Effects of Bacillus subtilis and Pseudomonas fluorescens Inoculation on Attributes of the Lettuce (Lactuca sativa L.) Soil Rhizosphere Microbial Community: The Role of the Management System

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Inoculum Preparation
2.3. Analyses of Soil Chemical and Biochemical Variables
2.4. Phospholipid Fatty Acid Analysis
2.5. Enzymatic Activity Analysis
2.6. Data Analysis
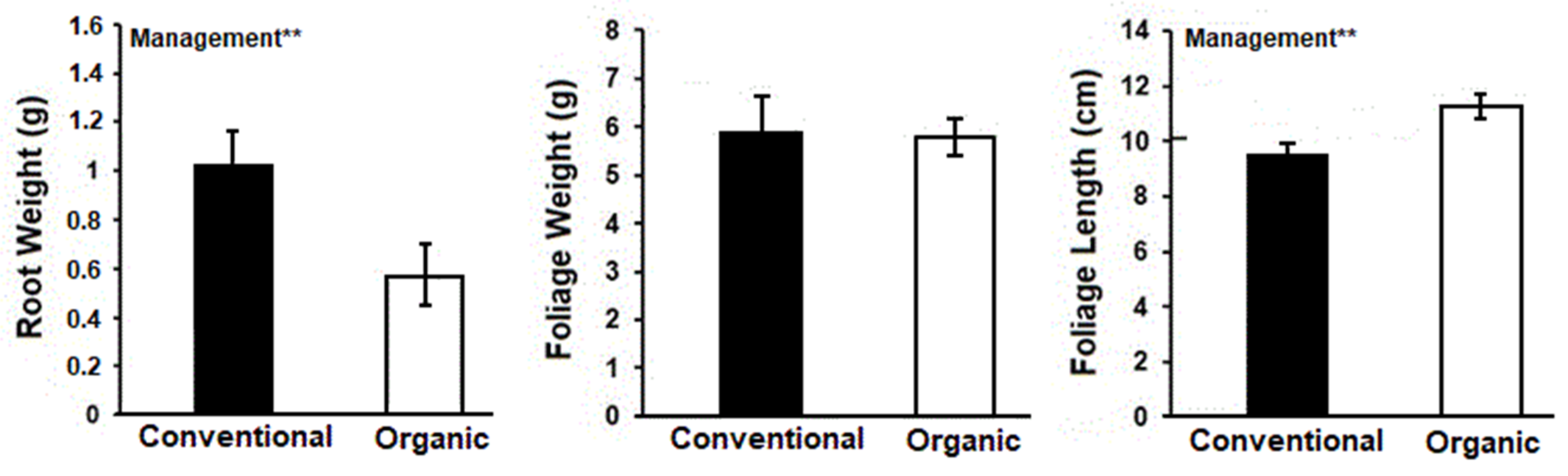
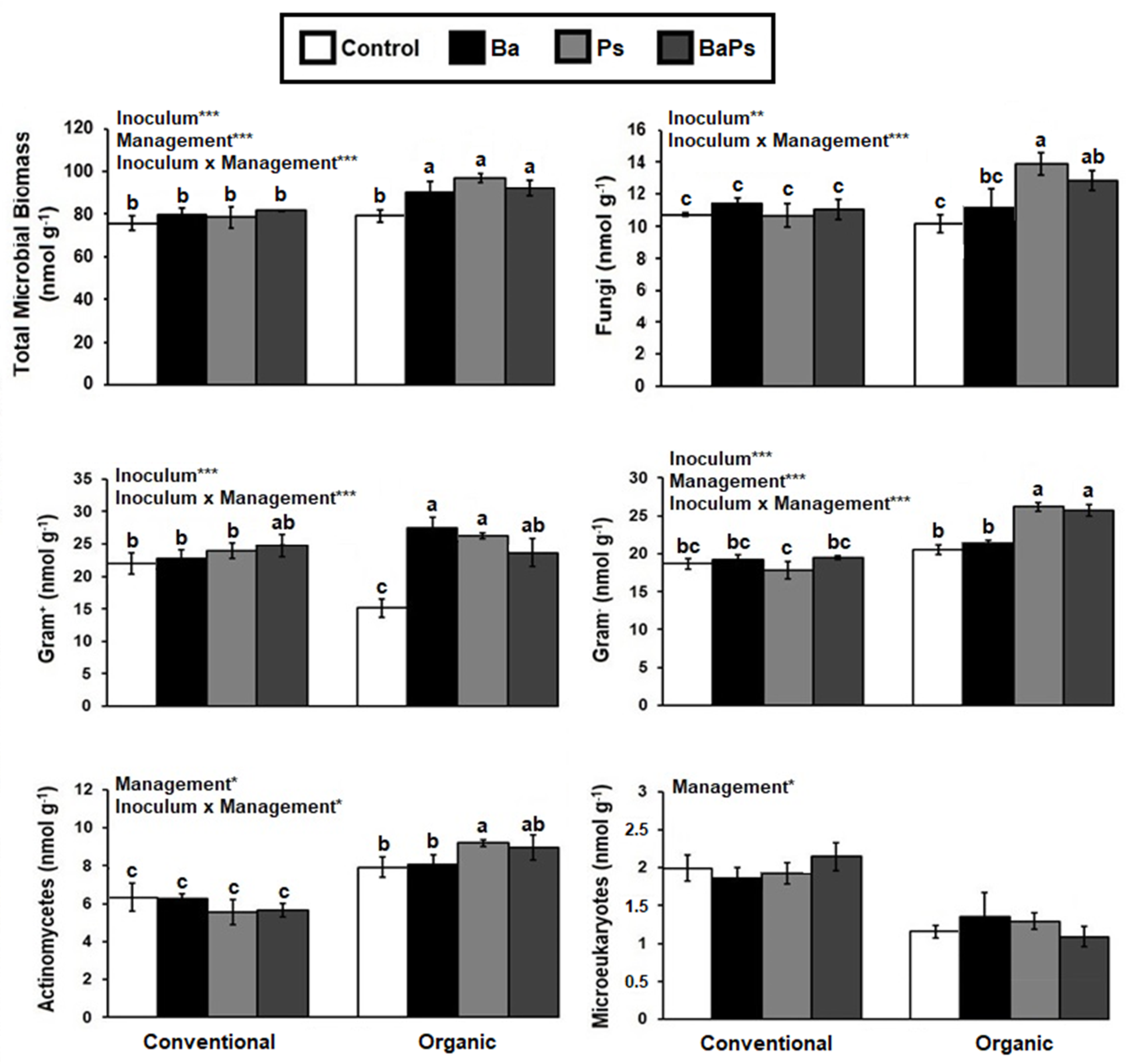
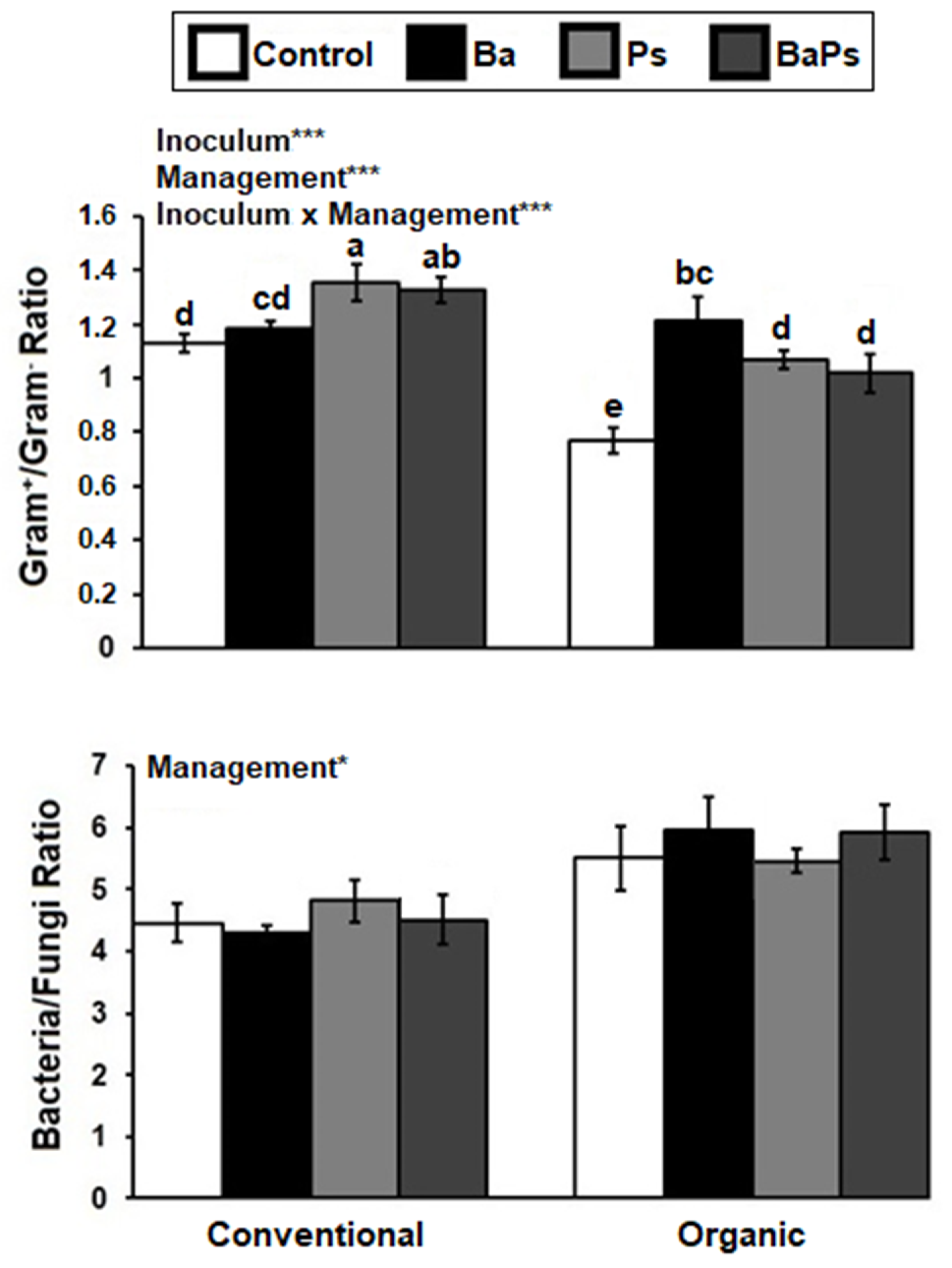
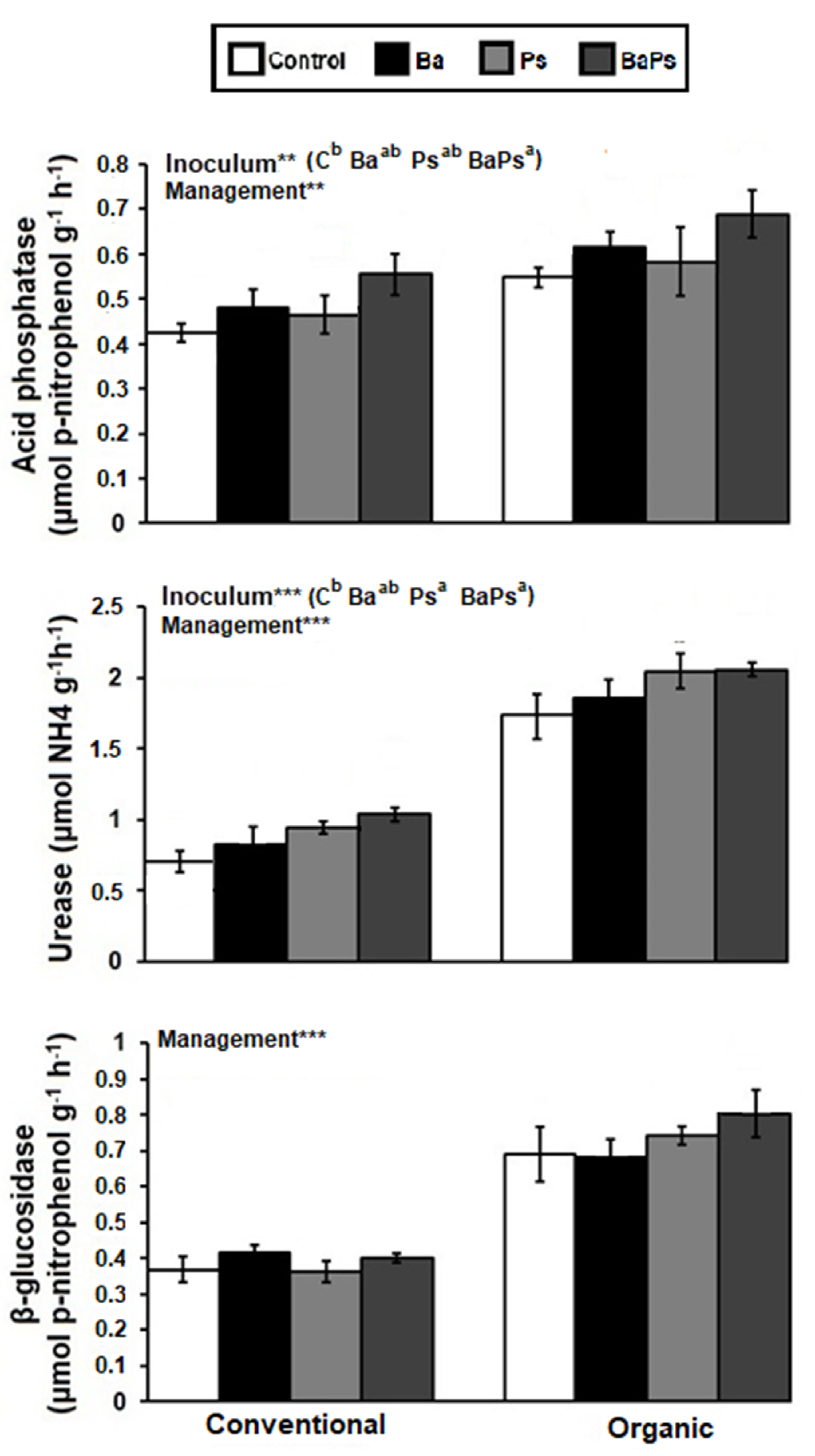
3. Results
4. Discussion
4.1. Plant Growth
4.2. Microbial Biomass, Composition, and Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tang, W.; Shan, B.; Zhang, H. Phosphorus buildup and release risk associated with agricultural intensification in the estuarine sediments of Chaohu Lake Valley, Eastern China. Clean Soil Air Water 2010, 38, 336–343. [Google Scholar] [CrossRef]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Farahani, E.; Kadner, S.; Seyboth, K.; Adier, A.; Baum, I.; Brunner, S.; Eickemeier, P. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambrige University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Laetz, C.A.; Baldwin, D.H.; Collier, T.K.; Hebert, V.; Stark, J.D.; Scholz, N.L. The synergistic toxicity of pesticide mixtures: Implications for risk assessment and the conservation of endangered Pacific salmon. Environ. Health Perspect. 2009, 117, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, R.; Sun, J.; Deng, F.; Wang, P.; Zhou, H.; Wei, Z.; Chen, M.; He, Z.; Lai, M.; Ye, T.; et al. Contamination of pyrethroids and atrazine in greenhouse and open-field agricultural soils in China. Sci. Total Environ. 