The Effect of Modified Porcine Surfactant Alone or in Combination with Polymyxin B on Lung Homeostasis in LPS-Challenged and Mechanically Ventilated Adult Rats

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. First Series
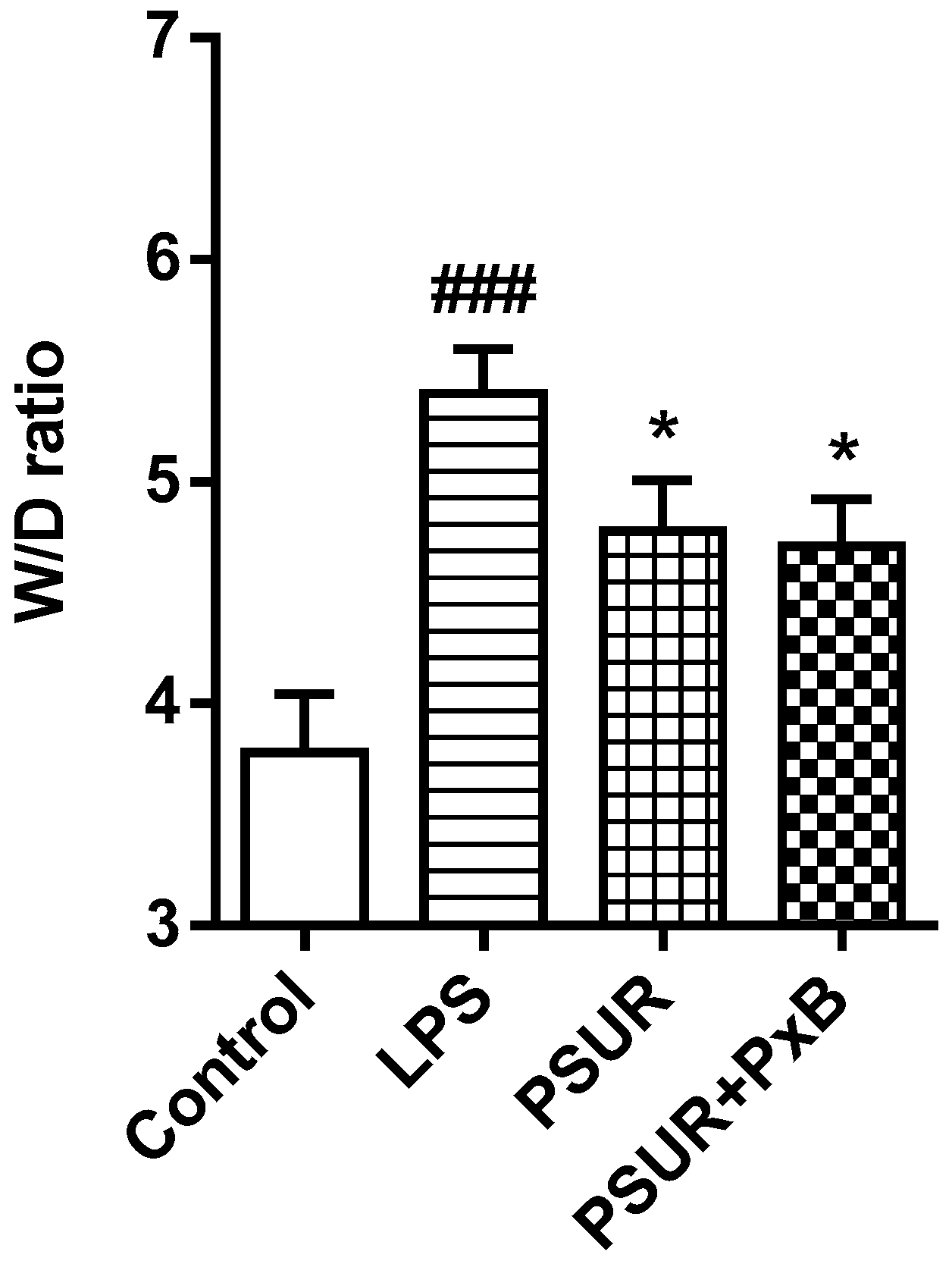
2.1.1. Lung Oedema Formation
2.1.2. Inflammatory Markers and Vascular-Specific Biomarker
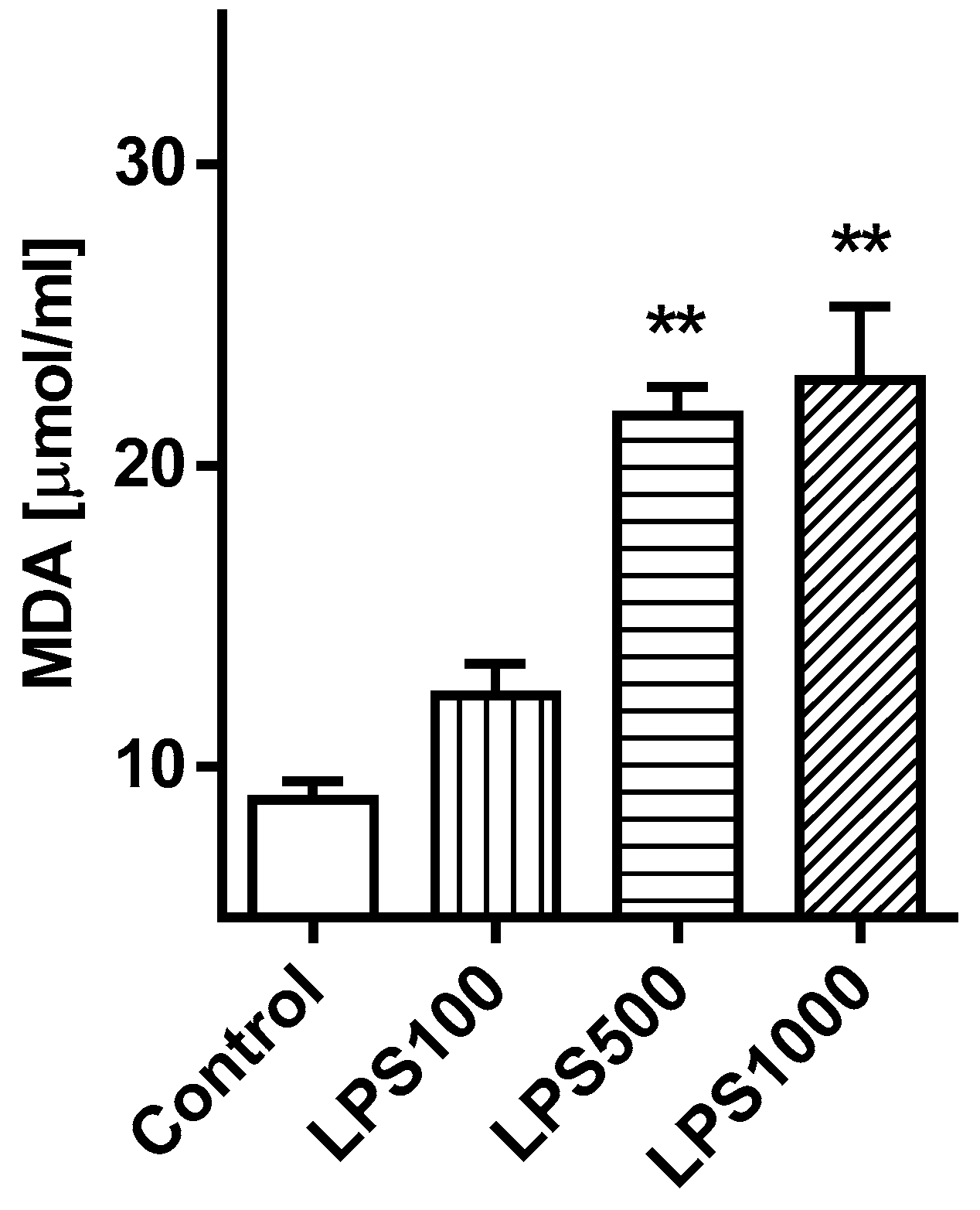
2.1.3. Oxidative Damage of the Lungs
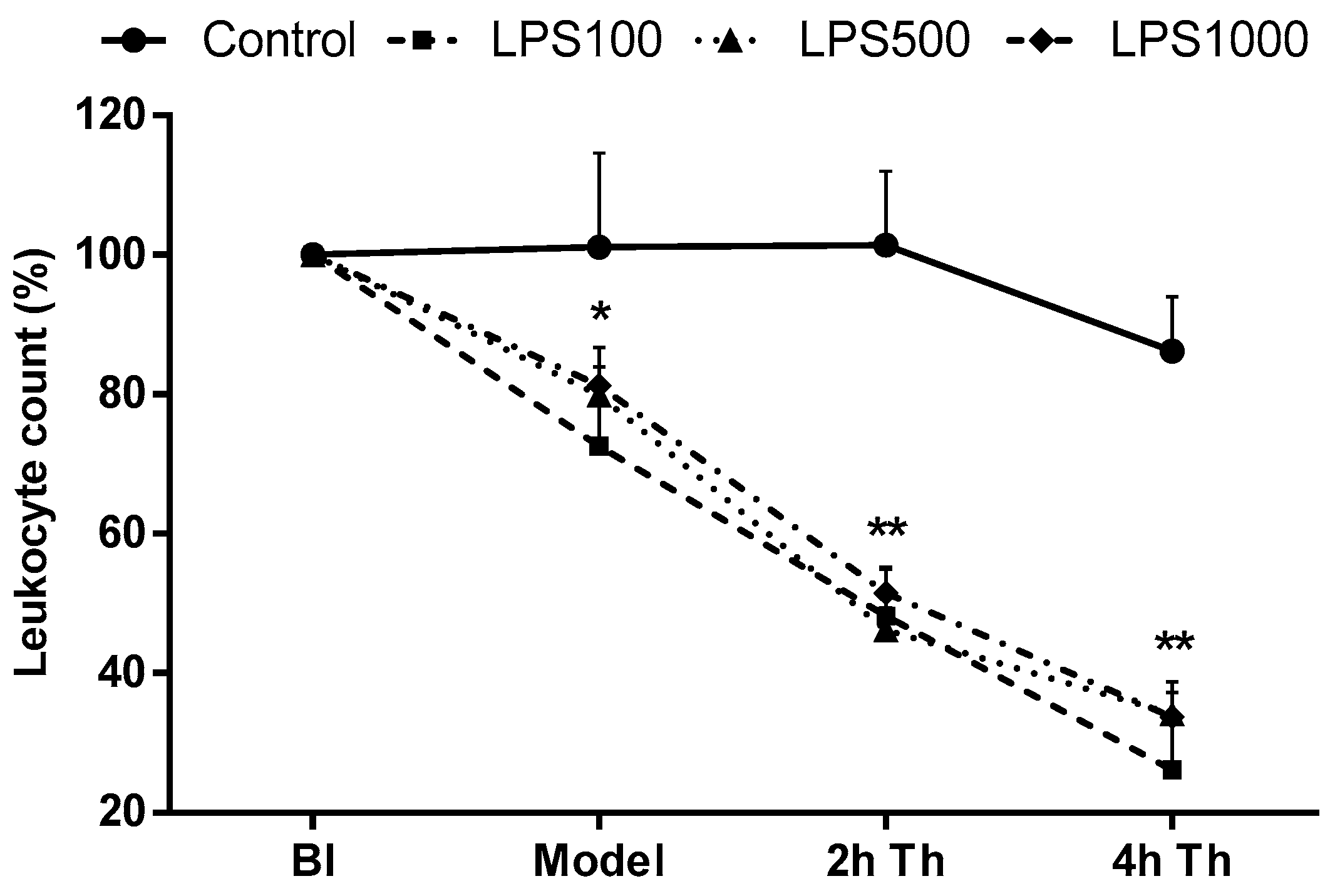
2.1.4. Total Leukocyte Count
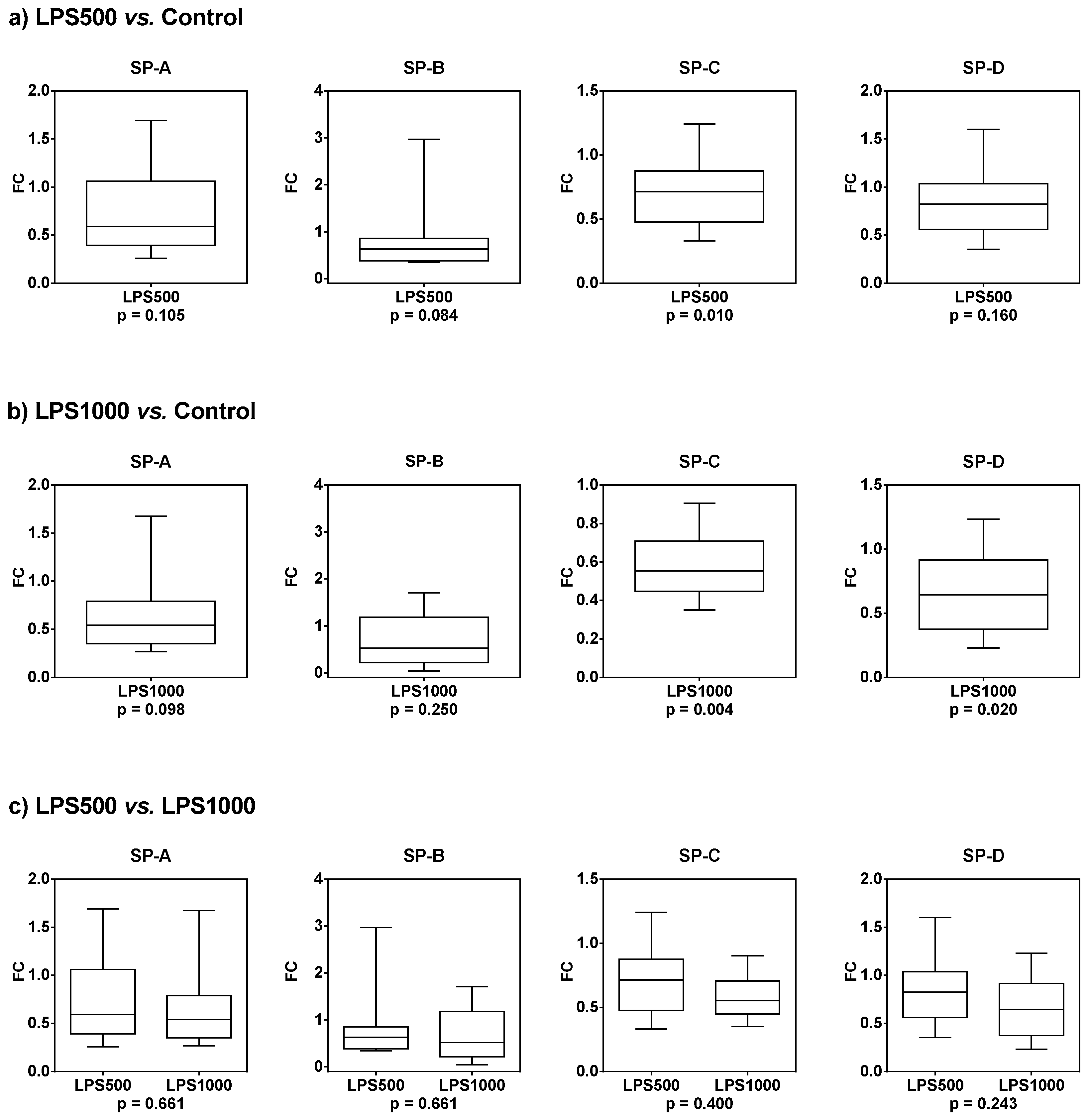
2.1.5. Gene Expression of Surfactant Proteins SP-A, SP-B, SP-C, and SP-D
2.2. Second Series
2.2.1. Lung Oedema Formation
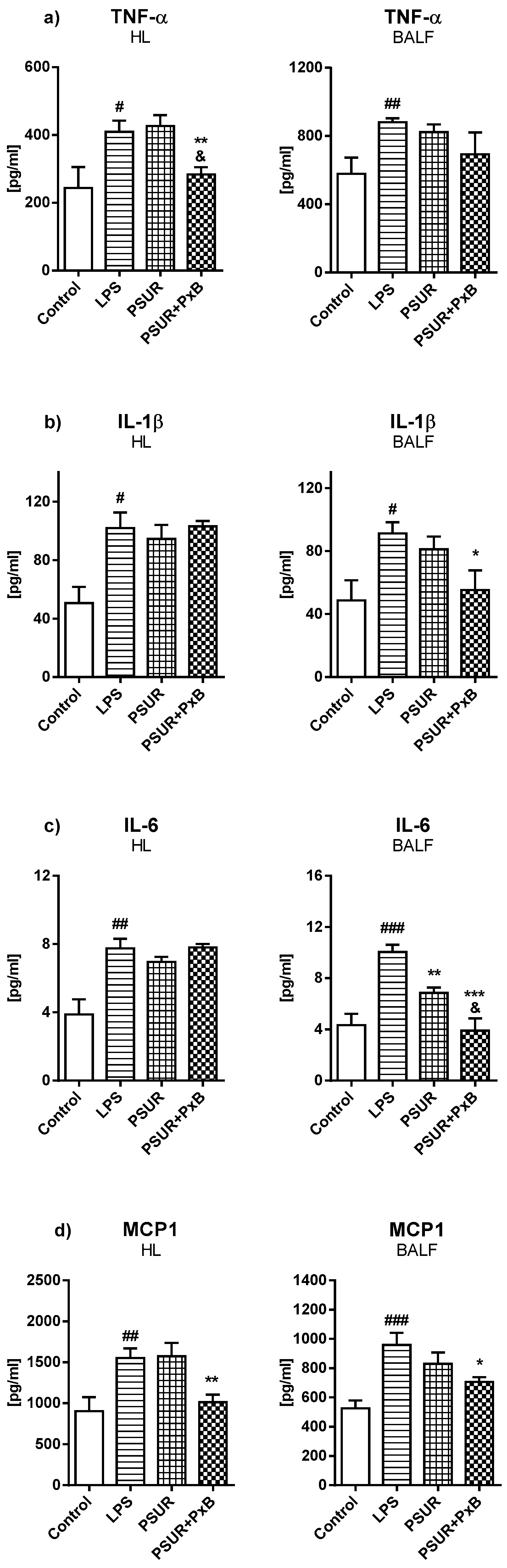
2.2.2. Cytokines and Inflammatory Markers
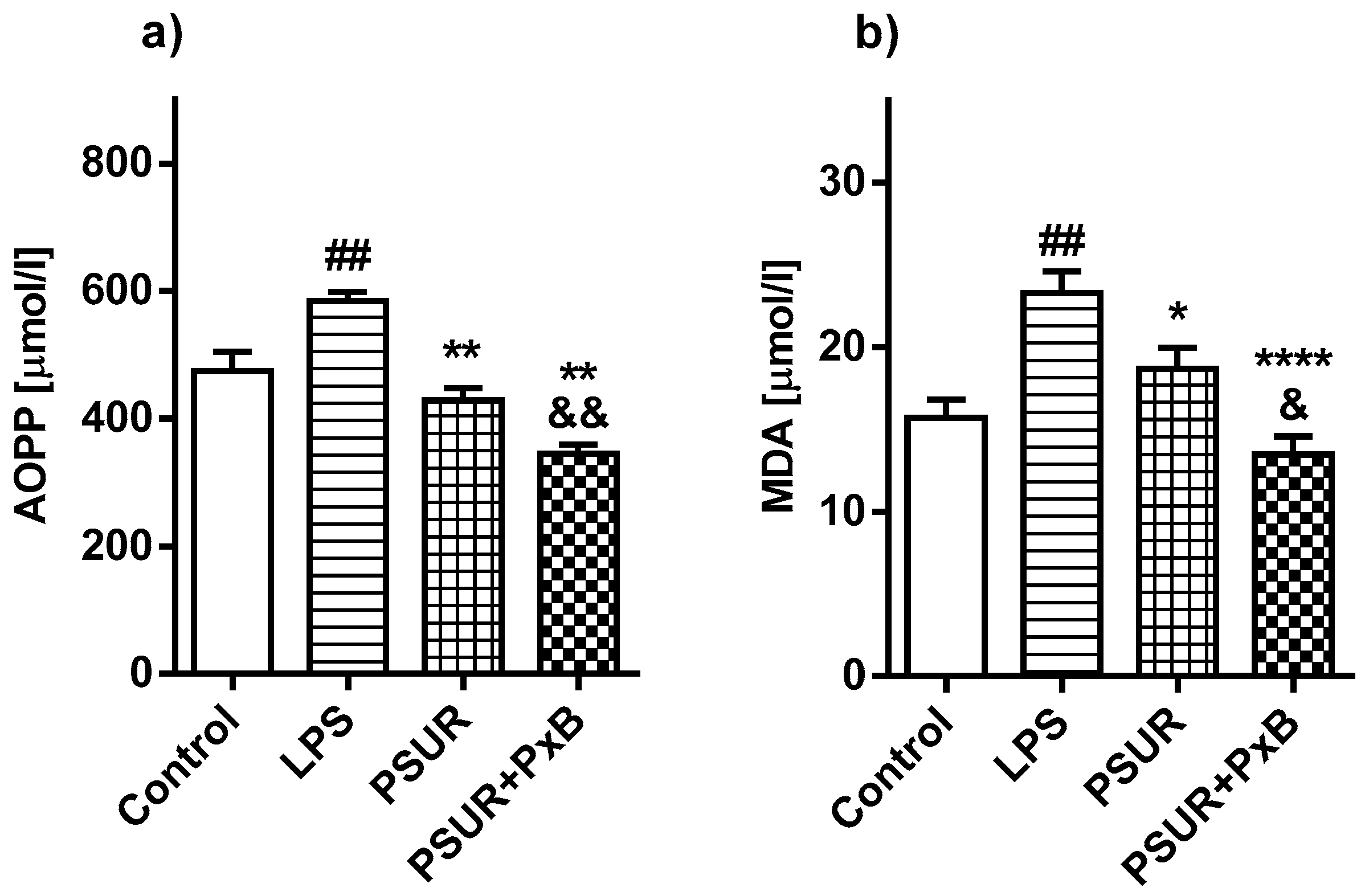
2.2.3. Oxidative Damage of the Lungs
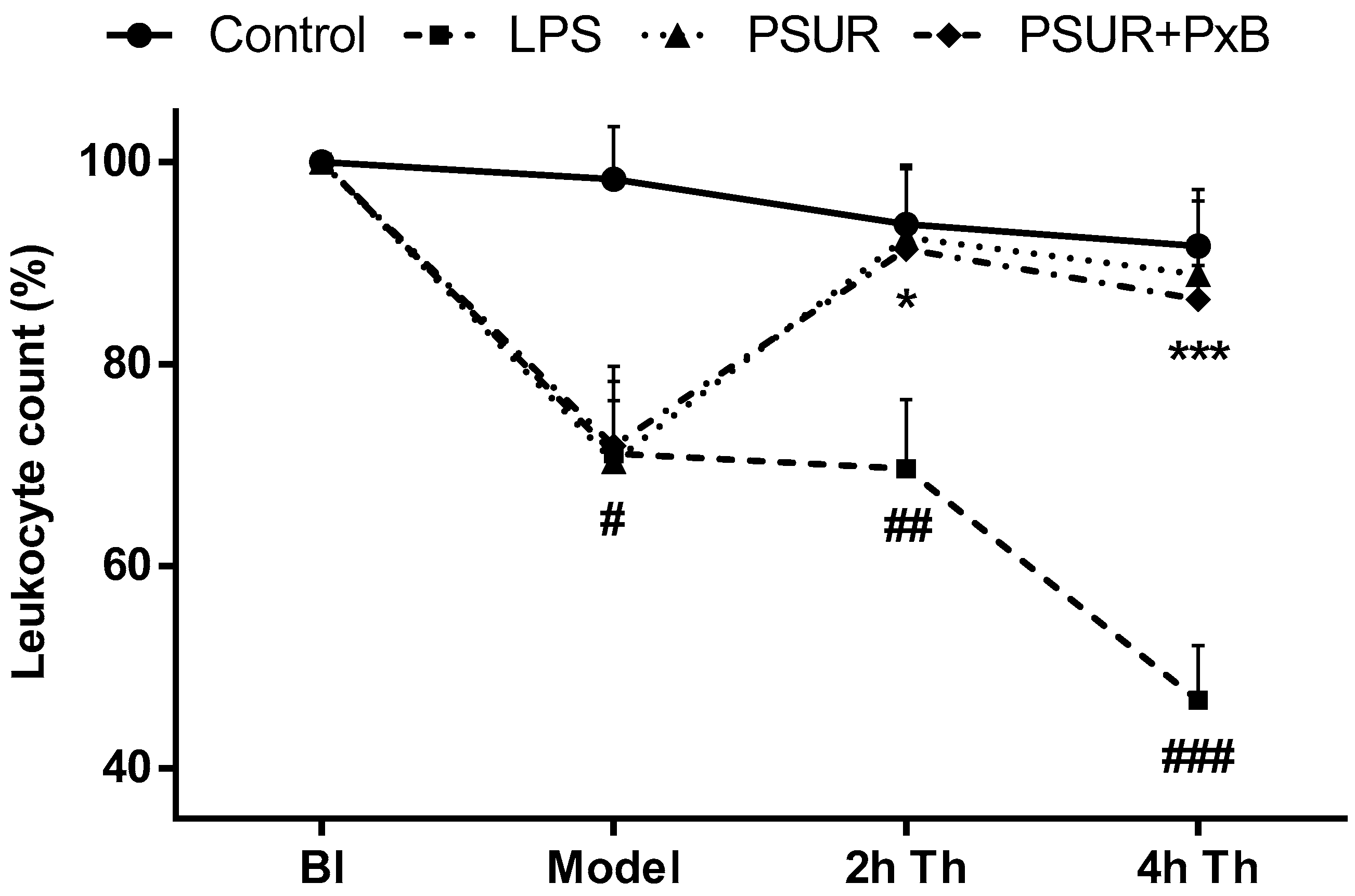
2.2.4. Total Leukocyte Count
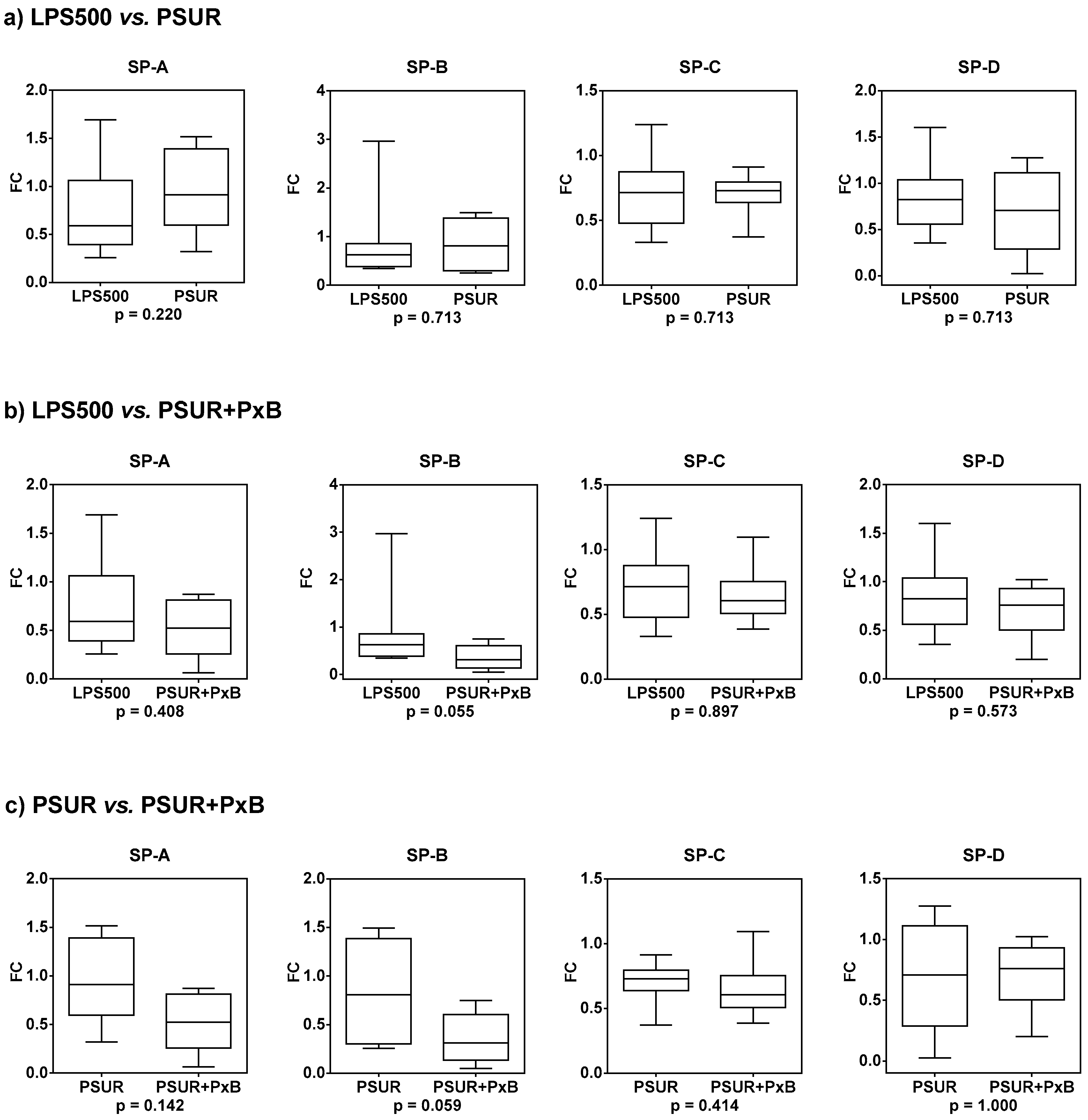
2.2.5. Gene Expression of Surfactant Proteins SP-A, SP-B, SP-C, and SP-D
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.2.1. Lipopolysaccharide (LPS)
4.2.2. Modified Porcine Surfactant
4.2.3. Polymyxin B (PxB)
4.3. First Series
4.4. Second Series
4.5. The Study Design
4.6. Assays
4.7. Evaluation of Lung Edema
4.8. Total Leukocyte Count
4.9. Evaluation of Gene Expression of Surfactant Proteins by Quantitative Real-Time PCR
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maldonado, R.F.; Sá-Correia, I.; Valvano, M.A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef]
- Xiang, M.; Fan, J.; Fan, J. Association of Toll-like receptor signaling and reactive oxygen species: A potential therapeutic target for posttrauma acute lung injury. Mediat. Inflamm. 2010, 2010, 916425. [Google Scholar] [CrossRef]
- Han, S.; Mallampalli, R.K. The Role of Surfactant in Lung Disease and Host Defense against Pulmonary Infections. Ann. Am. Thorac. Soc. 2015, 12, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Salvesen, B.; Stenvik, J.; Rossetti, C.; Saugstad, O.D.; Espevik, T.; Mollnes, T.E. Meconium-induced release of cytokines is mediated by the TRL4/MD-2 complex in a CD14-dependent manner. Mol. Immunol. 2010, 47, 1226–1234. [Google Scholar] [CrossRef]
