
Abstract
The influence of long-term storage on the evolution of social insect male ejaculates remains a puzzle. Here, we studied the influence of sperm storage on male ejaculates in the stingless bee Scaptotrigona aff. depilis, a species where queens mate with a single male and thus do not trigger sperm competition. We estimated quantitative genetic parameters by sampling males from six colonies and two consecutive generations (n = 172). Heritability for sperm length was lower than heritability estimates for other traits. Coefficients of additive genetic variance (CVa) suggest sperm viability and sperm counts are under directional selection, sperm length under stabilizing selection, and male size under weak selection. Both genetic and phenotypic correlations were weak and not significant, indicating independent trait evolution. Overall, our findings provide evidence of selection acting on social insect ejaculates and bring insights for stingless bee breeding programs.
Similar content being viewed by others


1 Introduction
The evolution of social insect male traits remains poorly understood, as few studies have assessed how sexual selection operates in this group (Abell et al. 1999; Boomsma et al. 2005; Couvillon et al. 2010; Jaffé and Moritz 2010; Koffler et al. 2016). Earlier attempts have been made to understand how competition for mating shapes male morphology (Abell et al. 1999, Couvillon et al. 2010). However, ejaculate evolution is expected to be under more specific selective pressures. Males of the haplodiploid social Hymenoptera (ants, bees, and wasps) originate from unfertilized eggs, so to achieve fitness gains, these haploid males must sire reproductive females (queens). This implies that most sperm are used for the production of sterile worker offspring until colonies are large enough to start producing queens (Oster and Wilson 1978). Male fitness gains can also be realized indirectly through the production of males by unmated workers, albeit worker reproduction is rare or absent in some groups (Wenseleers and Ratnieks 2006). As Hymenoptera queens mate only once in life and store sperm in a specialized organ (the spermatheca) for use in egg fertilization throughout their lives (Al-Lawati et al. 2009), ejaculate quality is expected to be under strong selection (Boomsma et al. 2005). Selection is also expected to shape more competitive ejaculates in species where queens mate with multiple males (den Boer et al. 2010; Fitzpatrick and Baer 2011), although selective pressures resulting from sperm competition are likely to differ. Studies aiming at disentangling these selective pressures and assessing the effect of long-term limited storage on male ejaculate should thus focus on singly mating (monandrous) species (Simmons 2001).
Stingless bees (Meliponini) comprise a diverse group with more than 500 social species exhibiting monandrous long-lived queens (Ascher and Pickering 2019; Carvalho-Zilse and Kerr 2004; Peters et al. 1999). Before mating, males congregate in large aggregations with hundreds of males (Engels and Engels 1984), where persistence at the aggregation is suggested to select more competitive males (Koffler et al. 2016, Figure. 1). Male traits involved in copula were positively related to fertilization traits in the stingless bee Scaptotrigona aff. depilis (Koffler et al. 2016). Specifically, males that persisted longer at the aggregations were smaller and showed higher sperm viability, shorter sperm length, and higher sperm length variation. However, to investigate patterns of selection acting on these traits, phenotypic and genotypic variation must be assessed, since only traits exhibiting additive genetic variance may respond to selection (Houle 1992; Simmons and Kotiaho 2002). Quantitative genetic studies also allow investigating the genetic architecture of traits, which is particularly interesting for S. aff. depilis males, since most studied traits were not correlated.

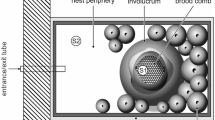
Aggregation of Scaptotrigona aff. depilis males over a colony of the same species (individual male on detail).
