
Diversity of Ginsenoside Profiles Produced by Various Processing Technologies

 , , ,
, , ,
Abstract
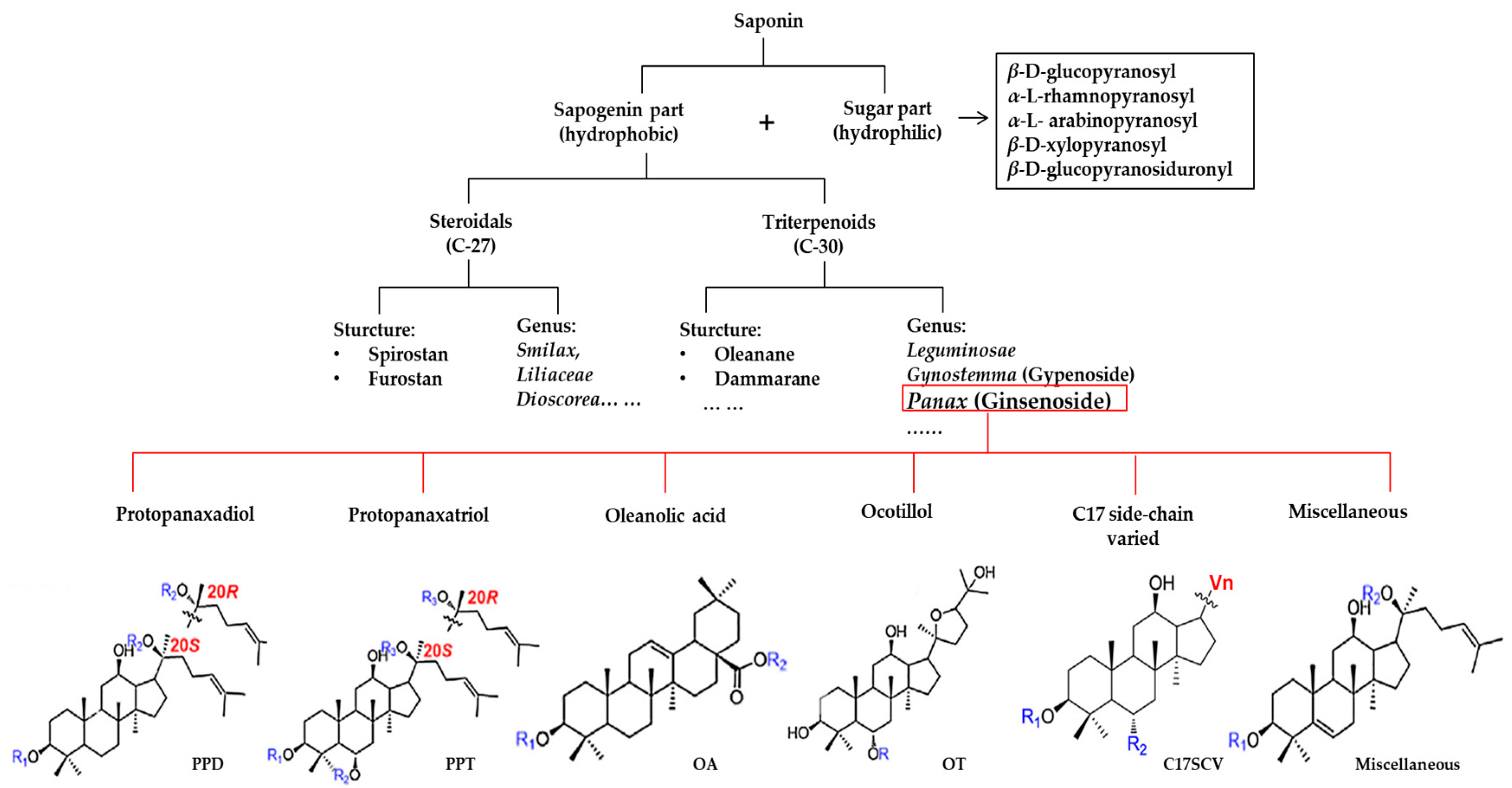
:1. Introduction
2. Comparison of the Major Ginsenosides in Various Panax Species
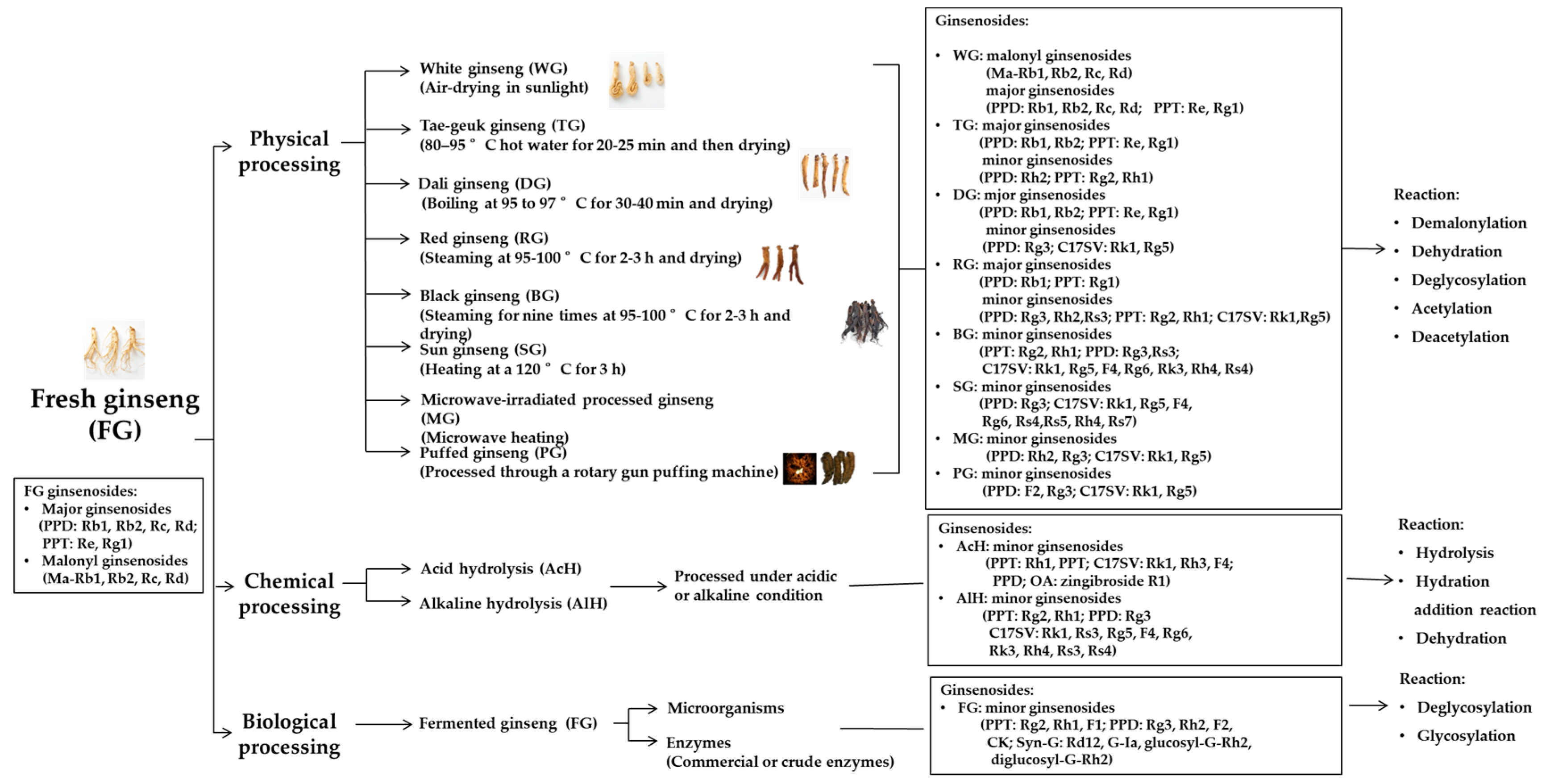
3. Variations in Ginsenoside Compositions due to Different Processing Technologies
3.1. Ginsenoside Variations in Physically Processed Ginseng
3.1.1. Fresh Ginseng (FG) and White Ginseng (WG)
3.1.2. Tae-Geuk Ginseng (TG) and Dali Ginseng (DG)
3.1.3. Red Ginseng (RG) and Black Ginseng (BG)
3.1.4. Sun Ginseng (SG)
3.1.5. Sulfur-Fumigated Ginseng
3.1.6. Microwave-Irradiated Ginseng and Puffed Ginseng
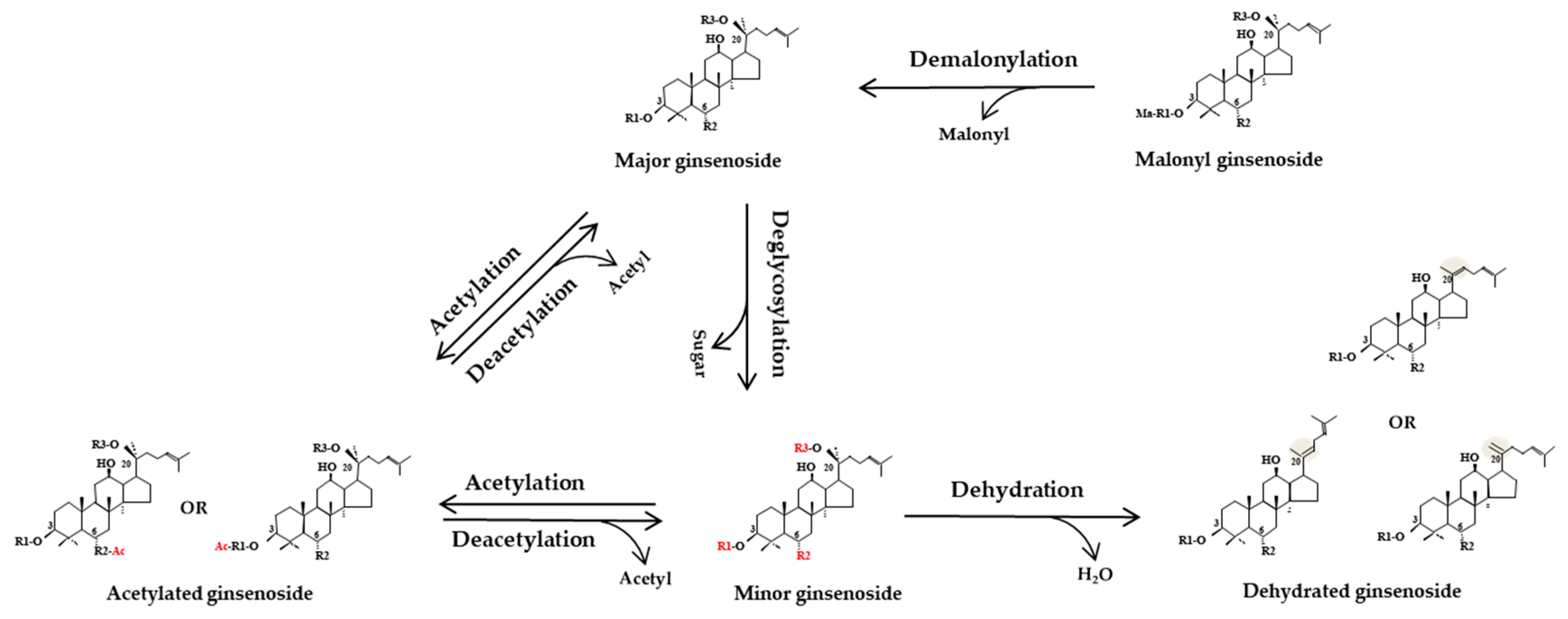
3.1.7. Transformation Pathways of Ginsenosides during Physical Processing
3.2. Ginsenoside Variation in Chemically Processed Ginseng
3.2.1. Ginsenoside Composition after Acid Hydrolysis
3.2.2. Ginsenoside Composition after Alkaline Hydrolysis
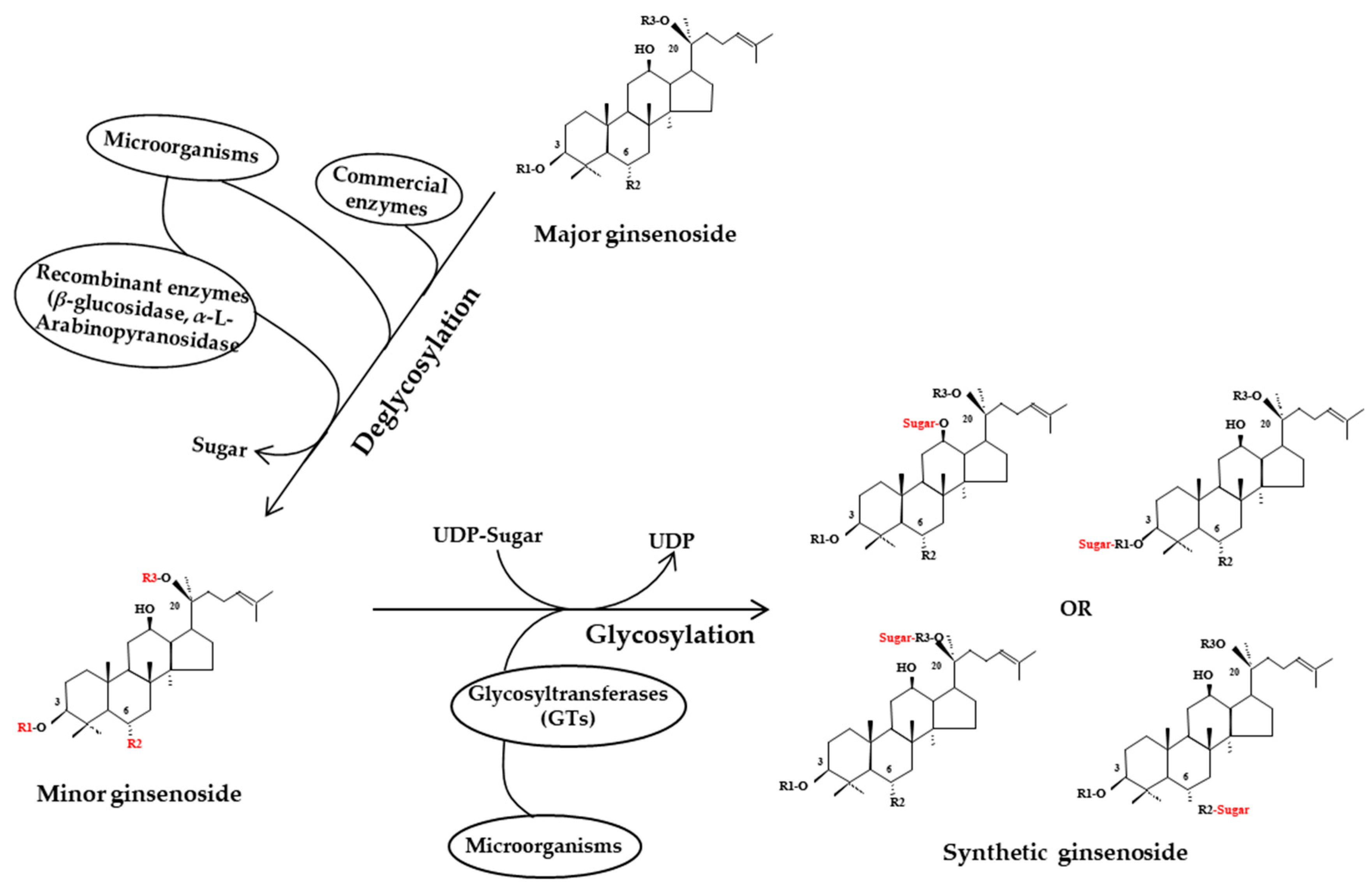
3.3. Ginsenoside Variation in Biologically Processed Ginseng
3.3.1. Microbial Ginseng Fermentation
3.3.2. Enzymatically Fermented Ginseng
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Güçlü-Üstündağ, Ö.; Mazza, G. Saponins: Properties, applications and processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef]
- Mishra, N.S.; Prakash, R.; Mishra, R.C. Phytomedicinal uses of saponin containing herbs. Int. J. Chem. Stud. 2017, 1, 12–16. [Google Scholar]
- Kregiel, D.; Berlowska, J.; Witonska, I.; Antolak, H.; Proestos, C.; Babic, M.; Babic, L.; Zhang, B. Saponin-based, biological-active surfactants from plants. Appl. Charact. Surfactants 2017, 183–205. [Google Scholar] [CrossRef]
- Tian, L.W.; Zhang, Z.; Long, H.L.; Zhang, Y.J. Steroidal Saponins from the genus Smilax and their biological activities. Nat. Prod. Bioprospect 2017, 7, 283–298. [Google Scholar] [CrossRef] [Green Version]
- Hao, D.; Gu, X.; Xiao, P.; Peng, Y. Chemical and biological research of Clematis medicinal resources. Chin. Sci. Bull. 2013, 58, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.Z.; Shi, X.J.; Yao, C.L.; Huang, Y.; Hou, J.J.; Han, S.M.; Feng, Z.J.; Wei, W.L.; Wu, W.Y.; Guo, D.A. A novel neutral loss/product ion scan-incorporated integral approach for the untargeted characterization and comparison of the carboxyl-free ginsenosides from Panax ginseng, Panax quinquefolius, and Panax notoginseng. J. Pharm. Biomed. Anal. 2020, 177, 112813. [Google Scholar] [CrossRef]
- Jung, J.; Lee, N.K.; Paik, H.D. Bioconversion, health benefits, and application of ginseng and red ginseng in dairy products. Food Sci. Biotechnol. 2017, 26, 1155–1168. [Google Scholar] [CrossRef]
- Yang, X. Triterpenoids in Panax ginseng. Mod. Chin. Med. 2016, 18, 7–15. [Google Scholar]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax L.(Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Christensen, L.P. Ginsenosides: Chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2008, 55, 1–99. [Google Scholar] [CrossRef]
- Zhang, H.; Abid, S.; Ahn, J.C.; Mathiyalagan, R.; Kim, Y.J.; Yang, D.C.; Wang, Y. Characteristics of Panax ginseng Cultivars in Korea and China. Molecules 2020, 25, 2635. [Google Scholar] [CrossRef] [PubMed]
- Baeg, I.H.; So, S.H. The world ginseng market and the ginseng (Korea). J. Ginseng Res. 2013, 37, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Qi, L.W.; Du, G.J.; Mehendale, S.R.; Wang, C.Z.; Yuan, C.S. Red notoginseng: Higher ginsenoside content and stronger anticancer potential than Asian and American ginseng. Food Chem. 2011, 125, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.Y.; Fan, Y.; Yu, Q.T.; Ge, Y.Z.; Yan, C.P.; Alolga, R.N.; Li, P.; Ma, Z.H.; Qi, L.W. Integrated evaluation of malonyl ginsenosides, amino acids and polysaccharides in fresh and processed ginseng. J. Pharm. Biomed. Anal. 2015, 107, 89–97. [Google Scholar] [CrossRef]
- Lim, C.Y.; Moon, J.M.; Kim, B.Y.; Lim, S.H.; Lee, G.S.; Yu, H.S.; Cho, S.I. Comparative study of Korean White Ginseng and Korean Red Ginseng on efficacies of OVA-induced asthma model in mice. J. Ginseng Res. 2015, 39, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.; Ryu, G.H. Effects of extrusion cooking on physicochemical properties of white and red ginseng (powder). J. Ginseng Res. 2014, 38, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Han, B.H.; Park, M.H.; Han, Y.N.; Shin, S.C. Studies on the antioxidant components of Korean ginseng (IV) Antifatigue active components. Yakhak Hoeji 1984, 28, 231–235. [Google Scholar]
- Bhattacharya, S.; Mitra, S. Anxiolytic activity of Panax ginseng roots: An experimental study. J. Ethnopharmacol. 1991, 34, 87–92. [Google Scholar] [CrossRef]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a North American ginseng extract. Mol. Cell Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef]