- Thompson, B.T.; Chambers, R.C.; Liu, K.D. Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2017, 377, 562–572. [Google Scholar] [CrossRef]
- Meng, S.S.; Chang, W.; Lu, Z.H.; Xie, J.F.; Qiu, H.B.; Yang, Y.; Guo, F.M. Effect of surfactant administration on outcomes of adult patients in acute respiratory distress syndrome: A meta-analysis of randomized controlled trials. BMC Pulm. Med. 2019, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- McAllister, S.M.; Alpar, H.O.; Brown, M.R. Antimicrobial properties of liposomal polymyxin B. J. Antimicrob. Chemother. 1999, 43, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaltash, S.; Palmblad, M.; Curstedt, T.; Johansson, J.; Persson, B. Pulmonary surfactant protein B: A structural model and a functional analogue. Biochim. Biophys. Acta 2000, 1466, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Calkovska, A.; Some, M.; Linderholm, B.; Johansson, J.; Curstedt, T.; Robertson, B. Biophysical and physiological properties of porcine surfactant enriched with polymyxin B. Neonatology 2005, 88, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kolomaznik, M.; Liskayova, G.; Kanjakova, N.; Hubcik, L.; Uhrikova, D.; Calkovska, A. The Perturbation of Pulmonary Surfactant by Bacterial Lipopolysaccharide and Its Reversal by Polymyxin B: Function and Structure. Int. J. Mol. Sci. 2018, 19, 1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Kasiakou, S.K. Toxicity of polymyxins: A systematic review of the evidence from old and recent studies. Crit. Care 2006, 10, R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, B.; Souza, L.M.P.; Pimentel, A.S.; Veldhuizen, R.A.W. New insights into exogenous surfactant as a carrier of pulmonary therapeutics. Biochem. Pharmacol. 2019, 164, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Stichtenoth, G.; Linderholm, B.; Björkman, M.H.; Walter, G.; Curstedt, T.; Herting, E. Prophylactic intratracheal polymyxin B/surfactant prevents bacterial growth in neonatal Escherichia coli pneumonia of rabbits. Pediatr. Res. 2010, 67, 