
- Onder, Y.; Green, C.B. Rhythms of metabolism in adipose tissue and mitochondria. Neurobiol. Sleep Circadian Rhythm. 2018, 4, 57–63. [Google Scholar] [CrossRef]
- Bray, M.; Young, M. The role of cell-specific circadian clocks in metabolism and disease. Obes. Rev. 2009, 10, 6–13. [Google Scholar] [CrossRef]
- Antoch, M.P.; Gorbacheva, V.Y.; Vykhovanets, O.; Toshkov, I.A.; Kondratov, R.V.; Kondratova, A.A.; Lee, C.; Nikitin, A.Y. Disruption of the circadian clock due to the Clock mutation has discrete effects on aging and carcinogenesis. Cell Cycle 2008, 7, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R. Obesity and metabolic syndrome in circadian Clock mutant mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Anea, C.B.; Zhang, M.; Stepp, D.W.; Simkins, G.B.; Reed, G.; Fulton, D.J.; Rudic, R.D. Vascular disease in mice with a dysfunctional circadian clock. Circulation 2009, 119, 1510–1517. [Google Scholar] [CrossRef] [Green Version]
- Shimba, S.; Ishii, N.; Ohta, Y.; Ohno, T.; Watabe, Y.; Hayashi, M.; Wada, T.; Aoyagi, T.; Tezuka, M. Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 12071–12076. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Chatterjee, S.; Li, L.; Kim, J.M.; Lee, J.; Yechoor, V.K.; Minze, L.J.; Hsueh, W.; Ma, K. The clock gene, brain and muscle Arnt-like 1, regulates adipogenesis via Wnt signaling pathway. FASEB J. 2012, 26, 3453–3463. [Google Scholar] [CrossRef]
- Nam, D.; Guo, B.; Chatterjee, S.; Chen, M.H.; Nelson, D.; Yechoor, V.K.; Ma, K. The adipocyte clock controls brown adipogenesis through the TGF-beta and BMP signaling pathways. J. Cell Sci. 2015, 128, 1835–1847. [Google Scholar] [CrossRef] [Green Version]
- Fournier, B.; Murray, B.; Gutzwiller, S.; Marcaletti, S.; Marcellin, D.; Bergling, S.; Brachat, S.; Persohn, E.; Pierrel, E.; Bombard, F.; et al. Blockade of the activin receptor IIb activates functional brown adipogenesis and thermogenesis by inducing mitochondrial oxidative metabolism. Mol. Cell. Biol. 2012, 32, 2871–2879. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.M.; Kim, S.; Tseng, C.Y.; Jeon, Y.H.; Choe, S.; Lee, D.K. BMP-9 as a potent brown adipogenic inducer with anti-obesity capacity. Biomaterials 2014, 35, 3172–3179. [Google Scholar] [CrossRef]
- Kondratov, R.V.; Vykhovanets, O.; Kondratova, A.A.; Antoch, M.P. Antioxidant N-acetyl-L-cysteine ameliorates symptoms of premature aging associated with the deficiency of the circadian protein BMAL1. Aging 2009, 1, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Chawla, A.; Lazar, M.A. Induction of Rev-ErbA alpha, an orphan receptor encoded on the opposite strand of the alpha-thyroid hormone receptor gene, during adipocyte differentiation. J. Biol. Chem. 1993, 268, 16265–16269. [Google Scholar]
- Bray, M.S.; Young, M.E. Circadian rhythms in the development of obesity: Potential role for the circadian clock within the adipocyte. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2007, 8, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Torra, I.P.; Tsibulsky, V.; Delaunay, F.; Saladin, R.; Laudet, V.; Fruchart, J.C.; Kosykh, V.; Staels, B. Circadian and glucocorticoid regulation of Rev-erbalpha expression in liver. Endocrinology 2000, 141, 3799–3806. [Google Scholar] [CrossRef]
- Delezie, J.; Dumont, S.; Dardente, H.; Oudart, H.; Grechez-Cassiau, A.; Klosen, P.; Teboul, M.; Delaunay, F.; Pevet, P.; Challet, E. The nuclear receptor REV-ERBalpha is required for the daily balance of carbohydrate and lipid metabolism. FASEB J. 2012, 26, 3321–3335. [Google Scholar] [CrossRef] [Green Version]
- Solt, L.A.; Wang, Y.; Banerjee, S.; Hughes, T.; Kojetin, D.J.; Lundasen, T.; Shin, Y.; Liu, J.; Cameron, M.D.; Noel, R.; et al. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature 2012, 485, 62–68. [Google Scholar] [CrossRef]
- Amador, A.; Wang, Y.; Banerjee, S.; Kameneka, T.M.; Solt, L.A.; Burris, T.P. Pharmacological and Genetic Modulation of REV-ERB Activity and Expression Affects Orexigenic Gene Expression. PLoS ONE 2016, 11, e0151014. [Google Scholar] [CrossRef]
- Gerhart-Hines, Z.; Feng, D.; Emmett, M.J.; Everett, L.J.; Loro, E.; Briggs, E.R.; Bugge, A.; Hou, C.; Ferrara, C.; Seale, P.; et al. The nuclear receptor Rev-erbalpha controls circadian thermogenic plasticity. Nature 2013, 503, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Chatterjee, S.; Yin, H.; Liu, R.; Lee, J.; Yechoor, V.K.; Ma, K. Novel Function of Rev-erbalpha in Promoting Brown Adipogenesis. Sci. Rep. 2015, 5, 11239. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 controls lipid metabolism by direct regulation of PPARgamma. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, A.; Costa, M.J.; Rivero-Gutierrez, B.; Ji, L.; Morgan, S.L.; Feldman, B.J. The Circadian Clock Regulates Adipogenesis by a Per3 Crosstalk Pathway to Klf15. Cell Rep. 2017, 21, 2367–2375. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Yong, Q.; Kurose, T.; Todo, T.; Mizunoya, W.; Fushiki, T.; Seino, Y.; Yamada, Y. Clock gene defect disrupts light-dependency of autonomic nerve activity. Biochem. Biophys. Res. Commun. 2007, 364, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Barclay, J.L.; Shostak, A.; Leliavski, A.; Tsang, A.H.; Johren, O.; Muller-Fielitz, H.; Landgraf, D.; Naujokat, N.; van der Horst, G.T.; Oster, H. High-fat diet-induced hyperinsulinemia and tissue-specific insulin resistance in Cry-deficient mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1053–E1063. [Google Scholar] [CrossRef] [Green Version]
- Narasimamurthy, R.; Hatori, M.; Nayak, S.K.; Liu, F.; Panda, S.; Verma, I.M. Circadian clock protein cryptochrome regulates the expression of proinflammatory cytokines. Proc. Natl. Acad. Sci. USA 2012, 109, 12662–12667. [Google Scholar] [CrossRef] [Green Version]
- Dashti, H.S.; Smith, C.E.; Lee, Y.C.; Parnell, L.D.; Lai, C.Q.; Arnett, D.K.; Ordovas, J.M.; Garaulet, M. CRY1 circadian gene variant interacts with carbohydrate intake for insulin resistance in two independent populations: Mediterranean and North American. Chronobiol. Int. 2014, 31, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Paschos, G.K.; Ibrahim, S.; Song, W.L.; Kunieda, T.; Grant, G.; Reyes, T.M.; Bradfield, C.A.; Vaughan, C.H.; Eiden, M.; Masoodi, M.; et al. Obesity in mice with adipocyte-specific deletion of clock component Arntl. Nat. Med. 2012, 18, 1768–1777. [Google Scholar] [CrossRef] [Green Version]
- Birky, T.L.; Bray, M.S. Understanding circadian gene function: Animal models of tissue-specific circadian disruption. IUBMB Life 2014, 66, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Xiong, W.; Zhao, X.; Fan, Y.; Guo, Y.; Garcia-Barrio, M.; Zhang, J.; Jiang, Z.; Lin, J.D.; Chen, Y.E. Bmal1 in Perivascular Adipose Tissue Regulates Resting-Phase Blood Pressure Through Transcriptional Regulation of Angiotensinogen. Circulation 2018, 138, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Metal ion-catalyzed oxidation of proteins: Biochemical mechanism and biological consequences. Free Radic. Biol. Med. 1990, 9, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Frohnert, B.I.; Sinaiko, A.R.; Serrot, F.J.; Foncea, R.E.; Moran, A.; Ikramuddin, S.; Choudry, U.; Bernlohr, D.A. Increased adipose protein carbonylation in human obesity. Obesity 2011, 19, 1735–1741. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Li, H.; Horke, S.; Forstermann, U. Vascular oxidative stress, nitric oxide and atherosclerosis. Atherosclerosis 2014, 237, 208–219. [Google Scholar] [CrossRef]