Whereas selection acts on traits that exhibit some degree of additive genetic variance, the ratio between genetic and environmental variance indicates how selection operates (Lynch and Walsh 1998). Studies using different animal groups revealed that fitness-related traits show lower heritability estimates (rate between additive genetic variance and total phenotypic variance) (Koffler et al. 2017; Mousseau and Roff 1987; Postma 2014), which was interpreted as a reduction in additive genetic variance due to fixation of optimum condition traits. However, when calculating coefficients of variation (standardizing the variance by trait mean), fitness-related traits exhibited high coefficients of additive genetic variance (CVa) (Houle 1992; Merilä and Sheldon 2000; Pomiankowski and Møller 1995). Coefficients of residual variance (CVr), which comprise non-additive genetic variance and environmental variance, were also high for these traits, thus resulting in low heritability estimates. The analysis of coefficients of variance allows estimating selection patterns acting on each trait, as traits exhibiting higher CVa are expected to be under directional selection, while traits with low CVa are expected to be under stabilizing selection (Houle 1992). In addition, when considering multiple traits, the occurrence of genetic correlations may favor or constrain trait evolution, requiring thus multivariate analyses.
Aiming to assess the incidence of selection on male ejaculates in a monandrous social insect, here we estimated quantitative genetic parameters (heritability, CVa, CVr, and genetic correlations) for males of the stingless bee S. aff. depilis. Mixed models were employed for parameter estimation since they provide more accurate estimates and do not require fixed breeding designs (Postma 2014; Wilson et al. 2010). This approach facilitates the inclusion of individuals scattered across complex pedigrees, which is especially well suited for studying social insect males, since they are haploid and do not produce direct male offspring. Following Koffler et al. (2016), we analyzed sperm viability, sperm counts, sperm length, and male size. Male size was analyzed to assess selection patterns on a morphological trait and to compare it with selection on ejaculate traits.
2 Material and methods
2.1 Sampling design and male traits
To obtain a gradient of relatedness coefficients from individuals sampled from different colonies and generations, we gathered six colonies of Scaptotrigona aff. depilis. Queens were removed and males were sampled before queen removal (first generation) and after queen supersedure (second generation) Fig. 2. Emerging males were collected and transferred to wooden boxes containing recently emerged workers from the same colony, pollen, and sugar syrup. These wooden boxes were kept inside an incubator at 28 °C (± 1°C), and food was replaced three times per week. Male sampling for the first generation was performed between May and June 2016, and between August 2016 and February 2017 for the second generation. Even though our design does not allow controlling for maternal effects, spermatogenesis and male size in Hymenoptera are determined during development (Boomsma et al. 2005), so we do not expect them to be affected by rearing conditions. Sperm viability, however, might be influenced by conditions as age and temperature (Sturup et al. 2013), which were standardized.

Sampling design showing the two generations from which males were collected. In each generation, males could have been produced by queens or workers
Males were dissected 14 to 16 days after emergence, allowing male maturation. Between 9 and 20 males were sampled from each colony in each generation (n = 172, Online Resource 1: Table I). To assess the ejaculate quality, we measured sperm viability (relative proportion of live sperm cells), sperm counts, and sperm length, following the protocols adapted to stingless bees (Meneses et al. 2014). Male’s seminal vesicles were dissected with forceps and placed in 120 μL Hayes solution (pH = 8.7). Sperm viability was measured staining 5 μL of the sperm solution with the LIVE DEAD® Sperm Viability Kit and classifying 400 cells as live (green), dead (red), or dying (green and red). Ten microliters of the sperm solution was spread on a slide, air dried, and stained with DAPI. Sperm length was measured using photographs taken at × 20 magnification. Sperm counts were estimated by diluting the sperm solution 10,000 times in Hayes solution and samples of 1 μL were placed on a slide, air dried, and stained with DAPI. For each male, we measured three sperm count replicates and five sperm cells, and mean values were used in the analyses. When dissection did not allow extracting all semen effectively (e.g., vesicles ruptured during dissection), the sample was not used for measuring sperm traits. Intertegular distance (the shortest distance between the bases of the tegulae) was used as an estimate of male size (Cane 1987).