369–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basabe-Burgos, O.; Zebialowicz, J.; Stichtenoth, G.; Curstedt, T.; Bergman, P.; Johansson, J.; Rising, A. Natural Derived Surfactant Preparation As a Carrier of Polymyxin E for Treatment of Pseudomonas aeruginosa Pneumonia in a Near-Term Rabbit Model. J. Aerosol Med. Pulm. Drug Deliv. 2019, 32, 110–118. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Velkov, T.; Lin, Y.W.; Yun, B.; Nowell, C.J.; Zhou, F.; Zhou, Q.T.; Chan, K.; Azad, M.A.K.; Li, J. Potential Toxicity of Polymyxins in Human Lung Epithelial Cells. Antimicrob. Agents Chemother. 2017, 61, e02690-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.U.; Velkov, T.; Zhou, Q.T.; Fulcher, A.J.; Callaghan, J.; Zhou, F.; Chan, K.; Azad, M.A.K.; Li, J. Intracellular localization of polymyxins in human alveolar epithelial cells. J. Antimicrob. Chemother. 2019, 74, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Stichtenoth, G.; Herting, E.; Rüdiger, M.; Wemhöner, A. Pulmonary surfactant preserves viability of alveolar type II cells exposed to polymyxin B in vitro. PLoS ONE. 2013, 8, e62105. [Google Scholar] [CrossRef] [Green Version]
- Whitsett, J.A.; Wert, S.E.; Weaver, T.E. Alveolar surfactant homeostasis and the pathogenesis of pulmonary disease. Annu. Rev. Med. 2010, 61, 105–119. [Google Scholar] [CrossRef] [Green Version]
- Harrison, L.H., Jr.; Beller, J.J.; Gill, C.C.; Hinshaw, L.B.; Coalson, J.J.; Greenfield, L.J. Effects of endotoxin on pulmonary capillary permeability and surfactant. Surg. Forum 1968, 19, 250–251. [Google Scholar]
- Brogden, K.A.; Cutlip, R.C.; Lehmkuhl, H.D. Complexing of bacterial lipopolysaccharide with lung surfactant. Infect. Immun. 1986, 52, 644–649. [Google Scholar] [CrossRef] [Green Version]
- Augusto, L.A.; Li, J.; Synguelakis, M.; Johansson, J.; Chaby, R. Structural basis for interactions between lung surfactant protein C and bacterial lipopolysaccharide. J. Biol. Chem. 2002, 277, 23484–23492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augusto, L.A.; Synguelakis, M.; Johansson, J.; Pedron, T.; Girard, R.; Chaby, R. Interaction of pulmonary surfactant protein C with CD14 and lipopolysaccharide. Infect. Immun. 