Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Methodology

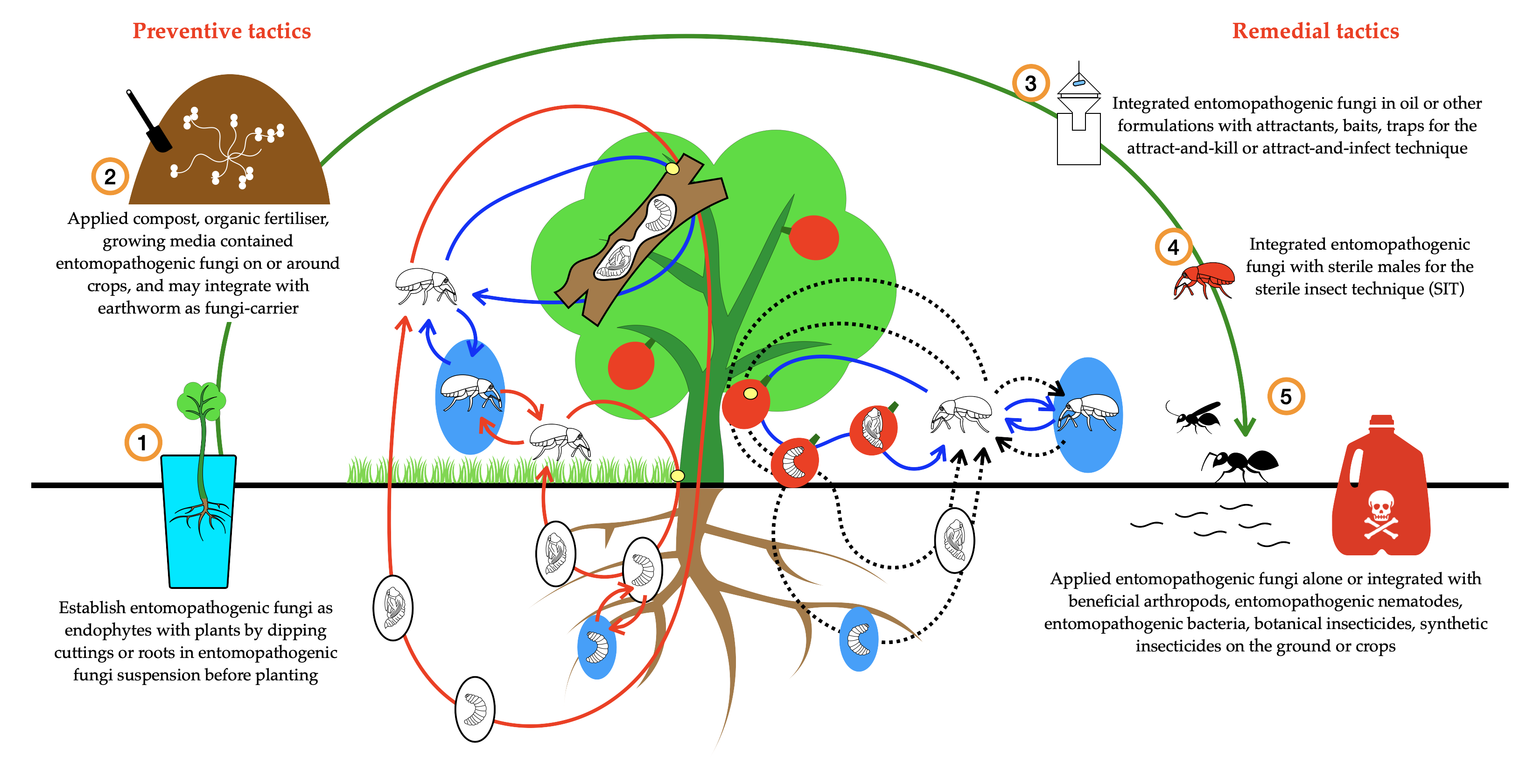{kind=link}
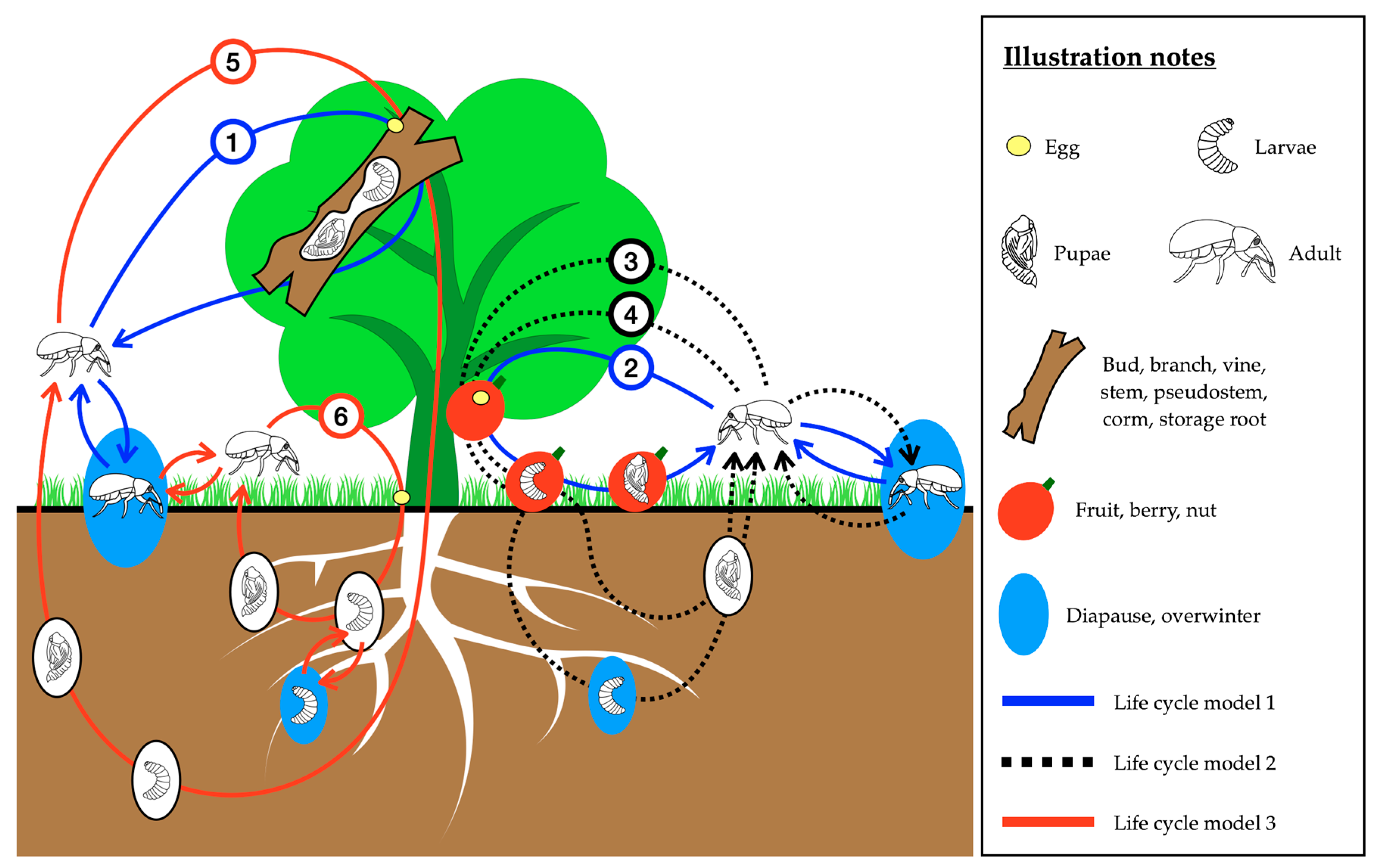{kind=link}
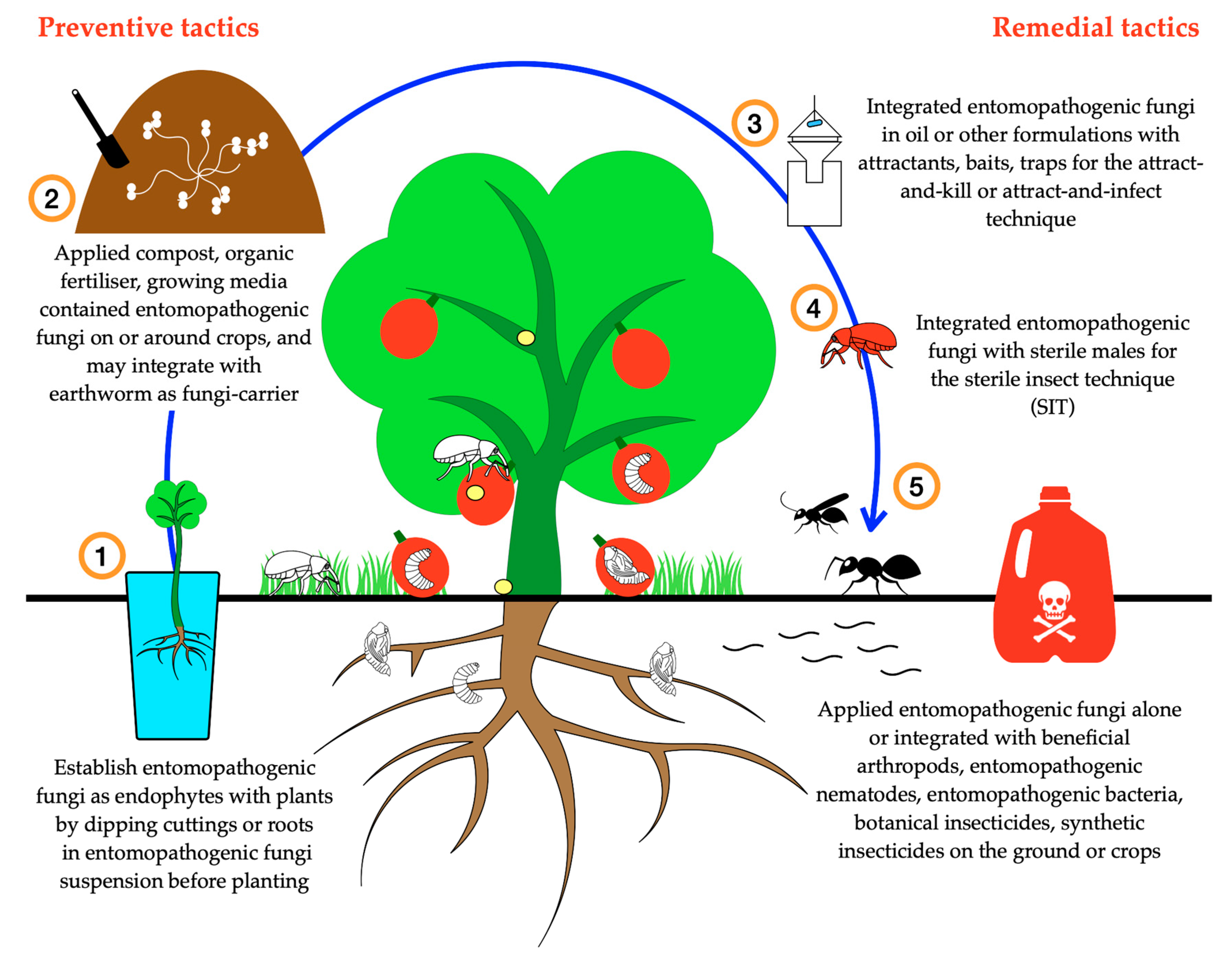{kind=link}
| Weevil Species | Common Name | Family: Subfamily 1 | Distribution 2 | Crops | Damaging Stages 3 | Economic Impact 4 | Ref. |
|---|---|---|---|---|---|---|---|
| Aclees sp. cf. foveatus (Voss) | Fig weevil | Cur: Mol | IT | Fig | A & L | n/a | [35] |
| Aegorhinus superciliosus (Guérin) | Raspberry weevil | Cur: Cyc | AR & CL | Blueberries, raspberries, strawberry | A & L | n/a | [36,37] |
| Anthonomus musculus (Say) | Cranberry weevil | Cur: Cur | North-Eastern US & CA | Blueberries, cranberries | A & L | n/a | [38] |
| Anthonomus piri (Kollar) | Apple bud weevil | Cur: Cur | EUR & GB | Apple, pears | A & L | n/a | [39] |
| Anthonomus pomorum (L.) | Apple blossom weevil | Cur: Cur | EUR | Apple, pears | A & L | n/a | [39] |
| Anthonomus rubi (Herbst) | Strawberry blossom weevil | Cur: Cur | EUR & GB | Strawberry, blackberry, raspberry | A & L | MCL between 36–90% | [40,41] |
| Anthonomus signatus (Say) | Strawberry bud weevil | Cur: Cur | US & CA | Strawberry | A & L | MCL up to 100% in New York & 70% in Quebec | [42] |
| Blosyrus asellus (Olivier) | Rough sweetpotato weevil | Cur: Ent | US | Sweetpotato | A & L | n/a | [43] |
| Conotrachelus nenuphar (Herbst) | Plum curculio | Cur: Mol | Eastern & central NAM (US, CA) | Pome & stone fruits | A & L | MCL up to 85% in unsprayed orchard | [44,45] |
| Conotrachelus psidii (Marshall) | Guava weevil | Cur: Mol | BO, BR, CO, MX, PY & VE | Guava | L | MCL up to 100% in Rio de Janeiro, Brazil | [46,47] |
| Cosmopolites sordidus (Germar) | Banana weevil | Cur: Dry | Tropical regions worldwide | Banana & plantain | A & L | MCL up to 50% | [31,48] |
| Curculio caryae (Horn) | Pecan weevil | Cur: Cur | Southern US | Pecan | A & L | MCL between 30–80% | [29,30] |
| Curculio caryatrypes (Boheman) | Larger chestnut weevil | Cur: Cur | Central-eastern US | Chestnut | A & L | n/a | [49] |
| Curculio elephas (Gyllenhal) | Chestnut weevil | Cur: Cur | Central & Southern EUR, North AFR | Chestnut | A & L | MCL up to 90% in Italy | [39,50] |
| Curculio nucum (L.) | Hazelnut weevil | Cur: Cur | PAL, also present in North AFR | Hazelnut | A & L | MCL up to 80% in the unprotected orchards in Spain | [51,52] |
| Curculio sayi (Gyllenhal) | Lesser chestnut weevil | Cur: Cur | Central-eastern US | Chestnut | A & L | n/a | [49] |
| Curculio sikkimensis (Heller) | Chestnut weevil | Cur: Cur | CN, IN, JP & KR | Chestnut | A & L | n/a | [53,54] |
| Cylas formicarius (F.) | Sweetpotato weevil | Bre: Bre | Tropical regions worldwide | Sweetpotato | A & L | MCL up to 100% | [31,55] |
| Cylas puncticollis (Boheman), C. brunneus (F.) | African sweetpotato weevil | Bre: Bre | AFR (sub-Saharan) | Sweetpotato | A & L | MCL up to 97% | [55,56] |
| Diaprepes abbreviatus (L.) | Citrus root weevil | Cur: Ent | US & several CAR | Citrus, sugarcane | L | n/a | [57] |
| Heilipus lauri (Boheman) | Avocado seed weevil | Cur: Mol | CO & MX | Avocado | A & L | MCL between 60–70% in Mexico | [58,59] |
| Hypothenemus hampei (Ferrari) | Coffee berry borer | Cur: Sco | AFR, ASI, OCE, SCA & US | Coffee | A & L | MCL between 40 - 90%. EAL around US$215–358 million in Brazil or around US$500 million worldwide | [31,32,60] |
| Kuschelorhynchus macadamiae (Jennings & Oberprieler) | Macadamia seed weevil | Cur: Cur | Eastern AU | Macadamia | A & L | MCL up to 15% | [61,62] |
| Odoiporus longicollis (Olivier) | Banana stem weevil | Cur: Dry | Tropical ASI | Banana & plantain | A & L | MCL between 10–90% | [63] |
| Otiorhynchus clavipes (Bonsdorff) | Red-legged weevil | Cur: Ent | Western EUR | Plum, apple, berry crops, grapevine | A & L | n/a | [39] |
| Otiorhynchus ovatus (L.) | Strawberry weevil | Cur: Ent | EUR & NAM | Strawberry, berry crops | A & L | MCL up to 100% in Saxony, Germany | [39] |
| Otiorhynchus rugifrons (Gyllenhal) | Strawberry root weevil | Cur: Ent | EUR | Strawberry | A & L | n/a | [39] |
| Otiorhynchus rugosostriatus (Goeze) | Rough strawberry root weevil | Cur: Ent | EUR, NAM & MED | Strawberry, berry crops | A & L | n/a | [39] |
| Otiorhynchus singularis (L.) | Clay-coloured weevil | Cur: Ent | EUR & NAM | Apple, pear, berry crops, grapevine | A & L | n/a | [39] |
| Otiorhynchus sulcatus (F.) | Black vine weevil | Cur: Ent | EUR, NAM & AUA | Grapevines, berry crops | A & L | n/a | [39,64] |
| Pantorhytes plutus (Oberthür) | Cacao weevil | Cur: Ent | PG | Cacao | L | n/a | [65,66] |
| Phlyctinus callosus (Schönherr) | Banded fruit weevil | Cur: Ent | AU, NZ & ZA | Grapevines, pome fruit, stone fruits | A & L | MCL up to 40% | [67,68] |
| Pityophthorus juglandis (Blackman) | Walnut twig beetle | Cur: Sco | south-western US & MX | Walnut | A & L | n/a | [69] |
| Rhynchophorus bilineatus (Montrouzier) | Black palm weevil | Cur: Dry | ID, PG & SB | Palm | L | n/a | [70] |
| Rhynchophorus cruentatus (F.) | Palmetto weevil | Cur: Dry | Florida & south-eastern US | Palm | L | n/a | [70] |
| Rhynchophorus ferrugineus (Olivier) | Red palm weevil | Cur: Dry | ASI, AU & MED | Palm | L | EAL around US$5–26 million in the Middle East | [71,72] |
| Rhynchophorus palmarum (L.) | American palm weevil | Cur: Dry | MX & SCA | Palm | L | MCL up to 15% | [70,73] |
| Rhynchophorus phoenicis (F.) | African palm weevil | Cur: Dry | AFR | Palm | L | n/a | [70] |
| Rhynchophorus quadrangulus (Queden) | n/a | Cur: Dry | AFR | Palm | L | n/a | [70] |
| Scolytus amygdali (Guérin-Méneville) | Almond bark beetle | Cur: Sco | MED | Almond, apricot, peach | A & L | n/a | [39] |
| Scolytus mali (Bechstein & Scharfenberg) | Large fruit bark beetle | Cur: Sco | EUR & PAL | Apple, plum, pear | A & L | n/a | [39] |
| Scolytus rugulosus (Müller) | Fruit bark beetle | Cur: Sco | EUR | Apple, pear, plum | A & L | n/a | [39] |
| Xyleborus affinis (Eichhoff) | Ambrosia beetle | Cur: Sco | MX & US | Avocado, mango, macadamia, walnut | A & L | n/a | [74,75,76] |
| Weevil Species | Egg (Days) | Larvae (Days) | Pupae (Days) | Adult (Days) | Generation | Ref. |
|---|---|---|---|---|---|---|
| Aclees sp. cf. foveatus | 10–20 | n/a | n/a | n/a | 2 generations/year | [35] |
| Anthonomus signatus | 6–14 | 21–28 | 5–8 | n/a | 32–64 days/generation, 1 generation/year | [42,77] |
| Conotrachelus nenuphar | 2–12 | 14–21 | 30 | n/a | 57 days/generation | [78] |
| Conotrachelus psidii | 2–6 | 8–27 | 14–18 | <418 | 108–280 days/generation | [79] |
| Cosmopolites sordidus | 5–8 | 14–21 | 5–7 | <730 | 1–6 months/generation | [31,80] |
| Curculio caryae | n/a | 30 | 270–1080 | n/a | 2–3 years/generation | [29,81] |
| Curculio elephas | n/a | 730–1095 | 90–150 | n/a | 1 generation/year in Italy | [50,82] |
| Curculio nucum | >7 | 28–35 | < 365 | 90 | 1 generation/year in Turkey | [39,83] |
| Cylas formicarius | 3–7 | 7–11 | 5–7 | <240 | 5–8 generations/year in United States | [31,55] |
| Cylas puncticollis | < 5 | <23 | <14 | <141 | 20–25 days/generation | [56] |
| Diaprepes abbreviatus | 7–10 | 240–450 | 15–30 | <147 | 5–18 months/generation | [84] |
| Heilipus lauri | <13 | <49 | <15 | n/a | 76 days/generation | [58] |
| Hypothenemus hampei | 5–9 | 10–26 | 4–9 | <157 | 25–35 days/generation, >8 generations/year in African countries, 2–3 generations/year in Colombia | [31,85] |
| Kuschelorhynchus macadamiae | 6 | 28 | 4 | n/a | At least 3 generations/year | [86] |
| Odoiporus longicollis | 3–8 | 30–60 | 17–22 | 50–95 | 53–95 days/generation | [87] |
| Otiorhynchus sulcatus | >8 | 84–211 | 10–50 | n/a | 1 generation/year | [64] |
| Pantorhytes plutus | n/a | 90–270 | 14 | 365–730 | 4–11 months/generation | [88] |
| Phlyctinus callosus | 6–15 | n/a | 7–21 | n/a | 1–2 generations/year | [89] |
| Pityophthorus juglandis | n/a | n/a | n/a | n/a | 7 weeks/generation, 2 generations/year | [90] |
| Rhynchophorus ferrugineus | 1–6 | 25–105 | 11–45 | n/a | 45 days/generation in the Philippines, 139 days/generation in Spain; 3–4 generations/year in India, up to 21 generations/year in Egypt | [71] |
| Scolytus amygdali | n/a | n/a | n/a | n/a | >3 generations/year in the Mediterranean area | [91] |
| Life Cycle Model | Subcategory | Weevil Species | Common Name |
|---|---|---|---|
