Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer


 and
and
Abstract
:1. Introduction
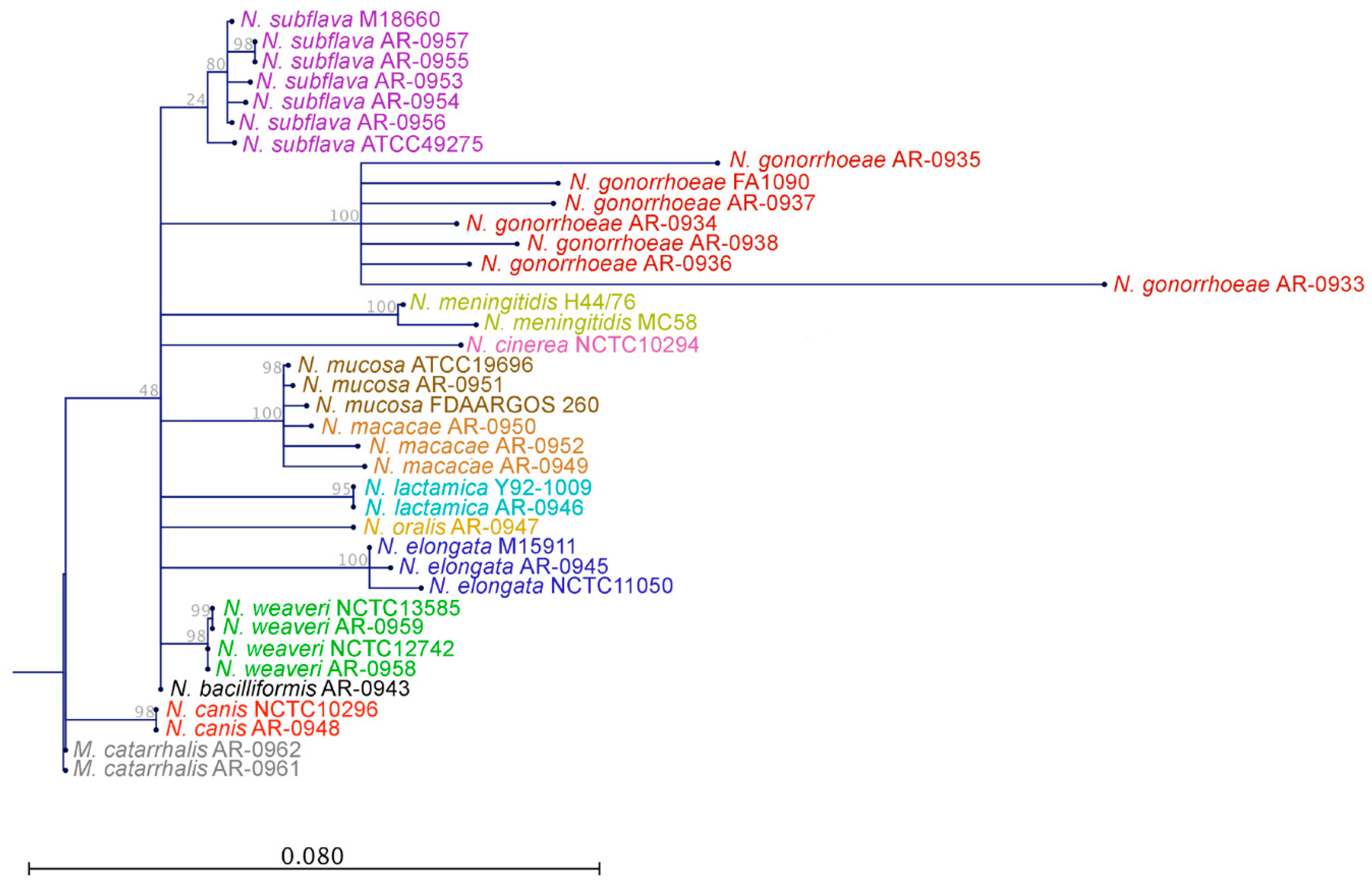
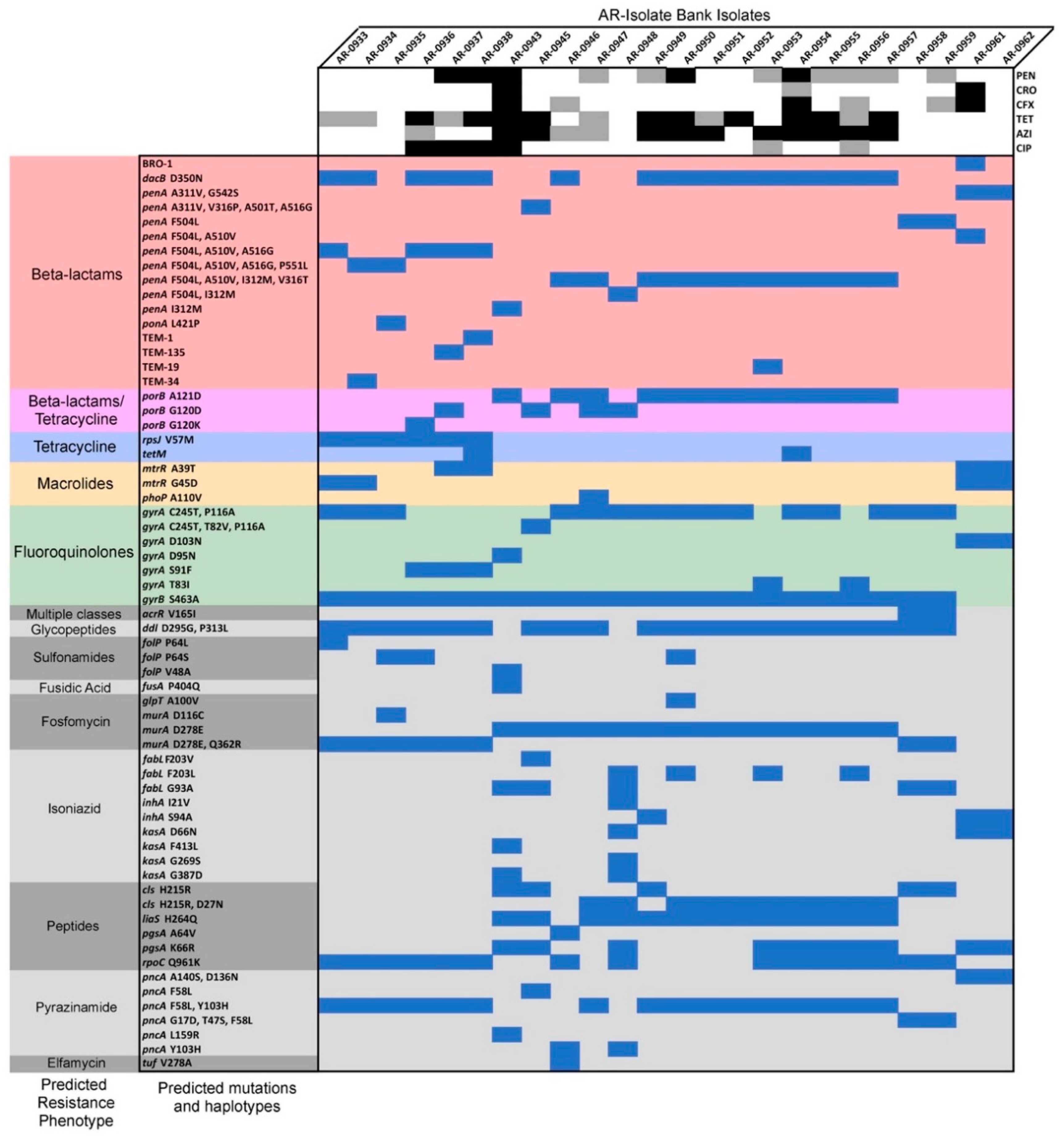
2. Results
2.1. Characterization of Phenotypic Resistance in the AR Isolate Bank Neisseria Panel
2.2. Genome Assemblies and Computational Analyses
2.3. Quinolone Reduced Susceptibility
2.4. Macrolide Resistance
2.5. Reduced Susceptibility to Tetracycline
2.6. Resistance to β-Lactams
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Minimum Inhibitory Concentration Testing
4.3. Library Preparation and Genomic Sequencing
4.4. Genome Assembly and Bioinformatic Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- U.S. Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019; pp. 1–148.