- Cancello, R.; Henegar, C.; Viguerie, N.; Taleb, S.; Poitou, C.; Rouault, C.; Coupaye, M.; Pelloux, V.; Hugol, D.; Bouillot, J.-L. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005, 54, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Lam, K.S.; Wang, Y.; Wu, D.; Lam, M.C.; Shen, J.; Wong, L.; Hoo, R.L.; Zhang, J.; Xu, A. Hypoxia dysregulates the production of adiponectin and plasminogen activator inhibitor-1 independent of reactive oxygen species in adipocytes. Biochem. Biophys. Res. Commun. 2006, 341, 549–556. [Google Scholar] [CrossRef]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1alpha Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Egg, M.; Koblitz, L.; Hirayama, J.; Schwerte, T.; Folterbauer, C.; Kurz, A.; Fiechtner, B.; Most, M.; Salvenmoser, W.; Sassone-Corsi, P.; et al. Linking Oxygen to Time: The Bidirectional Interaction Between the Hypoxic Signaling Pathway and the Circadian Clock. Chronobiol. Int. 2013, 30, 510–529. [Google Scholar] [CrossRef]
- Kusminski, C.M.; Bickel, P.E.; Scherer, P.E. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov. 2016, 15, 639–660. [Google Scholar] [CrossRef] [PubMed]
- Alzamendi, A.; Giovambattista, A.; Raschia, A.; Madrid, V.; Gaillard, R.C.; Rebolledo, O.; Gagliardino, J.J.; Spinedi, E. Fructose-rich diet-induced abdominal adipose tissue endocrine dysfunction in normal male rats. Endocrine 2009, 35, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [Green Version]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N.; Pejovic, S.; Zoumakis, E.; Lin, H.M.; Bixler, E.O.; Basta, M.; Fang, J.; Sarrigiannidis, A.; Chrousos, G.P. Daytime napping after a night of sleep loss decreases sleepiness, improves performance, and causes beneficial changes in cortisol and interleukin-6 secretion. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E253–E261. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N. Does obesity play a major role in the pathogenesis of sleep apnoea and its associated manifestations via inflammation, visceral adiposity, and insulin resistance? Arch. Physiol. Biochem. 2008, 114, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Tsai, J.Y.; Villegas-Montoya, C.; Boland, B.B.; Blasier, Z.; Egbejimi, O.; Kueht, M.; Young, M.E. Time-of-day-dependent dietary fat consumption influences multiple cardiometabolic syndrome parameters in mice. Int. J. Obes. 2010, 34, 1589–1598. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R.; Coto-Montes, A.; Poeggeler, B. Circadian rhythms, oxidative stress, and antioxidative defense mechanisms. Chronobiol. Int. 2003, 20, 921–962. [Google Scholar] [CrossRef]
- Teixeira, K.R.C.; Dos Santos, C.P.; de Medeiros, L.A.; Mendes, J.A.; Cunha, T.M.; De Angelis, K.; Penha-Silva, N.; de Oliveira, E.P.; Crispim, C.A. Night workers have lower levels of antioxidant defenses and higher levels of oxidative stress damage when compared to day workers. Sci. Rep. 2019, 9, 4455. [Google Scholar] [CrossRef] [Green Version]
- Punjabi, N.M.; Beamer, B.A. C-reactive protein is associated with sleep disordered breathing independent of adiposity. Sleep 2007, 30, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Lananna, B.V.; Musiek, E.S. The wrinkling of time: Aging, inflammation, oxidative stress, and the circadian clock in neurodegeneration. Neurobiol. Dis. 2020, 139, 104832. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Yang, G.; O’Donnell, J.C.; Hinson, M.D.; McCormack, S.E.; Falk, M.J.; La, P.; Robinson, M.B.; Williams, M.L.; Yohannes, M.T.; et al. The circadian gene Rev-erbalpha improves cellular bioenergetics and provides preconditioning for protection against oxidative stress. Free Radic. Biol. Med. 2016, 93, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, J.F.; Li, X.K.; Li, W.Q.; Gao, Q.; Zhang, Y.; Wang, X.M.; Fu, J.Q.; Cui, S.S.; Qu, J.H.; Zhao, X.; et al. Diurnal oscillations of endogenous H2O2 sustained by p66(Shc) regulate circadian clocks. Nat. Cell Biol. 2019, 21, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.-i. “Clocks” in the NAD World: NAD as a metabolic oscillator for the regulation of metabolism and aging. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 1584–1590. [Google Scholar] [CrossRef] [Green Version]
- Froy, O. Circadian rhythms and obesity in mammals. ISRN Obes. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Froy, O. Circadian rhythms, aging, and life span in mammals. Physiology 2011, 26, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Sandbichler, A.M.; Jansen, B.; Peer, B.A.; Paulitsch, M.; Pelster, B.; Egg, M. Metabolic Plasticity Enables Circadian Adaptation to Acute Hypoxia in Zebrafish Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1159–1174. [Google Scholar] [CrossRef]
- Rutter, J.; Reick, M.; Wu, L.C.; McKnight, S.L. Regulation of clock and NPAS2 DNA binding by the redox state of NAD cofactors. Science 2001, 293, 510–514. [Google Scholar] [CrossRef] [Green Version]
- Rey, G.; Valekunja, U.K.; Feeney, K.A.; Wulund, L.; Milev, N.B.; Stangherlin, A.; Ansel-Bollepalli, L.; Velagapudi, V.; O’Neill, J.S.; Reddy, A.B. The Pentose Phosphate Pathway Regulates the Circadian Clock. Cell Metab. 2016, 24, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Wible, R.S.; Ramanathan, C.; Sutter, C.H.; Olesen, K.M.; Kensler, T.W.; Liu, A.C.; Sutter, T.R. NRF2 regulates core and stabilizing circadian clock loops, coupling redox and timekeeping in Mus musculus. eLife 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Mills, E.L.; Pierce, K.A.; Jedrychowski, M.P.; Garrity, R.; Winther, S.; Vidoni, S.; Yoneshiro, T.; Spinelli, J.B.; Lu, G.Z.; Kazak, L.; et al. Accumulation of succinate controls activation of adipose tissue thermogenesis. Nature 2018, 560, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Kazak, L.; Jedrychowski, M.P.; Lu, G.Z.; Erickson, B.K.; Szpyt, J.; Pierce, K.A.; Laznik-Bogoslavski, D.; Vetrivelan, R.; Clish, C.B.; et al. Mitochondrial ROS regulate thermogenic energy expenditure and sulfenylation of UCP1. Nature 2016, 532, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, B.; Wu, Q.; Sun, C. Melatonin: A novel strategy for prevention of obesity and fat accumulation in peripheral organs through the improvements of circadian rhythms and antioxidative capacity. Melatonin Res. 2020, 3, 58–76. [Google Scholar] [CrossRef]
- Hacışevki, A.; Baba, B. An overview of melatonin as an antioxidant molecule: A biochemical approach. Melatonin Mol. Biol. Clin. Pharm. Approaches 2018, 59–85. [Google Scholar] [CrossRef] [Green Version]
- Tarocco, A.; Caroccia, N.; Morciano, G.; Wieckowski, M.R.; Ancora, G.; Garani, G.; Pinton, P. Melatonin as a master regulator of cell death and inflammation: Molecular mechanisms and clinical implications for newborn care. Cell Death Dis. 2019, 10, 317. [Google Scholar] [CrossRef] [Green Version]
- Fernandez Vazquez, G.; Reiter, R.J.; Agil, A. Melatonin increases brown adipose tissue mass and function in Zucker diabetic fatty rats: Implications for obesity control. J. Pineal Res. 2018, 64, e12472. [Google Scholar] [CrossRef]
- Jimenez-Aranda, A.; Fernandez-Vazquez, G.; Campos, D.; Tassi, M.; Velasco-Perez, L.; Tan, D.X.; Reiter, R.J.; Agil, A. Melatonin induces browning of inguinal white adipose tissue in Zucker diabetic fatty rats. J. Pineal Res. 2013, 55, 416–423. [Google Scholar] [CrossRef]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [Green Version]
- Vollmers, C.; Gill, S.; DiTacchio, L.; Pulivarthy, S.R.; Le, H.D.; Panda, S. Time of feeding and the intrinsic circadian clock drive rhythms in hepatic gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 21453–21458. [Google Scholar] [CrossRef] [Green Version]
