A Mutant of Vibrio parahaemolyticus pirABVP (+) That Carries Binary Toxin Genes but Does Not Cause Acute Hepatopancreatic Necrosis Disease


Abstract
:1. Introduction
2. Materials and Methods
2.1. Wild Type and Natural Mutants of AHPND-Causing V. parahaemolyticus
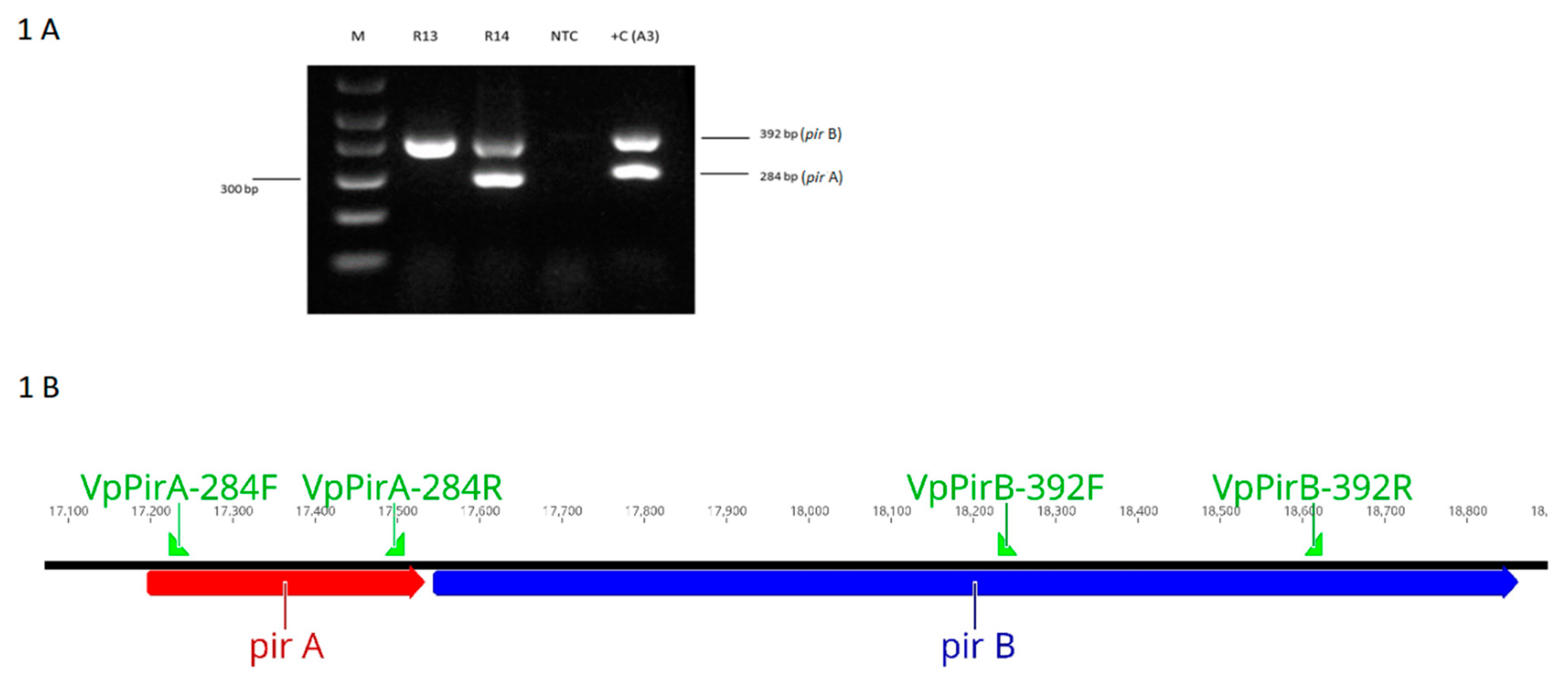
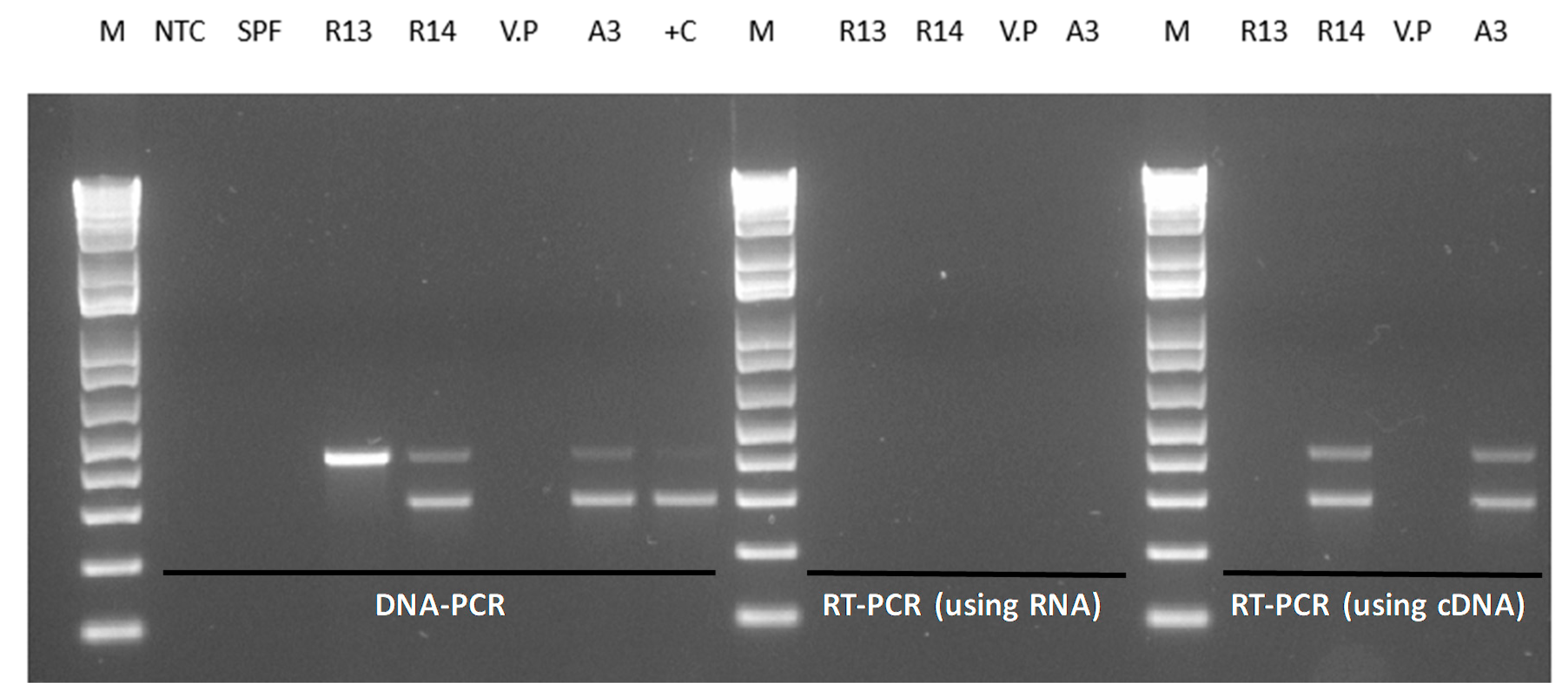
2.2. Duplex PCR Assay for the Detection of pirA- and pirB-Genes
2.3. Reverse Transcriptase-PCR for the Detection of the pirAB Genes
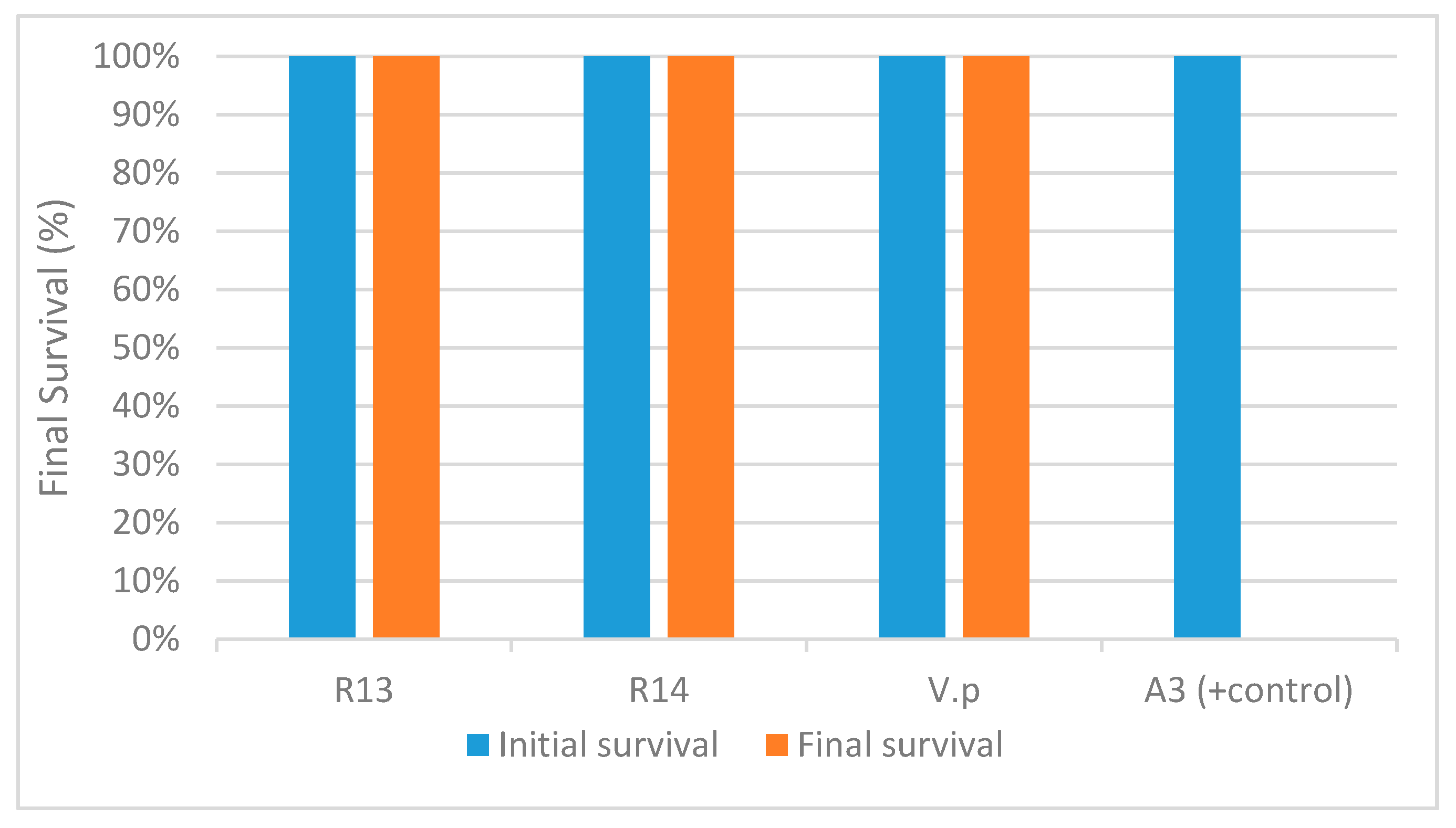
2.4. AHPND Challenge Test
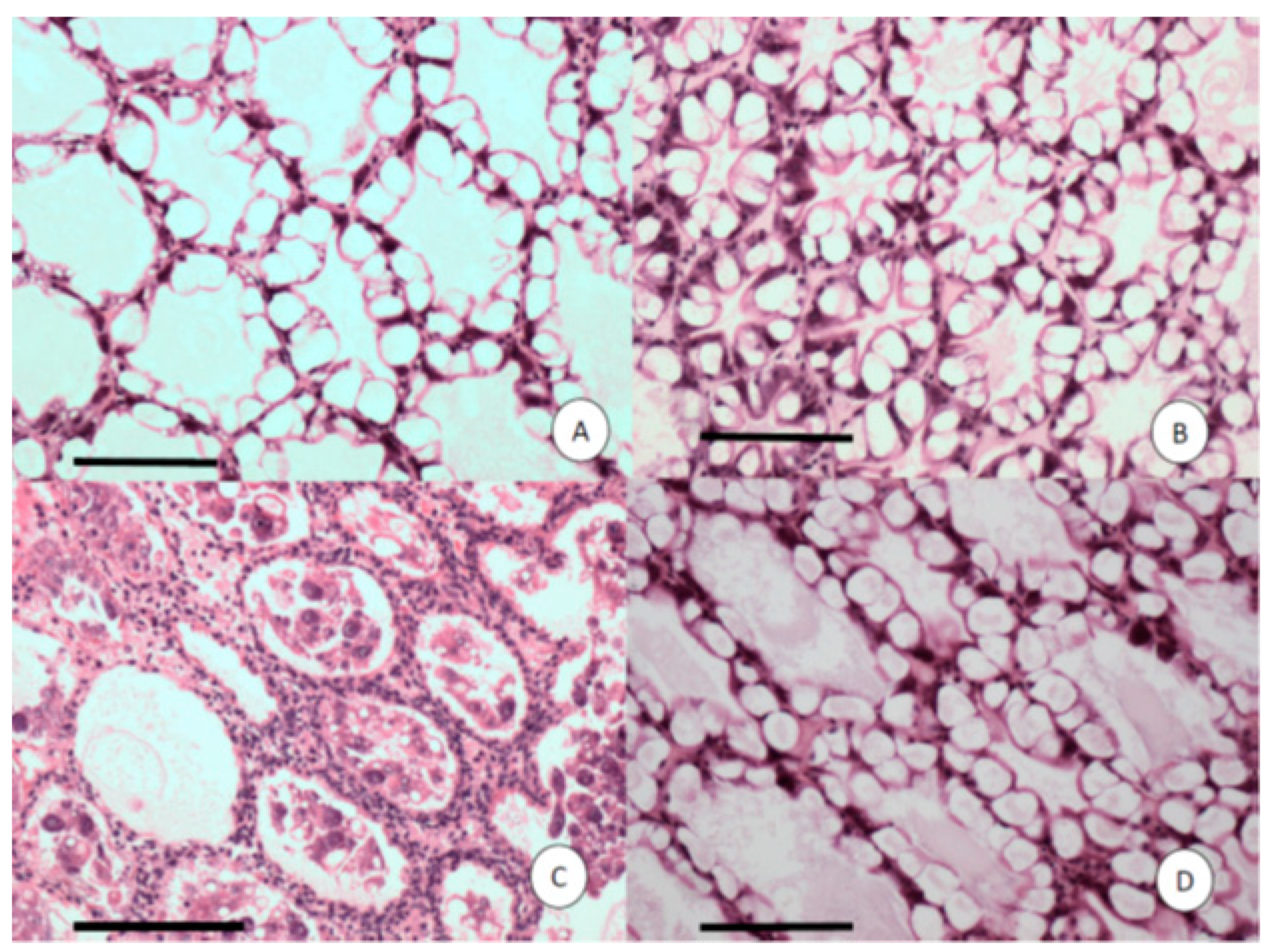
2.5. Histopathology
2.6. Sequence Analysis
2.7. Western Blot Analysis
3. Results
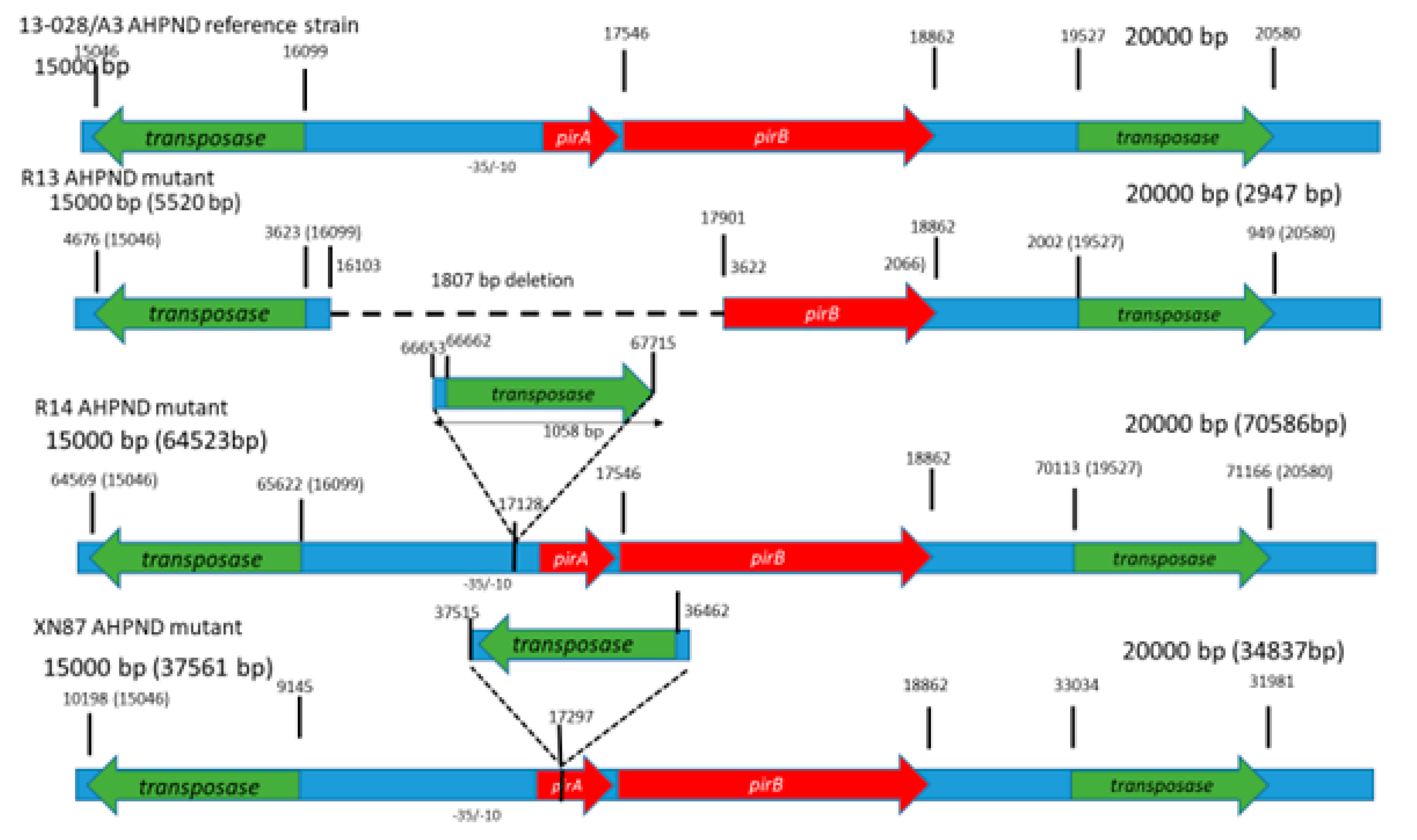
3.1. Duplex PCR for the Detection of the pirA and pirB-Like Genes
3.2. Challenge Test, Histopathology and PCR Results
3.3. Detection of pirA and pirB Transcripts by RT-PCR
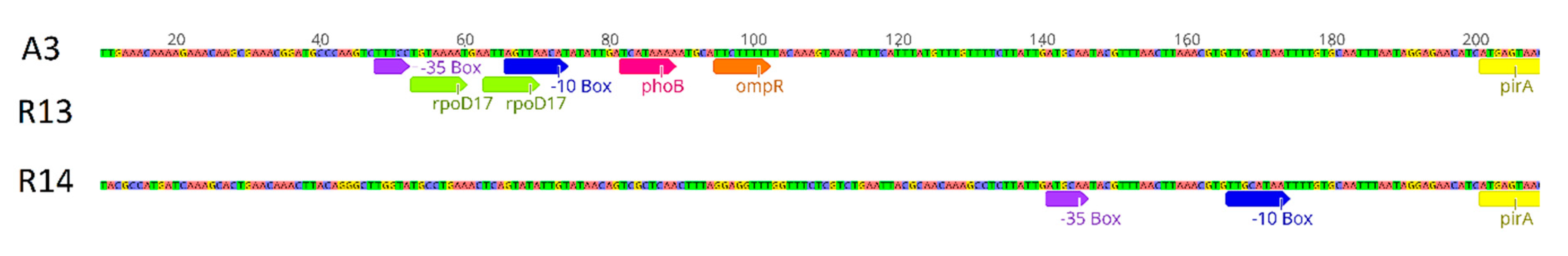
3.4. Sequence Analyses
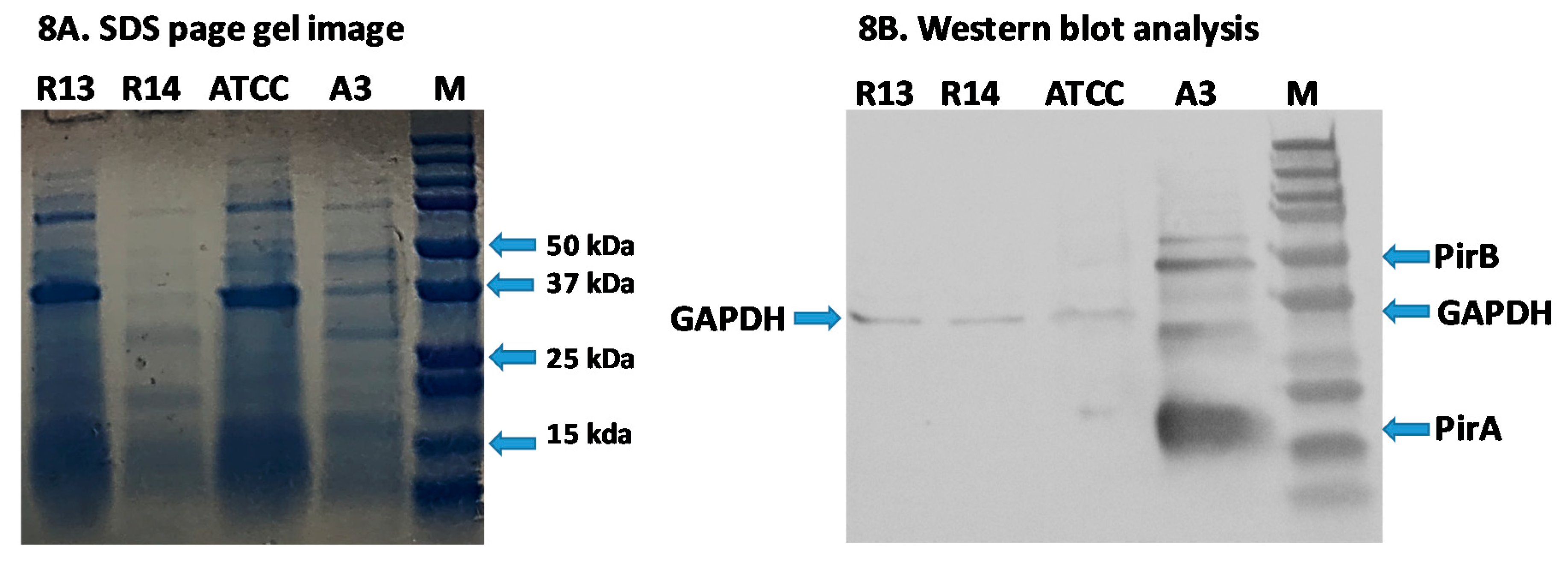
3.5. Detection of Binary Toxin by Western Blot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lightner, D.; Flegel, T. Diseases of Crustaceans–Acute Hepatopancreatic Necrosis Syndrome (AHPNS) (Disease Card). Asia Pacific Emergency Regional Consultation on Early Mortality Syndrome (EMS)/Acute Hepatopancreatic Necrosis Syndrome (AHPNS), 2014.