| Model 1: Larvae and Pupae in the Host Plant | 1 | Aclees sp. cf. foveatus | Fig weevil |
| Anthonomus signatus | Strawberry bud weevil | ||
| Blosyrus asellus | Rough sweetpotato weevil | ||
| Cosmopolites sordidus | Banana weevil | ||
| Cylas formicarius | Sweetpotato weevil | ||
| Cylas puncticollis | African Sweetpotato weevil | ||
| Odoiporus longicollis | Banana stem weevil | ||
| Pantorhytes plutus | Cacao weevil | ||
| Pityophthorus juglandis | Walnut twig beetle | ||
| Rhynchophorus ferrugineus | Red palm weevil | ||
| Rhynchophorus bilineatus | Black palm weevil | ||
| Scolytus amygdali | Almond bark beetle | ||
| Xyleborus affinis | Ambrosia beetle | ||
| 2 | Heilipus lauri | Avocado seed weevil | |
| Hypothenemus hampei | Coffee berry borer | ||
| Kuschelorhynchus macadamiae | Macadamia seed weevil | ||
| Model 2: Larvae in the Host Plant and Pupae under the Ground | 3 | Curculio caryae | Pecan weevil |
| Curculio elephas | Chestnut weevil | ||
| Curculio sikkimensis | Chestnut weevil | ||
| Curculio nucum | Hazelnut weevil | ||
| 4 | Conotrachelus nenuphar | Plum curculio | |
| Conotrachelus psidii | Guava weevil | ||
| Model 3: Larvae and Pupae under the Ground | 5 | Diaprepes abbreviatus | Citrus root weevil |
| 6 | Aegorhinus superciliosus | Raspberry weevil | |
| Otiorhynchus sulcatus | Black vine weevil | ||
| Phlyctinus callosus | Banded fruit weevil |
3. Life Cycle Patterns of Weevils Affecting Horticultural Crops
4. Effect of Fungal Entomopathogens on Weevils with Life Cycle Model 1: Larvae and Pupae in the Host Plant
5. Effect of Fungal Entomopathogens on Weevils with Life Cycle Model 2: Larvae in the Host Plant and Pupation under the Ground
6. Effect of Fungal Entomopathogens on Weevils with Life Cycle Model 3: Larvae and Pupae under the Ground
7. Integration of Fungal Entomopathogens in the Integrated Pest Management Programs and Future Research Directions
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hill, D.S. The Economic Importance of Insects; Springer: London, UK, 1997; pp. 1–5. [Google Scholar]
- Culliney, T.W. Chapter 8: Crop losses to arthropods. In Integrated Pest Management: Pesticide Problems; Pimentel, D., Peshin, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 3, pp. 201–225. [Google Scholar]
- Atwood, D.; Paisley-Jones, C. Pesticides Industry Sales and Usage: 2008–2012 Market Estimates; United States Environmental Protection Agency: Washington, DC, USA, 2017. Available online: https://www.epa.gov/sites/production/files/2017-01/documents/pesticides-industry-sales-usage-2016_0.pdf (accessed on 16 July 2020).
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchard IPM program. Biocontrol 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Adamson, D.; Furlong, M.J. The future of IPM: Whither or wither? Aust. J. Entomol. 2009, 48, 85–96. [Google Scholar] [CrossRef]
- Dutcher, J.D. A review of resurgence and replacement causing pest outbreaks in IPM. In General Concepts in Integrated Pest and Disease Management; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 27–43. [Google Scholar]
- Pimentel, D.; Acquay, H.; Biltonen, M.; Rice, P.; Silva, M.; Nelson, J.; Lipner, V.; Giordano, S.; Horowitz, A.; D’Amore, M. Environmental and economic costs of pesticide use. Bioscience 1992, 42, 750–760. [Google Scholar] [CrossRef]
- Pickering, J.; Dutcher, J.D.; Ekbom, B.S. The effect of a fungicide on fungal-induced mortality of pecan aphids (Homoptera: Aphididae) in the field. J. Econ. Entomol. 1990, 83, 1801–1805. [Google Scholar] [CrossRef]
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B 2011, 366, 1987–1998. [Google Scholar] [CrossRef]
- Leahy, J.; Mendelsohn, M.; Kough, J.; Jones, R.; Berckes, N. Biopesticide oversight and registration at the US Environmental Protection Agency. In Biopesticides: State of the Art and Future Opportunities; Gross, A.D., Coats, J.R., Duke, S.O., Seiber, J.N., Eds.; ACS Publications: Washington, DC, USA, 2014; pp. 3–18. [Google Scholar]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef]
- Olson, S. An analysis of the biopesticide market now and where it is going. Outlooks Pest Manag. 2015, 26, 203–206. [Google Scholar] [CrossRef]
- Dolinski, C.; Lacey, L.A. Microbial control of arthropod pests of tropical tree fruits. Neotrop. Entomol. 2007, 36, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Shapiro-Ilan, D.I. Microbial control of insect pests in temperate orchard systems: Potential for incorporation into IPM. Annu. Rev. Entomol. 2008, 53, 121–144. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [Green Version]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. Biocontrol 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Navarro-Llopis, V.; Ayala, I.; Sanchis, J.; Primo, J.; Moya, P. Field efficacy of a Metarhizium anisopliae-based attractant-contaminant device to control Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2015, 108, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Brandl, M.A.; Schumann, M.; Przyklenk, M.; Patel, A.; Vidal, S. Wireworm damage reduction in potatoes with an attract-and-kill strategy using Metarhizium brunneum. J. Pest Sci. 2017, 90, 479–493. [Google Scholar] [CrossRef]
- Toledo, J.; Flores, S.; Campos, S.; Villaseñor, A.; Enkerlin, W.; Liedo, P.; Valle, Á.; Montoya, P. Pathogenicity of three formulations of Beauveria bassiana and efficacy of autoinoculation devices and sterile fruit fly males for dissemination of conidia for the control of Ceratitis capitata. Entomol. Exp. Appl. 2017, 164, 340–349. [Google Scholar] [CrossRef]
- Sookar, P.; Bhagwant, S.; Khayrattee, F.B.; Chooneea, Y.; Ekesi, S. Mating compatibility of wild and sterile melon flies, Bactrocera cucurbitae (Diptera: Tephritidae) treated with entomopathogenic fungi. J. Appl. Entomol. 2014, 138, 409–417. [Google Scholar] [CrossRef]
- Novelo-Rincón, L.F.; Montoya, P.; Hernández-Ortiz, V.; Liedo, P.; Toledo, J. Mating performance of sterile Mexican fruit fly Anastrepha ludens (Dipt., Tephritidae) males used as vectors of Beauveria bassiana (Bals.) Vuill. J. Appl. Entomol. 2009, 133, 702–710. [Google Scholar] [CrossRef]
- Rossoni, C.; Kassab, S.O.; Loureiro, E.D.; Pereira, F.F.; Costa, D.P.; Barbosa, R.H.; Zanuncio, J.C. Metarhizium anisopliae and Beauveria bassiana (Hypocreales: Clavicipitaceae) are compatible with Cotesia flavipes (Hymenoptera: Braconidae). Fla. Entomol. 2014, 97, 1794–1804. [Google Scholar] [CrossRef]
- Labbé, R.M.; Gillespie, D.R.; Cloutier, C.; Brodeur, J. Compatibility of an entomopathogenic fungus with a predator and a parasitoid in the biological control of greenhouse whitefly. Biocontrol Sci. Technol. 2009, 19, 429–446. [Google Scholar] [CrossRef]
- Al Mazra’awi, M.S.; Shipp, J.L.; Broadbent, A.B.; Kevan, P.G. Dissemination of Beauveria bassiana by honey bees (Hymenoptera: Apidae) for control of tarnished plant bug (Hemiptera: Miridae) on canola. Environ. Entomol. 2006, 35, 1569–1577. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Characterization of the synergistic interaction between Beauveria bassiana strain GHA and Bacillus thuringiensis morrisoni strain tenebrionis applied against Colorado potato beetle larvae. J. Invertebr. Pathol. 2017, 144, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Sayed, A.M.M.; Behle, R.W. Evaluating a dual microbial agent biopesticide with Bacillus thuringiensis var. kurstaki and Beauveria bassiana blastospores. Biocontrol Sci. Technol. 2017, 27, 461–474. [Google Scholar]
- Duarte, R.T.; Gonçalves, K.C.; Espinosa, D.J.L.; Moreira, L.F.; De Bortoli, S.A.; Humber, R.A.; Polanczyk, R.A. Potential of entomopathogenic fungi as biological control agents of diamondback moth (Lepidoptera: Plutellidae) and compatibility with chemical insecticides. J. Econ. Entomol. 2016, 109, 594–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niassy, S.; Maniania, N.K.; Subramanian, S.; Gitonga, M.L.; Maranga, R.; Obonyo, A.B.; Ekesi, S. Compatibility of Metarhizium anisopliae isolate ICIPE 69 with agrochemicals used in French bean production. Int. J. Pest Manag. 2012, 58, 131–137. [Google Scholar] [CrossRef]
- Ree, B.; Knutson, A.E.; Harris, M. Controlling the Pecan Weevil. Texas Extension E-343. 2005. Available online: http://gregg.agrilife.org/files/2011/09/controllingthepecanweevil_1.pdf (accessed on 16 July 2020).