- Yamamuro, D.; Takahashi, M.; Nagashima, S.; Wakabayashi, T.; Yamazaki, H.; Takei, A.; Takei, S.; Sakai, K.; Ebihara, K.; Iwasaki, Y.; et al. Peripheral circadian rhythms in the liver and white adipose tissue of mice are attenuated by constant light and restored by time-restricted feeding. PLoS ONE 2020, 15, e0234439. [Google Scholar] [CrossRef]
- Oishi, K.; Kasamatsu, M.; Ishida, N. Gene- and tissue-specific alterations of circadian clock gene expression in streptozotocin-induced diabetic mice under restricted feeding. Biochem. Biophys. Res. Commun. 2004, 317, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Damiola, F.; Le Minh, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Goede, P.; Sen, S.; Oosterman, J.E.; Foppen, E.; Jansen, R.; la Fleur, S.E.; Challet, E.; Kalsbeek, A. Differential effects of diet composition and timing of feeding behavior on rat brown adipose tissue and skeletal muscle peripheral clocks. Neurobiol. Sleep Circadian Rhythm. 2018, 4, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319 e304. [Google Scholar] [CrossRef]
- Heilbronn, L.K.; Panda, S. Alternate-Day Fasting Gets a Safe Bill of Health. Cell Metab. 2019, 30, 411–413. [Google Scholar] [CrossRef]
- Heilbronn, L.K.; Civitarese, A.E.; Bogacka, I.; Smith, S.R.; Hulver, M.; Ravussin, E. Glucose tolerance and skeletal muscle gene expression in response to alternate day fasting. Obes. Res. 2005, 13, 574–581. [Google Scholar] [CrossRef]
- Hoddy, K.K.; Kroeger, C.M.; Trepanowski, J.F.; Barnosky, A.; Bhutani, S.; Varady, K.A. Meal timing during alternate day fasting: Impact on body weight and cardiovascular disease risk in obese adults. Obesity 2014, 22, 2524–2531. [Google Scholar] [CrossRef]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Kroeger, C.M.; Trepanowski, J.F.; Haus, J.M.; Hoddy, K.K.; Calvo, Y. Alternate day fasting for weight loss in normal weight and overweight subjects: A randomized controlled trial. Nutr. J. 2013, 12, 146. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Page, A.J.; Hatzinikolas, G.; Chen, M.; Wittert, G.A.; Heilbronn, L.K. Intermittent Fasting Improves Glucose Tolerance and Promotes Adipose Tissue Remodeling in Male Mice Fed a High-Fat Diet. Endocrinology 2019, 160, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Móez Al-Islam, E.F.; Jahrami, H.A.; Obaideen, A.A.; Madkour, M.I. Impact of diurnal intermittent fasting during Ramadan on inflammatory and oxidative stress markers in healthy people: Systematic review and meta-analysis. J. Nutr. Intermed. Metab. 2019, 15, 18–26. [Google Scholar]
- BaHammam, A.; Alrajeh, M.; Albabtain, M.; Bahammam, S.; Sharif, M. Circadian pattern of sleep, energy expenditure, and body temperature of young healthy men during the intermittent fasting of Ramadan. Appetite 2010, 54, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Madkour, M.I.; El-Serafi, A.T.; Jahrami, H.A.; Sherif, N.M.; Hassan, R.E.; Awadallah, S.; Faris, M.A.E. Ramadan diurnal intermittent fasting modulates SOD2, TFAM, Nrf2, and sirtuins (SIRT1, SIRT3) gene expressions in subjects with overweight and obesity. Diabetes Res. Clin. Pract. 2019, 155, 107801. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hutchison, A.T.; Wittert, G.A.; Thompson, C.H.; Lange, K.; Liu, B.; Heilbronn, L.K. Intermittent Fasting Does Not Uniformly Impact Genes Involved in Circadian Regulation in Women with Obesity. Obesity 2020, 28 (Suppl. 1), S63–S67. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, P.; Chinetti, G.; Fruchart, J.C.; Staels, B. Sorting out the roles of PPAR alpha in energy metabolism and vascular homeostasis. J. Clin. Investig. 2006, 116, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Braissant, O.; Foufelle, F.; Scotto, C.; Dauca, M.; Wahli, W. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology 1996, 137, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Downes, M.; Yu, R.T.; Bookout, A.L.; He, W.; Straume, M.; Mangelsdorf, D.J.; Evans, R.M. Nuclear receptor expression links the circadian clock to metabolism. Cell 2006, 126, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Lee, C.H.; Tiep, S.; Yu, R.T.; Ham, J.; Kang, H.; Evans, R.M. Peroxisome-proliferator-activated receptor delta activates fat metabolism to prevent obesity. Cell 2003, 113, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Kroon, T.; Harms, M.; Maurer, S.; Bonnet, L.; Alexandersson, I.; Lindblom, A.; Ahnmark, A.; Nilsson, D.; Gennemark, P.; O’Mahony, G.; et al. PPARgamma and PPARalpha synergize to induce robust browning of white fat in vivo. Mol. Metab. 2020, 36, 100964. [Google Scholar] [CrossRef]
- Chen, L.; Yang, G. PPARs Integrate the Mammalian Clock and Energy Metabolism. PPAR Res. 2014, 2014, 653017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltowski, J. Endogenous hydrogen sulfide in perivascular adipose tissue: Role in the regulation of vascular tone in physiology and pathology. Can. J. Physiol. Pharm. 2013, 91, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Shi, X.; Wang, H.; Fan, J.; Feng, Y.; Lin, X.; Yang, J.; Cui, Q.; Tang, C.; Xu, G.; et al. Cystathionine gamma lyase-hydrogen sulfide increases peroxisome proliferator-activated receptor gamma activity by sulfhydration at C139 site thereby promoting glucose uptake and lipid storage in adipocytes. Biochim. Biophys. Acta 2016, 1861, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Charoensuksai, P.; Xu, W. PPARs in Rhythmic Metabolic Regulation and Implications in Health and Disease. PPAR Res 2010, 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, K.; Shirai, H.; Ishida, N. CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated receptor alpha (PPARalpha) in mice. Biochem. J. 2005, 386, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Canaple, L.; Rambaud, J.; Dkhissi-Benyahya, O.; Rayet, B.; Tan, N.S.; Michalik, L.; Delaunay, F.; Wahli, W.; Laudet, V. Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol. Endocrinol. 2006, 20, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, K.; Yamada, T.; Tsukita, S.; Takahashi, K.; Ishigaki, Y.; Oka, Y.; Katagiri, H. Obesity alters circadian expressions of molecular clock genes in the brainstem. Brain Res. 2009, 1263, 58–68. [Google Scholar] [CrossRef]
- Yang, G.; Jia, Z.; Aoyagi, T.; McClain, D.; Mortensen, R.M.; Yang, T. Systemic PPARgamma deletion impairs circadian rhythms of behavior and metabolism. PLoS ONE 2012, 7, e38117. [Google Scholar] [CrossRef] [Green Version]
- Gutman, R.; Barnea, M.; Haviv, L.; Chapnik, N.; Froy, O. Peroxisome proliferator-activated receptor alpha (PPARalpha) activation advances locomotor activity and feeding daily rhythms in mice. Int. J. Obes. 2012, 36, 1131–1134. [Google Scholar] [CrossRef] [Green Version]
- Shirai, H.; Oishi, K.; Kudo, T.; Shibata, S.; Ishida, N. PPARalpha is a potential therapeutic target of drugs to treat circadian rhythm sleep disorders. Biochem. Biophys. Res. Commun. 2007, 357, 679–682. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Liu, J.Z.; Hu, J.X.; Chen, H.L.; Li, W.L.; Hai, C.X. Double antioxidant activities of rosiglitazone against high glucose-induced oxidative stress in hepatocyte. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2011, 25, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.F.; Ku, H.C.; Lin, H. PGC-1alpha as a Pivotal Factor in Lipid and Metabolic Regulation. Int. J. Mol. Sci. 2018, 19, 3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lin, J.D. Transcriptional control of circadian metabolic rhythms in the liver. Diabetes Obes. Metab. 2015, 17, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Li, S.; Liu, T.; Borjigin, J.; Lin, J.D. Transcriptional coactivator PGC-1alpha integrates the mammalian clock and energy metabolism. Nature 2007, 447, 477–481. [Google Scholar] [CrossRef]