2020, 701, 134916. [Google Scholar] [CrossRef] [PubMed]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. Response of soil microorganisms and enzymes to the foliar application of Helicur 250 EW fungicide on Horderum vulgare L. Chemosphere 2020, 242, 125163. [Google Scholar] [CrossRef]
- Satapute, P.; Kamble, M.V.; Adhikari, S.S.; Jogaiah, S. Influence of triazole pesticides on tillage soil microbial populations and metabolic changes. Sci. Total Environ. 2019, 651, 2334–2344. [Google Scholar] [CrossRef] [PubMed]
- Neuwirthová, N.; Trojan, M.; Svobodová, M.; Vašíčková, J.; Šimek, Z.; Hofman, J.; Bielská, L. Pesticide residues remaining in soils from previous growing season(s)—Can they accumulate in non-target organisms and contaminate the food web? Sci. Total Environ. 2019, 646, 1056–1062. [Google Scholar] [CrossRef]
- Birkhofer, K.; Bezemer, T.M.; Bloem, J.; Bonkowski, M.; Christensen, S.; Dubois, D.; Ekelund, F.; Fließbach, A.; Gunst, L.; Hedlund, K.; et al. Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biol. Biochem. 2008, 40, 2297–2308. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, W.; Liang, G.; Wang, X.; Sun, J.; He, P.; Li, L. Effects of different organic manures on the biochemical and microbial characteristics of albic paddy soil in a short-term experiment. PLoS ONE 2015, 10, e0124096. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ierna, A.; Mauromicale, G. Sustainable and profitable nitrogen fertilization management of potato. Agronomy 2019, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The role of microbial inoculants on plant protection, growth stimulation, and crop productivity of the olive tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Mirshad, P.P.; Puthur, J.T. Drought