- Mulder, P.G.; Harris, M.K.; Grantham, R.A. Biology and management of the pecan weevil (Coleoptera: Curculionidae). J. Integr. Pest Manag. 2012, 3, A1–A9. [Google Scholar] [CrossRef]
- Muniappan, R.; Shepard, B.M.; Carner, G.R.; Ooi, P.A.C. Arthropod Pests of Horticultural Crops in Tropical Asia; CABI: Wallingford, UK, 2012; pp. 52–134. [Google Scholar]
- Infante, F.; Pérez, J.; Vega, F.E. Redirect research to control coffee pest. Nature 2012, 489, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullan, P.J.; Cranston, P.S. The Insects: An. Outline of Entomology, 5th ed.; John Wiley & Sons: Oxford, UK, 2014; pp. 418–456. [Google Scholar]
- Oberprieler, R.G.; Marvaldi, A.E.; Anderson, R.S. Weevils, weevils, weevils everywhere. Zootaxa 2007, 1668, 491–520. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Meregalli, M.; Canale, A. Field observations on the mating behavior of Aclees sp. cf. foveatus Voss (Coleoptera: Curculionidae), an exotic pest noxious to fig orchards. J. Insect Behav. 2014, 27, 419–427. [Google Scholar]
- Parra, L.; Mutis, A.; Ceballos, R.; Lizama, M.; Pardo, F.; Perich, F.; Quiroz, A. Volatiles released from Vaccinium corymbosum were attractive to Aegorhinus superciliosus (Coleoptera: Curculionidae) in an olfactometric bioassay. Environ. Entomol. 2009, 38, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, J.; Urzúa, A.; Tampe, J.; Parra, L.; Quiroz, A. Repellent activity of the essential oil from the heartwood of Pilgerodendron uviferum (D. Don) Florin against Aegorhinus superciliosus (Coleoptera: Curculionidae). Molecules 2016, 21, 533. [Google Scholar] [CrossRef] [Green Version]
- Szendrei, Z.; Averill, A.; Alborn, H.; Rodriguez-Saona, C. Identification and field evaluation of attractants for the cranberry weevil, Anthonomus musculus Say. J. Chem. Ecol. 2011, 37, 387–397. [Google Scholar] [CrossRef]
- Alford, D.V. Pest of Fruit Crops: A Colour Handbook, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 152–174. [Google Scholar]
- Cross, J.V.; Burgess, C.M. Strawberry fruit yield and quality responses to flower bud removal: A simulation of damage by strawberry blossom weevil (Anthonomus rubi). J. Hortic. Sci. Biotechnol. 1998, 73, 676–680. [Google Scholar] [CrossRef]
- Cross, J.V.; Easterbrook, M.A.; Crook, A.M.; Crook, D.; Fitz Gerald, J.D.; Innocenzi, P.J.; Jay, C.N.; Solomon, M.G. Review: Natural enemies and biocontrol of pests of strawberry in northern and central Europe. Biocontrol Sci. Technol. 2001, 11, 165–216. [Google Scholar]
- Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Gregoire, J.C.; Miret, J.A.J.; Navarro, M.N.; et al. Pest categorisation of Anthonomus signatus. EFSA J. 2017, 15, 4882. [Google Scholar]
- Pulakkatu-thodi, I.; Motomura-Wages, S.; Miyasaka, S. Evaluation of insecticides for the management of rough sweetpotato weevil, Blosyrus asellus (Coleoptera: Curculionidae) in Hawai’i island. Crop Prot. 2018, 114, 223–227. [Google Scholar]
- Racette, G.; Chouinard, G.; Vincent, C.; Hill, S.B. Ecology and management of plum curculio, Conotrachelus nenuphar [Coleoptera: Curculionidae], in apple orchards. Phytoprotection 1992, 73, 85–100. [Google Scholar]
- Leskey, T.C.; Wright, S.E. Monitoring plum curculio, Conotrachelus nenuphar (Coleoptera: Curculionidae), populations in apple and peach orchards in the mid-Atlantic. J. Econ. Entomol. 2004, 97, 79–88. [Google Scholar]
- Da Rosa, J.M.; Boff, M.I.C.; Nunes, M.Z.; Agostinetto, L.; Boff, P. Damage caused by Conotrachelus psidii (Coleoptera: Curculionidae) to the fruits of feijoa (Acca sellowiana). Rev. Colomb. Entomol. 2015, 41, 12–17. [Google Scholar]
- Del Valle, E.E.; Dolinski, C.; Barreto, E.L.S.; Souza, R.M.; Samuels, R.I. Efficacy of Heterorhabditis baujardi LPP7 (Nematoda: Rhabditida) applied in Galleria mellonella (Lepidoptera: Pyralidae) insect cadavers to Conotrachelus psidii, (Coleoptera: Curculionidae) larvae. Biocontrol Sci. Technol. 2008, 18, 33–41. [Google Scholar]
- Rukazambuga, N.D.T.M.; Gold, C.S.; Gowen, S.R. Yield loss in East African highland banana (Musa spp., AAA-EA group) caused by the banana weevil, Cosmopolites sordidus Germar. Crop Prot. 1998, 17, 581–589. [Google Scholar]
- Keesey, I.W.; Barrett, B.A. Seasonal occurrence and soil distribution of the lesser chestnut weevil, Curculio sayi (Coleoptera: Curculionidae) in Mid-Missouri. J. Kans. Entomol. Soc. 2008, 81, 345–354. [Google Scholar]
- Paparatti, B.; Speranza, S. Biological control of chestnut weevil (Curculio elephas Gyll.; Coleoptera, Curculionidae) with the entomopathogen fungus Beauveria bassiana (Balsamo) Vuill. (Deuteromycotina, Hyphomycetes). In Proceedings of the 2nd International Symposium on Chestnut, Bordeaux, France, 19 October 1998; Salesses, G., Ed.; International Society for Horticultural Science: Leuven, Belgium, 1999; pp. 459–464. [Google Scholar]
- Guidone, L.; Valentini, N.; Rolle, L.; Me, G.; Tavella, L. Early nut development as a resistance factor to the attacks of Curculio nucum (Coleoptera: Curculionidae). Ann. Appl. Biol. 2007, 150, 323–329. [Google Scholar] [CrossRef]
- Batalla-Carrera, L.; Morton, A.; Garcia-del-Pino, F. Field efficacy against the hazelnut weevil, Curculio nucum and short-term persistence of entomopathogenic nematodes. Span. J. Agric. Res. 2013, 11, 1112–1119. [Google Scholar] [CrossRef]
- Pelsue, F.W.; Zhang, R.Z. A review of the Genus Curculio from China with descriptions of fourteen new species. Part IV. The Curculio sikkimensis (Heller) group (Coleoptera: Curculionidae: Curculioninae: Curculionini). Coleopt. Bull. 2003, 57, 311–333. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yoon, C.M.; Shin, S.C.; Choi, K.S.; Kim, G.H. Seasonal occurrence of the larvae and adults of chestnut weevil, Curculio sikkimensis (Coleoptera: Curculionidae). Korean J. Appl. Entomol. 2008, 47, 9–15. [Google Scholar] [CrossRef]
- Reddy, P.P. Plant Protection in Tropical Root and Tuber Crops; Springer: New Delhi, India, 2015; pp. 87–98. [Google Scholar]
- Smit, N.E.J.M.; van Huis, A. Biology of the African sweetpotato weevil species Cylas puncticollis (Boheman) and C. brunneus (Fabricius) (Coleoptera: Apionidae). Int. J. Trop. Insect Sci. 1998, 18, 93–100. [Google Scholar] [CrossRef]
- Weissling, T.J.; Peña, J.E.; Giblin-Davis, R.M.; Knapp, J.L., Jr. Diaprepes root weevil, Diaprepes abbreviatus (Linnaeus) (Insecta: Coleoptera: Curculionidae). IFAS Extension EENY-024. 2009. Available online: https://edis.ifas.ufl.edu/pdffiles/IN/IN15100.pdf (accessed on 16 July 2020).
- Diaz, V.; Caicedo, A.M.; Carabali, A. Life cycle and morphological description of Heilipus lauri Boheman (Coleoptera: Curculionidae) in Colombia. Acta Zool. Mex. 2017, 33, 231–242. [Google Scholar]
- Castañeda-Vildozola, Á.; Franco-Mora, O.; De Jesús Pérez-Lopez, D.; Nava-Díaz, C.; Carrasco, J.V.; Vargas-Rojas, L. Association of Heilipus lauri Boheman and Conotrachelus perseae Barber (Coleoptera: Curculionidae) on avocado in Mexico. Coleopt. Bull. 2013, 67, 116–118. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Economic impact of exotic insect pests in Brazilian agriculture. J. Appl. Entomol. 2013, 137, 1–15. [Google Scholar] [CrossRef]
- Bright, J. Macadamia Seed Weevil (Kuschelorhynchus macadamiae) Orchard Management. Primefact 1585. 2017. Available online: https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0008/731987/Macadamia-seed-weevil-update-orchard-management_2.pdf (accessed on 16 July 2020).
- Huwer, R. Ecology and Management of Sigastus Weevil in Macadamias; Horticulture Innovation Australia Limited: Sydney, Australia, 2016; Available online: https://www.horticulture.com.au/globalassets/laserfiche/assets/project-reports/mc15010/mc15010-final-report-514.pdf (accessed on 16 July 2020).
- Padmanaban, B.; Sathiamoorthy, S. The Banana Stem Weevil Odoiporus longicollis. Musa Pest Fact Sheet No. 5. 2001. Available online: https://www.bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/The_Banana_stem_weevil_Odoiporus_longicollis_756.pdf (accessed on 16 July 2020).
- Moorhouse, E.R.; Charnley, A.K.; Gillespie, A.T. A review of the biology and control of the vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae). Ann. Appl. Biol. 1992, 121, 431–454. [Google Scholar] [CrossRef]
- Prior, C.; Jollands, P.; Le Patourel, G. Infectivity of oil and water formulations of Beauveria bassiana (Deuteromycotina: Hyphomycetes) to the cocoa weevil pest Pantorhytes plutus (Coleoptera: Curculionidae). J. Invertebr. Pathol. 1988, 52, 66–72. [Google Scholar] [CrossRef]
- Gressitt, J.L. The weevil genus Pantorhytes (Coleoptera) involving cacao pests and epizoic symbiosis with cryptogamic plants and microfauna. Pac. Insects 1966, 8, 915–965. [Google Scholar]
- Ferreira, T.; Malan, A.P. Potential of entomopathogenic nematodes for the control of the banded fruit weevil, Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae). J. Helminthol. 2014, 88, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Witt, A.B.R.; Little, R.M.; Crowe, T.M. The effectiveness of helmeted guineafowl Numida meleagris (Linnaeus 1766) in controlling the banded fruit weevil Phlyctinus callosus (Schönherr 1826), and their impact on other invertebrates in apple orchards in the Western Cape Province, South Africa. Agric. Ecosyst. Environ. 1995, 55, 169–179. [Google Scholar] [CrossRef]
- Seybold, S.J.; Coleman, T.W.; Dallara, P.L.; Dart, N.L.; Graves, A.D.; Pederson, L.A.; Spichiger, S.E. Recent collecting reveals new state records and geographic extremes in the distribution of the walnut twig beetle, Pityophthorus juglandis Blackman (Coleoptera: Scolytidae), in the United States. Pan-Pac. Entomol. 2012, 88, 277–280. [Google Scholar] [CrossRef]
- Wattanapongsiri, A. A Revision to the Genera Rhynchophorus and Dynamis (Coleoptera: Curculionidae). Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1965. [Google Scholar]
- Faleiro, J.R. A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar]
- El-Sabea, A.M.R.; Faleiro, J.R.; Abo-El-Saad, M.M. The threat of red palm weevil Rhynchophorus ferrugineus to date plantations of the Gulf region in the Middle-East: An economic perspective. Outlooks Pest Manag. 2009, 20, 131–134. [Google Scholar] [CrossRef]
- Oehlschlager, A.C.; Chinchilla, C.; Castillo, G.; Gonzalez, L. Control of red ring disease by mass trapping of Rhynchophorus palmarum (Coleoptera: Curculionidae). Fla. Entomol. 2002, 85, 507–513. [Google Scholar] [CrossRef]
- Chang, V.C.S. Macadamia quick decline and Xyleborus beetles (Coleoptera: Scolytidae). Int. J. Pest Manag. 1993, 39, 144–148. [Google Scholar] [CrossRef]
- Lona, I.D.; Miller, D.G., III; Hatfield, C.A.; Rosecrance, R.C.; Nelson, L.J.; Audley, J.P.; Siefker, M.A.; Chen, Y.; Seybold, S.J. Host selection behavior mediated by differential landing rates of the walnut twig beetle, Pityophthorus juglandis, and associated subcortical insect species, on two western North American walnut species, Juglans californica and J. major. Entomol. Exp. Appl. 2020, 168, 240–258. [Google Scholar] [CrossRef]
- Castrejón-Antonio, J.E.; Tamez-Guerra, P.; Montesinos-Matias, R.; Ek-Ramos, M.J.; Garza-López, P.M.; Arredondo-Bernal, H.C. Selection of Beauveria bassiana (Hypocreales: Cordycipitaceae) strains to control Xyleborus affinis (Curculionidae: Scolytinae) females. PeerJ 2020, 8, e9472. [Google Scholar] [CrossRef]
- Mailloux, G.; Bostanian, N.J. Development of the strawberry bud weevil (Coleoptera: Curculionidae) in strawberry fields. Ann. Entomol. Soc Am. 1993, 86, 384–393. [Google Scholar] [CrossRef]
- Eaton, A.T. Plum Curculio. UNH Cooperative Extension. 2018. Available online: https://extension.unh.edu/resources/files/Resource002799_Rep4154.pdf (accessed on 16 July 2020).