- Grimaldi, B.; Sassone-Corsi, P. Circadian rhythms: Metabolic clockwork. Nature 2007, 447, 386–387. [Google Scholar] [CrossRef]
- Sonoda, J.; Mehl, I.R.; Chong, L.W.; Nofsinger, R.R.; Evans, R.M. PGC-1beta controls mitochondrial metabolism to modulate circadian activity, adaptive thermogenesis, and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2007, 104, 5223–5228. [Google Scholar] [CrossRef] [Green Version]
- Tanno, M.; Sakamoto, J.; Miura, T.; Shimamoto, K.; Horio, Y. Nucleocytoplasmic shuttling of the NAD+-dependent histone deacetylase SIRT1. J. Biol. Chem. 2007, 282, 6823–6832. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef]
- Donmez, G.; Guarente, L. Aging and disease: Connections to sirtuins. Aging Cell 2010, 9, 285–290. [Google Scholar] [CrossRef]
- Man, A.W.C.; Li, H.; Xia, N. The Role of Sirtuin1 in Regulating Endothelial Function, Arterial Remodeling and Vascular Aging. Front. Physiol. 2019, 10, 1173. [Google Scholar] [CrossRef] [Green Version]
- Gu, P.; Hui, H.; Vanhoutte, P.; Lam, K.; Xu, A. Deletion of SIRT1 in perivascular adipose tissue accelerates obesity-induced endothelial dysfunction. In Proceedings of the 1st ASCEPT-BPS Joint Scientific Meeting, Hong Kong, China, 19–21 May 2015. [Google Scholar]
- Minor, R.K.; Baur, J.A.; Gomes, A.P.; Ward, T.M.; Csiszar, A.; Mercken, E.M.; Abdelmohsen, K.; Shin, Y.K.; Canto, C.; Scheibye-Knudsen, M.; et al. SRT1720 improves survival and healthspan of obese mice. Sci. Rep. 2011, 1, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Li, J.; Xiao, N.; Wang, M.; Kou, J.; Qi, L.; Huang, F.; Liu, B.; Liu, K. Pharmacological activation of AMPK ameliorates perivascular adipose/endothelial dysfunction in a manner interdependent on AMPK and SIRT1. Pharm. Res. 2014, 89, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Fleenor, B.S.; Eng, J.S.; Sindler, A.L.; Pham, B.T.; Kloor, J.D.; Seals, D.R. Superoxide signaling in perivascular adipose tissue promotes age-related artery stiffness. Aging Cell 2014, 13, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.K.; Hwang, M.H.; Luttrell, M.J.; Kim, H.K.; Meade, T.H.; English, M.; Segal, M.S.; Christou, D.D. Higher levels of adiponectin in vascular endothelial cells are associated with greater brachial artery flow-mediated dilation in older adults. Exp. Gerontol. 2015, 63, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Shao, J. SIRT1 regulates adiponectin gene expression through Foxo1-C/enhancer-binding protein alpha transcriptional complex. J. Biol. Chem. 2006, 281, 39915–39924. [Google Scholar] [CrossRef] [Green Version]
- Masri, S.; Orozco-Solis, R.; Aguilar-Arnal, L.; Cervantes, M.; Sassone-Corsi, P. Coupling circadian rhythms of metabolism and chromatin remodelling. Diabetes Obes. Metab. 2015, 17, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell 2008, 134, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Bellet, M.M.; Nakahata, Y.; Boudjelal, M.; Watts, E.; Mossakowska, D.E.; Edwards, K.A.; Cervantes, M.; Astarita, G.; Loh, C.; Ellis, J.L. Pharmacological modulation of circadian rhythms by synthetic activators of the deacetylase SIRT1. Proc. Natl. Acad. Sci. USA 2013, 110, 3333–3338. [Google Scholar] [CrossRef] [Green Version]
- Belden, W.J.; Dunlap, J.C. SIRT1 is a circadian deacetylase for core clock components. Cell 2008, 134, 212–214. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.H.; Zhao, T.; Cui, K.; Hu, G.; Chen, Q.; Chen, W.; Wang, X.W.; Soto-Gutierrez, A.; Zhao, K.; Deng, C.X. Negative reciprocal regulation between Sirt1 and Per2 modulates the circadian clock and aging. Sci. Rep. 2016, 6, 28633. [Google Scholar] [CrossRef]
- Ramsey, K.M.; Yoshino, J.; Brace, C.S.; Abrassart, D.; Kobayashi, Y.; Marcheva, B.; Hong, H.-K.; Chong, J.L.; Buhr, E.D.; Lee, C. Circadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis. Science 2009, 324, 651–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian control of the NAD+ salvage pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Sahar, S.; Masubuchi, S.; Eckel-Mahan, K.; Vollmer, S.; Galla, L.; Ceglia, N.; Masri, S.; Barth, T.K.; Grimaldi, B.; Oluyemi, O. Circadian control of fatty acid elongation by SIRT1 protein-mediated deacetylation of acetyl-coenzyme A synthetase 1. J. Biol. Chem. 2014, 289, 6091–6097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdieri, L.; Zhang, M.; Vancura, A. Regulation of acetyl-CoA homeostasis and global histone acetylation (942.1). FASEB J. 2014, 28, 942-1. [Google Scholar]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [Green Version]
- Arany, Z.; Neinast, M. Branched Chain Amino Acids in Metabolic Disease. Curr. Diabetes Rep. 2018, 18, 76. [Google Scholar] [CrossRef]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yin, Y.; Tan, B.; Kong, X.; Wu, G. Leucine nutrition in animals and humans: mTOR signaling and beyond. Amino Acids 2011, 41, 1185–1193. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Zhenyukh, O.; Civantos, E.; Ruiz-Ortega, M.; Sanchez, M.S.; Vazquez, C.; Peiro, C.; Egido, J.; Mas, S. High concentration of branched-chain amino acids promotes oxidative stress, inflammation and migration of human peripheral blood mononuclear cells via mTORC1 activation. Free Radic. Biol. Med. 2017, 104, 165–177. [Google Scholar] [CrossRef]
- Cao, R.; Lee, B.; Cho, H.Y.; Saklayen, S.; Obrietan, K. Photic regulation of the mTOR signaling pathway in the suprachiasmatic circadian clock. Mol. Cell. Neurosci. 2008, 38, 312–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipton, J.O.; Yuan, E.D.; Boyle, L.M.; Ebrahimi-Fakhari, D.; Kwiatkowski, E.; Nathan, A.; Guttler, T.; Davis, F.; Asara, J.M.; Sahin, M. The Circadian Protein BMAL1 Regulates Translation in Response to S6K1-Mediated Phosphorylation. Cell 2015, 161, 1138–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragert, K.; Bhattacharya, I.; Hall, M.N.; Humar, R.; Battegay, E.; Haas, E. Basal mTORC2 activity and expression of its components display diurnal variation in mouse perivascular adipose tissue. Biochem. Biophys. Res. Commun. 2016, 473, 317–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragert, K.; Bhattacharya, I.; Pellegrini, G.; Seebeck, P.; Azzi, A.; Brown, S.A.; Georgiopoulou, S.; Held, U.; Blyszczuk, P.; Arras, M.; et al. Deletion of Rictor in brain and fat alters peripheral clock gene expression and increases blood pressure. Hypertension 2015, 66, 332–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Dong, W.; Shao, J.; Wang, Y.; Zhou, M.; Sun, H. Branched-Chain Amino Acid Negatively Regulates KLF15 Expression via PI3K-AKT Pathway. Front. Physiol. 2017, 8, 853. [Google Scholar] [CrossRef] [Green Version]
- Matoba, K.; Lu, Y.; Zhang, R.; Chen, E.R.; Sangwung, P.; Wang, B.; Prosdocimo, D.A.; Jain, M.K. Adipose KLF15 Controls Lipid Handling to Adapt to Nutrient Availability. Cell Rep. 2017, 21, 3129–3140. [Google Scholar] [CrossRef]
- Standl, E. GLP-1 receptor agonists and cardiovascular outcomes: An updated synthesis. Lancet Diabetes Endocrinol. 2019, 7, 741–743. [Google Scholar] [CrossRef]
- Han, L.; Yu, Y.; Sun, X.; Wang, B. Exendin-4 directly improves endothelial dysfunction in isolated aortas from obese rats through the cAMP or AMPK–eNOS pathways. Diabetes Res. Clin. Pract. 2012, 97, 453–460. [Google Scholar] [CrossRef]