- Walsh, F.; Duffy, B. The culturable soil antibiotic resistome: A community of multi-drug resistant bacteria. PLoS ONE 2013, 8, e65567. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The shared antibiotic resistome of soil bacteria and human pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [Green Version]
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [Green Version]
- Bezanson, G.S.; Khakhria, R.; Bollegraaf, E. Nosocomial outbreak caused by antibiotic-resistant strain of Salmonella typhimurium acquired from dairy cattle. Can. Med. Assoc. J. 1983, 128, 426–427. [Google Scholar]
- Spika, J.S.; Waterman, S.H.; Hoo, G.W.; St Louis, M.E.; Pacer, R.E.; James, S.M.; Bissett, M.L.; Mayer, L.W.; Chiu, J.Y.; Hall, B. Chloramphenicol-resistant Salmonella newport traced through hamburger to dairy farms. N. Engl. J. Med. 1987, 316, 565–570. [Google Scholar] [CrossRef]
- Levy, S.B.; FitzGerald, G.B.; Macone, A.B. Changes in intestinal flora of farm personnel after introduction of a tetracycline-supplemented feed on a farm. N. Engl. J. Med. 1976, 295, 583–588. [Google Scholar] [CrossRef]
- Rashid, H.; Rahman, M. Possible transfer of plasmid mediated third generation cephalosporin resistance between Escherichia coli and Shigella sonnei in the human gut. Infect. Gent. Evol. 2015, 30, 15–18. [Google Scholar] [CrossRef]
- Knudsen, P.K.; Gammelsrud, K.W.; Alfsnes, K.; Steinbakk, M.; Abrahamsen, T.G.; Müller, F.M.X.; Bohlin, J. Transfer of a blaCTX-M-1-carrying plasmid between different Escherichia coli strains within the human gut explored by whole genome sequencing analyses. Sci. Rep. 2017, 8, 1–10. [Google Scholar]
- Wadsworth, C.B.; Arnold, B.J.; Sater, M.R.A.; Grad, Y.H. Azithromycin resistance through interspecific acquisition of an epistasis-dependent efflux pump component and transcriptional regulator in Neisseria gonorrhoeae. mBio 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2014, 13, 42–51. [Google Scholar] [CrossRef]
- Eyre, D.W.; Sanderson, N.D.; Lord, E.; Regisford-Reimmer, N.; Chau, K.; Barker, L.; Morgan, M.; Newnham, R.; Golparian, D.; Unemo, M.; et al. Gonorrhoea treatment failure caused by a Neisseria gonorrhoeae strain with combined ceftriaxone and high-level azithromycin resistance, England, February 2018. Euro Surveill. 2018, 23, 1800323. [Google Scholar] [CrossRef]
- Jennison, A.V.; Whiley, D.; Lahra, M.M.; Graham, R.M.; Cole, M.J.; Hughes, G.; Fifer, H.; Andersson, M.; Edwards, A.; Eyre, D. Genetic relatedness of ceftriaxone-resistant and high-level azithromycin resistant Neisseria gonorrhoeae cases, United Kingdom and Australia, February to April 2018. Euro Surveill. 2019, 24, 1900118. [Google Scholar] [CrossRef] [Green Version]
- Corander, J.; Connor, T.R.; O’Dwyer, C.A.; Kroll, J.S.; Hanage, W.P. Population structure in the Neisseria, and the biological significance of fuzzy species. J. R. Soc. Interface 2011, 9, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Halter, R.; Pohlner, J.; Meyer, T.F. Mosaic-like organization of igA protease genes in Neisseria gonorrhoeae generated by horizontal genetic exchange in vivo. EMBO J. 1989, 8, 2737–2744. [Google Scholar] [CrossRef] [Green Version]
- Feavers, I.M.; Heath, A.B.; Bygraves, J.A.; Maiden, M.C. Role of horizontal genetic exchange in the antigenic variation of the class 1 outer membrane protein of Neisseria meningitidis. Mol. Microbiol. 1992, 6, 489–495. [Google Scholar] [CrossRef]
- Smith, J.M.; Smith, N.H.; O’Rourke, M.; Spratt, B.G. How clonal are bacteria? Proc. Natl. Acad. Sci. USA 1993, 90, 4384–4388. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Spratt, B.G. Sequence diversity within the argF, fbp and recA genes of natural isolates of Neisseria meningitidis: Interspecies recombination within the argF gene. Mol. Microbiol. 1992, 6, 2135–2146. [Google Scholar] [CrossRef]