- Hu, C.; Kitts, D.D. Free radical scavenging capacity as related to antioxidant activity and ginsenoside composition of Asian and North American ginseng extracts. J. Am. Oil Chem. Soc. 2001, 78, 249–255. [Google Scholar] [CrossRef]
- Angelova, N.; Kong, H.W.; Van Der Heijden, R.; Yang, S.Y.; Choi, Y.H.; Kim, H.K.; Wang, M.; Hankemeier, T.; Van Der Greef, J.; Xu, G. Recent methodology in the phytochemical analysis of ginseng. Phytochem. Anal. 2008, 19, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.C.; Shin, B.K.; Cho, I.; Koo, H.; Kim, M.; Han, J. Anti-obesity effect of carbon dioxide supercritical fluid extracts of Panax ginseng CA Meyer. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 738–743. [Google Scholar] [CrossRef]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.C. Molecular signaling of ginsenosides Rb1, Rg1, and Rg3 and their mode of actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Balan, P.; Popovich, D.G. Analysis of Ginsenoside Content (Panax ginseng) from Different Regions. Molecules 2019, 24, 3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.E.; Na, C.S.; Yoo, S.A.; Seo, S.H.; Son, H.S. Biotransformation of major ginsenosides in ginsenoside model culture by lactic acid bacteria. J. Ginseng Res. 2017, 41, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Qiao, X.; Li, K.; Fan, J.; Bo, T.; Guo, D.A.; Ye, M. Identification and differentiation of Panax ginseng, Panax quinquefolium, and Panax notoginseng by monitoring multiple diagnostic chemical markers. Acta Pharm. Sin. B 2016, 6, 568–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. A review of ginseng species in different regions as a multipurpose herb in traditional Chinese medicine, modern herbology and pharmacological science. J. Med. Plant. Res. 2019, 13, 213–226. [Google Scholar]
- Shin, B.K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H. Chemical diversity of Panax ginseng, Panax quinquifolium, and Panax notoginseng. J. Ginseng Res. 2012, 36, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Balan, P.; Popovich, D.G. Comparison of ginsenoside components of various tissues of New Zealand forest-grown Asian ginseng (Panax ginseng) and American ginseng (Panax quinquefolium L.). Biomolecules 2020, 10, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, K.; Zhu, S.; Meselhy, M.R.; Tohda, C.; Cai, S.; Komatsu, K. Dammarane-type saponins from Panax japonicus and their neurite outgrowth activity in SK-N-SH cells. J. Nat. Prod. 2002, 65, 1288–1292. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Zhu, S.; Tohda, C.; Cai, S.; Komatsu, K. Dammarane-type Triterpene Saponins from Panax japonicus. J. Nat. Prod. 2002, 65, 346–351. [Google Scholar] [CrossRef]
- Morita, T.; Kasai, R.; Tanaka, O.; Zhou, J.; Yang, T.; Shoji, J. Saponins of Zu-Tziseng, rhizomes of Panax japonicus CA Meyer var. major (BURK.) CY Wu et KM FENG, collected in Yunnan, China. Chem. Pharm. Bull. 1982, 30, 4341–4345. [Google Scholar] [CrossRef]
- Morita, T.; Tanaka, O.; Kohda, H. Saponin composition of rhizomes of Panax japonicus collected in South Kyushu, Japan, and its significance in oriental traditional medicine. Chem. Pharm. Bull. 1985, 33, 3852–3858. [Google Scholar] [CrossRef] [Green Version]
- Huong, N.T.T.; Matsumoto, K.; Yamasaki, K.; Duc, N.M.; Nham, N.T.; Watanabe, H. Crude sponin extracted from Vietnamese ginseng and its major constituent majonoside-R2 attenuate the psychological stress-and foot-shock stress-induced antinociception in mice. Pharm. Biochem. Behav. 1995, 52, 427–432. [Google Scholar] [CrossRef]
- Van Le, T.H.; Lee, S.Y.; Kim, T.R.; Kim, J.Y.; Kwon, S.W.; Nguyen, N.K.; Park, J.H.; Nguyen, M.D. Processed Vietnamese ginseng: Preliminary results in chemistry and biological activity. J. Ginseng Res. 2014, 38, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Huong, N.T.T.; Matsumoto, K.; Yamasaki, K.; Duc, N.M.; Nham, N.T.; Watanabe, H. Majonoside-R2, a major constituent of Vietnamese ginseng, attenuates opioid-induced antinociception. Pharm. Biochem. Behav. 1997, 57, 285–291. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, Z.; Wu, M.; Zhou, J.; Tanaka, O. Studies on saponins of rhizomes of Panax zingiberensis Wu et Feng. Acta Pharm. Sin. 1984, 19, 232–236. [Google Scholar]
- Tang, Q.Y.; Chen, G.; Song, W.L.; Fan, W.; Wei, K.H.; He, S.M.; Zhang, G.H.; Tang, J.R.; Li, Y.; Lin, Y. Transcriptome analysis of Panax zingiberensis identifies genes encoding oleanolic acid glucuronosyltransferase involved in the biosynthesis of oleanane-type ginsenosides. Planta 2019, 249, 393–406. [Google Scholar] [CrossRef]