tolerance of bioenergy grass Saccharum spontaneum L. enhanced by arbuscular mycorrhizae. Rhizosphere 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Multifunctional Pseudomonas putida strain FBKV2 from arid rhizosphere soil and its growth promotional effects on maize under drought stress. Rhizosphere 2016, 1, 4–13. [Google Scholar] [CrossRef]
- Deshmukh, Y.; Khare, P.; Patra, D. Rhizobacteria elevate principal basmati aroma compound accumulation in rice variety. Rhizosphere 2016, 1, 53–57. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Barea, J.M. Future challenges and perspectives for applying microbial biotechnology in sustainable agriculture based on a better understanding of plant-microbiome interactions. J. Soil Sci. Plant Nutr. 2015, 15, 261–282. [Google Scholar] [CrossRef]
- Kamou, N.N.; Karasali, H.; Menexes, G.; Kasiotis, K.M.; Bon, M.C.; Papadakis, E.N.; Tzelepis, G.D.; Lotos, L.; Lagopodi, A.L. Isolation screening and characterisation of local beneficial rhizobacteria based upon their ability to suppress the growth of Fusarium oxysporum f. sp. radicis-lycopersici and tomato foot and root rot. Biocontrol Sci. Technol. 2015, 25, 928–949. [Google Scholar] [CrossRef]
- Ruiu, L. Microbial biopesticides in agroecosystems. Agronomy 2018, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Mishra, S.; Dixit, V.; Kumar, M.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Leveau, J.H.J.; Lindow, S.E. Utilization of the plant hormone Indole-3-Acetic Acid for growth by. Society 2005, 71, 2365–2371. [Google Scholar]