- Gil-Lozano, M.; Mingomataj, E.L.; Wu, W.K.; Ridout, S.A.; Brubaker, P.L. Circadian secretion of the intestinal hormone GLP-1 by the rodent L cell. Diabetes 2014, 63, 3674–3685. [Google Scholar] [CrossRef] [Green Version]
- Gil-Lozano, M.; Hunter, P.M.; Behan, L.A.; Gladanac, B.; Casper, R.F.; Brubaker, P.L. Short-term sleep deprivation with nocturnal light exposure alters time-dependent glucagon-like peptide-1 and insulin secretion in male volunteers. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E41–E50. [Google Scholar] [CrossRef] [Green Version]
- Cantini, G.; Mannucci, E.; Luconi, M. Perspectives in GLP-1 Research: New Targets, New Receptors. Trends Endocrinol. Metab. 2016, 27, 427–438. [Google Scholar] [CrossRef]
- Han, F.; Hou, N.; Liu, Y.; Huang, N.; Pan, R.; Zhang, X.; Mao, E.; Sun, X. Liraglutide improves vascular dysfunction by regulating a cAMP-independent PKA-AMPK pathway in perivascular adipose tissue in obese mice. Biomed. Pharmacother. 2019, 120, 109537. [Google Scholar] [CrossRef] [PubMed]
- Trzaskalski, N.A.; Fadzeyeva, E.; Mulvihill, E.E. Dipeptidyl Peptidase-4 at the Interface Between Inflammation and Metabolism. Clin. Med. Insights Endocrinol Diabetes 2020, 13, 1179551420912972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.A.; Chowdhury, M.R.H.; Jain, P.; Sagor, M.A.T.; Reza, H.M. DPP-4 inhibitor sitagliptin prevents inflammation and oxidative stress of heart and kidney in two kidney and one clip (2K1C) rats. Diabetol. Metab. Syndr. 2015, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Liu, J.; Tian, X.Y.; Wong, W.T.; Lau, C.W.; Xu, A.; Xu, G.; Ng, C.F.; Yao, X.; Gao, Y. Uncoupling protein-2 mediates DPP-4 inhibitor-induced restoration of endothelial function in hypertension through reducing oxidative stress. Antioxid. Redox Signal. 2014, 21, 1571–1581. [Google Scholar] [CrossRef] [Green Version]
- Kothny, W.; Foley, J.; Kozlovski, P.; Shao, Q.; Gallwitz, B.; Lukashevich, V. Improved glycaemic control with vildagliptin added to insulin, with or without metformin, in patients with type 2 diabetes mellitus. Diabetes Obes. Metab. 2013, 15, 252–257. [Google Scholar] [CrossRef]
- Salim, H.M.; Fukuda, D.; Higashikuni, Y.; Tanaka, K.; Hirata, Y.; Yagi, S.; Soeki, T.; Shimabukuro, M.; Sata, M. Teneligliptin, a dipeptidyl peptidase-4 inhibitor, attenuated pro-inflammatory phenotype of perivascular adipose tissue and inhibited atherogenesis in normoglycemic apolipoprotein-E-deficient mice. Vasc. Pharmacol. 2017, 96, 19–25. [Google Scholar] [CrossRef]
- Ahren, B. Dipeptidyl peptidase-4 inhibitors: Clinical data and clinical implications. Diabetes Care 2007, 30, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Sufiun, A.; Rafiq, K.; Fujisawa, Y.; Rahman, A.; Mori, H.; Nakano, D.; Kobori, H.; Ohmori, K.; Masaki, T.; Kohno, M.; et al. Effect of dipeptidyl peptidase-4 inhibition on circadian blood pressure during the development of salt-dependent hypertension in rats. Hypertens Res. 2015, 38, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell. Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Um, J.H.; Pendergast, J.S.; Springer, D.A.; Foretz, M.; Viollet, B.; Brown, A.; Kim, M.K.; Yamazaki, S.; Chung, J.H. AMPK regulates circadian rhythms in a tissue- and isoform-specific manner. PLoS ONE 2011, 6, e18450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. AMP-Activated Protein Kinase (AMPK) Regulates Energy Metabolism through Modulating Thermogenesis in Adipose Tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almabrouk, T.A.; White, A.D.; Ugusman, A.B.; Skiba, D.S.; Katwan, O.J.; Alganga, H.; Guzik, T.J.; Touyz, R.M.; Salt, I.P.; Kennedy, S. High fat diet attenuates the anticontractile activity of aortic PVAT via a mechanism involving AMPK and reduced adiponectin secretion. Front. Physiol. 2018, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.D.; Lamia, K.A. AMPK at the crossroads of circadian clocks and metabolism. Mol. Cell Endocrinol. 2013, 366, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.P.; Carling, D.; Munday, M.R.; Hardie, D.G. Diurnal rhythm of phosphorylation of rat liver acetyl-CoA carboxylase by the AMP-activated protein kinase, demonstrated using freeze-clamping. Effects of high fat diets. Eur. J. Biochem. 1992, 203, 615–623. [Google Scholar] [CrossRef]
- Busino, L.; Bassermann, F.; Maiolica, A.; Lee, C.; Nolan, P.M.; Godinho, S.I.; Draetta, G.F.; Pagano, M. SCFFbxl3 controls the oscillation of the circadian clock by directing the degradation of cryptochrome proteins. Science 2007, 316, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Eng, G.W.L.; Edison; Virshup, D.M. Site-specific phosphorylation of casein kinase 1 delta (CK1delta) regulates its activity towards the circadian regulator PER2. PLoS ONE 2017, 12, e0177834. [Google Scholar] [CrossRef] [Green Version]
- Barnea, M.; Haviv, L.; Gutman, R.; Chapnik, N.; Madar, Z.; Froy, O. Metformin affects the circadian clock and metabolic rhythms in a tissue-specific manner. Biochim. Biophys. Acta 2012, 1822, 1796–1806. [Google Scholar] [CrossRef] [Green Version]
- Vieira, E.; Nilsson, E.C.; Nerstedt, A.; Ormestad, M.; Long, Y.C.; Garcia-Roves, P.M.; Zierath, J.R.; Mahlapuu, M. Relationship between AMPK and the transcriptional balance of clock-related genes in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1032–E1037. [Google Scholar] [CrossRef]
- Lamia, K.A.; Sachdeva, U.M.; DiTacchio, L.; Williams, E.C.; Alvarez, J.G.; Egan, D.F.; Vasquez, D.S.; Juguilon, H.; Panda, S.; Shaw, R.J.; et al. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science 2009, 326, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, Y. Targeting Endothelial SIRT1 for the Prevention of Arterial Aging. In Endothelial Dysfunction-Old Concepts and New Challenges; IntechOpen: London, UK, 2018. [Google Scholar]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2011, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Fukai, T.; Siegfried, M.R.; Ushio-Fukai, M.; Cheng, Y.; Kojda, G.; Harrison, D.G. Regulation of the vascular extracellular superoxide dismutase by nitric oxide and exercise training. J. Clin. Investig. 2000, 105, 1631–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallerath, T.; Deckert, G.; Ternes, T.; Anderson, H.; Li, H.; Witte, K.; Förstermann, U. Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and activity of endothelial nitric oxide synthase. Circulation 2002, 106, 1652–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, N.; Strand, S.; Schlufter, F.; Siuda, D.; Reifenberg, G.; Kleinert, H.; Förstermann, U.; Li, H. Role of SIRT1 and FOXO factors in eNOS transcriptional activation by resveratrol. Nitric Oxide 2013, 32, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef]
- Paschos, G.K.; FitzGerald, G.A. Circadian clocks and vascular function. Circ. Res. 2010, 106, 833–841. [Google Scholar] [CrossRef]
- Nernpermpisooth, N.; Qiu, S.; Mintz, J.D.; Suvitayavat, W.; Thirawarapan, S.; Rudic, D.R.; Fulton, D.J.; Stepp, D.W. Obesity alters the peripheral circadian clock in the aorta and microcirculation. Microcirculation 2015, 22, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Kunieda, T.; Minamino, T.; Miura, K.; Katsuno, T.; Tateno, K.; Miyauchi, H.; Kaneko, S.; Bradfield, C.A.; FitzGerald, G.A.; Komuro, I. Reduced nitric oxide causes age-associated impairment of circadian rhythmicity. Circ. Res. 2008, 102, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Viswambharan, H.; Carvas, J.M.; Antic, V.; Marecic, A.; Jud, C.; Zaugg, C.E.; Ming, X.F.; Montani, J.P.; Albrecht, U.; Yang, Z. Mutation of the circadian clock gene Per2 alters vascular endothelial function. Circulation 2007, 115, 2188–2195. [Google Scholar] [CrossRef] [Green Version]