- Arnold, B.; Sohail, M.; Wadsworth, C.; Corander, J.; Hanage, W.P.; Sunyaev, S.; Grad, Y.H. Fine-Scale haplotype structure reveals strong signatures of positive selection in a recombining bacterial pathogen. Mol. Biol. Evol. 2020, 37, 417–428. [Google Scholar] [CrossRef]
- Rouquette-Loughlin, C.E.; Reimche, J.L.; Balthazar, J.T.; Dhulipala, V.; Gernert, K.M.; Kersh, E.N.; Pham, C.D.; Pettus, K.; Abrams, A.J.; Trees, D.L.; et al. Mechanistic basis for decreased antimicrobial susceptibility in a clinical isolate of Neisseria gonorrhoeae poossessing a mosaic-Like mtr efflux pump locus. mBio 2018, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Spratt, B.G.; Bowler, L.D.; Zhang, Q.Y.; Zhou, J.; Smith, J.M. Role of interspecies transfer of chromosomal genes in the evolution of penicillin resistance in pathogenic and commensal Neisseria species. J. Mol. Evol. 1992, 34, 115–125. [Google Scholar] [CrossRef]
- Grad, Y.H.; Harris, S.R.; Kirkcaldy, R.D.; Green, A.G.; Marks, D.S.; Bentley, S.D.; Trees, D.; Lipsitch, M. Genomic epidemiology of gonococcal resistance to extended-spectrum cephalosporins, macrolides, and fluoroquinolones in the United States, 2000–2013. J. Infect. Dis. 2016, 214, 1579–1587. [Google Scholar] [CrossRef] [Green Version]
- CDC & FDA. Antibiotic Resistance Isolate Bank; CDC: Atlanta, GA, USA, 2020.
- Lindbäck, E.; Rahman, M.; Jalal, S.; Wretlind, B. Mutations in gyrA, gyrB, parC, and parE in quinolone-resistant strains of Neisseria gonorrhoeae. APMIS 2002, 110, 651–657. [Google Scholar] [CrossRef]
- Peterson, S.W.; Martin, I.; Demczuk, W.; Bharat, A.; Hoang, L.; Wylie, J.; Allen, V.; Lefebvre, B.; Tyrrell, G.; Horsman, G.; et al. Molecular assay for detection of ciprofloxacin resistance in Neisseria gonorrhoeae isolates from cultures and clinical nucleic acid amplification test specimens. J. Clin. Microbiol. 2015, 53, 3606–3608. [Google Scholar] [CrossRef] [Green Version]
- Hemarajata, P.; Yang, S.; Soge, O.O.; Humphries, R.M.; Klausner, J.D. Performance and verification of a real-time pcr assay targeting the gyrA gene for prediction of ciprofloxacin resistance in Neisseria gonorrhoeae. J. Clin. Microbiol. 2016, 54, 805–808. [Google Scholar] [CrossRef] [Green Version]
- Ebeyan, S.; Windsor, M.; Bordin, A.; Mhango, L.; Erskine, S.; Mokany, E.; Tan, L.Y.; Whiley, D.; GRAND2 Study Investigators; Guy, R.; et al. Evaluation of the ResistancePlus GC (beta) assay: A commercial diagnostic test for the direct detection of ciprofloxacin susceptibility or resistance in Neisseria gonorrhoeae. J. Antimicrob. Chemoth. 2019, 74, 1820–1824. [Google Scholar] [CrossRef]
- Bruchmann, S.; Dötsch, A.; Nouri, B.; Chaberny, I.F.; Häussler, S. Quantitative contributions of target alteration and decreased drug accumulation to Pseudomonas aeruginosa fluoroquinolone resistance. Antimicrob. Agents Chemother. 2013, 57, 1361–1368. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.K.; Martin, I.; Liu, G.; Bryden, L. Mutation in 23S rRNA associated with macrolide resistance in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2002, 46, 3020–3025. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.A.; Dave, J.; Ison, C.A. High-Level azithromycin resistance occurs in Neisseria gonorrhoeae as a result of a single point mutation in the 23S rRNA genes. Antimicrob. Agents Chemother. 2010, 54, 3812–3816. [Google Scholar] [CrossRef] [Green Version]
- Trembizki, E.; Doyle, C.; Jennison, A.; Smith, H.; Bates, J.; Lahra, M.; Whiley, D. A Neisseria gonorrhoeae strain with a meningococcal mtrR sequence. J. Med. Microbiol. 2014, 63, 1113–1115. [Google Scholar] [CrossRef] [Green Version]