- Srivastava, S.; Bist, V.; Srivastava, S.; Singh, P.C.; Trivedi, P.K.; Asif, M.H.; Chauhan, P.S.; Nautiyal, C.S. Unraveling aspects of Bacillus amyloliquefaciens mediated enhanced production of rice under biotic stress of Rhizoctonia solani. Front. Plant Sci. 2016, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants—With special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Aini, N.; Yamika, W.S.D.; Ulum, B. Effect of nutrient concentration, PGPR and AMF on plant growth, yield and nutrient uptake of hydroponic lettuce. Int. J. Agric. Biol. 2019, 21, 175–183. [Google Scholar]
- Hsu, C.K.; Micallef, S.A. Plant-mediated restriction of Salmonella enterica on tomato and spinach leaves colonized with Pseudomonas plant growth-promoting rhizobacteria. Int. J. Food Microbiol. 2017, 259, 1–6. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cipriano, M.A.P.; Lupatini, M.; Lopes-Santos, L.; da Silva, M.J.; Roesch, L.F.W.; Destéfano, S.A.L.; Freitas, S.S.; Kuramae, E.E. Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucu, M.A.; Gilardi, G.; Pugliese, M.; Matić, S.; Gisi, U.; Gullino, M.L.; Garibaldi, A. Influence of different biological control agents and compost on total and nitrification-driven microbial communities at rhizosphere and soil level in a lettuce—Fusarium oxysporum f. sp. lactucae pathosystem. J. Appl. Microbiol. 2019, 126, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Castro-Sowinski, S.; Herschkovitz, Y.; Okon, Y.; Jurkevitch, E. Effects of inoculation with plant growth-promoting rhizobacteria on resident rhizosphere microorganisms. FEMS Microbiol. Lett. 2007, 276, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troelstra, S.R.; Wagenaar, R.; Smant, W.; Peters, B.A.M. Interpretation of bioassays in the study of interactions between soil organisms and plants: Involvement of nutrient factors. New Phytol. 2001, 150, 697–706. [Google Scholar] [CrossRef]