- Tran, L.; Nunan, L.; Redman, R.; Mohney, L.; Pantoja, C.; Fitzsimmons, K.; Lightner, D. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Organ. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, L.; Cabillon, N.; Catedral, D.; Amar, E.; Usero, R.; Monotilla, W.; Calpe, A.; Fernandez, D.; Saloma, C. Acute hepatopancreatic necrosis disease (AHPND) outbreaks in Penaeus vannamei and P. monodon cultured in the Philippines. Dis. Aquat. Organ. 2015, 116, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Nunan, L.; Lightner, D.; Pantoja, C.; Gomez-Jimenez, S. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Dis. Aquat. Organ. 2014, 111, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, A.K.; Piamsomboon, P.; Aranguren Caro, L.F.; Kanrar, S.; Adami, R.; Juan, Y.-S. First report of acute hepatopancreatic necrosis disease (AHPND) occurring in the USA. Dis. Aquat. Organ. 2019, 132, 241–247. [Google Scholar] [CrossRef]
- Woo, J.; Eun, J.; Tang, K.F.J.; Lightner, D.V.; Kim, J.; Won, S.; Chang, S. Potential application of bacteriophage pVp-1: Agent combating Vibrio parahaemolyticus strains associated with acute hepatopancreatic necrosis disease ( AHPND ) in shrimp. Aquaculture 2016, 457, 100–103. [Google Scholar]