- Zarantonelli, L.; Borthagaray, G.; Lee, E.H.; Veal, W.; Shafer, W.M. Decreased susceptibility to azithromycin and erythromycin mediated by a novel mtr(R) promoter mutation in Neisseria gonorrhoeae. J. Antimicrob. Chemoth. 2001, 47, 651–654. [Google Scholar] [CrossRef] [Green Version]
- Ohneck, E.A.; Zalucki, Y.M.; Johnson, P.J.T.; Dhulipala, V.; Golparian, D.; Unemo, M.; Jerse, A.E.; Shafer, W.M. A novel mechanism of high-level, broad-spectrum antibiotic resistance caused by a single base pair change in Neisseria gonorrhoeae. mBio 2011, 2, e00187-11. [Google Scholar] [CrossRef] [Green Version]
- Hagman, K.E.; Pan, W.; Spratt, B.G.; Balthazar, J.T.; Judd, R.C.; Shafer, W.M. Resistance of Neisseria gonorrhoeae to antimicrobial hydrophobic agents is modulated by the mtrRCDE efflux system. Microbiology 1995, 141, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Demczuk, W.; Martin, I.; Peterson, S.; Bharat, A.; Van Domselaar, G.; Graham, M.; Lefebvre, B.; Allen, V.; Hoang, L.; Tyrrell, G.; et al. Genomic epidemiology and molecular resistance mechanisms of azithromycin-resistant Neisseria gonorrhoeae in Canada from 1997 to 2014. J. Clin. Microbiol. 2016, 54, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- Shafer, W.M.; Balthazar, J.T.; Hagman, K.E.; Morse, S.A. Missense mutations that alter the DNA-binding domain of the MtrR protein occur frequently in rectal isolates of Neisseria gonorrhoeae that are resistant to faecal lipids. Microbiology 1995, 141 Pt 4, 907–911. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.S.; Johnson, S.R.; Zenilman, J.M.; Roberts, M.C.; Morse, S.A. High-level tetracycline resistance resulting from TetM in strains of Neisseria spp., Kingella denitrificans, and Eikenella corrodens. Antimicrob. Agents Chemother. 1988, 32, 765–767. [Google Scholar] [CrossRef] [Green Version]
- Morse, S.A.; Johnson, S.R.; Biddle, J.W.; Roberts, M.C. High-level tetracycline resistance in Neisseria gonorrhoeae is result of acquisition of streptococcal tetM determinant. Antimicrob. Agents Chemother. 1986, 30, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Gascoyne, D.M.; Heritage, J.; Hawkey, P.M.; Turner, A.; van Klingeren, B. Molecular evolution of tetracycline-resistance plasmids carrying TetM found in Neisseria gonorrhoeae from different countries. J. Antimicrob. Chemoth. 1991, 28, 173–183. [Google Scholar] [CrossRef]
- Gill, M.J.; Simjee, S.; Al-Hattawi, K.; Robertson, B.D.; Easmon, C.S.F.; Ison, C.A. Gonococcal resistance to β-lactams and tetracycline involves mutation in loop 3 of the porin encoded at the penB locus. Antimicrob. Agents Chemother. 1998, 42, 2799–2803. [Google Scholar] [CrossRef] [Green Version]
- Sparling, P.F.; Sarubbi, F.A.; Blackman, E. Inheritance of low-level resistance to penicillin, tetracycline, and chloramphenicol in Neisseria gonorrhoeae. J. Bacteriol. 1975, 124, 740–749. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Spratt, B.G. Regulation of the permeability of the gonococcal cell envelope by the mtr system. Mol. Microbiol. 1994, 11, 769–775. [Google Scholar] [CrossRef]
- Veal, W.L.; Nicholas, R.A.; Shafer, W.M. Overexpression of the MtrC-MtrD-MtrE efflux pump due to an mtrR mutation is required for chromosomally mediated penicillin resistance in Neisseria gonorrhoeae. J. Bacteriol. 2002, 184, 5619–5624. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Nandi, S.; Davies, C.; Nicholas, R.A. High-Level chromosomally mediated tetracycline resistance in Neisseria gonorrhoeae results from a point mutation in the rpsJ gene encoding ribosomal protein S10 in combination with the mtrR and penB resistance determinants. Antimicrob. Agents Chemother. 2005, 49, 4327–4334. [Google Scholar] [CrossRef] [Green Version]