- Le Tran, Q.; Than, M.M.; Tezuka, Y.; Banskota, A.H.; Kouda, K.; Watanabe, H.; Zhu, S.; Komatsu, K.; Thet, M.M.; Swe, T. Wild ginseng grows in Myanmar. Chem. Pharm. Bull. (Tokyo) 2003, 51, 679–682. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Li, J.; Zhang, X.; Pei, J.; Huang, L. An integrated LC-MS-based strategy for the quality assessment and discrimination of three Panax species. Molecules 2018, 23, 2988. [Google Scholar] [CrossRef] [Green Version]
- Yang, C. Two new saponins of oleanolic acid from the underground part of Panax stipuleanus Tsai et Feng. Acta Bot. Yunnanica 1985, 7, 103–108. [Google Scholar]
- Komatsu, Z.K.Z.S.K. Analysis of Saponins of Panax Stipuleanatus by Using HPLC andAPIMS/MS Techniques. J. Univ. Hydraul. Electr. Eng. 2002, 24, 4. [Google Scholar]
- Liang, C.; Ding, Y.; Nguyen, H.T.; Kim, J.A.; Boo, H.J.; Kang, H.K.; Nguyen, M.C.; Kim, Y.H. Oleanane-type triterpenoids from Panax stipuleanatus and their anticancer activities. Bioorg. Med. Chem. Lett. 2010, 20, 7110–7115. [Google Scholar] [CrossRef]
- Tung, N.H.; Quang, T.H.; Ngan, N.T.T.; Van Minh, C.; Anh, B.K.; Long, P.Q.; Cuong, N.M.; Kim, Y.H. Oleanolic triterpene saponins from the roots of Panax bipinnatifidus. Chem. Pharm. Bull. (Tokyo) 2011, 59, 1417–1420. [Google Scholar] [CrossRef] [Green Version]
- Gurung, B.; Bhardwaj, P.K.; Rai, A.K.; Sahoo, D. Major ginsenoside contents in rhizomes of Panax sokpayensis and Panax bipinnatifidus. Nat. Prod. Res. 2018, 32, 234–238. [Google Scholar] [CrossRef]
- Yue, P.Y.K.; Mak, N.K.; Cheng, Y.K.; Leung, K.W.; Ng, T.B.; Fan, D.T.P.; Yeung, H.W.; Wong, R.N.S. Pharmacogenomics and the Yin/Yang actions of ginseng: Anti-tumor, angiomodulating and steroid-like activities of ginsenosides. Chin. Med. 2007, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, O. Recent studies on glycosides from plant drugs of Himalaya and south-western China: Chemo-geographical correlation of Panax species. Pure Appl. Chem. 1990, 62, 1281–1284. [Google Scholar] [CrossRef]
- Shukla, Y.N.; Thakur, R.S.; Pachaly, P. A bidesmosidic oleanolic acid saponin from Panax pseudo-ginseng. Phytochemistry 1992, 31, 1046–1048. [Google Scholar] [CrossRef]
- Tanaka, O.; Yahara, S. Dammarane saponins of leaves of Panax pseudo-ginseng subsp. himalaicus. Phytochemistry 1978, 17, 1353–1358. [Google Scholar] [CrossRef]
- Pandey, A.K.; Ali, M.A. Intraspecific variation in Panax assamicus Ban. populations based on internal transcribed spacer (ITS) sequences of nrDNA. Indian J. Biotechnol. 2012, 11, 30–38. [Google Scholar]
- Kharwanlang, L.; Das, M.C.; Kumaria, S.; Tandon, P. Histological and SEM studies on somatic embryogenesis in rhizome-derived callus of Panax assamicus. Ban. Pharm. Innov. J. 2016, 5, 93–99. [Google Scholar]
- Pandey, A.K.; Ali, M.A.; Biate, D.L.; Misra, A. Molecular systematics of Aralia-Panax complex (Araliaceae) in India based on ITS sequences of nrDNA. Proc. Natl. Acad. Sci. India 2009, 79, 255–261. [Google Scholar]
- Wen, J.; Zimmer, E.A. Phylogeny and biogeography of Panax L. (the ginseng genus, Araliaceae): Inferences from ITS sequences of nuclear ribosomal DNA. Mol. Phylogenet Evol. 1996, 6, 167–177. [Google Scholar] [CrossRef]
- Yun, T.K. Brief introduction of Panax ginseng CA Meyer. J. Korean Med. Sci. 2001, 16, S3–S5. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.M.; Marderosian, A.D. Two-dimensional TLC analysis of ginsenosides from root of dwarf ginseng (Panax trifolius L.) araliaceae. J. Pharm. Sci. 1981, 70, 89–91. [Google Scholar] [CrossRef]
- Lee, T.M.; Marderosian, A.D. A chemical study of the saponins and flavonoids of Dwarf ginseng (Panax trifolius L.) and its comparison to related species in the Araliaceae. In Proceedings of the Ginseng Society Conference; The Korean Society of Ginseng: Seoul, Korea, 1988; pp. 141–146. [Google Scholar]
- Nagulan, V.; Preeti, A. A unique type of endosperm in Panax wangianus SC Sun. J. Plant. Dev. 2013, 20, 45–50. [Google Scholar]
- Venugopal, N.; Ahuja, P. Relationship between age, size, fecundity and climatic factors in Panax wangianus an endangered medicinal plant in the sacred grove forest of North-East India. J. For. Res. 2011, 22, 427–435. [Google Scholar] [CrossRef]
- Venugopal, N.; Preeti, A. Seed germination of Panax wangianus SC Sun (Araliaceae): A critically endangered medicinal plant of Meghalaya, Northeast India. Int. J. Plant. Reprod. Biol. 2013, 5, 156–160. [Google Scholar]
- Zheng, M.M.; Xu, F.X.; Li, Y.J.; Xi, X.Z.; Cui, X.W.; Han, C.C.; Zhang, X.L. Study on transformation of ginsenosides in different methods. Biomed. Res. Int. 2017, 2017, 8601027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Li, X.; Liu, Z.; Wang, J.; Zheng, Y.; Yang, X. Isolation and structure characterization of malonyl-notoginsenoside-R4 from the root of Panax ginseng. Chem. J. Chin. Univ. 2007, 28, 1316. [Google Scholar]