- Restrepo, L.; Bayot, B.; Betancourt, I.; Pinzón, A. Draft genome sequence of pathogenic bacteria Vibrio parahaemolyticus strain Ba94C2, associated with acute hepatopancreatic necrosis disease isolate from South America. Genomics Data 2016, 9, 143–144. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, L.; Bayot, B.; Arciniegas, S.; Bajaña, L.; Betancourt, I.; Panchana, F.; Reyes Muñoz, A. PirVP genes causing AHPND identified in a new Vibrio species (Vibrio punensis) within the commensal Orientalis clade. Sci. Rep. 2018, 8, 13080. [Google Scholar] [CrossRef]
- Han, J.; Tang, K.; Lightner, D. Genotyping of virulence plasmid from Vibrio parahaemolyticus isolates causing acute hepatopancreatic necrosis disease in shrimp. Dis. Aquat. Organ. 2015, 115, 245–251. [Google Scholar] [CrossRef]
- Han, J.E.; Tang, K.F.J.; Aranguren, L.F.; Piamsomboon, P. Characterization and pathogenicity of acute hepatopancreatic necrosis disease natural mutants, pirABvp (-) V. parahaemolyticus, and pirABvp (+) V. campbellii strains. Aquaculture 2017, 470, 84–90. [Google Scholar] [CrossRef]
- Kondo, H.; Van, P.T.; Dang, L.T.; Hirono, I. Draft Genome Sequence of Non- Vibrio parahaemolyticus Acute Hepatopancreatic Necrosis Disease Strain KC13.17.5, Isolated from Diseased Shrimp in Vietnam. Genome Announc. 2015, 3, 2014–2015. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Liu, L.; Ke, Y.; Li, X.; Liu, Y.; Pan, Y.; Yan, S.; Wang, Y. Shrimp AHPND-causing plasmids encoding the PirAB toxins as mediated by pirAB-Tn903 are prevalent in various Vibrio species. Sci. Rep. 2017, 7, 42177. [Google Scholar] [CrossRef] [PubMed]