- Ashford, W.; Golash, R.; Hemming, V. Penicilunase-producing Neisseria gonorrhoeae. Lancet 2020, 308, 657–658. [Google Scholar] [CrossRef]
- Dowson, C.G.; Jephcott, A.E.; Gough, K.R.; Spratt, B.G. Penicillin-binding protein 2 genes of non-beta-lactamase-producing, penicillin-resistant strains of Neisseria gonorrhoeae. Mol. Microbiol. 1989, 3, 35–41. [Google Scholar] [CrossRef]
- Ropp, P.A.; Hu, M.; Olesky, M.; Nicholas, R.A. Mutations in ponA, the gene encoding Penicillin-Binding Protein 1, and a novel locus, penC, are required for high-level chromosomally mediated penicillin resistance in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2002, 46, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Tobiason, D.M.; Hu, M.; Seifert, H.S.; Nicholas, R.A. The penC mutation conferring antibiotic resistance in Neisseria gonorrhoeae arises from a mutation in the PilQ secretin that interferes with multimer stability. Mol. Microbiol. 2005, 57, 1238–1251. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, M.; Golparian, D.; Shimuta, K.; Saika, T.; Hoshina, S.; Iwasaku, K.; Nakayama, S.I.; Kitawaki, J.; Unemo, M. Is Neisseria gonorrhoeae initiating a future era of untreatable gonorrhea?: Detailed characterization of the first strain with high-level resistance to ceftriaxone. Antimicrob. Agents Chemother. 2011, 55, 3538–3545. [Google Scholar] [CrossRef] [Green Version]
- Ameyama, S.; Onodera, S.; Takahata, M.; Minami, S.; Maki, N.; Endo, K.; Goto, H.; Suzuki, H.; Oishi, Y. Mosaic-Like structure of Penicillin-Binding Protein 2 gene (penA) in clinical isolates of Neisseria gonorrhoeae with reduced susceptibility to cefixime. Antimicrob. Agents Chemother. 2002, 46, 3744–3749. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Deguchi, T.; Mizutani, K.-S.; Yasuda, M.; Yokoi, S.; Ito, S.-I.; Takahashi, Y.; Ishihara, S.; Kawamura, Y.; Ezaki, T. Emergence and spread of Neisseria gonorrhoeae clinical isolates harboring mosaic-like structure of Penicillin-Binding Protein 2 in Central Japan. Antimicrob. Agents Chemother. 2005, 49, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Fung, C.P.; Yeo, S.F.; Livermore, D.M. Susceptibility of Moraxella catarrhalis isolates to beta-lactam antibiotics in relation to beta-lactamase pattern. J. Antimicrob. Chemoth. 1994, 33, 215–222. [Google Scholar] [CrossRef]
- Khan, M.A.; Northwood, J.B.; Levy, F.; Verhaegh, S.J.C.; Farrell, D.J.; Van Belkum, A.; Hays, J.P. BRO β-lactamase and antibiotic resistances in a global cross-sectional study of Moraxella catarrhalis from children and adults. J. Antimicrob. Chemoth. 2009, 65, 91–97. [Google Scholar] [CrossRef]
- Kellogg, D.S.; Peacock, W.L.; Deacon, W.E.; Brown, l.; Pirkle, D. Neisseria gonorrhoeae. I. Virulence genetically linked to clonal variation. J. Bacteriol. 1963, 85, 1274–1279. [Google Scholar] [CrossRef] [Green Version]
- Papp, J.R.; Rowlinson, M.-C.; O’Connor, N.P.; Wholehan, J.; Razeq, J.H.; Glennen, A.; Ware, D.; Iwen, P.C.; Lee, L.V.; Hagan, C. Accuracy and reproducibility of the Etest to detect drug-resistant Neisseria gonorrhoeae to contemporary treatment. J. Med. Microbiol. 2018, 67, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 28 September 2020).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2010, 48, D517–D525. [Google Scholar] [CrossRef]
- Roberts, M.C. Plasmids of Neisseria gonorrhoeae and other Neisseria species. Clin. Microbiol. Rev. 1989, 2, S18–S23. [Google Scholar] [CrossRef]
- CLC Genomics Workbench 20.0. Available online: http://resources.qiagenbioinformatics.com/manuals/clcgenomicsworkbench/current/User_Manual.pdf (accessed on 28 September 2020).