2003, 71, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chroneos, Z.C.; Sever-Chroneos, Z.; Shepherd, V.L. Pulmonary surfactant: An immunological perspective. Cell. Physiol. Biochem. 2010, 25, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Kadiiska, M.B.; Ghio, A.J.; Corbett, J.; Fann, Y.C.; Holland, S.M.; Thurman, R.G.; Mason, R.P. In vivo lipid-derived free radical formation by NADPH oxidase in acute lung injury induced by lipopolysaccharide: A model for ARDS. FASEB J. 2002, 16, 1713–1720. [Google Scholar] [CrossRef]
- Jansson, A.H.; Eriksson, C.; Wang, X. Lung inflammatory responses and hyperinflation induced by an intratracheal exposure to lipopolysaccharide in rats. Lung 2004, 182, 163–171. [Google Scholar] [CrossRef]
- Liu, F.; Li, W.; Pauluhn, J.; Trübel, H.; Wang, C. Lipopolysaccharide-induced acute lung injury in rats: Comparative assessment of intratracheal instillation and aerosol inhalation. Toxicology 2013, 304, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Alm, A.S.; Li, K.; Chen, H.; Wang, D.; Andersson, R.; Wang, X. Variation of lipopolysaccharide-induced acute lung injury in eight strains of mice. Respir. Physiol. Neurobiol. 2010, 171, 157–164. [Google Scholar] [CrossRef]
- Aul, R.; Armstrong, J.; Duvoix, A.; Lomas, D.; Hayes, B.; Miller, B.E.; Jagger, C.; Singh, D. Inhaled LPS challenges in smokers: A study of pulmonary and systemic effects. Br. J. Clin. Pharmacol. 2012, 74, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Blumenthal, S.; Borgeat, A.; Pasch, T.; Reyes, L.; Booy, C.; Lambert, M.; Schimmer, R.C.; Beck-Schimmer, B. Ropivacaine decreases inflammation in experimental endotoxin-induced lung injury. Anesthesiology 2006, 104, 961–969. [Google Scholar] [CrossRef]
- Nova, Z.; Skovierova, H.; Calkovska, A. Alveolar-Capillary Membrane-Related Pulmonary Cells as a Target in Endotoxin-Induced Acute Lung Injury. Int. J. Mol. Sci. 2019, 20, 831. [Google Scholar] [CrossRef] [Green Version]