- Sirikharin, R.; Taengchaiyaphum, S.; Sanguanrut, P.; Chi, T.D.; Mavichak, R.; Proespraiwong, P.; Nuangsaeng, B.; Thitamadee, S.; Flegel, T.W.; Sritunyalucksana, K. Characterization and PCR Detection Of Binary, Pir-Like Toxins from Vibrio parahaemolyticus Isolates that Cause Acute Hepatopancreatic Necrosis Disease (AHPND) in Shrimp. PLoS ONE 2015, 10, e0126987. [Google Scholar] [CrossRef] [PubMed]
- Phiwsaiya, K.; Charoensapsri, W.; Taengphu, S.; Dong, H.T.; Sangsuriya, P.; Taengchaiyaphum, S.; Chaivisuthangkura, P.; Longyant, S.; Sithigorngul, P.; Senapin, S. A Natural Vibrio parahaemolyticus Produces neither Pir Vp Toxins nor Acute Hepatopancreatic Necrosis Disease. Appl. Environ. Microbiol. 2017, 83, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, L.; Caro, A.; Mai, H.N.; Noble, B.; Dhar, A.K. Acute hepatopancreatic necrosis disease (VP AHPND), a chronic disease in shrimp Penaeus vannamei population raised in latin America. J. Invertebr. Pathol. 2020, 174, 107424. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibuchi, M. Identification of Vibrio parahaemolyticus strains at the species level by PCR targeted to the toxR gene. J. Clin. Microbiol. 1999, 37, 1173–1177. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Tang, K.; Tran, L.; Lightner, D. Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp. Dis. Aquat. Organ. 2015, 113, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Bell, T.A.; Lightner, D.V. A Handbook of Normal Penaeid Shrimp Histology, 1st ed.; World Aquaculture Society: Baton Rouge, Louisiana, 1988. [Google Scholar]
- Mai, H.N.; Cruz-Flores, R.; Dhar, A.K. Development of an indirect Enzyme Linked Immunoassay (iELISA) using monoclonal antibodies against Photorhabdus insect related toxins, PirAVp and PirBVp released from Vibrio spp. J. Microbiol. Methods 2020, 176, 106002. [Google Scholar] [CrossRef]
- Kanrar, S.; Dhar, A.K. Complete Genome Sequence of a Novel Mutant Strain of Vibrio parahaemolyticus from Pacific White Shrimp (Penaeus vannamei). Genome Announc. 2018, 6, 19–20. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.T.; Ismail, A.M.; Camilli, A. PhoB regulates both environmental and virulence gene expression in Vibrio cholerae. Mol. Microbiol. 2010, 77, 1595–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.W.; Gao, Y.; Kim, D.; Szubin, R.; Yang, J.; Cho, B.-K.; Palsson, B.O. Revealing genome-scale transcriptional regulatory landscape of OmpR highlights its expanded regulatory roles under osmotic stress in Escherichia coli K-12 MG1655. Sci. Rep. 2017, 7, 2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, T.E.; Yu, J.; Belasco, J.G. mRNA stabilization by the ompA 5’ untranslated region: Two protective elements hinder distinct pathways for mRNA degradation. RNA 1998, 4, 319–330. [Google Scholar] [PubMed]