- Ruan, C.C.; Liu, Z.; Li, X.; Liu, X.; Wang, L.J.; Pan, H.Y.; Zheng, Y.N.; Sun, G.Z.; Zhang, Y.S.; Zhang, L.X. Isolation and characterization of a new ginsenoside from the fresh root of Panax ginseng. Molecules 2010, 15, 2319–2325. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.; Jang, O.H.; Hwang, K.H.; An, Y.N.; Moon, B. Effects of steaming and air-drying on ginsenoside composition of Korean ginseng (Panax ginseng CA Meyer). J. Food Process. Preserv. 2015, 39, 207–213. [Google Scholar] [CrossRef]
- Ning, X.; Han, C. Drying characteristics and quality of taegeuk ginseng (Panax ginseng C.A. M eyer) using far-infrared rays. Int. J. Food Sci. Technol. 2013, 48, 477–483. [Google Scholar] [CrossRef]
- Yoon, D.; Shin, W.C.; Lee, Y.S.; Kim, S.; Baek, N.I.; Lee, D.Y. A comparative study on processed Panax ginseng products using HR-MAS NMR-based metabolomics. Molecules 2020, 25, 1390. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Kim, J.W.; Seguin, P.; Jun, Y.M.; Kim, S.H. Ginsenosides and phenolics in fresh and processed Korean ginseng (Panax ginseng CA Meyer): Effects of cultivation location, year, and storage period. Food Chem. 2012, 130, 73–83. [Google Scholar] [CrossRef]
- Li, S.L.; Lai, S.F.; Song, J.Z.; Qiao, C.F.; Liu, X.; Zhou, Y.; Cai, H.; Cai, B.C.; Xu, H.X. Decocting-induced chemical transformations and global quality of Du–Shen–Tang, the decoction of ginseng evaluated by UPLC–Q-TOF-MS/MS based chemical profiling approach. J. Pharm. Biomed. Anal. 2010, 53, 946–957. [Google Scholar] [CrossRef]
- Chu, C.; Xu, S.; Li, X.; Yan, J.; Liu, L. Profiling the ginsenosides of three ginseng products by Lc-Q-Tof/Ms. J. Food Sci. 2013, 78, C653–C659. [Google Scholar] [CrossRef]
- Lee, J.W.; Ji, S.H.; Choi, B.R.; Choi, D.J.; Lee, Y.G.; Kim, H.G.; Kim, G.S.; Kim, K.; Lee, Y.H.; Baek, N.I. UPLC-QTOF/MS-Based metabolomics applied for the quality evaluation of four processed Panax ginseng products. Molecules 2018, 23, 2062. [Google Scholar] [CrossRef] [Green Version]
- In, G.; Ahn, N.G.; Bae, B.S.; Lee, M.W.; Park, H.W.; Jang, K.H.; Cho, B.G.; Han, C.K.; Park, C.K.; Kwak, Y.-S. In situ analysis of chemical components induced by steaming between fresh ginseng, steamed ginseng, and red ginseng. J. Ginseng Res. 2017, 41, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.S.; Kim, H.Y.; Pyo, J.S.; Yokozawa, T. Increase in the free radical scavenging activity of ginseng by heat-processing. Biol. Pharm. Bull. 2006, 29, 750–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Kim, Y.J.; Jeon, J.N.; Wang, C.; Min, J.W.; Noh, H.Y.; Yang, D.C. Effect of white, red and black ginseng on physicochemical properties and ginsenosides. Plant. Foods Hum. Nutr. 2015, 70, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Luan, X.; Dou, D.; Huang, L. Comparative analysis of ginsenosides and oligosaccharides in White Ginseng (WG), Red Ginseng (RG) and Black Ginseng (BG). J. Chromatogr. Sci. 2019, 57, 403–410. [Google Scholar] [CrossRef]
- Sun, B.S.; Gu, L.J.; Fang, Z.M.; Wang, C.Y.; Wang, Z.; Lee, M.R.; Li, Z.; Li, J.J.; Sung, C.K. Simultaneous quantification of 19 ginsenosides in black ginseng developed from Panax ginseng by HPLC–ELSD. J. Pharm. Biomed. Anal. 2009, 50, 15–22. [Google Scholar] [CrossRef]
- Song, G.Y.; Oh, H.J.; Roh, S.S.; Seo, Y.B.; Park, Y.J.; Myung, C.S. Effect of black ginseng on body weight and lipid profiles in male rats fed normal diets. Yakhak Hoeji 2006, 50, 381–385. [Google Scholar]
- Kang, K.S.; Kim, H.Y.; Yamabe, N.; Nagai, R.; Yokozawa, T. Protective effect of sun ginseng against diabetic renal damage. Biol. Pharm. Bull. 2006, 29, 1678–1684. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.Y.; Kim, J.M.; Han, S.B.; Lee, S.K.; Kim, N.D.; Park, M.K.; Kim, C.K.; Park, J.H. Steaming of ginseng at high temperature enhances biological activity. J. Nat. Prod. 2000, 63, 1702–1704. [Google Scholar] [CrossRef]
- Keum, Y.S.; Park, K.K.; Lee, J.M.; Chun, K.S.; Park, J.H.; Lee, S.K.; Kwon, H.; Surh, Y.J. Antioxidant and anti-tumor promoting activities of the methanol extract of heat-processed ginseng. Cancer Lett. 2000, 150, 41–48. [Google Scholar] [CrossRef]
- Park, I.H.; Han, S.B.; Kim, J.M.; Piao, L.; Kwon, S.W.; Kim, N.Y.; Kang, T.L.; Park, M.K.; Park, J.H. Four new acetylated ginsenosides from processed ginseng (sun ginseng). Arch. Pharm. Res. 2002, 25, 837. [Google Scholar] [CrossRef]
- Jin, X.; Zhu, L.Y.; Shen, H.; Xu, J.; Li, S.L.; Jia, X.B.; Cai, H.; Cai, B.C.; Yan, R. Influence of sulphur-fumigation on the quality of white ginseng: A quantitative evaluation of major ginsenosides by high performance liquid chromatography. Food Chem. 2012, 135, 1141–1147. [Google Scholar] [CrossRef]