- Ingenito, E.P.; Mora, R.; Cullivan, M.; Marzan, Y.; Haley, K.; Mark, L.; Sonna, L.A. Decreased surfactant protein-B expression and surfactant dysfunction in a murine model of acute lung injury. Am. J. Respir. Cell Mol. Biol. 2001, 25, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, N.; Sanyal, S.N. Cycloxygenase inhibition enhances the effects of surfactant therapy in endotoxin-induced rat model of ARDS. Inflammation 2011, 34, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.N.; Zhou, J.H.; Wang, P.; Zhang, X.J. The localization of SP-B and influences of lipopolysaccharide on it. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2338–2345. [Google Scholar] [PubMed]
- Zhu, T.; Li, C.; Zhang, X.; Ye, C.; Tang, S.; Zhang, W.; Sun, J.; Huang, N.; Wen, F.; Wang, D.; et al. GLP-1 Analogue Liraglutide Enhances SP-A Expression in LPS-Induced Acute Lung Injury through the TTF-1 Signaling Pathway. Mediat. Inflamm. 2018, 2018, 3601454. [Google Scholar] [CrossRef] [PubMed]
- Günther, A.; Ruppert, C.; Schmidt, R.; Markart, P.; Grimminger, F.; Walmrath, D.; Seeger, W. Surfactant alteration and replacement in acute respiratory distress syndrome. Respir. Res. 2001, 2, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Väyrynen, O.; Glumoff, V.; Hallman, M. Regulation of surfactant proteins by LPS and proinflammatory cytokines in fetal and newborn lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L803–L810. [Google Scholar] [CrossRef] [Green Version]
- Bein, K.; Di Giuseppe, M.; Mischler, S.E.; Ortiz, L.A.; Leikauf, G.D. LPS-treated macrophage cytokines repress surfactant protein-B in lung epithelial cells. Am. J. Respir. Cell Mol. Biol. 2013, 49, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.Y.; Chen, T.L.; Cherng, Y.G.; Tai, Y.T.; Chen, T.G.; Chen, R.M. Lipopolysaccharide induces apoptotic insults to human alveolar epithelial A549 cells through reactive oxygen species-mediated activation of an intrinsic mitochondrion-dependent pathway. Arch. Toxicol. 2011, 85, 209–218. [Google Scholar] [CrossRef]
- Lin, J.; Tian, J.; Wang, L.; Wu, W.; Li, H.; Wang, X.; Zeng, X.; Zhang, W. Apoptosis and surfactant protein-C expression inhibition induced by lipopolysaccharide in AEC II cell may associate with NF-κB pathway. J. Toxicol. Sci. 2017, 42, 53–61. [Google Scholar] [CrossRef]
