Antimicrobial Resistance in Escherichia coli and Resistance Genes in Coliphages from a Small Animal Clinic and in a Patient Dog with Chronic Urinary Tract Infection


Abstract
:1. Introduction
2. Results
2.1. Isolation of Coliphage and E. coli from the Clinical Environments
2.2. Antimicrobial Resistance in Isolated E. coli
2.3. Phages and Antimicrobial Resistance Gene Detection
2.4. Resistance Transduction
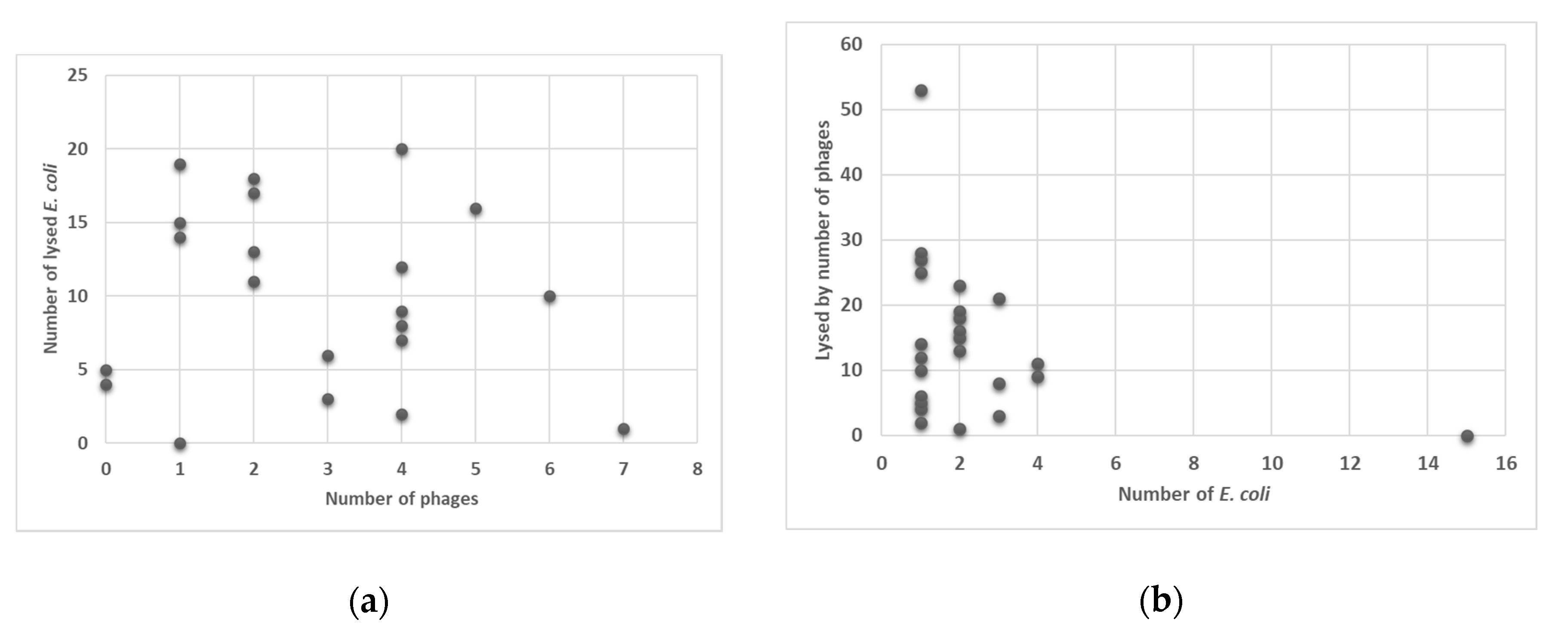
2.5. Phage Lysis
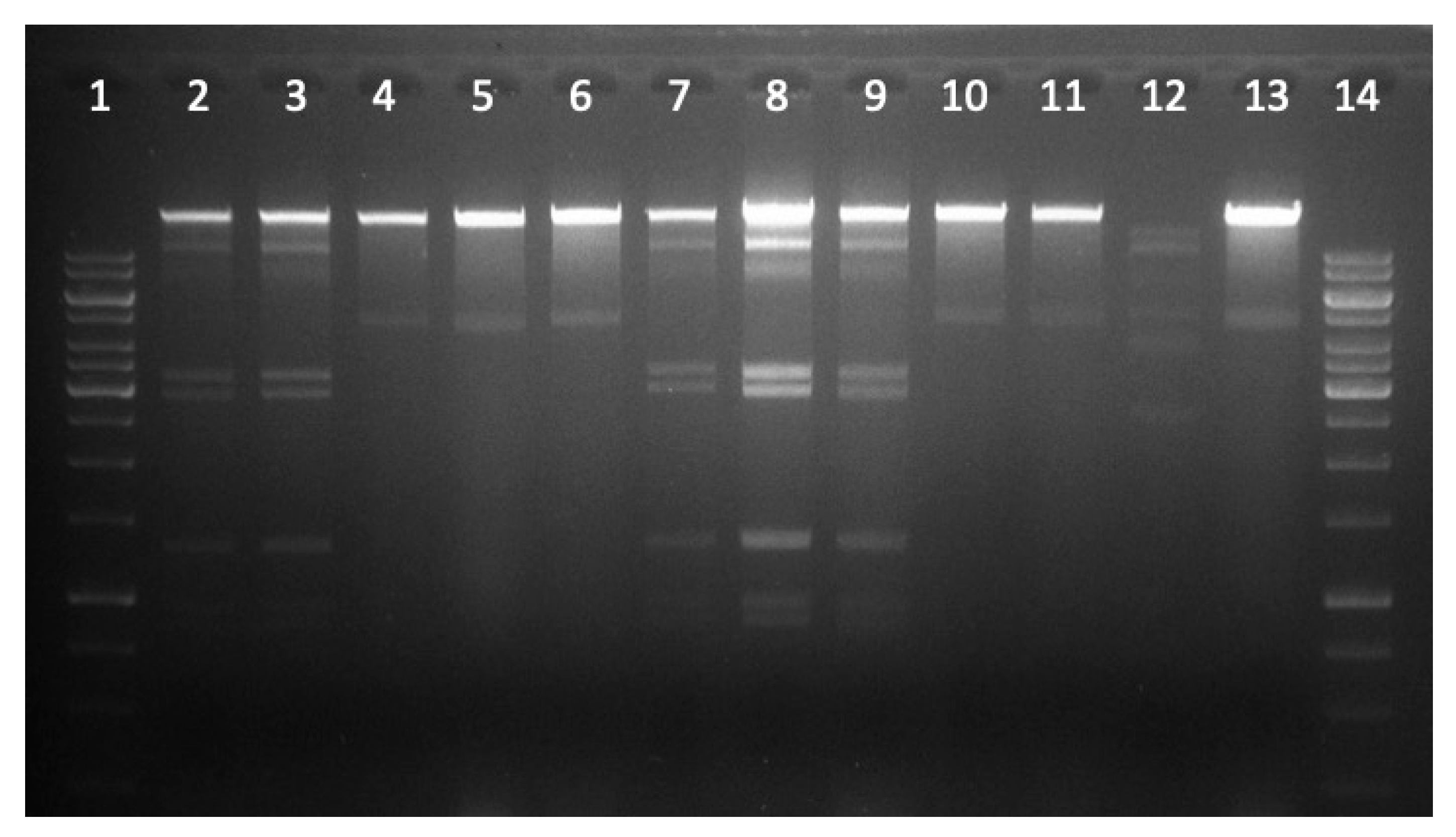
2.6. Restriction Analysis of Selected Phage DNA
2.7. Difference in Lysis Properties of Resistant (Harboring Resistant Genes) versus Non-Resistant E. coli
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Preparation
4.2. Isolation of E. coli and Bacteriophages
4.3. Antimicrobial Resistance Determination in E. coli Isolates
4.4. Antimicrobial Resistance Gene Detection in Phage Lysates
4.5. Transduction of Resistance Genes
4.6. Determining Phage Lysis Profile of E. coli Isolates and Lysis Property of Phages
4.7. Phage DNA Isolation and Restriction Profile
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boerlin, P.; Reid-Smith, R.J. Antimicrobial resistance: Its emergence and transmission. Anim. Health Res. Rev. 2008, 9, 115–126. [Google Scholar] [CrossRef]
- Wiedenbeck, J.; Cohan, F.M. Origins of Bacterial Diversity through horizontal genetic transfer and adaptation to new ecological niches. FEMS Microbiol. Rev. 2011, 35, 957–976. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Ellington, M.J. The Emergence of antibiotic resistance by mutation. Clin. Microbiol. Infect. 2007, 13, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Shousha, A.; Awaiwanont, N.; Sofka, D.; Smulders, F.J.M.; Paulsen, P.; Szostak, M.P.; Humphrey, T.; Hilbert, F. Bacteriophages isolated from chicken meat and the horizontal transfer of antimicrobial resistance genes. Appl. Environ. Microbiol. 2015, 81, 4600–4606. [Google Scholar] [CrossRef] [Green Version]