2.2 Data analyses
Heritability was estimated using the REMLF90 software, which relies on restricted maximum likelihood (REML) (Misztal et al. 2002). This analysis allows the estimation of the unknown additive genetic variances and covariances, given the phenotype and a relationship matrix (see examples of parameter and input files in Online Resource 3). Even though this approach has already been used to evaluate colony traits in social insects (Zakour et al. 2012), this is its first application in stingless bees or male characteristics. In our case, phenotype was set as the response variable and colony was included as fixed factor. Data were transformed to achieve normality and aid model convergence (sperm viability: logit + 10, sperm counts: log/100000). Maximum number of rounds was fixed at 100. Overfitting was assessed by systematically excluding part of the dataset (data from each colony, both generations, Online Resource 1: Table 1) and running the analyses to check the variation in parameter estimation.
To build the relationship matrix, we included the relatedness between all pairs of sampled males. Males could be produced by queens or workers, since unmated workers of S. aff. depilis can also lay reproductive eggs. Since we could not distinguish between queen or worker-produced males in each generation, we calculated weighted coefficients of relatedness, accounting for the probability of a male being produced by a queen or a worker. In each generation, all possible pairs of males were considered, with the respective relatedness coefficients: both males being worker sons (assuming different workers, r = 0.375), both males being queen’s sons (r = 0.5), and one being a worker son and one a queen son (r = 0.25) (Figure. 2). The same weighting was performed between generations and possible male pairs were both males being worker sons (assuming different workers, r = 0.1875), both males being queen sons (r = 0.25), queen son in the first generation and worker son in the second generation (r = 0.125), and worker son in the first generation and queen son in the second generation (r = 0.375) (Fig.2, Online Resource 1: Table II). Three distinct relationship scenarios were considered, following a study by Paxton et al. (2003), who studied male maternity in ten colonies of this species: (A) 13% of males produced by workers (mean proportion), (B) no worker reproduction (minimum observed), and (C) 51% of males produced by workers (maximum observed) (Online Resource 1: Table 2).
Heritability estimates were calculated as the ratio between genetic variance and phenotypic variance (genetic + residual variance). Additive genetic coefficients of variance (CVa) and residual coefficients of variance (CVr) were calculated following Houle (1992). Additionally, genetic correlations were obtained following the expression
where cov12 is the genetic covariance between traits 1 and 2, and var1 and var2 are the genetic variances of traits 1 and 2, respectively. Standard errors were approximated dividing the standard deviation from different estimates obtained using partial datasets (see overfitting analysis above) by the square root of the number of estimates. A likelihood ratio test was performed to test if genetic correlations were different from zero, by constraining to zero covariances between each pair of traits (Windig 1997). Phenotypic correlations were computed, using Spearman’s correlation.
3 Results
Measuring ejaculate and morphological traits and estimating their quantitative genetics parameters allowed us to infer selection patterns acting on Scaptotrigona aff. depilis males. Mean sperm viability was 56% (SD = 0.15), mean sperm counts were 1,372,245 (SD = 252,905), mean sperm length was 87.35 um (SD = 3.76), and mean male size was 1.92 mm (0.06) (Table 1). The assessed traits showed high heritability estimates, with the exception of sperm length which exhibited lower heritability (Table 1). Even though standard errors could not be computed due to low sample sizes, overfitting was not observed. Sperm viability and sperm counts showed high additive genetics coefficients of variance (CVa), while sperm length and male size showed lower CVa. Residual coefficients of variance (CVr) for male size were low; thus, heritability was high, while sperm length showed higher CVr and lower heritability. Distinct scenarios of worker reproduction resulted in similar estimates for all traits and no significant difference was observed between scenarios (scenario A (13%) × scenario B (null): LRT = 0, p = 1; scenario A (13%) × scenario C (51%): LRT = 0.001, p = 1; scenario B (null) × scenario C (51%): LRT = 0.001, p = 1).