- Bailez, O.E.; Viana-Bailez, A.M.; de Lima, J.O.G.; Moreira, D.D.O. Life-history of the guava weevil, Conotrachelus psidii Marshall (Coleoptera: Curculionidae), under laboratory conditions. Neotrop. Entomol. 2003, 32, 203–207. [Google Scholar] [CrossRef]
- Hill, D.S. Pests of Crops in Warmer Climates and Their Control; Springer: London, UK, 2008; p. 329. [Google Scholar]
- Cottrell, T.E.; Wood, B.W. Movement of adult pecan weevils Curculio caryae within pecan orchards. Agric. For. Entomol. 2008, 10, 363–373. [Google Scholar] [CrossRef]
- Venette, R.; Davis, E.; Heisler, H.; Larson, M. Mini Risk Assessment, Chestnut Weevil, Curculio elephas (Gyllenhal), [Coleoptera: Curculionidae]. 2003. Available online: http://download.ceris.purdue.edu/file/336 (accessed on 16 July 2020).
- Tuncer, C.; Ecevit, O. Current status of hazelnut pests in Turkey. In Proceedings of the 4th International Symposium on Hazelnut, Ordu, Turkey, 30 July 1996; Köksal, A.I., Okay, Y., Günes, N.T., Eds.; International Society for Horticultural Science: Leuven, Belgium, 1997; pp. 545–552. [Google Scholar]
- Grafton-Cardwell, E.; Godfrey, K.; Peña, J.; McCoy, C.; Luck, R. Diaprepes Root Weevil. ANR Publication 8131. 2004. Available online: https://ucanr.edu/datastoreFiles/391-265.pdf (accessed on 16 July 2020).
- Damon, A. A review of the biology and control of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entomol. Res. 2000, 90, 453–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bright, J. Sigastus Weevil Update. Part 1. Life Cycle and Monitoring Keys to Control; Australian Macadamia Society Ltd.: Lismore, Australia, 2017; Available online: https://www.horticulture.com.au/globalassets/hort-innovation/resource-assets/mc-ipm-program-sigastus-weevil-fact-sheet.pdf (accessed on 16 July 2020).
- Prabhavathi, M.K.; Ghosh, S.K. Studies on the interaction between Odoiporous longicollis and endophytic Beauveria bassiana by establishing fungal infection to bsw in the plant system. Int. J. Plant Prot. 2014, 7, 312–317. [Google Scholar] [CrossRef]
- Tsatsia, H.; Jackson, G. Cocoa Weevil Borer. Available online: http://www.pestnet.org/fact_sheets/cocoa_weevil_borer_061.htm (accessed on 23 May 2018).
- Dlamini, B.E.; Addison, P.; Malan, A.P. A review of the biology and control of Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae), with special reference to biological control using entomopathogenic nematodes and fungi. Afr. Entomol. 2019, 27, 279–288. [Google Scholar] [CrossRef]
- Mayfield, A.E.; Juzwik, J.; Scholer, J.; Vandenberg, J.D.; Taylor, A. Effect of bark application with Beauveria bassiana and permethrin insecticide on the walnut twig beetle (Coleoptera: Curculionidae) in black walnut bolts. J. Econ. Entomol. 2019, 112, 2493–2496. [Google Scholar] [CrossRef] [Green Version]
- Mendel, Z.; Ben-Yehuda, S.; Marcus, R.; Nestel, D. Distribution and extent of damage by Scolytus spp. to stone and pome fruit orchards in Israel. Int. J. Trop. Insect Sci. 1997, 17, 175–181. [Google Scholar] [CrossRef]
- Gargani, E.; Mazza, G.; Benvenuti, C.; Torrini, G.; Strangi, A.; Pennacchio, F.; Roversi, P.F. Biological control of Aclees sp. cf. foveatus and first recovery of an associate Beauveria bassiana strain. Redia 2016, 99, 29–33. [Google Scholar]
- Sabbahi, R.; Merzouki, A.; Guertin, C. Efficacy of Beauveria bassiana against the strawberry pests, Lygus lineolaris, Anthonomus signatus and Otiorhynchus ovatus. J. Appl. Entomol. 2008, 132, 151–160. [Google Scholar] [CrossRef]
- Lopes, R.B.; Michereff-Filho, M.; Tigano, M.S.; Neves, P.M.O.J.; López, E.L.; Fancelli, M.; da Silva, J.P. Virulence and horizontal transmission of selected Brazilian strains of Beauveria bassiana against Cosmopolites sordidus under laboratory conditions. Bull. Insectol. 2011, 64, 201–208. [Google Scholar]
- Lopes, R.B.; Mesquita, A.L.M.; Tigano, M.S.; Souza, D.A.; Martins, I.; Faria, M. Diversity of indigenous Beauveria and Metarhizium spp. in a commercial banana field and their virulence toward Cosmopolites sordidus (Coleoptera: Curculionidae). Fungal Ecol. 2013, 6, 356–364. [Google Scholar] [CrossRef]
- Fancelli, M.; Dias, A.B.; Delalibera, I.; de Jesus, S.C.; do Nascimento, A.S.; de Oliveira e Silva, S.; Caldas, R.C.; Ledo, C.A.S. Beauveria bassiana strains for biological control of Cosmopolites sordidus (Germ.) (Coleoptera: Curculionidae) in plantain. Biomed Res. Int. 2013, 2013, 184756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, D.N.; Chávez, M.A.A.; Gutiérrez, R.L.; Cupul, W.C.; Ochoa, J.M.; Velasco, E.G. Suitability of Cordyceps bassiana and Metarhizium anisopliae for biological control of Cosmopolites sordidus (Germar) (Coleoptera: Curculionidae) in an organic Mexican banana plantation: Laboratory and field trials. J. Plant Dis. Prot. 2018, 125, 73–81. [Google Scholar] [CrossRef]
- Omukoko, C.A.; Maniania, K.N.; Wesonga, J.M.; Kahangi, E.M.; Wamocho, L.S. Pathogenicity of isolates of beauveria bassiana to the banana weevil Cosmopolites sordidus. J. Agric. Sci. Technol. 2011, 13, 3–14. [Google Scholar]
- Omukoko, C.A.; Maniania, K.N.; Wesonga, J.M.; Kahangi, E.M.; Wamocho, L.S. Virulence of three strains of Beauveria bassiana against the banana weevil. J. Agric. Biol. Sci. 2014, 9, 333–336. [Google Scholar]
- Omukoko, C.A.; Wesonga, J.M.; Maniania, K.N.; Kahangi, E.M.; Wamocho, L.S. Screening of Beauveria bassiana isolates to the banana weevil and horizontal transmission under laboratory conditions. J. Agric. Sci. Technol. 2014, 16, 1–12. [Google Scholar]
- Maharaj, K.; Khan, A. Efficacy of banana spray oil, mineral oil and water formulations of Beauveria bassiana Balsamo for the control of Cosmopolites sordidus Germar (Coleoptera: Curculionidae) in Musa spp. Int. J. Trop. Agric. 2016, 34, 1455–1460. [Google Scholar]
- Membang, G.; Ambang, Z.; Mahot, H.C.; Kuate, A.F.; Fiaboe, K.K.M.; Hanna, R. Cosmopolites sordidus (Germar) susceptibility to indigenous Cameroonian Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metsch.) isolates. J. Appl. Entomol. 2020, 144, 468–480. [Google Scholar] [CrossRef]
- Dotaona, R.; Wilson, B.A.L.; Stevens, M.M.; Holloway, J.; Ash, G.J. Screening of tropical isolates of Metarhizium anisopliae (Hypocreales: Clavicipitaceae) for virulence to the sweetpotato weevil, Cylas formicarius (Coleoptera: Brentidae). Int. J. Trop. Insect Sci. 2015, 35, 153–163. [Google Scholar] [CrossRef]
- Saputro, T.B.; Prayogo, Y.; Rohman, F.L.; Alami, N.H. The virulence improvement of Beauveria bassiana in infecting Cylas formicarius modulated by various chitin based compounds. Biodiversitas 2019, 20, 2486–2493. [Google Scholar] [CrossRef] [Green Version]
- Ondiaka, S.; Maniania, N.K.; Nyamasyo, G.H.N.; Nderitu, J.H. Virulence of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae to sweetpotato weevil Cylas puncticollis and effects on fecundity and egg viability. Ann. Appl. Biol. 2008, 153, 41–48. [Google Scholar] [CrossRef]
- Clavijo, A.P.; Holguin, C.M. Pathogenicity of commercial entomopathogenic fungal strains on the avocado seed borer (ASB), Heilipus lauri (Coleoptera: Curculionidae) under laboratory conditions. Int. J. Trop. Insect Sci. 2020. [Google Scholar] [CrossRef]
- De la Rosa-Reyes, W.; Godinez-Aguilar, J.L.; Alatorre-Rosas, R. Biological activity of five strains of Metarhizium anisopliae, upon the coffee berry borer Hypothenemus hampei (Col.: Scolytidae). Entomophaga 1995, 40, 403–412. [Google Scholar] [CrossRef]
- De la Rosa, W.; Alatorre, R.; Trujillo, J.; Barrera, J.F. Virulence of Beauveria bassiana (Deuteromycetes) strains against the coffee berry borer (Coleoptera: Scolytidae). J. Econ. Entomol. 1997, 90, 1534–1538. [Google Scholar] [CrossRef]
- Haraprasad, N.; Niranjana, S.R.; Prakash, H.S.; Shetty, H.S.; Wahab, S. Beauveria bassiana—A potential mycopesticide for the efficient control of coffee berry borer, Hypothenemus hampei (Ferrari) in India. Biocontrol Sci. Technol. 2001, 11, 251–260. [Google Scholar] [CrossRef]
- Pava-Ripoll, M.; Posada, F.J.; Momen, B.; Wang, C.; St. Leger, R.J. Increased pathogenicity against coffee berry borer, Hypothenemus hampei (Coleoptera: Curculionidae) by Metarhizium anisopliae expressing the scorpion toxin (AaIT) gene. J. Invertebr. Pathol. 2008, 99, 220–226. [Google Scholar] [CrossRef]
- Samuels, R.I.; Pereira, R.C.; Gava, C.A.T. Infection of the coffee berry borer Hypothenemus hampei (Coleoptera: Scolytidae) by Brazilian isolates of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae (Deuteromycotina: Hyphomycetes). Biocontrol Sci. Technol. 2002, 12, 631–635. [Google Scholar] [CrossRef]
- Varela, A.; Morales, E. Characterization of some Beauveria bassiana isolates and their virulence toward the coffee berry borer Hypothenemus hampei. J. Invertebr. Pathol. 1996, 67, 147–152. [Google Scholar] [CrossRef]
- Vera, J.T.; Montoya, E.C.; Benavides, P.; Góngora, C.E. Evaluation of Beauveria bassiana (Ascomycota: Hypocreales) as a control of the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) emerging from fallen, infested coffee berries on the ground. Biocontrol Sci. Technol. 2011, 21, 1–14. [Google Scholar] [CrossRef]
- Cruz, L.P.; Gaitan, A.L.; Gongora, C.E. Exploiting the genetic diversity of Beauveria bassiana for improving the biological control of the coffee berry borer through the use of strain mixtures. Appl. Microbiol. Biotechnol. 2006, 71, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Mota, L.H.C.; Silva, W.D.; Sermarini, R.A.; Demétrio, C.G.B.; Bento, J.M.S.; Delalibera, I. Autoinoculation trap for management of Hypothenemus hampei (Ferrari) with Beauveria bassiana (Bals.) in coffee crops. Biol. Control 2017, 111, 32–39. [Google Scholar] [CrossRef]
- Posada-Flórez, F.J. Production of Beauveria bassiana fungal spores on rice to control the coffee berry borer, Hypothenemus hampei, in Colombia. J. Insect Sci. 2008, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, M.M.; Prakash, R.N. Infectivity of ten Metarhizium anisopliae isolates to the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae). J. Entomol. Zool. Stud. 2014, 2, 246–249. [Google Scholar]
- Belay, Y.C.; Tenkegna, T.A. Bioassay and pilot mass production of entomopathogenic fungus, Beauveria bassiana for the control of coffee berry borer (Hypothenemus hampei: Scolytidae), Ferrari. J. Appl. Biosci. 2017, 117, 11669–11683. [Google Scholar]
- Khun, K.K.; Ash, G.J.; Stevens, M.M.; Huwer, R.K.; Wilson, B.A.L. Response of the macadamia seed weevil Kuschelorhynchus macadamiae (Coleoptera: Curculionidae) to Metarhizium anisopliae and Beauveria bassiana in laboratory bioassays. J. Invertebr. Pathol. 2020, 174, 107437. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, B.; Thangavelu, R.; Gopi, M.; Mustaffa, M.M. Effect of mass multiplication media on sporulation, field efficacy and shelf life of Beauveria bassiana against rhizome and pseudostem weevils of banana. J. Biol. Control 2009, 23, 277–283. [Google Scholar]
- Alagesan, A.; Padmanaban, B.; Tharani, G.; Jawahar, S.; Manivannan, S. An assessment of biological control of the banana pseudostem weevil Odoiporus longicollis (Olivier) by entomopathogenic fungi Beauveria bassiana. Biocatal. Agric. Biotechnol. 2019, 20, 101262. [Google Scholar] [CrossRef]
- Awasthi, N.S.; Sridharan, S.; Mohankumar, S. In vitro evaluation of native isolate of Metarhizium anisopliae (Metchinkoff) Sorokin and its oil in water formulations against Odoiporus longicollis Olivier. J. Biol. Control 2017, 31, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, L.A.; Mayfield, A.E.; Griggs, M.H.; Camp, R.; Mudder, B.; Taylor, A.; Vandenberg, J.D. Mortality and reduced brood production in walnut twig beetles, Pityophthorus juglandis (Coleoptera: Curculionidae), following exposure to commercial strains of entomopathogenic fungi Beauveria bassiana and Metarhizium brunneum. Biol. Control 2017, 114, 79–86. [Google Scholar] [CrossRef]
- Abdel-Samad, S.S.M.; Mahmoud, B.A.; Abbas, M.S.T. Evaluation of the fungus, Beauveria bassiana (Bals.) Vuill as a bio-control agent against the red palm weevil, Rhynchophorus ferrugineus (Oliv.) (Coleoptera: Curculionidae). Egypt J. Biol. Pest Control 2011, 21, 125–129. [Google Scholar]
- Dembilio, Ó.; Quesada-Moraga, E.; Santiago-Álvarez, C.; Jacas, J.A. Potential of an indigenous strain of the entomopathogenic fungus Beauveria bassiana as a biological control agent against the red palm weevil, Rhynchophorus ferrugineus. J. Invertebr. Pathol. 2010, 104, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Gindin, G.; Levski, S.; Glazer, I.; Soroker, V. Evaluation of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana against the red palm weevil Rhynchophorus ferrugineus. Phytoparasitica 2006, 34, 370–379. [Google Scholar] [CrossRef]