- Kapali, S.; Gade, R.M.; Shitole, A.V.; Swathi, S. Isolation and characterization of Pseudomonas fluorescens and Bacillus subtilis and their in vitro evaluation. Adv. Life Sci. 2016, 5, 5856–5859. [Google Scholar]
- Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupski, J.R. Versalovic_MMCB 1994_Genomic Fingerprinting.pdf. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Allen, E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publishing: Oxford, UK, 1976. [Google Scholar]
- Ntalli, N.; Monokrousos, N.; Rumbos, C.; Kontea, D.; Zioga, D.; Argyropoulou, M.D.; Menkissoglu-Spiroudi, U.; Tsiropoulos, N.G. Greenhouse biofumigation with Melia azedarach controls Meloidogyne spp. and enhances soil biological activity. J. Pest Sci. 2018, 91, 29–40. [Google Scholar] [CrossRef]
- Ntalli, N.; Menkissoglu-Spiroudi, U.; Doitsinis, K.; Kalomoiris, M.; Papadakis, E.N.; Boutsis, G.; Dimou, M.; Monokrousos, N. Mode of action and ecotoxicity of hexanoic and acetic acids on Meloidogyne javanica. J. Pest Sci. 2020, 93, 867–877. [Google Scholar] [CrossRef]
- Panayiotou, E.; Dimou, M.; Monokrousos, N. The effects of grazing intensity on soil processes in a Mediterranean protected area. Environ. Monit. Assess. 2017, 189, 441. [Google Scholar] [CrossRef]
- Allison, S.D.; Jastrow, J.D. Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol. Biochem. 2006, 38, 3245–3256. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Reynolds, H.; Long, T.M. Rapid assay for amidohydrolase (urease) activity in environmental samples. Soil Biol. Biochem. 2000, 32, 2095–2097. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. Poorter and Nagel 1999 The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 1191. [Google Scholar]