In general, genetic and phenotypic correlations were weak (Table 2). Similar to heritability estimates, genetic correlations were not influenced by the amount of worker reproduction (Online Resource 1: Table 3), so only the results for the scenario A (13% of males produced by workers, Online Resource 1: Table 2) are reported. Both genetic and phenotypic correlations were not significantly different from zero.
4 Discussion
Our comparative analyses of Scaptotrigona aff. depilis male traits revealed that sperm length exhibits lower heritability, while sperm viability, sperm counts, and male size exhibit higher heritability estimates. Patterns of coefficients of variance for these traits suggest directional selection acting on sperm viability and sperm counts, stabilizing selection on sperm length, and weak selection on male size. Additionally, we found weak albeit not significant genetic correlations among these four male traits. Even though we did not assess coefficients of relatedness to estimate these parameters, employing three plausible relatedness scenarios did not alter our results, indicating that our estimates are robust. Despite accounting for the large variation in worker reproduction in this species (0 to 51% of males being worker sons), weighted coefficients of relatedness were similar among scenarios, probably due to the overall high relatedness in social insect colonies. Even though our dataset was restricted to males from few colonies, which might overestimate parameters, we show that REML is a suitable tool to estimate quantitative genetic parameters in groups that exhibit complex relationship structure. Since BLUPF90 is a family of programs developed in FORTRAN 90/95 and requires specific input files, we highlight that a similar approach can be obtained using the package ASReml-R on R (Butler et al. 2009).
High heritability estimates may be caused by a large additive genetic component, as was found to be the case for sperm viability and sperm counts. Both high CVa and CVr indicate sperm viability and sperm counts as life history traits that contribute to male fitness. High CVr for fitness components might be explained by the interaction of genetic and environmental effects during lifetime of the organism, or by selection favoring or being neutral on alleles affecting variance of fitness traits (Houle 1992). In addition, high CVa suggests these traits are under directional selection (Houle 1992; Pomiankowski and Møller 1995), confirming previous findings that sperm viability is under sexual selection in S. aff. depilis (Koffler et al. 2016). Directional selection was also suggested for sperm viability and sperm counts in the cockroach Nauphoeta cinerea (Moore et al. 2004) and ejaculate volume in the dung beetle Onthophagus taurus (Simmons and Kotiaho 2002), which may be related to sperm competition since these are polyandrous species. Likewise, sperm viability was also related to long-term sperm storage in the leafcutter ant Atta colombica (Stürup et al. 2011), another polyandrous species (Baer et al. 2009). In contrast, S. aff. depilis is a monandrous social bee where no sperm competition occurs; thus, directional selection in sperm viability and sperm counts is likely related to long-term storage of live sperm (Koffler et al. 2016). Similar to our results, heritability of sperm counts in A. colombica was high (Stürup et al. 2011).
In contrast, sperm length exhibited lower heritability, which is typical of fitness-related traits (Koffler et al. 2017; Mousseau and Roff 1987; Postma 2014). Most studies, however, have found high heritability estimates for sperm length in other species (Baer et al. 2006; Morrow and Gage 2001; Simmons and Kotiaho 2002; Stürup et al. 2011). Usually, mean sperm length is used in parameter estimation, which may inflate estimates. For instance, considering within male variance, which accounted for most of the observed variance in sperm length of Bombus terrestris, resulted in a lower heritability estimate (Baer et al. 2006). Thus, the heritability estimate reported here could be even lower, since within male variance could not be included in our multi-trait analyses. Sperm length also showed low CVa, suggesting this trait is under stabilizing selection (Houle 1992; Pomiankowski and Møller 1995). This result also confirms previous findings that sperm length presents low evolvability, even in polyandrous species where sperm length could contribute to sperm competition (Baer et al. 2006; Dobler and Hosken 2010; Pitnick and Miller 2000; Simmons and Kotiaho 2002). Stabilizing selection on sperm length may thus be related to a constraint imposed by the female, since the size of the sperm-storage organ and sperm length were found to be correlated in Drosophila (Pitnick et al. 1999). In S. aff. depilis, competitive males exhibit shorter sperm which would allow the storage of higher amounts of sperm by the queen (Koffler et al. 2016), since the spermatheca presents a thick cuticle and thus has a limited volume (da Cruz-Landim et al. 2003). On the other hand, production constraints may also affect sperm length. In fungus-growing ants, most variance on sperm length was explained by sexual dimorphism instead of storage constraints, and in species with small males relative to queens, sperm was shorter (Baer et al. 2009). Since sperm production is restricted in small males by size or resources, to produce a certain amount of sperm, sperm cells must be shorter (Baer et al. 2009), a process that is also compatible with stabilizing selection on sperm length. This pattern may also hold for stingless bees, which also exhibit sexual dimorphism with males small relative to queens, even though not so extreme as in ants.