- Nova, Z.; Skovierova, H.; Strnadel, J.; Halasova, E.; Calkovska, A. Short-Term versus Long-Term Culture of A549 Cells for Evaluating the Effects of Lipopolysaccharide on Oxidative Stress, Surfactant Proteins and Cathelicidin LL-37. Int. J. Mol. Sci. 2020, 21, 1148. [Google Scholar] [CrossRef] [Green Version]
- Kinniry, P.; Pick, J.; Stephens, S.; Jain, D.; Solomides, C.C.; Niven, R.; Segal, R.; Christofidou-Solomidou, M. KL4-surfactant prevents hyperoxic and LPS-induced lung injury in mice. Pediatr. Pulmonol. 2006, 41, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Van Helden, H.P.; Kuijpers, W.C.; Langerwerf, P.E.; Langen, R.C.; Haagsman, H.P.; Bruijnzeel, P.L. Efficacy of Curosurf in a rat model of acute respiratory distress syndrome. Eur. Respir. J. 1998, 12, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolomaznik, M.; Nova, Z.; Calkovska, A. Pulmonary surfactant and bacterial lipopolysaccharide: The interaction and its functional consequences. Physiol. Res. 2017, 66 (Suppl. 2), S147–S157. [Google Scholar] [CrossRef] [PubMed]
- Stichtenoth, G.; Jung, P.; Walter, G.; Johansson, J.; Robertson, B.; Curstedt, T.; Herting, E. Polymyxin B/pulmonary surfactant mixtures have increased resistance to inactivation by meconium and reduce growth of gram-negative bacteria in vitro. Pediatr. Res. 2006, 59, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Payne, M.S.; Miura, Y.; Ireland, D.J.; Stock, S.; Kallapur, S.G.; Kannan, P.S.; Newnham, J.P.; Kramer, B.W.; Jobe, A.H.; et al. Polymyxin B agonist capture therapy for intrauterine inflammation: Proof-of-principle in a fetal ovine model. Reprod. Sci. 2014, 21, 623–631. [Google Scholar] [CrossRef]
- Calkovska, A.; Haegerstrand-Björkman, M.; Linderholm, B.; Curstedt, T. The effect of surfactant/polymyxin B in double-hit model of neonatal lung injury. Neonatology 2017, 111, 441–442. [Google Scholar]
