X-Linked Duchenne-Type Muscular Dystrophy in Jack Russell Terrier Associated with a Partial Deletion of the Canine DMD Gene

 , ,
, ,  , , , and
, , , and 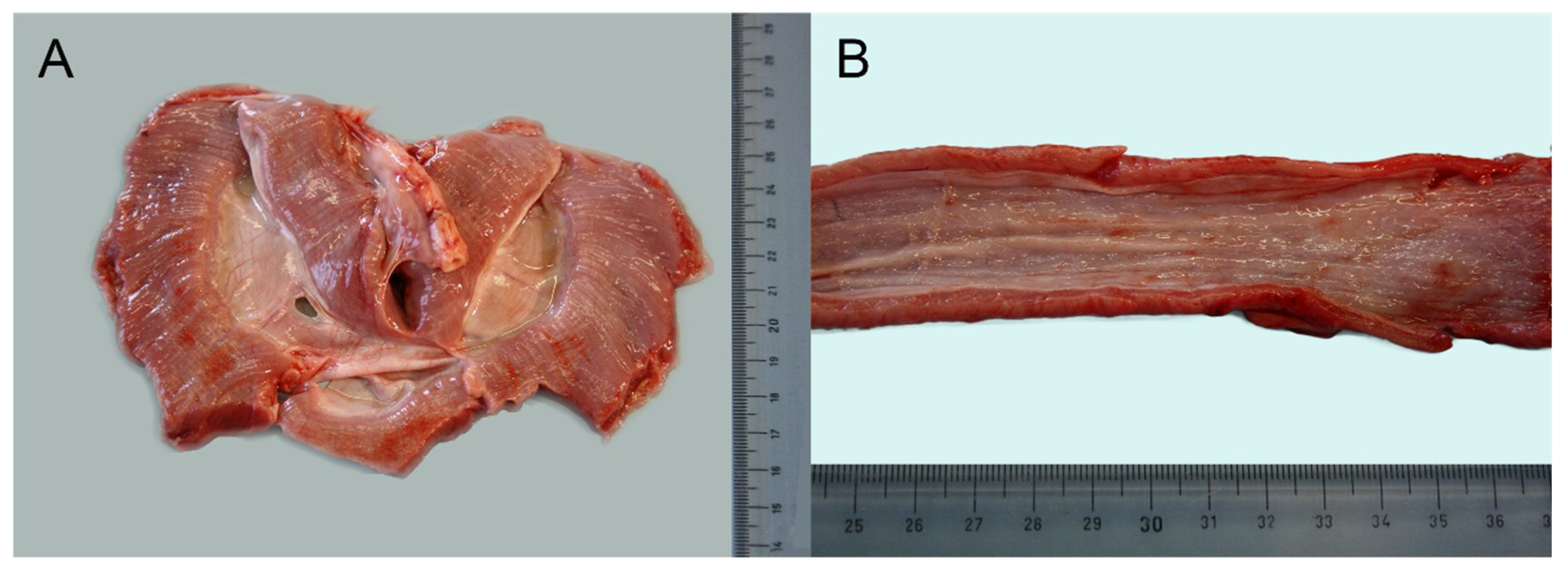{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Clinical History and Necropsy Request
2.3. Histopathology
2.4. Immunohistochemistry
2.5. Transmission Electron Microscopy
2.6. Whole-Genome Sequencing (WGS)
3. Results
3.1. Necropsy Examination
3.2. Histopathology
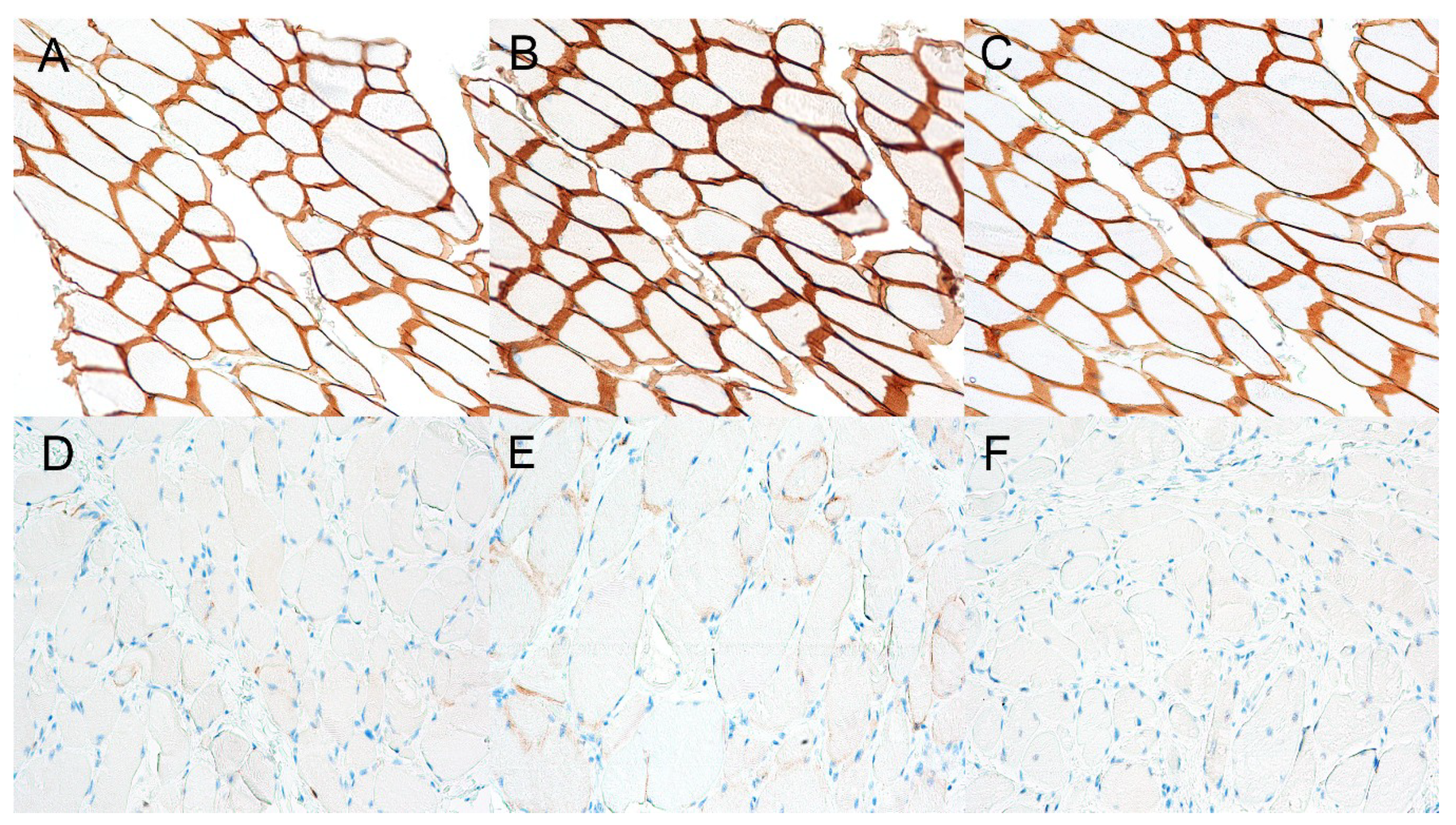
3.3. Immunohistochemistry
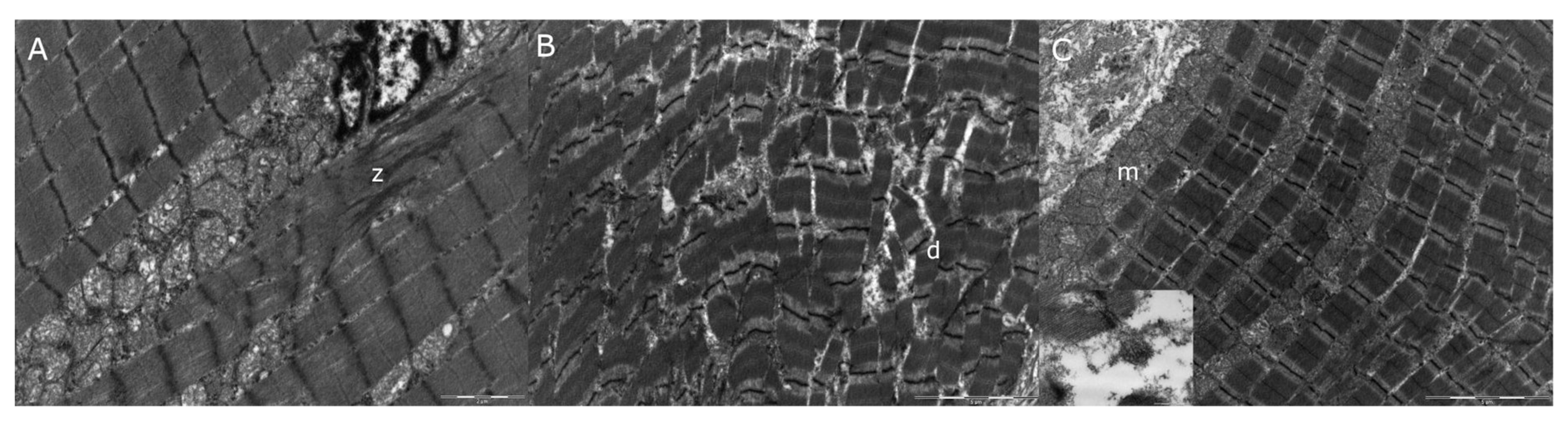
3.4. Transmission Electron Microscopy
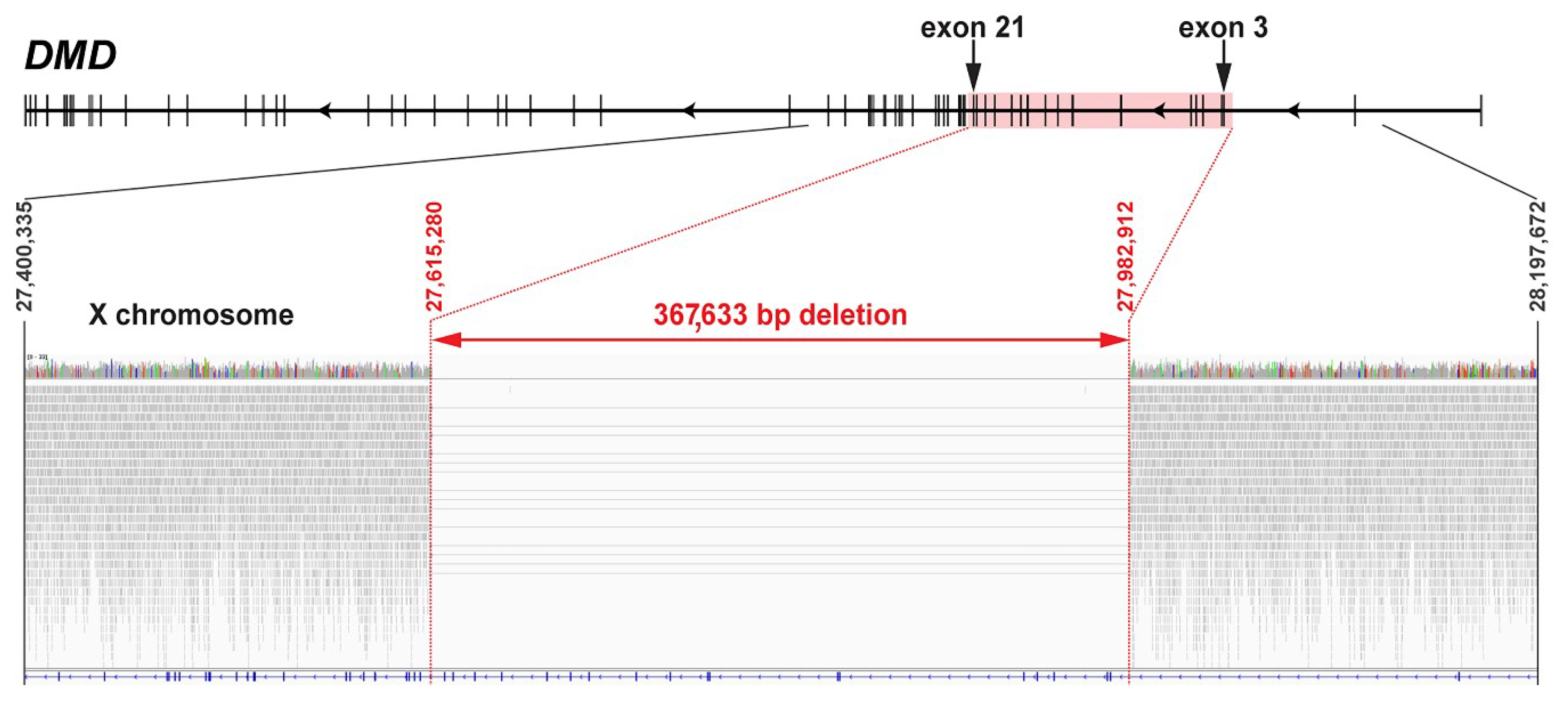
3.5. Whole-Genome Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, H.F.; Bryen, S.J.; Waddell, L.B.; Bournazos, A.; Davis, M.; Farrar, M.A.; Mclean, C.A.; Mowat, D.R.; Sampaio, H.; Woodcock, I.R.; et al. Importance of muscle biopsy to establish pathogenicity of DMD missense and splice variants. Neuromuscul. Disord. 