- Wagner, L.A.; Gesteland, R.F.; Dayhuff, T.J.; Weiss, R.B. An efficient Shine-Dalgarno sequence but not translation is necessary for lacZ mRNA stability in Escherichia coli. J. Bacteriol. 1994, 176, 1683–1688. [Google Scholar] [CrossRef] [Green Version]
- Karhumaa, K.; Rutberg, B.; Hambraeus, G. A 5′ stem–loop and ribosome binding but not translation are important for the stability of Bacillus subtilis aprE leader mRNA. Microbiology 2002, 148, 1795–1803. [Google Scholar]
- Epshtein, V.; Nudler, E. Cooperation Between RNA Polymerase Molecules in Transcription Elongation. Science (80-. ) 2003, 300, 801–805. [Google Scholar] [CrossRef]
- Deana, A. Lost in translation: The influence of ribosomes on bacterial mRNA decay. Genes Dev. 2005, 19, 2526–2533. [Google Scholar] [CrossRef] [Green Version]
- Iost, I.; Dreyfus, M. The stability of Escherichia coli lacZ mRNA depends upon the simultaneity of its synthesis and translation. EMBO J. 1995, 14, 3252–3261. [Google Scholar] [CrossRef]
- Abo, T.; Inada, T.; Ogawa, K.; Aiba, H. SsrA-mediated tagging and proteolysis of LacI and its role in the regulation of lac operon. EMBO J. 2000, 19, 3762–3769. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Kuwana, R.; Takamatsu, H. kinA mRNA is missing a stop codon in the undomesticated Bacillus subtilis strain ATCC 6051. Microbiology 2008, 154, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Roche, E.D.; Sauer, R.T. SsrA-mediated peptide tagging caused by rare codons and tRNA scarcity. EMBO J. 1999, 18, 4579–4589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yagi, M.; Morita, T.; Aiba, H. Cleavage of mRNAs and role of tmRNA system under amino acid starvation in Escherichia coli. Mol. Microbiol. 2008, 68, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Garza-Sánchez, F.; Gin, J.G.; Hayes, C.S. Amino Acid Starvation and Colicin D Treatment Induce A-site mRNA Cleavage in Escherichia coli. J. Mol. Biol. 2008, 378, 505–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunohara, T.; Jojima, K.; Tagami, H.; Inada, T.; Aiba, H. Ribosome Stalling during Translation Elongation Induces Cleavage of mRNA Being Translated in Escherichia coli. J. Biol. Chem. 2004, 279, 15368–15375. [Google Scholar] [CrossRef] [Green Version]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.A.; Ohneck, E.A.; Ryan, C.; Alonzo, F.; Smith, H.; Narechania, A.; Kolokotronis, S.-O.; Satola, S.W.; Uhlemann, A.-C.; Sebra, R.; et al. Evolution of hypervirulence by a MRSA clone through acquisition of a transposable element. Mol. Microbiol. 2014, 93, 664–681. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Features | R13 | R14 | V.P (ATCC) | A3 | XN87 * |