- Shen, H.; Zhang, L.; Xu, J.D.; Ding, Y.F.; Zhou, J.; Wu, J.; Zhang, W.; Mao, Q.; Liu, L.F.; Zhu, H. Effect of sulfur-fumigation process on ginseng: Metabolism and absorption evidences. J. Ethnopharmacol. 2020, 256, 112799. [Google Scholar] [CrossRef]
- Zhu, H.; Shen, H.; Xu, J.; Xu, J.D.; Zhu, L.Y.; Wu, J.; Chen, H.B.; Li, S.L. Comparative study on intestinal metabolism and absorption in vivo of ginsenosides in sulphur-fumigated and non-fumigated ginseng by ultra performance liquid chromatography quadruple time-of-flight mass spectrometry based chemical profiling approach. Drug Test. Anal. 2015, 7, 320–330. [Google Scholar] [CrossRef]
- Ren, G.; Chen, F. Degradation of ginsenosides in American ginseng (Panax quinquefolium) extracts during microwave and conventional heating. J. Agric. Food Chem. 1999, 47, 1501–1505. [Google Scholar] [CrossRef]
- Bai, Y.; Zhao, L.; Qu, C.; Meng, X.; Zhang, H. Microwave degradation of floatation-enriched ginsenoside extract from Panax quinquefolium L. leaf. J. Agric. Food Chem. 2009, 57, 10252–10260. [Google Scholar] [CrossRef]
- Park, J.Y.; Choi, P.; Kim, H.K.; Kang, K.S.; Ham, J. Increase in apoptotic effect of Panax ginseng by microwave processing in human prostate cancer cells: In vitro and in vivo studies. J. Ginseng Res. 2016, 40, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Ahn, S.C.; Choi, S.W.; Hur, N.Y.; Kim, B.Y.; Baik, M.Y. Changes in effective components of ginseng by puffing. J. Korean Soc. Appl. Biol. Chem. 2008, 51, 188–193. [Google Scholar]
- Wang, Y.; Li, X.; Jin, H.; Yu, Y.; You, J.; Zhang, K.; Ding, L.; Zhang, H. Degradation of ginsenosides in root of Panax ginseng CA Meyer. by high-pressure microwave-assisted extraction. Chem. J. Chin. Univ. 2007, 28, 2264–2269. [Google Scholar]
- Lee, S.M.; Shon, H.J.; Choi, C.S.; Hung, T.M.; Min, B.S.; Bae, K. Ginsenosides from heat processed ginseng. Chem. Pharm. Bull. 2009, 57, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M. Thermal conversion pathways of ginsenoside in red ginseng processing. Nat. Prod. Sci. 2014, 20, 119–125. [Google Scholar]
- Chang, K.H.; Park, S.H.; Jee, H.S.; Kim, K.T.; Paik, H.D.; Lee, J.Y. Enhancement of the cytotoxicities and antioxidative activities of white ginseng extract by hydrolysis under mild acidic conditions. Food Sci. Biotechnol. 2014, 23, 173–178. [Google Scholar] [CrossRef]
- Quan, K.; Liu, Q.; Wan, J.Y.; Zhao, Y.J.; Guo, R.Z.; Alolga, R.N.; Li, P.; Qi, L.W. Rapid preparation of rare ginsenosides by acid transformation and their structure-activity relationships against cancer cells. Sci. Rep. 2015, 5, 8598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Qin, Q.; Guo, Y.; Sun, J.; Liu, S. Studies on the chemical transformation of 20 (S)-protopanaxatriol (PPT)-type ginsenosides Re, Rg2, and Rf using rapid resolution liquid chromatography coupled with quadruple-time-of-flight mass spectrometry (RRLC-Q-TOF-MS). J. Agric. Food Chem. 2012, 60, 10007–10014. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.Y.; Kim, M.Y.; Lee, Y.J.; Li, M.; Shin, Y.S.; Lee, J.; Jeong, H.S. Influence of organic acids and heat treatment on ginsenoside conversion. J. Ginseng Res. 2018, 42, 532–539. [Google Scholar] [CrossRef]
- Kim, M.H.; Hong, H.D.; Kim, Y.C.; Rhee, Y.K.; Kim, K.T.; Rho, J.H. Ginsenoside changes in red ginseng manufactured by acid impregnation treatment. J. Ginseng Res. 2010, 34, 93–97. [Google Scholar] [CrossRef]
- Bae, E.A.; Han, M.J.; Kim, E.J.; Kim, D.H. Transformation of ginseng saponins to ginsenoside Rh 2 by acids and human intestinal bacteria and biological activities of their transformants. Arch. Pharm. Res. 2004, 27, 61–67. [Google Scholar] [CrossRef]
- Zhang, C.; Guan, F.; Zhang, L. A preliminary study on the improvement of protopanaxadiol yield by alkaline hydrolysis. J. Baotou Med. Coll. 2011, 27, 1–5. [Google Scholar]
- Ma, L.Y.; Zhou, Q.L.; Yang, X.W. New SIRT1 activator from alkaline hydrolysate of total saponins in the stems-leaves of Panax ginseng. Bioorg. Med. Chem. Lett. 2015, 25, 5321–5325. [Google Scholar] [CrossRef]
- Ma, L.; Yang, X. 20 (R)-Ginsenoside-Rh19, a novel ginsenoside from alkaline hydrolysates of total saponins in stems-leaves of Panax ginseng. Chin. Tradit. Herb. Drugs 2016, 47, 6–14. [Google Scholar] [CrossRef]
- Cui, J.F.; Eneroth, P.; Bruhn, J.G.; Arihara, S.; Yoshikawa, K. Alkaline cleavage of gypenosides and characterization of dammarane-type aglycones by gas chromatography–mass spectrometry. Phytochem. Anal. 1998, 9, 128–133. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, D.; Zhou, D.; Ye, Z. Study on hydrolysis kinetics of ginsenoside-Ro in alkaline medium and structural analysis of its hydrolyzate. China J. Chin. Mater. Med. 2014, 39, 867–872. [Google Scholar]