{kind=link}
{kind=link}
| Minimum Inhibitory Concentration (MIC) a,b | |||||||
|---|---|---|---|---|---|---|---|
| AR Bank # | Species | Penicillin (PEN) | Ceftriaxone (CRO) | Cefixime (CFX) | Tetracycline (TET) | Azithromycin (AZI) | Ciprofloxacin (CIP) |
| AR-0933 | Neisseria gonorrhoeae | 0.19 | 0.004 | <0.016 | 1.5 | 0.25 | 0.004 |
| AR-0934 | Neisseria gonorrhoeae | 0.38 | 0.016 | 0.023 | 1.5 | 0.125 | 0.006 |
| AR-0935 | Neisseria gonorrhoeae | 0.19 | 0.006 | <0.016 | 0.75 | 0.064 | 0.006 |
| AR-0936 | Neisseria gonorrhoeae | 0.38 | 0.047 | <0.016 | 2 | 1.5 | >32 |
| AR-0937 | Neisseria gonorrhoeae | >32 | 0.016 | <0.016 | 1.5 | 0.38 | 6 |
| AR-0938 | Neisseria gonorrhoeae | >32 | 0.008 | <0.016 | 32 | 0.094 | 4 |
| AR-0943 | Neisseria bacilliformis | 32 | 16 | 0.5 | 4 | 4 | 6 |
| AR-0944 | Neisseria cinerea | 0.38 | 0.094 | <0.016 | 2 | 8 | 0.032 |
| AR-0945 | Neisseria elongata | 0.25 | 0.094 | 0.032 | 0.38 | 0.5 | 0.25 |
| AR-0946 | Neisseria lactamica | 0.75 | 0.023 | 0.25 | 0.75 | 1.5 | 0.008 |
| AR-0947 | Neisseria oralis | 1.5 | 0.047 | 0.064 | 1.5 | 1.5 | 0.016 |
| AR-0948 | Neisseria canis | 0.25 | 0.008 | <0.016 | 0.5 | 0.38 | 0.008 |
| AR-0949 | Neisseria macacae | 1.5 | 0.064 | 0.064 | 2 | 8 | 0.125 |
| AR-0950 | Neisseria macacae | 3 | 0.047 | 0.125 | 3 | 8 | 0.032 |
| AR-0951 | Neisseria mucosa | 0.25 | 0.032 | 0.125 | 1 | 3 | 0.023 |
| AR-0952 | Neisseria macacae | 0.38 | 0.023 | 0.047 | 2 | 0.5 | 0.012 |
| AR-0953 | Neisseria subflava | 1.5 | 0.019 | 0.38 | 0.5 | 2 | 0.75 |
| AR-0954 | Neisseria subflava | 3 | 0.25 | 0.5 | 48 | 4 | 0.047 |
| AR-0955 | Neisseria subflava | 1 | 0.032 | 0.064 | 6 | 12 | 0.125 |
| AR-0956 | Neisseria subflava | 1.5 | 0.125 | 0.25 | 1 | 6 | 0.75 |
| AR-0957 | Neisseria subflava | 1 | 0.047 | 0.064 | 4 | 8 | 0.064 |
| AR-0958 | Neisseria weaveri | 0.38 | 0.064 | 0.047 | 0.5 | 0.25 | 0.006 |
| AR-0959 | Neisseria weaveri | 1 | 0.19 | 0.023 | 0.75 | 0.75 | 0.023 |
| AR-0960 | Kingella denitrificans | 0.32 | 0.064 | 0.25 | 0.75 | 2 | 0.094 |
| AR-0961 | Moraxella catarrhalis | 0.19 | 1 | 2 | 0.5 | 0.25 | 0.094 |
| AR-0962 | Moraxella catarrhalis | 0.094 | 0.012 | 0.064 | 0.75 | 0.25 | 0.125 |
| Isolate | SRA Read Accession | Genome Size (bp) | GC Content (%) | No. of Contigs | Estimated Coverage (x) | No. of ORFs | No. of tRNAs |
|---|---|---|---|---|---|---|---|