Even though male size showed high heritability, CVa was low. Morphological traits, opposite to life history traits, tend to exhibit high heritability estimates (Koffler et al. 2017; Mousseau and Roff 1987; Postma 2014). For instance, high heritability estimates for male body size were also found for other social insects and solitary bees (Koffler et al. 2017). Both low CVa and CVr suggest male size is under weak selection, with CVa magnitude similar to other studies (Moore et al. 2004; Simmons and Kotiaho 2002). In addition, low CVa would confer lower evolvability to this trait (Houle 1992). We nevertheless expected that male size would show patters of strong selection in our study species, because body size was related to competitive ability in male aggregations (Koffler et al. 2016). This result highlights the importance of combining the analyses of phenotypic and genotypic variance and estimating levels of additive genetic variance in traits suspected to be under selection (Houle 1992; Simmons and Kotiaho 2002).
Although associations between traits are usually assessed by phenotypic correlations, these estimates may not represent evolutionary trait integration, since phenotypic and genetic correlations may differ (Moore et al. 2004). However, the four analyzed male traits in our study were neither phenotypically nor genetically correlated. Even though we found evidence that sperm traits are under selection in S. aff. depilis, absence of genetic correlations between traits suggests independence in trait evolution. This result contrasted to general predictions for ejaculate evolution, since ejaculate traits are expected to function as an integrated unit to assure fitness (Moore et al. 2004). However, other studies also did not observe genetic correlations between ejaculate traits (Simmons and Kotiaho 2002; Stürup et al. 2011), and negative genetic correlations were only found when there was a clear trade-off between traits related to female manipulation and sperm competition (Moore et al. 2004). Body size was not genetically correlated to ejaculate traits also in N. cinerea and O. taurus (Moore et al. 2004; Simmons and Kotiaho 2002); however, genetic correlations between male condition and ejaculate traits suggest condition dependence on ejaculate evolution (Simmons and Kotiaho 2002). In other social insects, phenotypic correlations were found between male size and sperm length; however, correlations were either weak or showed variation among related species (Baer et al. 2003; Stürup et al. 2011).
Our study revealed selective forces related to long-term sperm storage acting on ejaculate traits in a monandrous social bee. Given the scarcity of studies assessing quantitative genetic estimates of male traits in social insects, we cannot extrapolate our findings to the entire group. Stabilizing selection inferred for sperm length may not only reflect storage constraints but also physiological trade-offs. Our work also provides important insights for stingless bee breeding programs, as males could be selected on the basis of ejaculate traits to enhance queen fecundity and speed up genetic improvement programs, since sperm quality is related to colony performance (Pettis et al. 2016). Specifically, sperm viability and sperm counts, which we found to be under directional selection, are good candidates for breeding. On the other hand, sperm length would not be susceptible to selection, since this trait exhibits low levels of additive genetic variance and thus low evolvability.
Data availability
We have included our complete dataset as supplementary material.