- Akao, T.; Kanaoka, M.; Kobashi, K. Appearance of compound K, a major metabolite of ginsenoside Rb1 by intestinal bacteria, in rat plasma after oral administration: Measurement of compound K by enzyme immunoassay. Biol. Pharm. Bull. 1998, 21, 245–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.M.; Lee, J.Y.; Lee, Y.G.; Baek, S.; Kim, M.R. Enhanced production of compound K in fermented ginseng extracts by Lactobacillus brevis. Food Sci. Biotechnol. 2019, 28, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.I.; Cho, C.W.; Choi, U.K.; Kim, Y.C. Antioxidant activity and ginsenoside pattern of fermented white ginseng. J. Ginseng Res. 2010, 34, 168–174. [Google Scholar] [CrossRef]
- Kim, B.G.; Choi, S.Y.; Kim, M.R.; Suh, H.J.; Park, H.J. Changes of ginsenosides in Korean red ginseng (Panax ginseng) fermented by Lactobacillus plantarum M1. Process. Biochem. 2010, 45, 1319–1324. [Google Scholar] [CrossRef]
- Bae, E.A.; Hyun, Y.J.; Choo, M.K.; Oh, J.K.; Ryu, J.H.; Kim, D.H. Protective effect of fermented red ginseng on a transient focal ischemic rats. Arch. Pharm. Res. 2004, 27, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Pham, Q.L.; Jang, H.J.; Kim, K.B. Anti-wrinkle effect of fermented black ginseng on human fibroblasts. Int. J. Mol. Med. 2017, 39, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Bak, M.J.; Jeong, W.S.; Kim, K.B. Detoxifying effect of fermented black ginseng on H2O2-induced oxidative stress in HepG2 cells. Int. J. Mol. Med. 2014, 34, 1516–1522. [Google Scholar] [CrossRef] [Green Version]
- Chi, H.; Kim, D.H.; Ji, G.E. Transformation of ginsenosides Rb2 and Rc from Panax ginseng by food microorganisms. Biol. Pharm. Bull. 2005, 28, 2102–2105. [Google Scholar] [CrossRef] [Green Version]
- Park, C.S.; Yoo, M.H.; Noh, K.H.; Oh, D.K. Biotransformation of ginsenosides by hydrolyzing the sugar moieties of ginsenosides using microbial glycosidases. Appl. Microbiol. Biotechnol. 2010, 87, 9–19. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, S.Y.; Park, Y.; Jung, E.Y.; Suh, H.J. Enzymatic transformation of ginsenosides in Korean Red Ginseng (Panax ginseng Meyer) extract prepared by Spezyme and Optidex. J. Ginseng Res. 2014, 38, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.W.; Han, S.H.; Lee, S.W.; Choi, H.S.; Suh, H.J.; Hong, K.B. Enzymatic hydrolysis increases ginsenoside content in Korean red ginseng (Panax ginseng CA Meyer) and its biotransformation under hydrostatic pressure. J. Sci. Food Agric. 2019, 99, 6806–6813. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Lim, S.; Kim, S.O.; Ahn, S.H.; Choi, Y.J. Optimization of enzymatic treatment for compound K production from white ginseng extract by response surface methodology. Biosci. Biotechnol. Biochem. 2013, 77, 1138–1140. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Yang, Y.Y.; Ouyang, D.S.; Yang, G.P. A review of biotransformation and pharmacology of ginsenoside compound K. Fitoterapia 2015, 100, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.H.; Jee, H.S.; Lee, N.K.; Park, S.H.; Lee, N.W.; Paik, H.D. Optimization of the enzymatic production of 20 (S)-ginsenoside Rg3 from white ginseng extract using response surface methodology. New Biotechnol. 2009, 26, 181–186. [Google Scholar] [CrossRef]
- Sunwoo, H.H.; Gujral, N.; Huebl, A.C.; Kim, C.T. Application of high hydrostatic pressure and enzymatic hydrolysis for the extraction of ginsenosides from fresh ginseng root (Panax ginseng CA Myer). Food Bioprocess. Technol. 2014, 7, 1246–1254. [Google Scholar] [CrossRef]
- Wang, Y.; Choi, K.D.; Yu, H.; Jin, F.; Im, W.T. Production of ginsenoside F1 using commercial enzyme Cellulase KN. J. Ginseng Res. 2016, 40, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lee, S.J.; Yuan, Q.P.; Im, W.T.; Kim, S.C.; Han, N.S. Production of bioactive ginsenoside Rg3 (S) and compound K using recombinant Lactococcus lactis. J. Ginseng Res. 2018, 42, 412–418. [Google Scholar] [CrossRef]
- Quan, L.H.; Min, J.W.; Jin, Y.; Wang, C.; Kim, Y.J.; Yang, D.C. Enzymatic biotransformation of ginsenoside Rb1 to compound K by recombinant β-glucosidase from Microbacterium esteraromaticum. J. Agric. Food Chem. 2012, 60, 3776–3781. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, X.M.; Yan, H.J.; Liu, X.Y.; Zhou, Q.; Luo, Z.Y.; Tan, X.N.; Zhang, B.L. Highly selective production of compound K from ginsenoside Rd by hydrolyzing glucose at C-3 glycoside using β-glucosidase of Bifidobacterium breve ATCC 15700. J. Microbiol. Biotechnol. 2019, 29, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Huq, M.A.; Siraj, F.M.; Kim, Y.J.; Yang, D.C. Enzymatic transformation of ginseng leaf saponin by recombinant β-glucosidase (bgp1) and its efficacy in an adipocyte cell line. Biotechnol. Appl. Biochem. 2016, 63, 532–538. [Google Scholar] [CrossRef]