| AR-0933 | SAMN15454039 | 2,100,787 | 52.6 | 65 | 438 | 2059 | 52 |
| AR-0934 | SAMN15454040 | 2,102,184 | 52.7 | 64 | 458 | 2081 | 49 |
| AR-0935 | SAMN15454041 | 2,215,614 | 52.35 | 54 | 470 | 2224 | 51 |
| AR-0936 | SAMN15454042 | 2,158,950 | 52.38 | 71 | 380 | 2117 | 50 |
| AR-0937 | SAMN15454043 | 2,138,399 | 52.53 | 69 | 498 | 2102 | 49 |
| AR-0938 | SAMN15454044 | 2,152,496 | 52.5 | 71 | 491 | 2124 | 49 |
| AR-0943 | SAMN15454045 | 2,342,297 | 59.34 | 91 | 421 | 2151 | 52 |
| AR-0945 | SAMN15454046 | 2,572,594 | 53.81 | 45 | 214 | 2511 | 55 |
| AR-0946 | SAMN15454047 | 2,145,323 | 52.48 | 53 | 366 | 1994 | 54 |
| AR-0947 | SAMN15454048 | 2,497,075 | 52.8 | 19 | 247 | 2302 | 56 |
| AR-0948 | SAMN15454049 | 2,399,311 | 48.41 | 42 | 341 | 2261 | 52 |
| AR-0949 | SAMN15454050 | 2,938,382 | 50.84 | 98 | 228 | 2749 | 58 |
| AR-0950 | SAMN15454051 | 1,762,877 | 51.47 | 18 | 344 | 2184 | 55 |
| AR-0951 | SAMN15454052 | 2,580,111 | 51.1 | 98 | 295 | 2330 | 54 |
| AR-0952 | SAMN15454053 | 2,446,961 | 51.04 | 66 | 319 | 2216 | 53 |
| AR-0953 | SAMN15454054 | 2,201,301 | 49.57 | 16 | 921 | 2096 | 54 |
| AR-0954 | SAMN15454055 | 2,359,025 | 49.13 | 57 | 390 | 2276 | 54 |
| AR-0955 | SAMN15454056 | 2,188,966 | 49.37 | 16 | 327 | 2033 | 55 |
| AR-0956 | SAMN15454057 | 2,238,415 | 49.02 | 34 | 397 | 2106 | 55 |
| AR-0957 | SAMN15454058 | 2,187,919 | 49.37 | 15 | 326 | 2029 | 55 |
| AR-0958 | SAMN15454059 | 2,268,810 | 49.1 | 30 | 478 | 2132 | 49 |
| AR-0959 | SAMN15454060 | 2,148,848 | 49.01 | 24 | 382 | 1962 | 49 |
| AR-0961 | SAMN15454061 | 1,882,811 | 41.81 | 13 | 577 | 1728 | 44 |
| AR-0962 | SAMN15454062 | 1,843,431 | 41.64 | 12 | 432 | 1684 | 43 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiore, M.A.; Raisman, J.C.; Wong, N.H.; Hudson, A.O.; Wadsworth, C.B. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics 2020, 9, 656. https://doi.org/10.3390/antibiotics9100656
Fiore MA, Raisman JC, Wong NH, Hudson AO, Wadsworth CB. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics. 2020; 9(10):656. https://doi.org/10.3390/antibiotics9100656
Chicago/Turabian StyleFiore, Michael A., Jordan C. Raisman, Narayan H. Wong, André O. Hudson, and Crista B. Wadsworth. 2020. "Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer" Antibiotics 9, no. 10: 656. https://doi.org/10.3390/antibiotics9100656