- Volkova, V.V.; Lu, Z.; Besser, T.; Gröhn, Y.T. Modeling the infection dynamics of bacteriophages in enteric Escherichia coli: Estimating the contribution of transduction to antimicrobial gene spread. Appl. Environ. Microbiol. 2014, 80, 4350–4362. [Google Scholar] [CrossRef] [Green Version]
- von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Sidjabat, H.E.; Townsend, K.M.; Lorentzen, M.; Gobius, K.S.; Fegan, N.; Chin, J.J.; Bettelheim, K.A.; Hanson, N.D.; Bensink, J.C.; Trott, D.J. Emergence and spread of two distinct clonal groups of multidrug-resistant Escherichia coli in a veterinary teaching hospital in Australia. J. Med. Microbiol. 2006, 55, 1125–1134. [Google Scholar] [CrossRef]
- So, J.H.; Kim, J.; Bae, I.K.; Jeong, S.H.; Kim, S.H.; Lim, S.K.; Park, Y.H.; Lee, K. Dissemination of multidrug-resistant Escherichia coli in Korean veterinary hospitals. Diagn. Microbiol. Infect. Dis. 2012, 73, 195–199. [Google Scholar] [CrossRef]
- LeCuyer, T.E.; Byrne, B.A.; Daniels, J.B.; Diaz-Campos, D.V.; Hammac, G.K.; Miller, C.B.; Besser, T.E.; Davis, M.A. Population Structure and Antimicrobial Resistance of Canine Uropathogenic Escherichia Coli. J. Clin. Microbiol. 2018, 56, e00788-18. [Google Scholar] [CrossRef] [Green Version]
- Walther, B.; Tedin, K.; Lübke-Becker, A. Multidrug-resistant opportunistic pathogens challenging veterinary infection control. Vet. Microbiol. 2017, 200, 71–78. [Google Scholar] [CrossRef]
- Hilbert, M.; Csadek, I.; Auer, U.; Hilbert, F. Antimicrobial Resistance-Transducing Bacteriophages Isolated from Surfaces of Equine Surgery Clinics – A Pilot Study. Eur. J. Microbiol. Immunol. 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pacífico, C.; Hilbert, M.; Sofka, D.; Dinhopl, N.; Pap, I.J.; Aspöck, C.; Hilbert, F. Characterization of bacteria and inducible phages in an intensive care unit. J. Clin. Med. 2019, 8, 1433. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, D.H. Reservoirs of antimicrobial resistance in pet animals. Clin. Infect. Dis. 2007, 45 (Suppl. 2), 148–152. [Google Scholar] [CrossRef]