- Tunctan, B.; Weigl, Y.; Dotan, A.; Peleg, L.; Zengil, H.; Ashkenazi, I.; Abacioglu, N. Circadian variation of nitric oxide synthase activity in mouse tissue. Chronobiol. Int. 2002, 19, 393–404. [Google Scholar] [CrossRef]
- Anea, C.B.; Cheng, B.; Sharma, S.; Kumar, S.; Caldwell, R.W.; Yao, L.; Ali, M.I.; Merloiu, A.M.; Stepp, D.W.; Black, S.M.; et al. Increased superoxide and endothelial NO synthase uncoupling in blood vessels of Bmal1-knockout mice. Circ. Res. 2012, 111, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastronardi, C.A.; Yu, W.H.; McCann, S.M. Resting and circadian release of nitric oxide is controlled by leptin in male rats. Proc. Natl. Acad. Sci. USA 2002, 99, 5721–5726. [Google Scholar] [CrossRef] [Green Version]
- Kriegsfeld, L.J.; Drazen, D.L.; Nelson, R.J. Circadian organization in male mice lacking the gene for endothelial nitric oxide synthase (eNOS-/-). J. Biol. Rhythm. 2001, 16, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Arraj, M.; Lemmer, B. Endothelial nitric oxide is not involved in circadian rhythm generation of blood pressure: Experiments in wild-type C57 and eNOS knock-out mice under light-dark and free-run conditions. Chronobiol. Int. 2007, 24, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Förstermann, U.; Li, H. Effects of resveratrol on eNOS in the endothelium and the perivascular adipose tissue. Ann. N. Y. Acad. Sci. 2017, 1403, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Galvin, V.B.; Barakat, H.; Kemeny, G.; Macdonald, K.G.; Pories, W.J.; Hickner, R.C. Endothelial nitric oxide synthase content in adipose tissue from obese and lean African American and white American women. Metabolism 2005, 54, 1368–1373. [Google Scholar] [CrossRef]
- Virdis, A.; Duranti, E.; Rossi, C.; Dell’Agnello, U.; Santini, E.; Anselmino, M.; Chiarugi, M.; Taddei, S.; Solini, A. Tumour necrosis factor-alpha participates on the endothelin-1/nitric oxide imbalance in small arteries from obese patients: Role of perivascular adipose tissue. Eur. Heart J. 2015, 36, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Xia, N.; Weisenburger, S.; Koch, E.; Burkart, M.; Reifenberg, G.; Forstermann, U.; Li, H.G. Restoration of perivascular adipose tissue function in diet-induced obese mice without changing bodyweight. Br. J. Pharmacol. 2017, 174, 3443–3453. [Google Scholar] [CrossRef] [Green Version]
- Xia, N.; Horke, S.; Habermeier, A.; Closs, E.I.; Reifenberg, G.; Gericke, A.; Mikhed, Y.; Munzel, T.; Daiber, A.; Forstermann, U.; et al. Uncoupling of Endothelial Nitric Oxide Synthase in Perivascular Adipose Tissue of Diet-Induced Obese Mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi-Utsumi, K.; Gao, B.; Ohinata, H.; Hashimoto, M.; Yamamoto, N.; Kuroshima, A. Enhanced gene expression of endothelial nitric oxide synthase in brown adipose tissue during cold exposure. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2002, 282, R623–R626. [Google Scholar] [CrossRef] [Green Version]
- Csiszar, A.; Labinskyy, N.; Pinto, J.T.; Ballabh, P.; Zhang, H.; Losonczy, G.; Pearson, K.; De Cabo, R.; Pacher, P.; Zhang, C. Resveratrol induces mitochondrial biogenesis in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H13–H20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Ortega, M.; Stucchi, P.; Guzmán-Ruiz, R.; Cano, V.; Arribas, S.; González, M.C.; Ruiz-Gayo, M.; Fernández-Alfonso, M.S.; Somoza, B. Adaptative nitric oxide overproduction in perivascular adipose tissue during early diet-induced obesity. Endocrinology 2010, 151, 3299–3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Li, L.; Shao, Y.; Bai, X.; Bai, T.; Huang, X. Methotrexate improves perivascular adipose tissue/endothelial dysfunction via activation of AMPK/eNOS pathway. Mol. Med. Rep. 2017, 15, 2353–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Molecular clock mechanism in adipose tissue. The molecular clockwork in adipose tissue relies on the dimerization of BMAL1 and CLOCK proteins, whereas the heterodimer of CLOCK:BMAL1 binds to E-box sequence (5′-CACGTG-3′) in the promoters and activates the transcription of Per1–3, Cry1/2, ROR and Rev-ERB. When the cytoplasmic levels of PERs and CRYs reach a certain threshold, PERs and CRYs dimerize and translocate into the nucleus and inhibit the CLOCK:BMAL1-mediated transcription. REV-ERB negatively regulates Bmal1 and Clock expression, whereas RORs positively regulate Bmal1 and Clock expression via ROR response elements (RORE) at the promotor regions. The molecular circadian clock also leads to rhythmic expression of clock-controlled genes (CCGs) through CLOCK:BMAL1-mediated activation of circadian promoter elements including E-boxes, D-boxes, and ROR response elements. BMAL1, brain and muscle Aryl hydrocarbon receptor nuclear translocator--like protein 1; CLOCK, circadian locomotor output cycles kaput; CRY, Cryptochrome; PER, Period; REV-ERB, reverse ERB; ROR, retinoic acid receptor-related orphan receptors.
Figure 1.
Molecular clock mechanism in adipose tissue. The molecular clockwork in adipose tissue relies on the dimerization of BMAL1 and CLOCK proteins, whereas the heterodimer of CLOCK:BMAL1 binds to E-box sequence (5′-CACGTG-3′) in the promoters and activates the transcription of Per1–3, Cry1/2, ROR and Rev-ERB. When the cytoplasmic levels of PERs and CRYs reach a certain threshold, PERs and CRYs dimerize and translocate into the nucleus and inhibit the CLOCK:BMAL1-mediated transcription. REV-ERB negatively regulates Bmal1 and Clock expression, whereas RORs positively regulate Bmal1 and Clock expression via ROR response elements (RORE) at the promotor regions. The molecular circadian clock also leads to rhythmic expression of clock-controlled genes (CCGs) through CLOCK:BMAL1-mediated activation of circadian promoter elements including E-boxes, D-boxes, and ROR response elements. BMAL1, brain and muscle Aryl hydrocarbon receptor nuclear translocator--like protein 1; CLOCK, circadian locomotor output cycles kaput; CRY, Cryptochrome; PER, Period; REV-ERB, reverse ERB; ROR, retinoic acid receptor-related orphan receptors.

Figure 2.
Oxidative stress and circadian rhythm are interrelated in modulating adipose tissue function. Circadian clock regulates the rhythmic oscillation of cellular redox function, while changes in redox state in the cell can influence the clock machinery. Oxidative stress occurs when the production of oxidant molecules exceeds the capacity of antioxidants to defend against these insults. The cellular redox state has been known to oscillate over circadian. Animal models of circadian disruption by hypoxia, clock genes knockout or mutant, night-shift workers and patients with sleeping disorder are reported to have increased oxidative stress and reduced antioxidant defence. Disruption of circadian and increased oxidative stress lead to reduction of adipokine productions, lipid metabolism and browning process in adipose tissue. Metabolic and cardiovascular diseases that are caused by disruption of circadian rhythm and increased oxidative stress can be prevented by antioxidants treatment and time-restricted feeding. ROS, reactive oxygen species; TNF-α, Tumor necrosis factor alpha; IL-6, interleukin 6.