References
Abell, A.J., Cole, B.J., Reyes, R., Wiernasz, D.C., 1999. Sexual selection on body size and shape in the western harvester ant, Pogonomyrmex occidentalis Cresson, Evolution (N. Y). 53, 535–545. https://doi.org/10.1111/j.1558-5646.1999.tb03788.x
Al-Lawati, H., Kamp, G., Bienefeld, K., 2009. Characteristics of the spermathecal contents of old and young honeybee queens. J. Insect Physiol. 55, 117–122. https://doi.org/10.1016/j.jinsphys.2008.10.010
Ascher, J.S., Pickering, J., 2019. Discover Life bee species guide and world checklist (Hymenoptera: Apoidea: Anthophila) [WWW Document]. URL http://www.discoverlife.org/mp/20q?guide=Apoidea_species&flags=HAS
Baer, B., Schmid-Hempel, P., Høeg, J.T., Boomsma, J.J., 2003. Sperm length, sperm storage and mating system characteristics in bumblebees. Insectes Soc. 50, 101–108. https://doi.org/10.1007/s00040-003-0641-0
Baer, B., de Jong, G., Schmid-Hempel, R., Schmid-Hempel, P., Høeg, J.T., Boomsma, J.J., 2006. Heritability of sperm length in the bumblebee Bombus terrestris. Genetica 127, 11–23. https://doi.org/10.1007/s10709-005-2157-1
Baer, B., Dijkstra, M.B., Mueller, U.G., Nash, D.R., Boomsma, J.J., 2009. Sperm length evolution in the fungus-growing ants. Behav. Ecol. 20, 38–45. https://doi.org/10.1093/beheco/arn112
Boomsma, J.J., Baer, B., Heinze, J., 2005. The evolution of male traits in social insects. Annu. Rev. Entomol. 50, 395–420. https://doi.org/10.1146/annurev.ento.50.071803.130416
Butler, D.G., Cullis, B.R., Gilmour, A.R., Gogel, B.J., 2009. ASReml-R reference manual. The State of Queensland, Department of Primary Industries and Fisheries, Brisbane.
Cane, J.H., 1987. Estimation of bee size using intertegular span (Apoidea). J. Kansas Entomol. Soc. 60, 145–147.
Carvalho-Zilse, G.A., Kerr, W.E., 2004. Natural substitutions of queens and flight distance of males in tiuba (Melipona compressipes fasciculata Smith, 1854) and uruçu (Melipona scutellaris Latreille, 1811) (Apidae, Meliponini). Acta Amaz. 34, 649–652.
Couvillon, M.J., Hughes, W.O.H., Perez-Sato, J.A., Martin, S.J., Roy, G.G.F., Ratnieks, F.L.W., 2010. Sexual selection in honey bees: colony variation and the importance of size in male mating success. Behav. Ecol. 21, 520–525. https://doi.org/10.1093/beheco/arq016
Cruz-Landim, C. da, Yabuki, A.T., Iamonte, M., 2003. Ultrastructure of the spermatheca of Melipona bicolor bicolor Lep. (Hymenoptera, Apinae, Meliponini). Biosci. J. 19, 57–64.
den Boer, S.P.A., Baer, B., Boomsma, J.J., 2010. Seminal Fluid Mediates Ejaculate Competition in Social Insects. Science (80-. ). 327, 1506–1509. https://doi.org/10.1126/science.1184709
Dobler, R., Hosken, D.J., 2010. Response to selection and realized heritability of sperm length in the yellow dung fly (Scathophaga stercoraria). Heredity (Edinb). 104, 61–6. https://doi.org/10.1038/hdy.2009.93
Engels, E., Engels, W., 1984. Drohnen-ansammlungen bei nestern der stachellosen Biene Scaptotrigona postica. Apidologie 15, 315–328.