- Ruan, C.C.; Zhang, H.; Zhang, L.X.; Liu, Z.; Sun, G.Z.; Lei, J.; Qin, Y.X.; Zheng, Y.N.; Li, X.; Pan, H.Y. Biotransformation of ginsenoside Rf to Rh1 by recombinant β-glucosidase. Molecules 2009, 14, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Jung, S.Y.; Kim, Y.J.; Lee, D.Y.; Aceituno, V.C.; Wang, C.; Yang, D.C. Microbial deglycosylation and ketonization of ginsenoside by Cladosporium cladosporioide and their anticancer activity. Antonie Van Leeuwenhoek 2016, 109, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Jung, S.Y.; Kim, Y.J.; Lee, D.Y.; Min, J.W.; Wang, C.; Yang, D.C. Microbial ketonization of ginsenosides F1 and C–K by Lactobacillus brevis. Antonie Van Leeuwenhoek 2014, 106, 1215–1221. [Google Scholar] [CrossRef]
- Kim, J.H.; Oh, J.M.; Chun, S.; Park, H.Y.; Im, W.T. Enzymatic Biotransformation of Ginsenoside Rb 2 into Rd by Recombinant α-L-Arabinopyranosidase from Blastococcus saxobsidens. J. Microbiol. Biotechnol. 2020, 30, 391–397. [Google Scholar] [CrossRef]
- Yang, J.L.; Hu, Z.F.; Zhang, T.T.; Gu, A.D.; Gong, T.; Zhu, P. Progress on the studies of the key enzymes of ginsenoside biosynthesis. Molecules 2018, 23, 589. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.S.; Lee, H.J.; Mathiyalagan, R.; Kim, Y.J.; Yang, D.U.; Lee, D.Y.; Min, J.W.; Jimenez, Z.; Yang, D.C. Synthesis of a novel α-glucosyl ginsenoside F1 by cyclodextrin glucanotransferase and its in vitro cosmetic applications. Biomolecules 2018, 8, 142. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Kim, Y.J.; Baek, N.I.; Mathiyalagan, R.; Wang, C.; Jin, Y.; Xu, X.Y.; Yang, D.C. Glycosyltransformation of ginsenoside Rh2 into two novel ginsenosides using recombinant glycosyltransferase from Lactobacillus rhamnosus and its in vitro applications. J. Ginseng Res. 2019, in press. [Google Scholar] [CrossRef]
- Hu, Y.; Li, H.; Qu, Y.; Zhang, X.; Zhang, J.; Dai, L. Biocatalytic synthesis of a novel bioactive ginsenoside using UDP-glycosyltransferase from Bacillus Subtilis 168. Catalysts 2020, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Jin, Y.; Wang, C.; Kim, Y.J.; Perez, Z.E.J.; Baek, N.I.; Mathiyalagan, R.; Markus, J.; Yang, D.C. Rare ginsenoside Ia synthesized from F1 by cloning and overexpression of the UDP-glycosyltransferase gene from Bacillus subtilis: Synthesis, characterization, and in vitro melanogenesis inhibition activity in BL6B16 cells. J. Ginseng Res. 2018, 42, 42–49. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Geographical Distribution | Main Saponin Types | Distinctiveness | Ref. |
|---|---|---|---|---|---|
| P. ginseng | Korean ginseng | Asian countries | PPT and PPD | G–Rf (PPT)and G–Rs1 (PPD) | [24,25,26,28,29,30] |
| P. quinquefolius | American ginseng | America | PPT and PPD | P–F11 (OT) | [26,29,31] |
| P. notoginseng | Chinese (Sanchi) ginseng | China | PPT and PPD | Noto–R1 (PPT) | [26,29] |
| P. japonicas | Japanese ginsengor Ye–sanchi | China and Japan | OT | Yesanchinosides (OT) | [32,33,34,35] |
| P. vietnamensis | Vietnam ginseng | Vietnam | PPT, PPD, and OT | Majon–R2 (OT) | [36,37,38] |
| P. zingiberensis | Ginger ginseng or Myanmar ginseng | China | OA and PPT | - | [39,40,41,42] |
| P. stipuleanatus | Pingpien ginseng | China and Vietnam | OA | - | [43,44,45] |
| P. bipinnatifidus | Feather-leaf bamboo ginseng | China, Eastern Himalayas, and Nepal | OA | - | [46,47,48] |
| P. sokpayensis | - | India | PPT and PPD | - | [47] |
| P. omeiensis | Omei ginseng | China, Eastern Himalayas, and Nepal | - | - | [11,48] |
| P. pseudoginseng | Himalayan ginseng | China, Eastern Himalayas, and Nepal | PPT, PPD, and OA | [11,48,49,50,51] | |
| P. assamicus | - | India and West Bengal | - | - | [52,53,54] |
| P. shangianus | - | China | - | - | [11,54] |
| P. sinensis | - | China | - | - | [11,55,56] |
| P. trifolius | Dwarf ginseng | Ohio andPennsylvania | PPT, PPD, and OA | - | [48,57,58] |
| P. variabilis | - | China and India | - | - | [11,54] |
| P. wangianus | Narrow-leaved pseudoginseng | China and India | - | - | [48,59,60,61] |
| Types | Name | R1 | R2 | R3 | Remark |
|---|---|---|---|---|---|
PPD | G–Rb1 | glc(2–1)glc | H | glc(6–1)glc | Major ginsenoside |
| G–Rb2 | glc(2–1)glc | H | glc(6–1)ara(p) | Major ginsenoside | |
| G–Rc | glc(2–1)glc | H | glc(6–1)ara(f) | Major ginsenoside | |
| G–Rd | glc(2–1)glc | H | glc(6–1) | Major ginsenoside | |
| Ma–Rb1 | glc(2–1)glc(6)Ma | H | glc(6–1)glc | Malonyl ginsenoside | |
| Ma–Rb2 | glc(2–1)glc(6)Ma | H | glc(6–1)ara(p) | Malonyl ginsenoside | |