- Lu, X.; Chan, T.; Xu, C.; Zhu, L.; Zhou, Q.T.; Roberts, K.D.; Chan, H.K.; Li, J.; Zhou, F. Human oligopeptide transporter 2 (PEPT2) mediates cellular uptake of polymyxins. J. Antimicrob. Chemother. 2016, 71, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Bosquillon, C. Drug transporters in the lung—Do they play a role in the biopharmaceutics of inhaled drugs? J. Pharm. Sci. 2010, 99, 2240–2255. [Google Scholar] [CrossRef]
- De Fátima Fernandes Vattimo, M.; Watanabe, M.; da Fonseca, C.D.; Neiva, L.B.; Pessoa, E.A.; Borges, F.T. Polymyxin B Nephrotoxicity: From Organ to Cell Damage. PLoS ONE 2016, 11, e0161057. [Google Scholar] [CrossRef] [Green Version]
- Ozyilmaz, E.; Ebinc, F.A.; Derici, U.; Gulbahar, O.; Goktas, G.; Elmas, C.; Oguzulgen, I.K.; Sindel, S. Could nephrotoxicity due to colistin be ameliorated with the use of N-acetylcysteine? Intensive Care Med. 2011, 37, 141–146. [Google Scholar] [CrossRef]
- Yousef, J.M.; Chen, G.; Hill, P.A.; Nation, R.L.; Li, J. Ascorbic acid protects against the nephrotoxicity and apoptosis caused by colistin and affects its pharmacokinetics. J. Antimicrob. Chemother. 2012, 67, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.F.; Jäkel, C.; Haberstroh, J.; Schulte-Mönting, J.; Leititis, J.U.; Orlowska-Volk, M. Alveolar recruitment promotes homogeneous surfactant distribution in a piglet model of lung injury. Pediatr. Res. 2001, 50, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Schmiedl, A.; Grützner, D.; Hoffmann, T.; von Hörsten, S.; Stephan, M. DPP4 inhibitors increase differentially the expression of surfactant proteins in Fischer 344 rats. Acta Physiol. 2014, 212, 248–261. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 16 June 2020).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SEGA Publications, Inc.: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 16 June 2020).
Sample Availability: Samples of the compounds are not available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LPS500 vs. Control | LPS1000 vs. Control | LPS500 vs. LPS1000 | ||||
|---|---|---|---|---|---|---|
| Median FC | p-Value | Median FC | p-Value | Median FC | p-Value | |
| SP-A | 0.658 | 0.105 | 0.558 | 0.098 | 0.846 | 0.661 |
| SP-B | 0.627 | 0.084 | 0.623 | 0.250 | 0.916 | 0.661 |
| SP-C | 0.706 | 0.010 * | 0.557 | 0.004 * | 0.838 | 0.400 |