- Hajjar, M.J.; Ajlan, A.M.; Al-Ahmad, M.H. New approach of Beauveria bassiana to control the red palm weevil (Coleoptera: Curculionidae) by trapping technique. J. Econ. Entomol. 2015, 108, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; Ahmed, S.; Al-Jabr, A.M. Effect of Beauveria bassiana infection on the feeding performance and antioxidant defence of red palm weevil, Rhynchophorus ferrugineus. Biocontrol 2015, 60, 849–859. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; AlJabr, A.M. Susceptibility and immune defence mechanisms of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) against entomopathogenic fungal infections. Int. J. Mol. Sci. 2016, 17, 1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Verde, G.; Torta, L.; Mondello, V.; Caldarella, C.G.; Burruano, S.; Caleca, V. Pathogenicity bioassays of isolates of Beauveria bassiana on Rhynchophorus ferrugineus. Pest Manag. Sci. 2015, 71, 323–328. [Google Scholar] [CrossRef]
- Merghem, A. Susceptibility of the red palm weevil, Rhynchophorus ferrugineus (Olivier) to the green muscardine fungus, Metarhizium anisopliae (Metsch.) in the laboratory and in palm trees orchards. Egypt J. Biol. Pest Control 2011, 21, 179–183. [Google Scholar]
- Ricaño, J.; Güerri-Agulló, B.; Serna-Sarriás, M.J.; Rubio-Llorca, G.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Evaluation of the pathogenicity of multiple isolates of Beauveria bassiana (Hypocreales: Clavicipitaceae) on Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) for the assessment of a solid formulation under simulated field conditions. Fla. Entomol. 2013, 96, 1311–1324. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.D.; Yan, W.; Qin, W.Q.; Zhang, J.; Niu, X.Q.; Ma, G.C.; Li, F.H. Screening of tropical isolates of Metarhizium anisopliae for virulence to the red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae). SpringerPlus 2016, 5, 1100. [Google Scholar] [CrossRef] [Green Version]
- Yasin, M.; Wakil, W.; Ghazanfar, M.U.; Qayyum, M.A.; Tahir, M.; Bedford, G.O. Virulence of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against red palm weevil, Rhynchophorus ferrugineus (Olivier). Entomol. Res. 2019, 49, 3–12. [Google Scholar] [CrossRef] [Green Version]
- El Husseini, M.M. Efficacy of the fungus Beauveria bassiana (Balsamo) Vuillemin on the red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) larvae and adults under laboratory conditions. Egypt J. Biol. Pest Control 2019, 29, 58. [Google Scholar] [CrossRef]
- Abdel-Raheem, M.A.; Alghamdi, H.A.; Reyad, N.F. Virulence of fungal spores and silver nanoparticles from entomopathogenic fungi on the red palm weevil, Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae). Egypt J. Biol. Pest Control 2019, 29, 97. [Google Scholar] [CrossRef]
- Aldossary, A.A.; Shehata, S.T.; Hegazy, G.; Salem, M.A.; Faiza, M.A.M. Assessment of the entomopathogenic fungus Beauveria bassiana Saudi Arabian isolate (B-SA3) against the developmental stages of the red palm weevil, Rhynchophorus ferrugineus (Oliv.). Arab Univ. J. Agric. Sci. 2009, 17, 227–237. [Google Scholar]
- Hou, F.J.; Addis, S.N.K.; Azmi, W.A. Virulence evaluation of entomopathogenic fungi against the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Dryopthoridae). Malays. Appl. Biol. 2018, 47, 25–30. [Google Scholar]
- Qayyum, M.A.; Saleem, M.A.; Saeed, S.; Wakil, W.; Ishtiaq, M.; Ashraf, W.; Ahmed, N.; Ali, M.; Ikram, R.M.; Yasin, M.; et al. Integration of entomopathogenic fungi and eco-friendly insecticides for management of red palm weevil, Rhynchophorus ferrugineus (Olivier). Saudi J. Biol. Sci. 2020, 27, 1811–1817. [Google Scholar] [CrossRef]
- Abdel-Raheem, M.A.; Reyad, N.F.; Alghamdi, H.A. Virulence of nanoparticle preparation of entomopathogenic fungi and entomopathogenic bacteria against red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Rom. Biotechnol. Lett. 2020, 25, 1151–1159. [Google Scholar] [CrossRef]
- Ishak, I.; Ng, L.C.; Haris-Hussain, M.; Jalinas, J.; Idris, A.B.; Azlina, Z.; Samsudin, A.; Wahizatul, A.A. Pathogenicity of an indigenous strain of the entomopathogenic fungus Metarhizium anisopliae (Hypocreales: Clavicipitaceae) (MET-GRA4 Strain) as a potential biological control agent against the red palm weevil (Coleoptera: Dryophthoridae). J. Econ. Entomol. 2020, 113, 43–49. [Google Scholar] [CrossRef]
- Cheong, J.L.; Azmi, W.A. Dataset on the influence of relative humidity on the pathogenicity of Metarhizium anisopliae isolates from Thailand and Malaysia against red palm weevil (Rhynchophorus ferrugineus, Olivier) adult. Data Brief 2020, 30, 105482. [Google Scholar] [CrossRef]
- Al-Keridis, L.A.; Gaber, N.M.; Aldawood, A.S. Pathogenicity of Saudi Arabian fungal isolates against egg and larval stages of Rhynchophorus ferrugineus under laboratory conditions. Int. J. Trop. Insect Sci. 2020. [Google Scholar] [CrossRef]
- Batta, Y.A. Biocontrol of almond bark beetle (Scolytus amygdali Geurin-Meneville, Coleoptera: Scolytidae) using Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: Hyphomycetes). J. Appl. Microbiol. 2007, 103, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Kaaya, G.P.; Seshu-Reddy, K.V.; Kokwaro, E.D.; Munyinyi, D.M. Pathogenicity of Beauveria bassiana, Metarhizium anisopliae and Serratia marcescens to the banana weevil Cosmopolites sordidus. Biocontrol Sci. Technol. 1993, 3, 177–187. [Google Scholar] [CrossRef]
- Magara, E.; Nankinga, C.M.K.; Gold, C.S.; Kyamanywa, S.; Ragama, P.; Tushemereirwe, W.K.; Moore, D.; Gowen, S.R. Efficacy of Beauveria bassiana substrates and formulations for the control of banana weevil. Uganda J. Agric. Sci. 2004, 9, 900–905. [Google Scholar]
- Francardi, V.; Benvenuti, C.; Roversi, P.F.; Rumine, P.; Barzanti, G. Entomopathogenicity of Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metsch.) Sorokin isolated from different sources in the control of Rhynchophorus ferrugineus (Oliver) (Coleoptera Curculionidae). Redia 2012, 95, 49–55. [Google Scholar]
- Francardi, V.; Benvenuti, C.; Barzanti, G.P.; Roversi, P.F. Autocontamination trap with entomopathogenic fungi: A possible strategy in the control of Rhynchophorus ferrugineus (Olivier) (Coleoptera Curculionidae). Redia 2013, 96, 57–67. [Google Scholar]
- Francardi, V.; Benvenuti, C.; Barzanti, G.P.; Roversi, P.F. Metarhizium anisopliae biopesticides and fungus isolates: Control efficacy against Rhynchophorus ferrugineus (Olivier) (Coleoptera Dryophthoridae) on different contamination substrata. Redia 2015, 98, 25–29. [Google Scholar]
- Cito, A.; Mazza, G.; Strangi, A.; Benvenuti, C.; Barzanti, G.P.; Dreassi, E.; Turchetti, T.; Francardi, V.; Roversi, P.F. Characterization and comparison of Metarhizium strains isolated from Rhynchophorus ferrugineus. FEMS Microbiol. Lett. 2014, 355, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Peña, J.E.; Gilbin-Davis, R.M.; Duncan, R. Impact of indigenous Beauveria bassiana (Balsamo) Vuillemin on banana weevil and rotten sugarcane weevil (Coleoptera: Curculionidae) populations in banana in Florida. J. Agric. Entomol. 1995, 12, 163–167. [Google Scholar]
- Monzón, A.J.; Guharay, F.; Klingen, I. Natural occurrence of Beauveria bassiana in Hypothenemus hampei (Coleoptera: Curculionidae) populations in unsprayed coffee fields. J. Invertebr. Pathol. 2008, 97, 134–141. [Google Scholar] [CrossRef]
- Wraight, S.P.; Galaini-Wraight, S.; Howes, R.L.; Castrillo, L.A.; Carruthers, R.I.; Smith, R.H.; Matsumoto, T.K.; Keith, L.M. Prevalence of naturally-occurring strains of Beauveria bassiana in populations of coffee berry borer Hypothenemus hampei on Hawai’i Island, with observations on coffee plant-H. hampei-B. bassiana interactions. J. Invertebr. Pathol. 2018, 156, 54–72. [Google Scholar] [CrossRef]
- Asiry, K.A.; Sulieman, A.M.E.; Al-Anazi, N.A.; Veettil, V.N.; Abdelgadir, M.; Alkhregi, I. Isolation, phenotypic and genotypic characterization of indigenous Beauveria bassiana isolates from date palm infested with Rhynchophorus ferrugineus in Hail region, Saudi Arabia. Biosci. Biotechnol. Res. Commun. 2018, 11, 393–401. [Google Scholar] [CrossRef]
- Güerri-Agulló, B.; López-Follana, R.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Use of a solid formulation of Beauveria bassiana for biocontrol of the red palm weevil (Rhynchophorus ferrugineus) (Coleoptera: Dryophthoridae) under field conditions in SE Spain. Fla. Entomol. 2011, 94, 737–747. [Google Scholar] [CrossRef]
- Prior, C.; Arura, M. The infectivity of Metarhizium anisopliae to two insect pests of coconuts. J. Invertebr. Pathol. 1985, 45, 187–194. [Google Scholar] [CrossRef]
- Sabbahi, R.; Merzouki, A.; Guertin, C. Potential effect of Beauveria bassiana (Hypocreales: Clavicipitaceae) on Anthonomus signatus (Coleoptera: Curculionidae) in strawberries. Biocontrol Sci. Technol. 2009, 19, 729–741. [Google Scholar] [CrossRef]
- Schoeman, P.S.; Botha, H. Field management of the banana weevil, Cosmopolites sordidus (Coleoptera: Curculionidae), with Beauveria bassiana. Afr. Plant Prot. 2003, 9, 1–3. [Google Scholar]
- Hlerema, I.; Laurie, S.; Eiasu, B. Preliminary observations on use of Beauveria bassiana for the control of the sweetpotato weevil (Cylas sp.) in South Africa. Open Agric. 2017, 2, 595–599. [Google Scholar]
- de la Rosa, W.; Alatorre, R.; Barrera, J.F.; Toriello, C. Effect of Beauveria bassiana and Metarhizium anisopliae (Deuteromycetes) upon the coffee berry borer (Coleoptera: Scolytidae) under field conditions. J. Econ. Entomol. 2000, 93, 1409–1414. [Google Scholar] [CrossRef]
- Edgington, S.; Segura, H.; de la Rosa, W.; Williams, T. Photoprotection of Beauveria bassiana: Testing simple formulations for control of the coffee berry borer. Int. J. Pest Manag. 2000, 46, 169–176. [Google Scholar] [CrossRef]
- Greco, E.B.; Wright, M.G.; Burgueño, J.; Jaronski, S.T. Efficacy of Beauveria bassiana applications on coffee berry borer across an elevation gradient in Hawaii. Biocontrol Sci. Technol. 2018, 28, 995–1013. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Aristizábal, L.F.; Shriner, S.; Mascarin, G.M.; Moral, R.D.; Arthurs, S.P. Incorporating Beauveria bassiana into an integrated pest management plan for coffee berry borer in Hawaii. Front. Sustain. Food Syst. 2020, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- El-Sufty, R.; Al-Awash, S.A.; Al Bgham, S.; Shahdad, A.S.; Al Bathra, A.H. Pathogenicity of the fungus Beauveria bassiana (Bals.) Vuill to the red palm weevil, Rhynchophorus ferrugineus (Oliv.) (Col.: Curculionidae) under laboratory and field conditions. Egypt J. Biol. Pest Control 2009, 19, 81–85. [Google Scholar]
- Sewify, G.H.; Belal, M.H.; Al-Awash, S.A. Use of the entomopathogenic fungus, Beauveria bassiana for the biological control of the red palm weevil, Rhynchophorus ferrugineus Olivier. Egypt J. Biol. Pest Control 2009, 19, 157–163. [Google Scholar]
- Su, C.Y.; Tzean, S.S.; Ko, W.H. Beauveria bassiana as the lethal factor in a Taiwanese soil pernicious to sweetpotato weevil, Cylas formicarius. J. Invertebr. Pathol. 1988, 52, 195–197. [Google Scholar] [CrossRef]
- Nankinga, C.M.; Moore, D. Reduction of banana weevil populations using different formulations of the entomopathogenic fungus Beauveria bassiana. Biocontrol Sci. Technol. 2000, 10, 645–657. [Google Scholar] [CrossRef]
- Godonou, I.; Green, K.R.; Oduro, K.A.; Lomer, C.J.; Afreh-Nuamah, K. Field evaluation of selected formulations of Beauveria bassiana for the management of the banana weevil (Cosmopolites sordidus) on plantain (Musa spp., AAB group). Biocontrol Sci. Technol. 2000, 10, 779–788. [Google Scholar] [CrossRef]
- Bustillo, A.E.; Bernal, M.G.; Benavides, P.; Chaves, B. Dynamics of Beauveria bassiana and Metarhizium anisopliae infecting Hypothenemus hampei (Coleoptera: Scolytidae) populations emerging from fallen coffee berries. Fla. Entomol. 1999, 82, 491–498. [Google Scholar] [CrossRef]
- Malik, M.A.; Ahmad, S.J.N.; Ahmad, J.N.; Abbasi, A.; Sufyan, M.; Arif, M.J. Efficacy of Bacillus thuringiensis and Beauveria bassiana against red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae). Afr. Entomol. 2019, 27, 386–394. [Google Scholar] [CrossRef]
- Saleh, M.M.E.; Hegazy, G.; Salem, M.; Hanounik, S.B.; Al Mohanna, O.; Alheji, M.A. Persistence of Steinernema carpocapsae (Nematoda: Steinernematidae) and Beauveria bassiana (Deuteromycotina: Hyphomycetes) in soil around date palm trunks and their effect on adults of Rhynchophorus ferrugineus. Egypt J. Biol. Pest Control 2004, 14, 141–145. [Google Scholar]