Figure 2.
Oxidative stress and circadian rhythm are interrelated in modulating adipose tissue function. Circadian clock regulates the rhythmic oscillation of cellular redox function, while changes in redox state in the cell can influence the clock machinery. Oxidative stress occurs when the production of oxidant molecules exceeds the capacity of antioxidants to defend against these insults. The cellular redox state has been known to oscillate over circadian. Animal models of circadian disruption by hypoxia, clock genes knockout or mutant, night-shift workers and patients with sleeping disorder are reported to have increased oxidative stress and reduced antioxidant defence. Disruption of circadian and increased oxidative stress lead to reduction of adipokine productions, lipid metabolism and browning process in adipose tissue. Metabolic and cardiovascular diseases that are caused by disruption of circadian rhythm and increased oxidative stress can be prevented by antioxidants treatment and time-restricted feeding. ROS, reactive oxygen species; TNF-α, Tumor necrosis factor alpha; IL-6, interleukin 6.

Figure 3.
Crosstalk between adipose tissue clock and important signaling targets. Considerable evidence has been shown to support the hypothesis that oxidative stress and circadian rhythm are interrelated. Identification of important CGGs and pathway involved in adipose tissue function is important to improve the understanding of clock involvement in the pathophysiology of metabolic diseases. The intact circadian oscillations of these targets are important in maintaining the balance between oxidative status and energy metabolism in the adipose tissue. mTOR, mammalian target of rapamycin; PPARs, peroxisome proliferator-activated receptors; PGC1, PPARγ coactivator 1; GLP-1, glucagon-like petide-1; DPP-IV, dipeptidyl peptidase-IV; eNOS, endothelial nitric oxide synthase; NO, nitric oxide; SIRT1, sirtuin 1; NAD; nicotinamide adenine dinucleotide; NAMPT; nicotinamide phosphoribosyltransferase; AMPK, 5’ adenosine monophosphate-activated protein kinase; KLF15, Krüppel-like factor 15; BCAA, branched-chain amino acid.
Figure 3.