- Khosravi, A.; Zarei, M.; Ronaghi, A. Effect of PGPR, Phosphate sources and vermicompost on growth and nutrients uptake by lettuce in a calcareous soil. J. Plant Nutr. 2018, 41, 80–89. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Glaes, J.; Spaepen, S.; Bodson, B.; du Jardin, P.; Delaplace, P. Biostimulant effects of Bacillus strains on wheat from in vitro towards field conditions are modulated by nitrogen supply. J. Plant Nutr. Soil Sci. 2019, 182, 325–334. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Lynch, W.H. Effect of temperature on Pseudomonas fluorescens chemotaxis. J. Bacteriol. 1980, 143, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef]
- Thirup, L.; Ekelund, F.; Johnsen, K.; Jacobsen, C.S. Population dynamics of the fast-growing sub-populations of Pseudomonas and total bacteria, and their protozoan grazers, revealed by fenpropimorph treatment. Soil Biol. Biochem. 2000, 32, 1615–1623. [Google Scholar] [CrossRef]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef]
- Kozdrój, J.; Trevors, J.T.; Van Elsas, J.D. Influence of introduced potential biocontrol agents on maize seedling growth and bacterial community structure in the rhizosphere. Soil Biol. Biochem. 2004, 36, 1775–1784. [Google Scholar] [CrossRef]
- Han, L.; Wang, Z.; Li, N.; Wang, Y.; Feng, J.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- Zhang, L.; Lueders, T. Micropredator niche differentiation between bulk soil and rhizosphere of an agricultural soil depends on bacterial prey. FEMS Microbiol. Ecol. 2017, 93, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Viollet, A.; Pivato, B.; Mougel, C.; Cleyet-Marel, J.C.; Gubry-Rangin, C.; Lemanceau, P.; Mazurier, S. Pseudomonas fluorescens C7R12 type III secretion system impacts mycorrhization of Medicago truncatula and associated microbial communities. Mycorrhiza 2017, 27, 23–33. [Google Scholar] [CrossRef]