- Yao, H.; Wu, D.; Lei, L.; Shen, Z.; Wang, Y.; Liao, K. The detection of fosfomycin resistance genes in Enterobacteriaceae from pets and their owners. Vet. Microbiol. 2016, 193, 67–71. [Google Scholar] [CrossRef]
- Emmerson, A.M.; Jones, A.M. The quinolones: Decades of development and use. J. Antimicrob. Chemother. 2003, 51 (Suppl. 1), 13–20. [Google Scholar] [CrossRef] [Green Version]
- Iravani, A.; Richard, G.A.; Baer, H.; Fennell, R. Comparative efficacy and safety of nalidixic acid versus trimethoprim/sulfamethoxazole in treatment of acute urinary tract infections in college-age women. Antimicrob. Agents Chemother. 1981, 19, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Zalewska-Piątek, B.; Piątek, R. Phage therapy as a novel strategy in the treatment of urinary tract infections caused by E. Coli. Antibiotics (Basel) 2020, 9, 304. [Google Scholar] [CrossRef]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Pistelli, L.; Mancianti, F. Antimicrobial Activity of Five Essential Oils against Bacteria and Fungi Responsible for Urinary Tract Infections. Molecules 2018, 23, 1668. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Ho, D.V.; Doan, T.Q.; Le, A.T.; Raal, A.; Usai, D.; Sanna, G.; Carta, A.; Rappelli, P.; Diaz, N.; et al. Biological Activities of Essential Oils from Leaves of Paramignya trimera (Oliv.) Guillaum and Limnocitrus littoralis (Miq.) Swingle. Antibiotics (Basel) 2020, 9, 207. [Google Scholar]
- Xiao, S.; Cui, P.; Shi, W.; Zhang, Y. Identification of essential oils with strong activity against stationary phase uropathogenic Escherichia Coli. Discov. Med. 2019, 28, 179–188. [Google Scholar]
- Azam, A.H.; Tanji, Y. Bacteriophage-host arm race: An update on the mechanism of phage resistance in bacteria and revenge of the phage with the perspective for phage therapy. Appl. Microbiol. Biotechnol. 2019, 103, 2121–2131. [Google Scholar] [CrossRef]