2019, 29, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, L.; Beltrán, E.; De Stefani, A.; Guo, L.T.; Shea, A.; Shelton, G.D.; De Risio, L.; Burmeister, L.M. Clinical and genetic characterisation of dystrophin-deficient muscular dystrophy in a family of Miniature Poodle dogs. PLoS ONE 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freund, A.A.; Scola, R.H.; Arndt, R.C.; Lorenzoni, P.J.; Kay, C.K.; Werneck, L.C. Duchenne and Becker muscular dystrophy: A molecular and immunohistochemical approach. Arq. Neuropsiquiatr. 2007, 65, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Deisch, J.K. Muscle and Nerve Development in Health and Disease, 6th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 4. [Google Scholar]
- Mata López, S.; Hammond, J.J.; Rigsby, M.B.; Balog-Alvarez, C.J.; Kornegay, J.N.; Nghiem, P.P. A novel canine model for Duchenne muscular dystrophy (DMD): Single nucleotide deletion in DMD gene exon 20. Skelet. Muscle 2018, 8, 18–23. [Google Scholar] [CrossRef]
- Elhawary, N.A.; Jiffri, E.H.; Jambi, S.; Mufti, A.H.; Dannoun, A.; Kordi, H.; Khogeer, A.; Jiffri, O.H.; Elhawary, A.N.; Tayeb, M.T. Molecular characterization of exonic rearrangements and frame shifts in the dystrophin gene in Duchenne muscular dystrophy patients in a Saudi community. Hum. Genom. 2018, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchenne and Becker Muscular Dystrophy. Available online: https://ghr.nlm.nih.gov/condition/duchenne-and-becker-muscular-dystrophy#sourcesforpage (accessed on 7 October 2020).
- Shieh, P.B. Muscular Dystrophies and Other Genetic Myopathies. Neurol. Clin. 2013, 31, 1009–1029. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.J.; Valentine, B. Muscle and Tendon. In Jubb, Kennedy, and Palmer’s Pathology of Domestic Animals; Elsevier: St. Louis, MO, USA, 2016; ISBN 0702053228. [Google Scholar]
- Walmsley, G.L.; Arechavala-Gomeza, V.; Fernandez-Fuente, M.; Burke, M.M.; Nagel, N.; Holder, A.; Stanley, R.; Chandler, K.; Marks, S.L.; Muntoni, F.; et al. A duchenne muscular dystrophy gene hot spot mutation in dystrophin-deficient Cavalier King Charles Spaniels is amenable to exon 51 skipping. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, V.; Drögemüller, C.; Leeb, T.; Aguirre, G.; André, C.; Bannasch, D.; Becker, D.; Davis, B.; Ekenstedt, K.; Faller, K.; et al. A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Nucleotide Archive (ENA). Available online: www.ebi.ac.uk/ena (accessed on 7 October 2020).