- Wakil, W.; Yasin, M.; Shapiro-Ilan, D.I. Effects of single and combined applications of entomopathogenic fungi and nematodes against Rhynchophorus ferrugineus (Olivier). Sci. Rep. 2017, 7, 5971. [Google Scholar] [CrossRef]
- Tinzaara, W.; Gold, C.S.; Dicke, M.; Van Huis, A.; Nankinga, C.M.; Kagezi, G.H.; Ragama, P.E. The use of aggregation pheromone to enhance dissemination of Beauveria bassiana for the control of the banana weevil in Uganda. Biocontrol Sci. Technol. 2007, 17, 111–124. [Google Scholar] [CrossRef]
- Lopes, R.B.; Laumann, R.A.; Moore, D.; Oliveira, M.W.M.; Faria, M. Combination of the fungus Beauveria bassiana and pheromone in an attract-and-kill strategy against the banana weevil, Cosmopolites sordidus. Entomol. Exp. Appl. 2014, 151, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Tinzaara, W.; Emudong, P.; Nankinga, C.; Tushemereirwe, W.; Kagezi, G.; Gold, C.S.; Dicke, M.; Van Huis, A.; Karamura, E. Enhancing dissemination of Beauveria bassiana with host plant base incision trap for the management of the banana weevil Cosmopolites sordidus. Afr. J. Agric. Res. 2015, 10, 3878–3884. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K. Auto-infection system for the sweetpotato weevil, Cylas formicarius (Fabricius) (Coleoptera: Curculionidae) with entomopathogenic fungi, Beauveria bassiana using a modified sex pheromone trap in the field. Appl. Entomol. Zool. 1999, 34, 501–505. [Google Scholar] [CrossRef] [Green Version]
- El-Sufty, R.; Al Bgham, S.; Al-Awash, S.; Shahdad, A.; Al Bathra, A. A Trap for auto-dissemination of the entomopathogenic fungus Beauveria bassiana by red palm weevil adults in date palm plantations. Egypt J. Biol. Pest Control 2011, 21, 271–276. [Google Scholar]
- Sewify, G.H.; Belal, M.H.; Saeed, M.Q. Using pheromone mass-trapping and the entomopathogenic fungus Beauveria bassiana in IPM programs for controlling the red palm weevil, Rhynchophorus ferrugineus Olivier (Coleoptera: Rhynchophoridae). Egypt J. Biol. Pest Control 2014, 24, 197–202. [Google Scholar]
- Dembilio, Ó.; Moya, P.; Vacas, S.; Ortega-Garcia, L.; Quesada-Moraga, E.; Jaques, J.A.; Navarro-Llopis, V. Development of an attract-and-infect system to control Rhynchophorus ferrugineus with the entomopathogenic fungus Beauveria bassiana. Pest. Manag. Sci. 2018, 74, 1861–1869. [Google Scholar] [CrossRef]
- El-Sufty, R.; Al Bgham, S.; Al-Awash, S.; Shahdad, A.; Al Bathra, A. A study on a trap for autodissemination of the entomopathogenic fungus Beauveria bassiana by red palm weevil adults in date palm plantations. J. Basic. Appl. Mycol. 2010, 1, 61–65. [Google Scholar]
- Landolt, P.J. Sex attractant and aggregation pheromones of male phytophagous insects. Am. Entomol. 1997, 43, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Uemura-Lima, D.H.; Ventura, M.U.; Mikami, A.Y.; da Silva, F.C.; Morales, L. Responses of coffee berry borer, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae), to vertical distribution of methanol:ethanol traps. Neotrop. Entomol. 2010, 39, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.E.; Vilela, E.F.; Tinoco, R.S.; de Lima, J.O.G.; Fantine, A.K.; Morais, E.G.F.; França, C.F.M. Correlation between numbers captured and infestation levels of the coffee berry-borer, Hypothenemus hampei: A preliminary basis for an action threshold using baited traps. Int. J. Pest Manag. 2012, 58, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Sewify, G.H.; Belal, M.H.; Qaed, M.S. Food-baited aggregation pheromone traps for management of the red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae). Egypt J. Biol. Pest Control 2014, 24, 431–436. [Google Scholar]
- Vacas, S.; Abad-Payá, M.; Primo, J.; Navarro-Llopis, V. Identification of pheromone synergists for Rhynchophorus ferrugineus trapping systems from Phoenix canariensis palm volatiles. J. Agric. Food Chem. 2014, 62, 6053–6064. [Google Scholar] [CrossRef]
- Dotaona, R.; Wilson, B.A.L.; Stevens, M.M.; Holloway, J.; Ash, G.J. Chronic effects and horizontal transmission of Metarhizium anisopliae strain QS155 infection in the sweetpotato weevil, Cylas formicarius (Coleoptera: Brentidae). Biol. Control 2017, 114, 24–29. [Google Scholar] [CrossRef]
- Schoeman, P.S.; Schoeman, M.H. Transmission of Beauveria bassiana from infected to uninfected adults of the banana weevil Cosmopolites sordidus (Coleoptera: Curculionidae). Afr. Plant Prot. 1999, 5, 53–54. [Google Scholar]
- Llácer, E.; Santiago-Álvarez, C.; Jacas, J.A. Could sterile males be used to vector a microbiological control agent? The case of Rhynchophorus ferrugineus and Beauveria bassiana. Bull. Entomol. Res. 2013, 103, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Bojke, A.; Tkaczuk, C.; Stepnowski, P.; Golebiowski, M. Comparison of volatile compounds released by entomopathogenic fungi. Microbiol. Res. 2018, 214, 129–136. [Google Scholar] [CrossRef]
- Herrera, J.M.; Pizzolitto, R.P.; Zunino, M.P.; Dambolena, J.S.; Zygadlo, J.A. Effect of fungal volatile organic compounds on a fungus and an insect that damage stored maize. J. Stored Prod. Res. 2015, 62, 74–80. [Google Scholar] [CrossRef]
- Dotaona, R.; Wilson, B.A.L.; Ash, G.J.; Holloway, J.; Stevens, M.M. Sweetpotato weevil, Cylas formicarius (Fab.) (Coleoptera: Brentidae) avoids its host plant when a virulent Metarhizium anisopliae isolate is present. J. Invertebr. Pathol. 2017, 148, 67–72. [Google Scholar] [CrossRef]
- Leng, P.H.; Reddy, G.V.P. Bioactivity of selected eco-friendly pesticides against Cylas formicarius (Coleoptera: Brentidae). Fla. Entomol. 2012, 95, 1040–1047. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Effect of endophytic Beauveria bassiana on populations of the banana weevil, Cosmopolites sordidus, and their damage in tissue-cultured banana plants. Entomol. Exp. Appl. 2008, 129, 157–165. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Endophytic Beauveria bassiana in banana (Musa spp.) reduces banana weevil (Cosmopolites sordidus) fitness and damage. Crop Prot. 2008, 27, 1437–1441. [Google Scholar] [CrossRef]
- Arab, Y.A.; El-Deeb, H.M. The use of endophyte Beauveria bassiana for bio-protection of date palm seedlings against red palm weevil and rhizoctonia root-rot disease. Sci. J. King Faisal Univ. 2012, 13, 91–101. [Google Scholar]
- Villacarlos, L.T.; Granados-Polo, M.F.U. Potential of Metarhizium anisopliae for the control of the sweetpotato weevil, Cylas formicarius (F.) (Curculionidae: Coleoptera). Philipp. J. Crop Sci. 1989, 14, 109–114. [Google Scholar]
- El Kichaoui, A.Y.; Abu Asaker, B.A.; El-Hindi, M.W. Isolation, molecular identification and under lab evaluation of the entomopathogenic fungi M. anisopliae and B. bassiana against the red palm weevil R. ferrugineus in Gaza Strip. Adv. Microbiol. 2017, 7, 109–124. [Google Scholar] [CrossRef] [Green Version]
- McPhie, D.; Burrack, H.J. Effects of microbial, organically acceptable, and reduced risk insecticides on Anthonomus signatus (Curculionidae: Coleoptera) in strawberries (Fragaria x ananassa). Crop Prot. 2016, 89, 255–258. [Google Scholar] [CrossRef]
- Irulandi, S.; Aiyanathan, K.E.A.; Bhuvaneswari, S.S.B. Assessment of biopesticides and insecticide against pseudostem weevil Odoiporus longicollis Oliver in red banana. J. Biopestic. 2012, 5, 68–71. [Google Scholar]
- Reddy, G.V.P.; Zhao, Z.H.; Humber, R.A. Laboratory and field efficacy of entomopathogenic fungi for the management of the sweetpotato weevil, Cylas formicarius (Coleoptera: Brentidae). J. Invertebr. Pathol. 2014, 122, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.A.; Qayyum, M.A.; Ali, M.; Amin, M.; Tayyab, M.; Maqsood, S. Effect of sub-lethal doses of Beauveria bassiana and nitenpyram on the development of red palm weevil, Rhynchophorus ferrugineus (Olivier). Pak. J. Zool. 2019, 51, 559–565. [Google Scholar] [CrossRef]
- Malik, M.A.; Manzoor, M.; Ali, H.; Muhammad, A.; ul Islam, S.; Qasim, M.; Ahmad, N.; Idrees, A.; Muhammad, A.; Saqib, H.S.A. Evaluation of imidacloprid and entomopathogenic fungi, Beauveria bassiana against the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae). J. Entomol. Zool. Stud. 2016, 4, 262–268. [Google Scholar]
- Alston, D.G.; Rangel, D.E.N.; Lacey, L.A.; Golez, H.G.; Kim, J.J.; Roberts, D.W. Evaluation of novel fungal and nematode isolates for control of Conotrachelus nenuphar (Coleoptera: Curculionidae) larvae. Biol. Control 2005, 35, 163–171. [Google Scholar] [CrossRef]
- Harrison, R.D.; Gardner, W.A.; Kinard, D.J. Relative susceptibility of pecan weevil fourth instars and adults to selected isolates of Beauveria bassiana. Biol. Control 1993, 3, 34–38. [Google Scholar] [CrossRef]
- Cheng, Y.Q.; Liu, T.; Zhao, Y.X.; Geng, W.T.; Chen, L.T.; Liu, J.F. Evaluation of pathogenicity of the fungi Metarhizium anisopliae and Beauveria bassiana in hazelnut weevil (Curculio nucum L., Coleoptera, Curculionidae) larvae. Indian J. Microbiol. 2016, 56, 405–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihara, F.; Toyama, M.; Sato, T. Pathogenicity of Metarhizium anisopliae to the chestnut weevil larvae under laboratory and field conditions. Appl. Entomol. Zool. 2003, 38, 461–465. [Google Scholar] [CrossRef]
- Ihara, F.; Toyama, M.; Higaki, M.; Mishwo, K.; Yaginuma, K. Comparison of pathogenicities of Beauveria bassiana and Metarhizium anisopliae to chestnut pests. Appl. Entomol. Zool. 2009, 44, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Tedders, W.L.; Weaver, D.J.; Wehunt, E.J. Pecan weevil: Suppression of larvae with the fungi Metarhizium anisopliae and Beauveria bassiana and the nematode Neoaplectana dutkyi. J. Econ. Entomol. 1973, 66, 723–725. [Google Scholar] [CrossRef]
- Tedders, W.L.; Weaver, D.J.; Wehunt, E.J.; Gentry, C.R. Bioassay of Metarhizium anisopliae, Beauveria bassiana, and Neoaplectana carpocapsae against larvae of the plum curculio, Conotrachelus nenuphar (Herbst) (Coleoptera: Curculionidae). Environ. Entomol. 1982, 11, 901–904. [Google Scholar] [CrossRef]
- Batalla-Carrera, L.; Morton, A.; Santamaria, S.; Garcia-del-Pino, F. Isolation and virulence of entomopathogenic fungi against larvae of hazelnut weevil Curculio nucum (Coleoptera, Curculionidae) and the effects of combining Metarhizium anisopliae with entomopathogenic nematodes in the laboratory. Biocontrol Sci. Technol. 2013, 23, 101–125. [Google Scholar] [CrossRef]
- Champlin, F.R.; Cheung, P.Y.K.; Pekrul, S.; Smith, R.J.; Burton, R.L.; Grula, E.A. Virulence of Beauveria bassiana mutants for the pecan weevil. J. Econ. Entomol. 1981, 74, 617–621. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Tedders, W.L. Colonization, transmission, and longevity of Beauveria bassiana and Metarhizium anisopliae (Deuteromycotina: Hypomycetes) on pecan weevil larvae (Coleoptera: Curculionidae) in the soil. Environ. Entomol. 1984, 13, 557–560. [Google Scholar] [CrossRef]
- Torrini, G.; Benvenuti, C.; Binazzi, F.; Marianelli, L.; Paoli, F.; Peverieri, G.S.; Roversi, P.F. Entomopathogenic fungi and nematodes against larvae of the chestnut weevil, Curculio elephas (Coleoptera: Curculionidae): A laboratory evaluation. Int. J. Pest Manag. 2018, 64, 287–293. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Fuxa, J.R.; Wood, B.W.; Nguyen, K.B.; Adams, B.J.; Humber, R.A.; Hall, M.J. Survey of entomopathogenic nematodes and fungi endemic to pecan orchards of the Southeastern United States and their virulence to the pecan weevil (Coleoptera: Curculionidae). Environ. Entomol. 2003, 32, 187–195. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Brown, I. Earthworms as phoretic hosts for Steinernema carpocapsae and Beauveria bassiana: Implications for enhanced biological control. Biol. Control 2013, 66, 41–48. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Tedders, W.L. Suppression of pecan weevil (Coleoptera: Curculionidae) populations with entomopathogenic fungi. Environ. Entomol. 1983, 12, 471–474. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Gardner, W.A.; Leland, J.; Behles, R.W. Laboratory mortality and mycosis of adult Curculio caryae (Coleoptera: Curculionidae) following application of Metarhizium anisopliae in the laboratory or field. J. Entomol. Sci. 2009, 44, 24–36. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Wells, L.; Wood, B.W. Cumulative impact of a clover cover crop on the persistence and efficacy of Beauveria bassiana in suppressing the pecan weevil (Coleoptera: Curculionidae). Environ. Entomol. 2012, 41, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Sarraquigne, J.P.; Couturié, E.; Fernandez, M.M. Integrated control of hazelnut weevil (Curculio nucum): An evaluation of entomopathogenic nematodes and parasitic fungi. Acta Hortic. 2009, 845, 555–560. [Google Scholar] [CrossRef]