Crosstalk between adipose tissue clock and important signaling targets. Considerable evidence has been shown to support the hypothesis that oxidative stress and circadian rhythm are interrelated. Identification of important CGGs and pathway involved in adipose tissue function is important to improve the understanding of clock involvement in the pathophysiology of metabolic diseases. The intact circadian oscillations of these targets are important in maintaining the balance between oxidative status and energy metabolism in the adipose tissue. mTOR, mammalian target of rapamycin; PPARs, peroxisome proliferator-activated receptors; PGC1, PPARγ coactivator 1; GLP-1, glucagon-like petide-1; DPP-IV, dipeptidyl peptidase-IV; eNOS, endothelial nitric oxide synthase; NO, nitric oxide; SIRT1, sirtuin 1; NAD; nicotinamide adenine dinucleotide; NAMPT; nicotinamide phosphoribosyltransferase; AMPK, 5’ adenosine monophosphate-activated protein kinase; KLF15, Krüppel-like factor 15; BCAA, branched-chain amino acid.


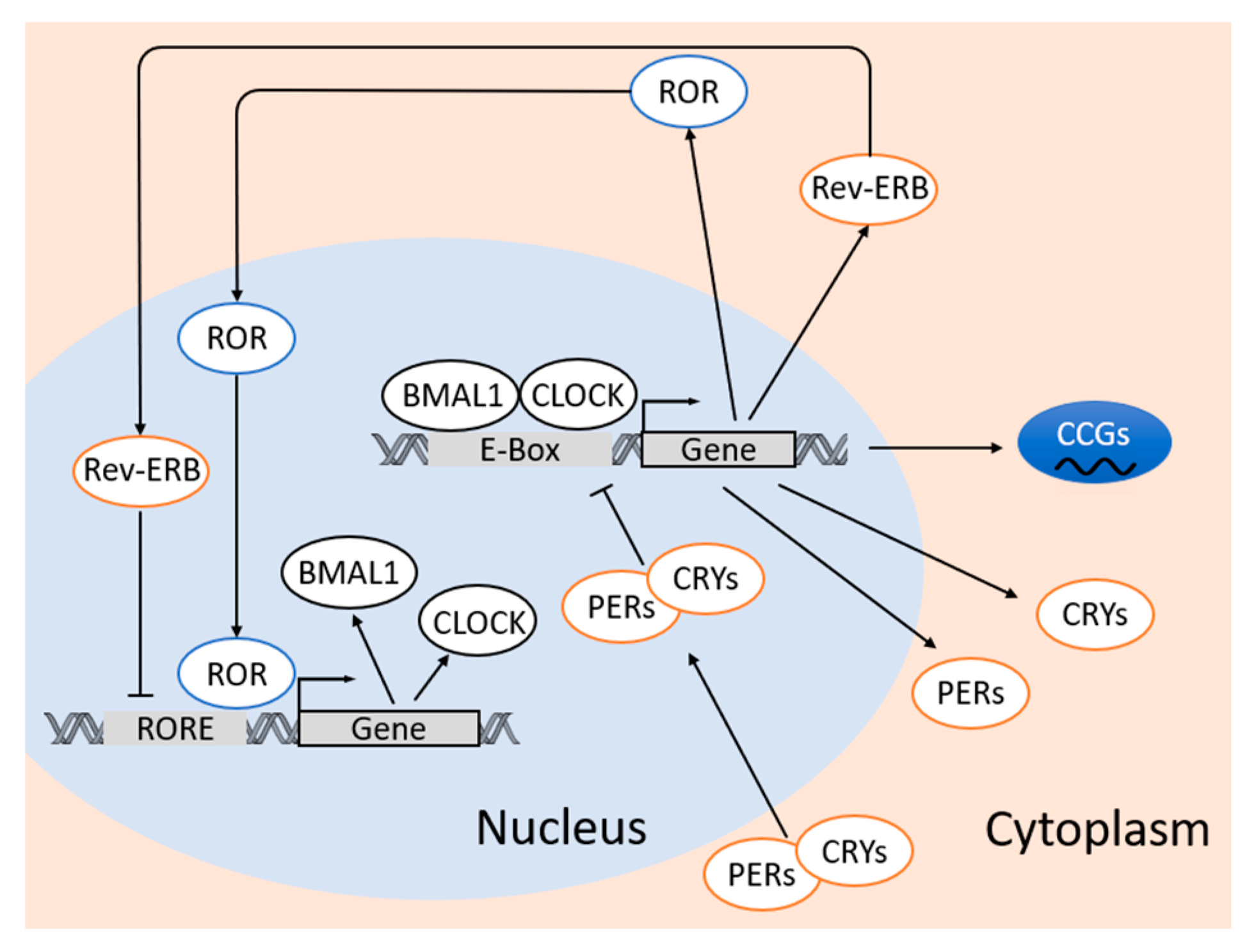{kind=link}
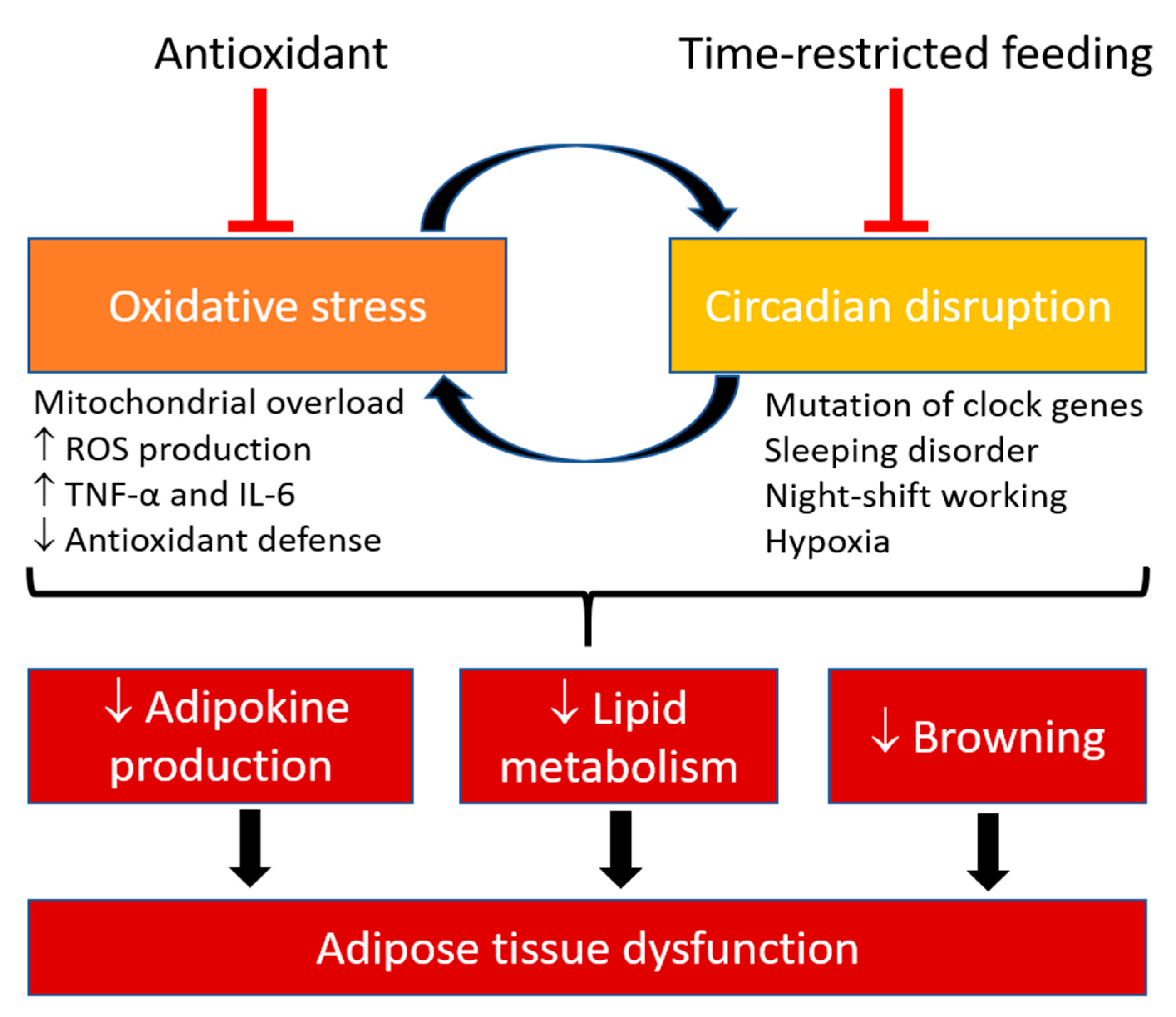{kind=link}
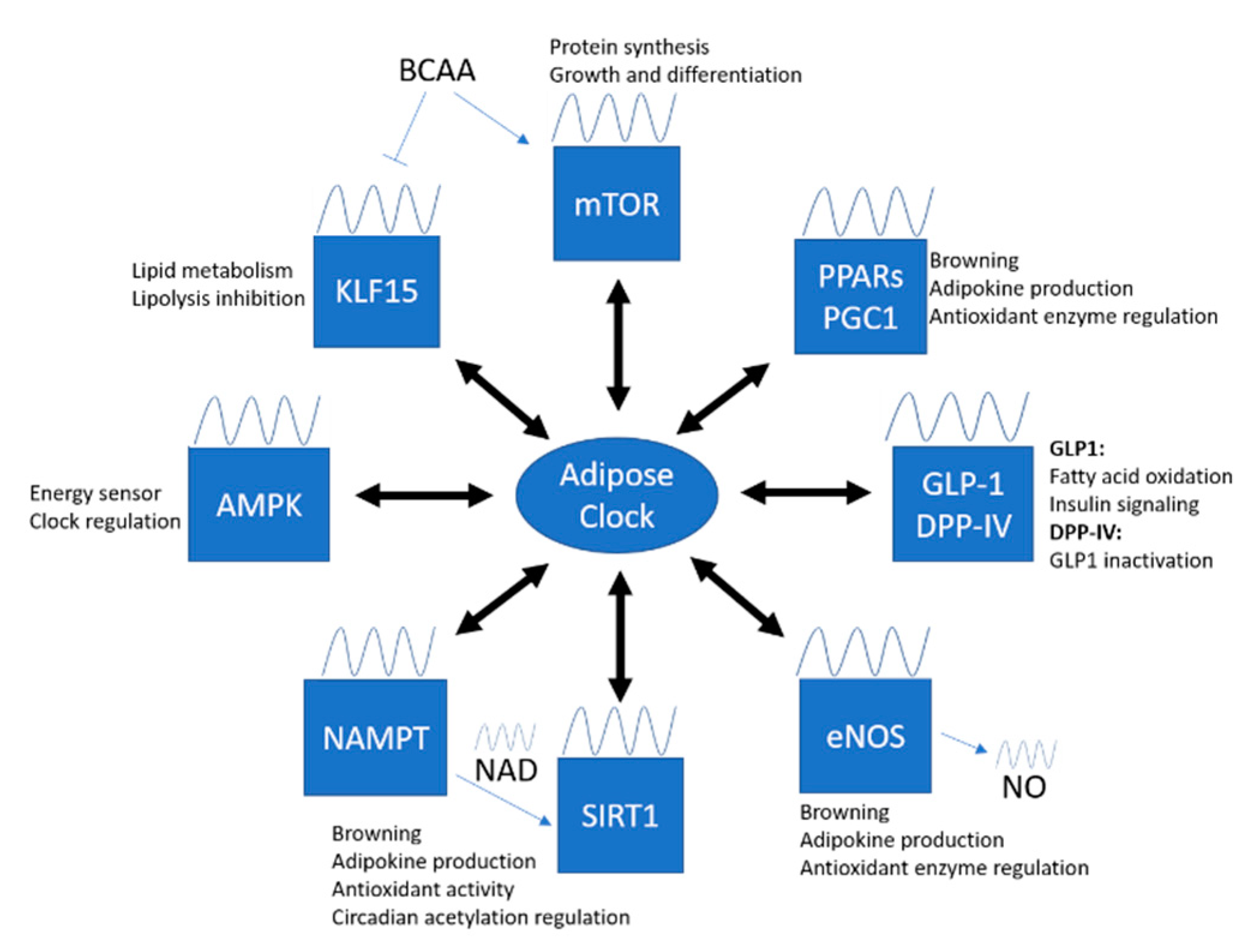{kind=link}
Table 1.
Studies using adipocyte-specific clock genes mutant/knockout mice.
| Gene Mutation/Knockout | Mice Phenotype | Effect on Circadian Rhythm and Oxidative Stress | Refs |
|---|---|---|---|
| Bmal1−/− driven by aP2 promoter | ↑ HFD-induced obesity ↑ HFD-induced adiposity ↑ plasma leptin level ↓ leptin signaling ↓ energy expenditure ↓ polysaturated fatty acids |
| [120] |
| ClockΔ19 driven by aP2 promoter | ↑ young mortality rate ↓ rate of glucose tolerance | Not mentioned | [121] |
| Bmal1−/− driven by UCP1 | ↓ PVAT-induced vasoconstriction | ↓ blood pressure during resting phase ↓ angiotensin and angiotensinogen levels in PVAT | [122] |
Bmal1, brain and muscle aryl hydrocarbon receptor nuclear translocator-like protein 1; aP2, adipocyte protein 2; HFD, high fat diet; BAT, brown adipose tissue; WAT, white adipose tissue; TG, triglyceride; Clock, circadian locomotor output cycles kaput; UCP1, uncoupling protein 1; PVAT, perivascular adipose tissue.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Man, A.W.C.; Xia, N.; Li, H. Circadian Rhythm in Adipose Tissue: Novel Antioxidant Target for Metabolic and Cardiovascular Diseases. Antioxidants 2020, 9, 968. https://doi.org/10.3390/antiox9100968
AMA Style
Man AWC, Xia N, Li H. Circadian Rhythm in Adipose Tissue: Novel Antioxidant Target for Metabolic and Cardiovascular Diseases. Antioxidants. 2020; 9(10):968. https://doi.org/10.3390/antiox9100968
Chicago/Turabian StyleMan, Andy W. C., Ning Xia, and Huige Li. 2020. "Circadian Rhythm in Adipose Tissue: Novel Antioxidant Target for Metabolic and Cardiovascular Diseases" Antioxidants 9, no. 10: 968. https://doi.org/10.3390/antiox9100968
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.