- Nguyen, F.; Cherel, Y.; Guigand, L.; Goubault-Leroux, I.; Wyers, M. Muscle lesions associated with dystrophin deficiency in neonatal golden retriever puppies. J. Comp. Pathol. 2002, 126, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Schatzberg, S.J.; Anderson, L.V.B.; Wilton, S.D.; Kornegay, J.N.; Mann, C.J.; Solomon, G.G.; Sharp, N.J.H. Alternative dystrophin gene transcripts in golden retriever muscular dystrophy. Muscle Nerve 1998, 21, 991–998. [Google Scholar] [CrossRef]
- Vieira, N.M.; Elvers, I.; Alexander, M.S.; Moreira, Y.B.; Eran, A.; Gomes, J.P.; Marshall, J.L.; Karlsson, E.K.; Verjovski-Almeida, S.; Lindblad-Toh, K.; et al. Jagged 1 Rescues the Duchenne Muscular Dystrophy Phenotype. Cell 2015, 163, 1204–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nghiem, P.P.; Bello, L.; Balog, C.; Sara, A.; López, M.; Bettis, A.; Barnett, H.; Hernandez, B.; Schatzberg, S.J.; Piercy, R.J.; et al. Whole genome sequencing reveals a 7 base-pair deletion in DMD exon 42 in a dog with muscular dystrophy. Mamm. Genome 2017, 28, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, H.; Zamecnik, J. Diagnostic immunohistology of muscle diseases. J. Neuropathol. Exp. Neurol. 2005, 64, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGreevy, J.W.; Hakim, C.H.; McIntosh, M.A.; Duan, D. Animal models of Duchenne muscular dystrophy: From basic mechanisms to gene therapy. Dis. Model. Mech. 2015, 8, 195–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornegay, J.N.; Bogan, J.R.; Bogan, D.J.; Childers, M.K.; Li, J.; Nghiem, P.; Detwiler, D.A.; Larsen, C.A.; Grange, R.W.; Bhavaraju-Sanka, R.K.; et al. Canine models of Duchenne muscular dystrophy and their use in therapeutic strategies. Mamm. Genome 2012, 23, 85–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrader, S.M.; Jung, S.; Denney, T.S.; Smith, B.F. Characterization of Australian Labradoodle dystrophinopathy. Neuromuscul. Disord. 2018, 28, 927–937. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunetti, B.; Muscatello, L.V.; Letko, A.; Papa, V.; Cenacchi, G.; Grillini, M.; Murgiano, L.; Jagannathan, V.; Drögemüller, C. X-Linked Duchenne-Type Muscular Dystrophy in Jack Russell Terrier Associated with a Partial Deletion of the Canine DMD Gene. Genes 2020, 11, 1175. https://doi.org/10.3390/genes11101175
Brunetti B, Muscatello LV, Letko A, Papa V, Cenacchi G, Grillini M, Murgiano L, Jagannathan V, Drögemüller C. X-Linked Duchenne-Type Muscular Dystrophy in Jack Russell Terrier Associated with a Partial Deletion of the Canine DMD Gene. Genes. 2020; 11(10):1175. https://doi.org/10.3390/genes11101175
Chicago/Turabian StyleBrunetti, Barbara, Luisa V. Muscatello, Anna Letko, Valentina Papa, Giovanna Cenacchi, Marco Grillini, Leonardo Murgiano, Vidhya Jagannathan, and Cord Drögemüller. 2020. "X-Linked Duchenne-Type Muscular Dystrophy in Jack Russell Terrier Associated with a Partial Deletion of the Canine DMD Gene" Genes 11, no. 10: 1175. https://doi.org/10.3390/genes11101175