|---|---|---|---|---|---|
| Size (kb) | 71 | 75.713 | − | 69 | − |
| pirA | − | + | − | + | − |
| pirB | Truncated (nt 361–1317) | + | − | + | + |
| Deletion/Insertion | Deletion | Insertion | − | − | Insertion |
| Mortality (%) | 0% | 0% | 0% | 100% | 47% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aranguren Caro, L.F.; Mai, H.N.; Kanrar, S.; Cruz-Flores, R.; Dhar, A.K. A Mutant of Vibrio parahaemolyticus pirABVP (+) That Carries Binary Toxin Genes but Does Not Cause Acute Hepatopancreatic Necrosis Disease. Microorganisms 2020, 8, 1549. https://doi.org/10.3390/microorganisms8101549
Aranguren Caro LF, Mai HN, Kanrar S, Cruz-Flores R, Dhar AK. A Mutant of Vibrio parahaemolyticus pirABVP (+) That Carries Binary Toxin Genes but Does Not Cause Acute Hepatopancreatic Necrosis Disease. Microorganisms. 2020; 8(10):1549. https://doi.org/10.3390/microorganisms8101549
Chicago/Turabian StyleAranguren Caro, Luis Fernando, Hung N. Mai, Siddhartha Kanrar, Roberto Cruz-Flores, and Arun K. Dhar. 2020. "A Mutant of Vibrio parahaemolyticus pirABVP (+) That Carries Binary Toxin Genes but Does Not Cause Acute Hepatopancreatic Necrosis Disease" Microorganisms 8, no. 10: 1549. https://doi.org/10.3390/microorganisms8101549