- Pereault, R.J.; Whalon, M.E.; Alston, D.G. Field efficacy of entomopathogenic fungi and nematodes targeting caged last-instar plum curculio (Coleoptera: Curculionidae) in Michigan cherry and apple orchards. Environ. Entomol. 2009, 38, 1126–1134. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Gardner, W.A. Trunk perimeter applications of Beauveria bassiana to suppress adult Curculio caryae (Coleoptera: Curculionidae). J. Entomol. Sci. 2004, 39, 337–349. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Mizell, R.F. An insect pupal cell with antimicrobial properties that suppress an entomopathogenic fungus. J. Invertebr. Pathol. 2015, 124, 114–116. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Cottrell, T.E.; Behle, R.W.; Wood, B.W. Comparison of application methods for suppressing the pecan weevil (Coleoptera: Curculionidae) with Beauveria bassiana under field conditions. Environ. Entomol. 2008, 37, 162–171. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Gardner, W.A.; Behle, R.W.; Ree, B.; Harris, M.K. Efficacy of entomopathogenic fungi in suppressing pecan weevil, Curculio caryae (Coleoptera: Curculionidae), in commercial pecan orchards. Southwest Entomol. 2009, 34, 111–120. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Wells, L.; Cottrell, T.E.; Behle, R.W.; Wood, B.W. Effects of entomopathogenic fungus species, and impact of fertilizers, on biological control of pecan weevil (Coleoptera: Curculionidae). Environ. Entomol. 2013, 42, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Jackson, M.; Reilly, C.C.; Hotchkiss, M.W. Effects of combining an entomopathogenic fungi or bacterium with entomopathogenic nematodes on mortality of Curculio caryae (Coleoptera: Curculionidae). Biol. Control 2004, 30, 119–126. [Google Scholar] [CrossRef]
- Asan, C.; Hazir, S.; Cimen, H.; Ulug, D.; Taylor, J.; Butt, T.; Karagoz, M. An innovative strategy for control of the chestnut weevil Curculio elephas (Coleoptera: Curculionidae) using Metarhizium brunneum. Crop Prot. 2017, 102, 147–153. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Bock, C.; Mai, K.; Boykin, D.; Wells, L.; Hudson, W.G.; Mizell, R.F. Control of pecan weevil with microbial biopesticides. Environ. Entomol. 2017, 46, 1299–1304. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Wood, B.W. Effects of combining microbial and chemical insecticides on mortality of the pecan weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2011, 104, 14–20. [Google Scholar] [CrossRef]
- Brito, E.S.; de Paula, A.R.; Vieira, L.P.; Dolinski, C.; Samuels, R.I. Combining vegetable oil and sub-lethal concentrations of imidacloprid with Beauveria bassiana and Metarhizium anisopliae against adult guava weevil Conotrachelus psidii (Coleoptera: Curculionidae). Biocontrol Sci. Technol. 2008, 18, 665–673. [Google Scholar] [CrossRef]
- Sepulveda, M.; Vargas, M.; Gerding, M.; Ceballos, R.; Oyarzua, P. Molecular, morphological and pathogenic characterization of six strains of Metarhizium spp. (Deuteromycotina: Hyphomycetes) for the control of Aegorhinus superciliosus (Coleoptera: Curculionidae). Chil. J. Agric. Res. 2016, 76, 77–83. [Google Scholar] [CrossRef]
- McCoy, C.W.; Boucias, D.G. Selection of Beauveria bassiana pathotypes as potential microbial control agents of soil-inhabiting citrus weevils. Mem. Inst. Oswaldo Cruz 1989, 84, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. Laboratory selection of Metarhizium spp. isolates for control of vine weevil larvae (Otiorhynchus sulcatus). J. Invertebr. Pathol. 1993, 62, 15–21. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. The influence of temperature on the susceptibility of vine weevil, Otiorhynchus sulcatus (Fabricius) (Coleoptera: Curculionidae), larvae to Metarhizium anisopliae (Deuteromycotina: Hyphomycetes). Ann. Appl. Biol. 1994, 124, 185–193. [Google Scholar] [CrossRef]
- Bruck, D.J. Natural occurrence of entomopathogens in pacific northwest nursery soils and their virulence to the black vine weevil, Otiorhynchus sulcatus (F.) (Coleoptera: Curculionidae). Environ. Entomol. 2004, 33, 1335–1343. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, J.; Reineke, A. Efficiency of commercial entomopathogenic fungal species against different members of the genus Otiorhynchus (Coleoptera: Curculionidae) under laboratory and semi-field conditions. J. Plant Dis. Prot. 2014, 121, 211–218. [Google Scholar] [CrossRef]
- Klingen, I.; Westrum, K.; Meyling, N.V. Effect of Norwegian entomopathogenic fungal isolates against Otiorhynchus sulcatus larvae at low temperatures and persistence in strawberry rhizospheres. Biol. Control 2015, 81, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pope, T.W.; Hough, G.; Arbona, C.; Roberts, H.; Bennison, J.; Buxton, J.; Prince, G.; Chandler, D. Investigating the potential of an autodissemination system for managing populations of vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae) with entomopathogenic fungi. J. Invertebr. Pathol. 2018, 154, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morera-Margarit, P.; Karley, A.J.; Mitchell, C.; Graham, R.I.; Pope, T.W. Geographic origin may not influence vine weevil Otiorhynchus sulcatus (Fabricius) susceptibility to the entomopathogenic fungus Metarhizium brunneum (Petch). Biocontrol Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Marchal, M.; Robert, P.H. Comparative susceptibility of Otiorhynchus sulcatus and Sitona lineatus (Coleoptera: Curculionidae) early stages to five entomopathogenic Hyphomycetes. Environ. Entomol. 1985, 14, 247–253. [Google Scholar] [CrossRef]
- Soares Jr, G.G.; Marchal, M.; Ferron, P. Susceptibility of Otiorhynchus sulcatus (Coleoptera: Curculionidae) larvae to Metarhizium anisopliae and Metarhizium flavoviride (Deuteromycotina: Hyphomycetes) at two different temperatures. Environ. Entomol. 1983, 12, 1887–1891. [Google Scholar] [CrossRef]
- Dlamini, B.E.; Malan, A.P.; Addison, P. Control of the banded fruit weevil, Phlyctinus callosus (Schonherr) (Coleoptera: Curculionidae), using entomopathogenic fungi. Afr. Entomol. 2020, 28, 106–114. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. Effect of potting media on the control of Otiorhynchus sulcatus larvae on outdoor strawberry plants using the entomogenous fungus Metarhizium anisopliae. Biol. Control 1992, 2, 238–243. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Easterbrook, M.A.; Gillespie, A.T.; Charnley, A.K. Control of Otiorhynchus sulcatus (Fabricius) (Coleoptera: Curculionidae) larvae on a range of hardy ornamental nursery stock species using the entomogenous fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 1993, 3, 63–72. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. Application of Metarhizium anisopliae (Metsch.) Sor. conidia to control Otiorhynchus sulcatus (F) (Coleoptera: Curculionidae) larvae on glasshouse pot plants. Ann. Appl. Biol. 1993, 122, 623–636. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. The development of Otiorhynchus sulcatus (Fabricius) (Coleoptera: Curculionidae) larvae on a range of ornamental pot-plant species and the potential for control using Metarhizium anisopliae. J. Hortic. Sci. 1993, 68, 627–635. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Charnley, A.K. Selection of virulent and persistent Metarhizium anisopliae isolates to control black vine weevil (Otiorhynchus sulcatus) larvae on glasshouse Begonia. J. Invertebr. Pathol. 1993, 62, 47–52. [Google Scholar] [CrossRef]
- Bruck, D.J. Ecology of Metarhizium anisopliae in soilless potting media and the rhizosphere: Implications for pest management. Biol. Control 2005, 32, 155–163. [Google Scholar] [CrossRef]
- Bruck, D.J.; Donahue, K.M. Persistence of Metarhizium anisopliae incorporated into soilless potting media for control of the black vine weevil, Otiorhynchus sulcatus in container-grown ornamentals. J. Invertebr. Pathol. 2007, 95, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.; Dobrovin-Pennington, A.; Fitzgerald, J.; Dew, K.; Wilson, C.; Ross, K.; Perkins, C. Improving biocontrol of black vine weevil (Otiorhynchus sulcatus) with entomopathogenic fungi in growing media by incorporating spent mushroom compost. Biocontrol 2018, 63, 697–706. [Google Scholar] [CrossRef]
- Cross, J.V.; Burgess, C.M. Localised insecticide treatment for the control of vine weevil larvae (Otiorhynchus sulcatus) on field-grown strawberry. Crop Prot. 1997, 16, 565–574. [Google Scholar] [CrossRef]
- Oddsdottir, E.S.; Eilenberg, J.; Sen, R.; Halldorsson, G. The effects of insect pathogenic soil fungi and ectomycorrhizal inoculation of birch seedlings on the survival of Otiorhynchus larvae. Agric. For. Entomol. 2010, 12, 319–324. [Google Scholar] [CrossRef]
- Vainio, A.; Hokkanen, H.M.T. The potential of entomopathogenic fungi and nematodes against Otiorhynchus ovatus L. and O. dubius Ström (Col., Curculionidae) in the field. J. Appl. Entomol. 1993, 115, 379–387. [Google Scholar] [CrossRef]
- Bruck, D.J. Effect of potting media components on the infectivity of Metarhizium anisopliae against the black vine weevil (Coleoptera: Curculionidae). J. Environ. Hortic. 2006, 24, 91–94. [Google Scholar] [CrossRef]
- Bruck, D.J. Efficacy of Metarhizium anisopliae as a curative application for black vine weevil (Otiorhynchus sulcatus) infesting container-grown nursery crops. J. Environ. Hortic. 2007, 25, 150–156. [Google Scholar] [CrossRef]
- Booth, S.R.; Shanks Jr, C.H. Potential of a dried rice/mycelium formulation of entomopathogenic fungi to suppress subterranean pests in small fruits. Biocontrol Sci. Technol. 1998, 8, 197–206. [Google Scholar] [CrossRef]
- Booth, S.R.; Tanigoshi, L.; Dewes, I. Potential of a dried mycelium formulation of an indigenous strain of Metarhizium anisopliae against subterranean pests of cranberry. Biocontrol Sci. Technol. 2000, 10, 659–668. [Google Scholar] [CrossRef]
- Easter-brook, M.A.; Cantwell, M.P.; Chandler, D. Control of the black vine weevil, Otiorhynchus sulcatus, with the fungus Metarhizium anisopliae. Phytoparasitica 1992, 20, S17–S19. [Google Scholar] [CrossRef]
- Roberts, J.M.; Jahir, A.; Graham, J.; Pope, T.W. Catch me if you can: The influence of refuge/trap design, previous feeding experience, and semiochemical lures on vine weevil (Coleoptera: Curculionidae) monitoring success. Pest Manag. Sci. 2019, 76, 553–560. [Google Scholar] [CrossRef]
- van Tol, R.W.H.M.; Elberse, I.A.M.; Bruck, D.J. Development of a refuge-kairomone device for monitoring and control of the vine weevil, Otiorhynchus sulcatus, by lure-and-kill and lure-and-infect. Crop Prot. 2020, 129, 105045. [Google Scholar] [CrossRef]
- Rondot, Y.; Reineke, A. Association of Beauveria bassiana with grapevine plants deters adult black vine weevils, Otiorhynchus sulcatus. Biocontrol Sci. Technol. 2017, 27, 811–820. [Google Scholar] [CrossRef]
- Kepler, R.M.; Bruck, D.J. Examination of the interaction between the black vine weevil (Coleoptera: Curculionidae) and an entomopathogenic fungus reveals a new tritrophic interaction. Environ. Entomol. 2006, 35, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.A.; Shah, F.A.; Butt, T.M. Combined use of entomopathogenic nematodes and Metarhizium anisopliae as a new approach for black vine weevil, Otiorhynchus sulcatus, control. Entomol. Exp. Appl. 2008, 129, 340–347. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Butt, T.M. The entomopathogenic nematode Steinernema kraussei and Metarhizium anisopliae work synergistically in controlling overwintering larvae of the black vine weevil, Otiorhynchus sulcatus, in strawberry growbags. Biocontrol Sci. Technol. 2010, 20, 99–105. [Google Scholar] [CrossRef]
- Ansari, M.A.; Butt, T.M. Influence of the application methods and doses on the susceptibility of black vine weevil larvae Otiorhynchus sulcatus to Metarhizium anisopliae in field-grown strawberries. Biocontrol 2013, 58, 257–267. [Google Scholar] [CrossRef]
- Quintela, E.D.; McCoy, C.W. Pathogenicity enhancement of Metarhizium anisopliae and Beauveria bassiana to first instars of Diaprepes abbreviatus (Coleoptera: Curculionidae) with sublethal doses of imidacloprid. Environ. Entomol. 1997, 26, 1173–1182. [Google Scholar] [CrossRef]