- Aidley, J.; Rajopadhye, S.; Akinyemi, N.M.; Lango-Scholey, L.; Jones, M.A.; Bayliss, C.D. Nonselective Bottlenecks Control the Divergence and Diversification of Phase-Variable Bacterial Populations. mBio 2017, 8, e02311-16. [Google Scholar] [CrossRef] [Green Version]
- Henderson, I.R.; Owen, P.; Nataro, J.P. Molecular switches—The ON and OFF of bacterial phase variation. Mol. Microbiol. 1999, 33, 919–932. [Google Scholar] [CrossRef]
- Bäckhed, F.; Alsén, B.; Roche, N.; Angström, J.; von Euler, A.; Breimer, M.E.; Westerlund-Wikström, B.; Teneberg, S.; Richter-Dahlfors, A. Identification of target tissue glycosphingolipid receptors for uropathogenic, F1C-fimbriated Escherichia coli and its role in mucosal inflammation. J. Biol. Chem. 2002, 277, 18198–18205. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.K.; Gunther IV, N.W.; Zhao, H.; Johnson, D.E.; Keay, S.K.; Mobley, H.L.T. In vivo phase variation of Escherichia coli type 1 fimbrial genes in women with urinary tract infection. Infect. Immun. 1998, 66, 3303–3310. [Google Scholar] [CrossRef] [Green Version]
- Safi, H.; Gopal, P.; Lingaraju, S.; Ma, S.; Levine, C.; Dartois, V.; Yee, M.; Li, L.; Blanc, L.; Ho Liang, H.P.; et al. Phase variation in Mycobacterium tuberculosis glpK produces transiently heritable drug tolerance. Proc. Natl. Acad. Sci. USA 2019, 116, 19665–19674. [Google Scholar] [CrossRef] [Green Version]
- ISO (International Organization for Standardization). ISO 10705-2:2000. Water Quality—Detection and Enumeration of Bacteriophages—Part 2: Enumeration of Somatic Coliphages; International Organization for Standardization: Geneva, Switzerland, 2000. [Google Scholar]
- Groisman, E.A. In vivo genetic engeneering with bacteriophage Mu. Escherichia coli and Salmonella typhimurium. Methods Enzymol. 1991, 204, 180–212. [Google Scholar]
- CLSI. VET01. Performance Standards for Antimicrobial Disc and Dilution Susceptibility Tests for Bacteria Isolates from Animals, 5th ed.; CLSI Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MIC’s and Zone Diameters. Version 10.0. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 9 January 2020).