| Ma–Rc | glc(2–1)glc(6)Ma | H | glc(6–1)ara(f) | Malonyl ginsenoside | |
| Ma–Rd | glc(2–1)glc(6)Ma | H | glc(6–1) | Malonyl ginsenoside | |
| G–Rb3 | glc(2–1)glc | H | glc(6–1)xyl | Ginsenoside | |
| G–Ra3 | glc(2–1)glc | H | glc(6–1)glc(3–1)xyl | Ginsenoside | |
| Noto–R4 | glc | H | glc(6–1)glc(6–1)xyl | Notoginsenoside | |
| G–Rs1 | glc(2–1)glc(6)Ac | H | glc(6–1)ara(p) | Acetylated ginsenoside | |
| G–Rs3 | glc(2–1)glc(6)Ac | H | H | Acetylated ginsenoside | |
| G–F2 | glc | H | glc | Minor ginsenoside | |
| G–Rg3 | glc(2–1)glc | H | H | Minor ginsenoside | |
| G–Rh2 | glc | H | H | Minor ginsenoside | |
| G–Mc | H | H | glc(6–1)ara(f) | Minor ginsenoside | |
| G–Compound O | glc | H | glc(6–1)ara(p) | Minor ginsenoside | |
| G–Compound Y | H | H | glc(6–1)ara(p) | Minor ginsenoside | |
| G–Compound K | H | H | glc | Minor ginsenoside | |
PPT | G–Re | H | Oglc(2–1)rha | glc | Major Ginsenoside |
| G–Rg1 | H | Oglc | glc | Major Ginsenoside | |
| G–Rf | H | Oglc(2–1)glc | H | Major Ginsenoside | |
| G–Rg2 | H | Oglc(2–1)rha | H | Minor Ginsenoside | |
| G–Rh1 | H | Oglc | H | Minor Ginsenoside | |
| G–F1 | H | OH | glc | Minor Ginsenoside | |
| G–F3 | H | OH | glc(6–1)ara(p) | Minor Ginsenoside | |
| Noto–R1 | H | Oglc(2–1)xyl | glc | Notoginsenoside | |
C17SCV–1 | G–Rg5 | glc(2–1)glc | H | Dehydrated ginsenoside | |
| G–F4 | H | Oglc(2–1)rha | Dehydrated ginsenoside | ||
| G–Rh4 | H | Oglc | Dehydrated ginsenoside | ||
| G–Rh3 | glc | H | Dehydrated ginsenoside | ||
| (20E)–G–Rg9 | H | Oglc(2–1)glc | Dehydrated ginsenoside | ||
| G–Rs4 | glc(2–1)glc(6)Ac | H | Acetylated ginsenoside | ||
C17SCV–2 | G–Rs6 | H | Oglc(6)Ac | Acetylated ginsenoside | |
| G–Rk1 | glc(2–1)glc | H | Dehydrated ginsenoside | ||
| G–Rk3 | H | Oglc | Dehydrated ginsenoside | ||
| G–Rg6 | H | Oglc(2–1)rha | Dehydrated ginsenoside | ||
| G–Rk2 | glc | H | Dehydrated ginsenoside | ||
| G–Rg10 | H | Oglc(2–1)glc | Dehydrated ginsenoside | ||
| G–Rs5 | glc(2–1)glc(6)Ac | H | Acetylated ginsenoside | ||
| G–Rs7 | H | Oglc(6)Ac | Acetylated ginsenoside | ||
C17SCV–3 | G–Rz1 | glc(2–1)glc | H | Dehydrated ginsenoside | |
| (20Z) –G–Rg9 | H | Oglc(2–1)glc | Dehydrated ginsenoside | ||
| G–Rh16 | glc | H | Dehydrated ginsenoside | ||
C17SCV–4 | G–Rf2 | H | Oglc(2–1)rha | Hydration addition reaction | |
| G–Rf3 | H | Oglc(2–1)glc | Hydration addition reaction | ||
OT | P–F11 | H | Oglc(2–1)rha | Pseudoginsenoside | |
| Majon–R1 | H | Oglc(2–1)glc | Majonoside | ||
| Majon–R2 | H | Oglc(2–1)xyl | Majonoside | ||
| Vina–R1 | H | O6–Ac–glc(2–1)rha | Vinaginsenoside | ||
| Vina–R2 | H | O6–Ac–glc(2–1)xyl | Vinaginsenoside | ||
OA | G–Ro | glcUA(2-1)glc | -glc | Ginsenoside | |
| CS–IV | glcUA(4-1)ara | -glc | Chikusetsu saponin | ||
| CS–IVa | glcUA | -glc | Chikusetsu saponin | ||
| Zing–R1 | glc(2-1)glc | H | Zingibroside | ||
| Spina-A methyl ester | 6-CH3-glcA(3-1)glc | H | Spinasaponin | ||
| Stipul–R1 | [3-O-glc]-glcA(4-1)ara(f) | H | Stipuleanoside | ||
| Stipul–R2 | [3-O-glc]-glcA(4-1)ara(f) | -glc | Stipuleanoside | ||
| Pseudo–RP1 | glcA(2-1)xyl | H | Pesudoginsenoside | ||
| Pseudo–RT1 | glcA(2-1)xyl | -glc | Pesudoginsenoside | ||
| Bifi-A | -6-CH3-glcA(2-1)ara(p) | H | Bifinoside | ||
| Bifi-B | -6-CH3-glcA(2-1)glc(6-1)xyl | H | Bifinoside | ||
| Bifi-C | -6-CH3-glcA [3-ara(p)]glc(2-1)xyl | -glc | Bifinoside | ||
Dammarane | Glucosyl-G- Rh2 | -glc(6-1)glc | H | H | Synthetic ginsenoside |
| Diglucosyl-G-Rh2 | -glc(6-1)glc(6-1)glc | H | H | Synthetic ginsenoside | |
| G-Ia | -glc | -OH | glc | Synthetic ginsenoside | |
| α-Glycosylated-G F1 | H | H | -glc(1-2)α-D-glucopyranoside | Synthetic ginsenoside | |
Miscellaneous-1 | 3-oxo-CK | - | H | -glc | Ketonization ginsenoside |
| 3-oxo-PPD | - | H | H | Ketonization ginsenoside | |
Miscellaneous-2 | G-Rd12 | -glc(2-1)glc | H | -glc | Synthetic ginsenoside |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piao, X.M.; Huo, Y.; Kang, J.P.; Mathiyalagan, R.; Zhang, H.; Yang, D.U.; Kim, M.; Yang, D.C.; Kang, S.C.; Wang, Y.P. Diversity of Ginsenoside Profiles Produced by Various Processing Technologies. Molecules 2020, 25, 4390. https://doi.org/10.3390/molecules25194390
Piao XM, Huo Y, Kang JP, Mathiyalagan R, Zhang H, Yang DU, Kim M, Yang DC, Kang SC, Wang YP. Diversity of Ginsenoside Profiles Produced by Various Processing Technologies. Molecules. 2020; 25(19):4390. https://doi.org/10.3390/molecules25194390
Chicago/Turabian StylePiao, Xiang Min, Yue Huo, Jong Pyo Kang, Ramya Mathiyalagan, Hao Zhang, Dong Uk Yang, Mia Kim, Deok Chun Yang, Se Chan Kang, and Ying Ping Wang. 2020. "Diversity of Ginsenoside Profiles Produced by Various Processing Technologies" Molecules 25, no. 19: 4390. https://doi.org/10.3390/molecules25194390