- Quintela, E.D.; McCoy, C.W. Conidial attachment of Metarhizium anisopliae and Beauveria bassiana to the larval cuticle of Diaprepes abbreviatus (Coleoptera: Curculionidae) treated with imidacloprid. J. Invertebr. Pathol. 1998, 72, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintela, E.D.; McCoy, C.W. Synergistic effect of imidacloprid and two entomopathogenic fungi on the behavior and survival of larvae of Diaprepes abbreviatus (Coleoptera: Curculionidae) in soil. J. Econ. Entomol. 1998, 91, 110–122. [Google Scholar] [CrossRef]
- Shah, F.A.; Ansari, M.A.; Prasad, M.; Butt, T.M. Evaluation of black vine weevil (Otiorhynchus sulcatus) control strategies using Metarhizium anisopliae with sublethal doses of insecticides in disparate horticultural growing media. Biol. Control 2007, 40, 246–252. [Google Scholar] [CrossRef]
- Shah, F.A.; Gaffney, M.; Ansari, M.A.; Prasad, M.; Butt, T.M. Neem seed cake enhances the efficacy of the insect pathogenic fungus Metarhizium anisopliae for the control of black vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae). Biol. Control 2008, 44, 111–115. [Google Scholar] [CrossRef]
- Gillett-Kaufman, J.L.; Kimbrough, J.W. A modified method to visualize infection sites of spores of the entomopathogen Beauveria bassiana (Deuteromycotina: Hyphomycetes) on the exoskeleton of citrus root weevil Diaprepes abbreviatus (Coleoptera: Curculionidae) adults. Fla. Entomol. 2009, 92, 623–628. [Google Scholar]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Wang, L.D. Fungal endophytes: Beyond herbivore management. Front. Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef] [Green Version]
- Ramakuwela, T.; Hatting, J.; Bock, C.; Vega, F.E.; Wells, L.; Mbata, G.N.; Shapiro-Ilan, D.I. Establishment of Beauveria bassiana as a fungal endophyte in pecan (Carya illinoinensis) seedlings and its virulence against pecan insect pests. Biol. Control 2020, 140, 104102. [Google Scholar] [CrossRef]
- Posada, F.; Vega, F.E. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedlings (Theobroma cacao). Mycologia 2005, 97, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Vidal, S.; Lopez-Llorca, L.V.; Jansson, H.B.; Salinas, J. Endophytic colonization of date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. Micron 2006, 37, 624–632. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Qasim, M.; Aguila, L.C.R.; Wang, F.F.; Keppanan, R.; Wang, L.D. Endophytic Beauveria bassiana in foliar-treated Citrus limon plants acting as a growth suppressor to three successive generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects 2019, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Bing, L.A.; Lewis, L.C. Occurrence of the entomopathogen Beauveria bassiana (Balsamo) Vuillemin in different tillage regimes and in Zea mays L and virulence towards Ostrinia nubilalis (Hubner). Agric. Ecosyst. Environ. 1993, 45, 147–156. [Google Scholar] [CrossRef]
- Oliveira, I.; Pereira, J.A.; Quesada-Moraga, E.; Lino-Neto, T.; Bento, A.; Baptista, P. Effect of soil tillage on natural occurrence of fungal entomopathogens associated to Prays oleae Bern. Sci. Hortic. 2013, 159, 190–196. [Google Scholar] [CrossRef]
- Milner, R.J.; Samson, P.; Morton, R. Persistence of conidia of Metarhizium anisopliae in sugarcane fields: Effect of isolate and formulation on persistence over 3.5 years. Biocontrol Sci. Technol. 2003, 13, 507–516. [Google Scholar] [CrossRef]
- Mayerhofer, J.; Enkerli, J.; Zelger, R.; Strasser, H. Biological control of the European cockchafer: Persistence of Beauveria brongniartii after long-term applications in the Euroregion Tyrol. Biocontrol 2015, 60, 617–629. [Google Scholar] [CrossRef]
- Swiergiel, W.; Meyling, N.V.; Porcel, M.; Rämert, B. Soil application of Beauveria bassiana GHA against apple sawfly, Hoplocampa testudinea (Hymenoptera: Tenthredinidae): Field mortality and fungal persistence. Insect Sci. 2016, 23, 854–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombes, C.A.; Hill, M.P.; Moore, S.D.; Dames, J.F. Entomopathogenic fungi as control agents of Thaumatotibia leucotreta in citrus orchards: Field efficacy and persistence. Biocontrol 2016, 61, 729–739. [Google Scholar] [CrossRef]
- Baxter, I.H.; Howard, N.; Armsworth, C.G.; Barton, L.E.E.; Jackson, C. The potential of two electrostatic powders as the basis for an auto dissemination control method of Plodia interpunctella (Hubner). J. Stored Prod. Res. 2008, 44, 152–161. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Vassilakos, T.N.; Dutton, A.C.; Jessop, N.; Sherwood, D.; Pease, G.; Brglez, A.; Storm, C.; Trdan, S. Combining electrostatic powder with an insecticide: Effect on stored-product beetles and on the commodity. Pest Manag. Sci. 2016, 72, 2208–2217. [Google Scholar] [CrossRef] [PubMed]
- Athanassiou, C.G.; Rumbos, C.I.; Sakka, M.; Potin, O.; Storm, C.; Dillon, A.B. Delivering Beauveria bassiana with electrostatic powder for the control of stored-product beetles. Pest Manag. Sci. 2017, 73, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Meikle, W.G.; Mercadier, G.; Holst, N.; Nansen, C.; Girod, V. Duration and spread of an entomopathogenic fungus, Beauveria bassiana (Deuteromycota: Hyphomycetes), used to treat varroa mites (Acari: Varroidae) in honey bee (Hymenoptera: Apidae) hives. J. Econ. Entomol. 2007, 100, 1–10. [Google Scholar] [CrossRef]
- Andriessen, R.; Snetselaar, J.; Suer, R.A.; Osinga, A.J.; Deschietere, J.; Lyimo, I.N.; Mnyone, L.L.; Brooke, B.D.; Ranson, H.; Knols, B.G.J.; et al. Electrostatic coating enhances bioavailability of insecticides and breaks pyrethroid resistance in mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 12081–12086. [Google Scholar] [CrossRef] [Green Version]
- Mazza, G.; Inghilesi, A.F.; Stasolla, G.; Cini, A.; Cervo, R.; Benvenuti, C.; Francardi, V.; Cristofaro, M.; Arnone, S.; Roversi, P.F. Sterile Rhynchophorus ferrugineus males efficiently impair reproduction while maintaining their sexual competitiveness in a social context. J. Pest Sci. 2016, 89, 459–468. [Google Scholar] [CrossRef]
- Sookar, P.; Alleck, M.; Ahseek, N.; Bhagwant, S. Sterile male peach fruit flies, Bactrocera zonata (Saunders) (Diptera: Tephritidae), as a potential vector of the entomopathogen Beauveria bassiana (Balsamo) Vuillemin in a SIT programme. Afr. Entomol. 2014, 22, 488–498. [Google Scholar] [CrossRef]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. Biocontrol 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Hedimbi, M.; Kaaya, G.P.; Singh, S.; Chimwamurombe, P.M.; Gindin, G.; Glazer, I.; Samish, M. Protection of Metarhizium anisopliae conidia from ultra-violet radiation and their pathogenicity to Rhipicephalus evertsi evertsi ticks. Exp. Appl. Acarol. 2008, 46, 149–156. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.G.P.; Lopes, R.B.; Rezende, J.M.; Delalibera, I. Increased tolerance of Beauveria bassiana and Metarhizium anisopliae conidia to high temperature provided by oil-based formulations. J. Invertebr. Pathol. 2018, 151, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Rangel, D.E.N.; Braga, G.U.L.; Anderson, A.J.; Roberts, D.W. Variability in conidial thermotolerance of Metarhizium anisopliae isolates from different geographic origins. J. Invertebr. Pathol. 2005, 88, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, È.K.K.; Rangel, D.E.N.; Moraes, Á.M.L.; Bittencourt, V.R.E.P.; Roberts, D.W. Variability in tolerance to UV-B radiation among Beauveria spp. isolates. J. Invertebr. Pathol. 2007, 96, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Rangel, D.E.N.; Braga, G.U.L.; Fernandes, È.K.K.; Keyser, C.A.; Hallsworth, J.E.; Roberts, D.W. Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation. Curr. Genet. 2015, 61, 383–404. [Google Scholar] [CrossRef] [PubMed]
- Rangel, D.E.N.; Roberts, D.W. Possible source of the high UV-B and heat tolerance of Metarhizium acridum (isolate ARSEF 324). J. Invertebr. Pathol. 2018, 157, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Ekobu, M.; Solera, M.; Kyamanywa, S.; Mwanga, R.O.M.; Odongo, B.; Ghislain, M.; Moar, W.J. Toxicity of seven Bacillus thuringiensis cry proteins against Cylas puncticollis and Cylas brunneus (Coleoptera: Brentidae) using a novel artificial diet. J. Econ. Entomol. 2010, 103, 1493–1502. [Google Scholar] [CrossRef]
- Mahmoud, S.B.; Ramos, J.E.; Shatters, R.G., Jr.; Hall, D.G.; Lapointe, S.L.; Niedz, R.P.; Rougé, P.; Cave, R.D.; Borovsky, D. Expression of Bacillus thuringiensis cytolytic toxin (Cyt2Ca1) in citrus roots to control Diaprepes abbreviatus larvae. Pestic. Biochem. Phys. 2017, 136, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Anbesse, S.A.; Adge, B.J.; Gebru, W.M. Laboratory screening for virulent entomopathogenic nematodes (Heterorhabditis bacteriophora and Steinernema yirgalemense) and fungi (Metarhizium anisopliae and Beauveria bassiana) and assessment of possible synergistic effects of combined use against grubs of the barley chafer Coptognathus curtipennis. Nematology 2008, 10, 701–709. [Google Scholar]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2006, 31, 162–171. [Google Scholar] [CrossRef]
- de Oliveira, F.Q.; Batista, J.D.; Malaquias, J.B.; de Brito, C.H.; Dos Santos, E.P. Susceptibility of the predator Euborellia annulipes (Dermaptera: Anisolabididae) to mycoinsecticides. Rev. Colomb. Entomol. 2011, 37, 234–237. [Google Scholar]
- Cottrell, T.E.; Shapiro-Ilan, D.I. Susceptibility of endemic and exotic North American ladybirds (Coleoptera: Coccinellidae) to endemic fungal entomopathogens. Eur. J. Entomol. 2008, 105, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Kim, J.J. Target-oriented dissemination of Beauveria bassiana conidia by the predators, Harmonia axyridis (Coleoptera: Coccinellidae) and Chrysoperla carnea (Neuroptera: Chrysopidae) for biocontrol of Myzus persicae. Biocontrol Sci. Technol. 2012, 22, 393–406. [Google Scholar] [CrossRef]
- Cottrell, T.E.; Shapiro-Ilan, D.I. Susceptibility of a native and an exotic lady beetle (Coleoptera: Coccinellidae) to Beauveria bassiana. J. Invertebr. Pathol. 2003, 84, 137–144. [Google Scholar] [CrossRef]
- de la Rosa, W.; Segura, H.R.; Barrera, J.F.; Williams, T. Laboratory evaluation of the impact of entomopathogenic fungi on Prorops nasuta (Hymenoptera: Bethylidae), a parasitoid of the coffee berry borer. Environ. Entomol. 2000, 29, 126–131. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Lozano, E.; Roman, J.C.; Pietrowski, V.; Neves, P.M.O.J. Interactions between Beauveria bassiana and Trichogramma pretiosum under laboratory conditions. Entomol. Exp. Appl. 2015, 154, 213–221. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Lozano, E.R.; Bonini, A.K.; Neves, P.M.O.J. Potential side effects of the entomopathogenic fungus Metarhizium anisopliae on the egg parasitoid Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) under controlled conditions. J. Econ. Entomol. 2017, 110, 2318–2324. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Haas, J.; da Silva, E.R.L.; Daros, A.; Pietrowski, V.; Neves, P.M.O.J. Selectivity of Beauveria bassiana and Metarhizium anisopliae to Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Neotrop. Entomol. 2009, 38, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.; Gómez, J.; Infante, F.; Vega, F.E. Susceptibility of the parasitoid Phymastichus coffea LaSalle (Hymenoptera: Eulophidae) to Beauveria bassiana under laboratory conditions. Neotrop. Entomol. 2009, 38, 665–670. [Google Scholar] [CrossRef]
- De la Rosa, W.; Godinez, J.L.; Alatorre, R.; Trujillo, J. Susceptibility of the parasitoid Cephalonomia stephanoderis to Beauveria bassiana and Metarhizium anisopliae strains. Southwest Entomol. 1997, 22, 233–242. [Google Scholar]
- Khun, K.K.; Ash, G.J.; Stevens, M.M.; Huwer, R.K.; Wilson, B.A.L. Compatibility of Metarhizium anisopliae and Beauveria bassiana with insecticides and fungicides used in macadamia production in Australia. Pest Manag. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khun, K.K.; Wilson, B.A.L.; Stevens, M.M.; Huwer, R.K.; Ash, G.J. Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops. Insects 2020, 11, 659. https://doi.org/10.3390/insects11100659
Khun KK, Wilson BAL, Stevens MM, Huwer RK, Ash GJ. Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops. Insects. 2020; 11(10):659. https://doi.org/10.3390/insects11100659
Chicago/Turabian StyleKhun, Kim Khuy, Bree A. L. Wilson, Mark M. Stevens, Ruth K. Huwer, and Gavin J. Ash. 2020. "Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops" Insects 11, no. 10: 659. https://doi.org/10.3390/insects11100659