- Chen, S.; Zhao, S.; White, D.G.; Schroeder, C.M.; Lu, R.; Yang, H.; McDermott, P.F.; Ayers, S.; Meng, J. Characterization of multiple-antimicrobial-resistant Salmonella serovars isolated from retail meats. Appl. Environ. Microbiol. 2004, 70, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Navia, M.M.; Ruiz, J.; Sanchez-Cespedes, J.; Vila, J. Detection of dihydrofolate reductase genes by PCR and RFLP. Diagn. Microbiol. Infect. Dis. 2003, 46, 295–298. [Google Scholar] [CrossRef]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic resistance in the ECOR collection: Integrons and identification of a novel aad gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Kerrn, M.B.; Klemmensen, T.; Frimodt-Møller, N.; Espersen, F. Susceptibility of Danish Escherichia coli strains isolated from urinary tract infections and bacteraemia, and distribution of sul genes conferring sulphonamide resistance. J. Antimicrob. Chemother. 2002, 50, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Maidhof, H.; Guerra, B.; Abbas, S.; Elsheikha, H.M.; Whittam, T.S.; Beutin, L. A multiresistant clone of Shiga toxin-producing Escherichia coli O118:[H16] is spread in cattle and humans over different European countries. Appl. Environ. Microbiol. 2002, 68, 5834–5842. [Google Scholar] [CrossRef] [Green Version]
- Colombet, J.; Robin, A.; Lavie, L.; Bettarel, Y.; Cauchie, H.M.; Sime-Ngando, T. Virioplankton ‘pegylation’: Use of PEG (polyethylene glycol) to concentrate and purify viruses in pelagic ecosystems. J. Microbiol. Methods 2007, 71, 212–219. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Resistance | AM R | NA R | S R | S3 R | W R |
|---|---|---|---|---|---|
| blaTEM | 39 | 39 | 34 | 34 | 1 |
| dfr | 1 | 1 | 0 | 0 | 1 |
| sulI | 2 | 2 | 2 | 2 | 0 |
| sulII | 34 | 34 | 34 | 34 | 0 |
| strA | 34 | 34 | 34 | 34 | 0 |
| RP1 | RP2 | RP3 | RP4 | RP5 | RP6 1 |
|---|---|---|---|---|---|
| PNap5 | PNap7 | PNap50 | PNap22 | PNap13 | PNap16 |
| PNap6 | PNap11 | PNap15 | PNap17 | ||
| PNap29 | PNap12 | PNap44 | PNap20 | ||
| PNap31 | PNap19 | PNap45 | PNap56 | ||
| PNap33 | PNap25 | PNap46 | |||
| PNap47 | PNap26 | PNap54 | |||
| PNap49 | PNap28 | ||||
| PNap38 | |||||
| PNap43 | |||||
| PNap53 |
| Primer | Sequence | Target Gene | Amplicon Size | Annealing (°C) | Reference |
|---|---|---|---|---|---|
| blaTEM-1-f | 5′-cagcggtaagatccttgaga-3′ | blaTEM | 643 | 55 | [31] |
| blaTEM-1-r | 5′-actccccgtcgtgtagataa-3′ | ||||
| blaCMY-f | 5′-tggccgttgccgttatctac-3′ | blaCMY | 870 | 55 | [31] |
| blaCMY-r | 5′-cccgttttatgcacccatga-3′ | ||||
| dfr1-f dfr1-r | 5′-gtgaaactatcactaatgg-3′ 5′-ttaacccttttgccagattt-3′ | dfrA1, dfrA5, dfrA15, dfrA15b, dfrA16, dfrA16b | 474 | 55 | [32] |
| sulI-f sulI-r | 5′-tggtgacggtgttcggcattc3′ 5′-gcgaaggtttccgagaaggtg-3′ | sulI | 790 | 63 | [33] |
| sulII-f sulII-r | 5′-gcgctcaaggcagatggcatt-3′ 5′-gcgtttgataccggcacccgt-3′ | sulII | 293 | 60 | [34] |
| strA-B-f strA-B-r | 5′-ccaatcgcagatagaaggcaag-3′ 5′-atcaactggcaggaggaacagg-3′ | strA | 580 | 65 | [35] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zechner, V.; Sofka, D.; Paulsen, P.; Hilbert, F. Antimicrobial Resistance in Escherichia coli and Resistance Genes in Coliphages from a Small Animal Clinic and in a Patient Dog with Chronic Urinary Tract Infection. Antibiotics 2020, 9, 652. https://doi.org/10.3390/antibiotics9100652
Zechner V, Sofka D, Paulsen P, Hilbert F. Antimicrobial Resistance in Escherichia coli and Resistance Genes in Coliphages from a Small Animal Clinic and in a Patient Dog with Chronic Urinary Tract Infection. Antibiotics. 2020; 9(10):652. https://doi.org/10.3390/antibiotics9100652
Chicago/Turabian StyleZechner, Veronika, Dmitrij Sofka, Peter Paulsen, and Friederike Hilbert. 2020. "Antimicrobial Resistance in Escherichia coli and Resistance Genes in Coliphages from a Small Animal Clinic and in a Patient Dog with Chronic Urinary Tract Infection" Antibiotics 9, no. 10: 652. https://doi.org/10.3390/antibiotics9100652