| SP-D | 0.824 | 0.160 | 0.645 | 0.020 * | 0.702 | 0.243 |
| LPS500 vs. PSUR | LPS500 vs. PSUR + PxB | PSUR vs. PSUR + PxB | ||||
|---|---|---|---|---|---|---|
| Median FC | p-Value | Median FC | p-Value | Median FC | p-Value | |
| SP-A | 1.374 | 0.220 | 0.798 | 0.408 | 0.572 | 0.142 |
| SP-B | 1.151 | 0.713 | 0.508 | 0.055 | 0.437 | 0.059 |
| SP-C | 1.022 | 0.713 | 0.950 | 0.897 | 0.894 | 0.414 |
| SP-D | 0.822 | 0.713 | 0.866 | 0.573 | 0.985 | 1.000 |
| Gene | Primer Sequence Forward and Reverse Primer | Size PCR Product (bp) |
|---|---|---|
| SFTPA | F5´-CTAAGTGCTGCCCTCTGACC-3´ R5´-AGGAGCCATACATGCCAAAC-3´ | 247 |
| SFTPB | F5´-CTGTGCCAAGAGTGTGAGGA-3´ R5´-CAAGCAGCTTCAAGGGTAGG-3’ | 124 |
| SFTPC | F5´-CAGCTCCAGGAACCTACTGC-3´ R5´-CTCTCCACACAAGGTGCTCA-3´ | 218 |
| SFTPD | F5´-ATGGCCAAAGTGTTGGAGAC-3´ R5´-CGTGCCCACATCTGTCATAC-3´ | 194 |
| RPL13a | F5´-CCCTCCACCCTATGACAAGA-3´ R5´-TTCCGGTAATGGATCTTTGC-3´ | 186 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolomaznik, M.; Kopincova, J.; Nova, Z.; Topercerova, J.; Zila, I.; Mikolka, P.; Kosutova, P.; Matasova, K., Jr.; Skovierova, H.; Grendar, M.; et al. The Effect of Modified Porcine Surfactant Alone or in Combination with Polymyxin B on Lung Homeostasis in LPS-Challenged and Mechanically Ventilated Adult Rats. Molecules 2020, 25, 4356. https://doi.org/10.3390/molecules25194356
Kolomaznik M, Kopincova J, Nova Z, Topercerova J, Zila I, Mikolka P, Kosutova P, Matasova K Jr., Skovierova H, Grendar M, et al. The Effect of Modified Porcine Surfactant Alone or in Combination with Polymyxin B on Lung Homeostasis in LPS-Challenged and Mechanically Ventilated Adult Rats. Molecules. 2020; 25(19):4356. https://doi.org/10.3390/molecules25194356
Chicago/Turabian StyleKolomaznik, Maros, Jana Kopincova, Zuzana Nova, Juliana Topercerova, Ivan Zila, Pavol Mikolka, Petra Kosutova, Katarina Matasova, Jr., Henrieta Skovierova, Marian Grendar, and et al. 2020. "The Effect of Modified Porcine Surfactant Alone or in Combination with Polymyxin B on Lung Homeostasis in LPS-Challenged and Mechanically Ventilated Adult Rats" Molecules 25, no. 19: 4356. https://doi.org/10.3390/molecules25194356