Fitzpatrick, J.L., Baer, B., 2011. Polyandry reduces sperm length in social insects. Evolution (N. Y). 65, 3006–3012. https://doi.org/10.1111/j.1558-5646.2011.01343.x
Houle, D., 1992. Comparing evolvability and variability of quantitative traits. Genetics 130, 195–204. https://doi.org/citeulike-article-id:10041224
Jaffé, R., Moritz, R.F.A., 2010. Mating flights select for symmetry in honeybee drones (Apis mellifera). Naturwissenschaften 97, 337–343. https://doi.org/10.1007/s00114-009-0638-2
Koffler, S., Meneses, H.M., Kleinert, A. de M. P., Jaffé, R., 2016. Competitive males have higher quality sperm in a monogamous social bee. BMC Evol. Biol. 16, 195. https://doi.org/10.1186/s12862-016-0765-2
Koffler, S., Kleinert, A. de M. P., Jaffé, R., 2017. Quantitative conservation genetics of wild and managed bees. Conserv. Genet. 18, 689–700. https://doi.org/10.1007/s10592-016-0904-8
Lynch, M., Walsh, B., 1998. Genetics and analysis of quantitative traits. Sinauer
Meneses, H.M., Koffler, S., Freitas, B.M., Imperatriz-Fonseca, V.L., Jaffé, R., 2014. Assessing Sperm Quality in Stingless Bees. Sociobiology 61, 517–522.
Merilä, J., Sheldon, B.C., 2000. Lifetime Reproductive Success and Heritability in Nature. Am. Nat. 155, 301–310. https://doi.org/10.1086/303330
Misztal, I., Tsuruta, S., Strabel, T., Auvray, B., Druet, T., Lee, D.H., 2002. BLUF90 and related programs (BGF90), in: Proc. 7th World Congr. Genet. Appl. Livest. Prod. Montpelier, France.
Moore, P.J., Harris, W.E., Montrose, V.T., Levin, D., Moore, A.J., 2004. Constraints on evolution and postcopulatory sexual selection: Trade-offs among ejaculate characteristics. Evolution (N. Y). 58, 1773–1780. https://doi.org/10.1554/04-116
Morrow, E.H., Gage, M.J.G., 2001. Artificial selection and heritability of sperm length in Gryllus bimaculatus. Heredity (Edinb). 79, 356–362.
Mousseau, T.A., Roff, D.A., 1987. Natural selection and the heritability of fitness components. Heredity (Edinb). 59, 181–197. https://doi.org/10.1038/hdy.1987.113
Oster, G.F., Wilson, E.O., 1978. Caste and ecology in the social insects, Monographs in Population Biology. Princeton University Press, Princeton, N.J.
Paxton, R.J., Bego, L.R., Shah, M.M., Mateus, S., 2003. Low mating frequency of queens in the stingless bee Scaptotrigona postica and worker maternity of males. Behav. Ecol. Sociobiol. 53, 174–181. https://doi.org/10.1007/s00265-002-0561-4
Peters, J.M., Queller, D.C., Imperatriz-Fonseca, V.L., Roubik, D.W., Strassmann, J.E., 1999. Mate number, kin selection and social conflicts in stingless bees and honeybees. Proc. R. Soc. London Ser. B-Biological Sci. 266, 379–384.
Pettis, J.S., Rice, N., Joselow, K., Van Engelsdorp, D., Chaimanee, V., 2016. Colony failure linked to low sperm viability in honey bee (Apis mellifera) queens and an exploration of potential causative factors. PLoS One 11, e0147220. https://doi.org/10.1371/journal.pone.0147220
Pitnick, S., Miller, G.T., 2000. Correlated response in reproductive and life history traits to selection on testis length in Drosophila hydei. Heredity (Edinb). 84, 416–426. https://doi.org/10.1046/j.1365-2540.2000.00679.x
Pitnick, S., Markow, T., Spicer, G.S., 1999. Evolution of multiple kinds of female sperm-storage organs in Drosophila. Evolution (N. Y). 53, 1804–1822. https://doi.org/10.1111/j.1558-5646.1999.tb04564.x
Pomiankowski, A., Møller, A.P., 1995. A resolution of the lek paradox. Proc. R. Soc. London. Ser. B Biol. Sci. 260, 21–29. https://doi.org/10.1098/rspb.1995.0054
Postma, E., 2014. Four decades of estimating heritabilities in wild vertebrate populations: improved methods, more data, better estimates?, in: Quantitative Genetics in the Wild. pp. 16–33.