- El Arbi, A.; Rochex, A.; Chataigné, G.; Béchet, M.; Lecouturier, D.; Arnauld, S.; Gharsallah, N.; Jacques, P. The Tunisian oasis ecosystem is a source of antagonistic Bacillus spp. producing diverse antifungal lipopeptides. Res. Microbiol. 2016, 167, 46–57. [Google Scholar] [CrossRef]
- Moeskops, B.; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Nacamulli, C.; Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Chiarini, L. Perturbation of maize rhizosphere microflora following seed bacterization with Burkholderia cepacia MCI 7. FEMS Microbiol. Ecol. 1997, 23, 183–193. [Google Scholar] [CrossRef]
- Nunan, N.; Leloup, J.; Ruamps, L.S.; Pouteau, V.; Chenu, C. Effects of habitat constraints on soil microbial community function. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Fujita, M.; Nakashima, K.; Achal, V.; Kawasaki, S. Whole-cell evaluation of urease activity of Pararhodobacter sp. isolated from peripheral beachrock. Biochem. Eng. J. 2017, 124, 1–5. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
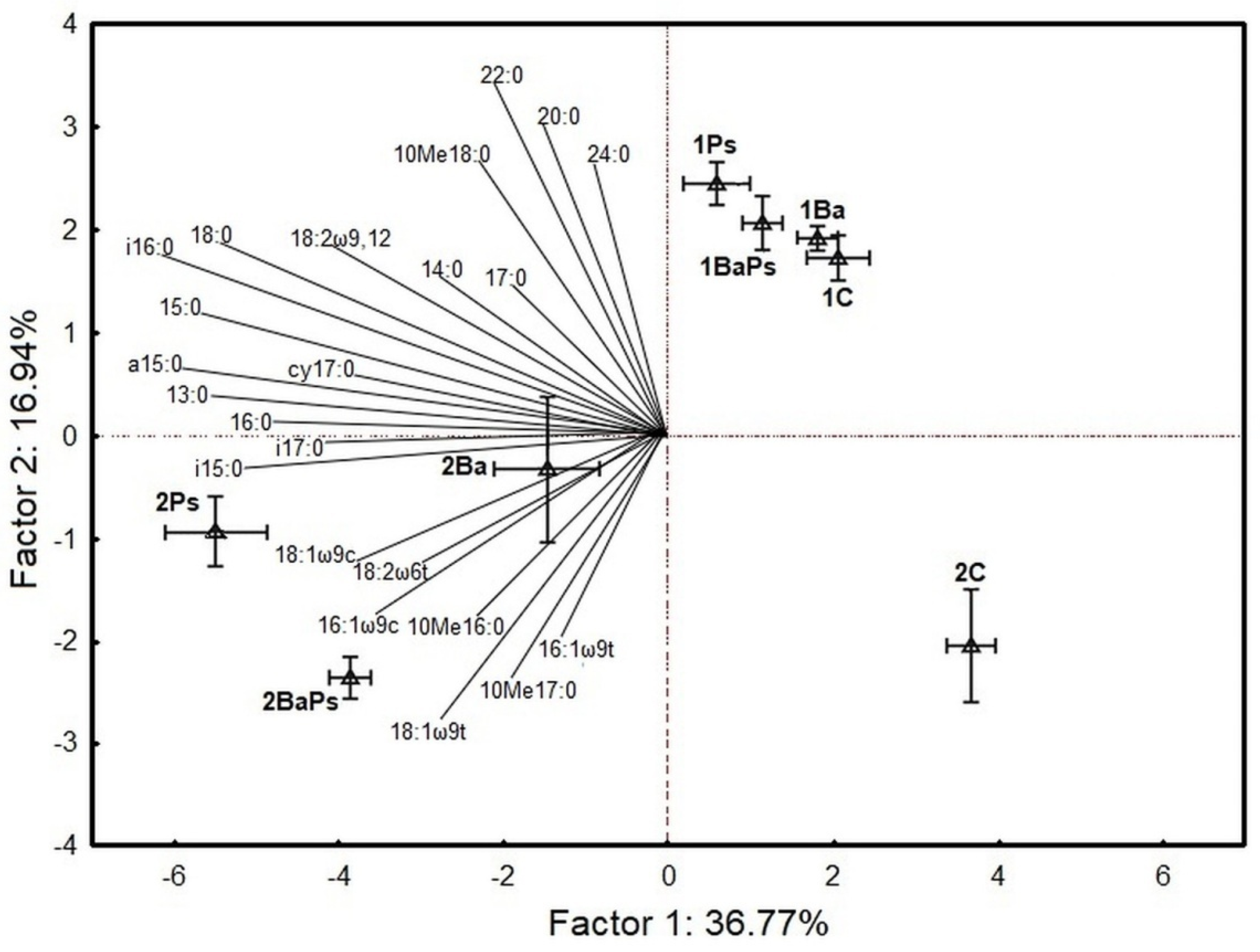{kind=link}
| Conventionally Managed System | Organically Managed System | p-Value | |
|---|---|---|---|
| pH | 6.60 ± 0.054 | 7.81 ± 0.035 | *** |
| EC (mS cm−1) | 0.93 ± 0.107 | 1.19 ± 0.049 | ns |
| Organic matter (%) | 2.29 ± 0.159 | 3.24 ± 0.351 | ** |
| Nitric Nitrogen (mg kg−1) | 14.13 ± 6.955 | 32.36 ± 2.377 | ** |
| Pext (mg kg−1) | 26.33 ± 0.609 | 133.75 ± 3.609 | *** |
| K (mg kg−1) | 153.66 ± 9.769 | 347.33 ± 10.170 | *** |
| Mg (mg kg−1) | 188.33 ± 7.264 | 352.33 ± 7.838 | *** |