Simmons, L.W., 2001. Sperm competition and its evolutionary consequences in the insects, Monographs in behavior and ecology. Princeton University Press, Princeton, USA.
Simmons, L.W., Kotiaho, J.S., 2002. Evolution of ejaculates: patterns of phenotypic and genotypic variation and condition dependence in sperm competition traits. Evolution (N. Y). 56, 1622–1631. https://doi.org/10.1111/j.0014-3820.2002.tb01474.x
Stürup, M., den Boer, S., Nash, D., Boomsma, J., Baer, B., 2011. Variation in male body size and reproductive allocation in the leafcutter ant Atta colombica: estimating variance components and possible trade-offs. Insectes Soc. 58, 47–55. https://doi.org/10.1007/s00040-010-0115-0
Sturup, M., Baer-Imhoof, B., Nash, D.R., Boomsma, J.J., Baer, B., 2013. When every sperm counts: factors affecting male fertility in the honeybee Apis mellifera. Behav. Ecol. 24, 1192–1198. https://doi.org/10.1093/beheco/art049
Wenseleers, T., Ratnieks, F.L.W., 2006. Comparative Analysis of Worker Reproduction and Policing in Eusocial Hymenoptera Supports Relatedness Theory. Am. Nat. 168, E163–E179. https://doi.org/10.1086/508619
Wilson, A.J., Réale, D., Clements, M.N., Morrissey, M.M., Postma, E., Walling, C.A., Kruuk, L.E.B., Nussey, D.H., 2010. An ecologist’s guide to the animal model. J. Anim. Ecol. 79, 13–26. https://doi.org/10.1111/j.1365-2656.2009.01639.x
Windig, J.J., 1997. The calculation and significance testing of genetic correlations across environments. J. Evol. Biol. 10, 853–874. https://doi.org/10.1007/s000360050058
Zakour, M.K., Ehrhardt, K., Bienefeld, K., 2012. First estimate of genetic parameters for the Syrian honey bee Apis mellifera syriaca. Apidologie 43, 600–607. https://doi.org/10.1007/s13592-012-0130-4
Acknowledgments
We thank Gracie Zhipei Du, Manuel Plate, and Richard Bernstein for helping implementing the quantitative genetic analyses, Eduardo Santos and Denise Alves for comments during the development of this study, Laís Flores for editing Fig. 2, Prof. Ana Castrucci for providing access to the fluorescence microscope, and two anonymous reviewers for comments on the manuscript.
Funding
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior–CAPES (SK: Finance Code 001), Santander Mobilidade 04/2016 (SK), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (RJ: CNPq 478982/2013-5 and 301616/2017-5).
Author information
Authors and Affiliations
Contributions
SK, AK, and RJ conceived this research and sampling design; SK collected the data and performed the analyses; AH and KB supervised data analyses; SK wrote the paper. All authors read, participated in revisions, and approved the final manuscript.
Corresponding author
Additional information
Handling Editor: Mathieu Lihoreau
Le stockage à long terme affecte la qualité de l'éjaculat de l'abeille monogame sans dard ( Scaptotrigona aff. depilis ).
reproduction / héritabilité / sélection sexuelle / Hymenoptera.
Die Langzeitaufbewahrung beeinfusst die Qualität des Ejakulats bei der monogamen Stachellosen Biene Scaptotrigona aff. Depilis.
Züchtung / Vererbbarkeit / sexuelle Selektion / Hymenoptera.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Koffler, S., Hoppe, A., Bienefeld, K. et al. Long-term storage shapes ejaculate traits in a monogamous stingless bee (Scaptotrigona aff. depilis). Apidologie 52, 242–251 (2021). https://doi.org/10.1007/s13592-020-00813-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-020-00813-x