| Zn (mg kg−1) | 5.64 ± 0.065 | 5.61 ± 0.196 | ns |
| Mn (mg kg−1) | 28.04 ± 2.256 | 26.40 ± 1.307 | ns |
| B (mg kg−1) | 0.84 ± 0.070 | 0.81 ± 0.051 | ns |
| CS-C | 0.174 | 0.116 | 0.024 | 0.030 | 0.027 | 0.026 | 0.061 | |
| CS-Ba | 0.174 | 0.318 | 0.284 | 0.029 | 0.061 | 0.030 | 0.027 | |
| CS-Ps | 0.116 | 0.318 | 0.055 | 0.030 | 0.028 | 0.030 | 0.027 | |
| CS-BaPs | 0.024 | 0.284 | 0.055 | 0.029 | 0.028 | 0.029 | 0.028 | |
| OS-C | 0.030 | 0.029 | 0.030 | 0.029 | 0.028 | 0.031 | 0.033 | |
| OS-Ba | 0.027 | 0.061 | 0.028 | 0.028 | 0.028 | 0.026 | 0.057 | |
| OS-Ps | 0.026 | 0.030 | 0.030 | 0.029 | 0.031 | 0.026 | 0.222 | |
| OS-BaPs | 0.061 | 0.027 | 0.027 | 0.028 | 0.033 | 0.057 | 0.222 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelina, E.; Papatheodorou, E.M.; Demirtzoglou, T.; Monokrousos, N. Effects of Bacillus subtilis and Pseudomonas fluorescens Inoculation on Attributes of the Lettuce (Lactuca sativa L.) Soil Rhizosphere Microbial Community: The Role of the Management System. Agronomy 2020, 10, 1428. https://doi.org/10.3390/agronomy10091428
Angelina E, Papatheodorou EM, Demirtzoglou T, Monokrousos N. Effects of Bacillus subtilis and Pseudomonas fluorescens Inoculation on Attributes of the Lettuce (Lactuca sativa L.) Soil Rhizosphere Microbial Community: The Role of the Management System. Agronomy. 2020; 10(9):1428. https://doi.org/10.3390/agronomy10091428
Chicago/Turabian StyleAngelina, Eirini, Efimia M. Papatheodorou, Triantafyllia Demirtzoglou, and Nikolaos Monokrousos. 2020. "Effects of Bacillus subtilis and Pseudomonas fluorescens Inoculation on Attributes of the Lettuce (Lactuca sativa L.) Soil Rhizosphere Microbial Community: The Role of the Management System" Agronomy 10, no. 9: 1428. https://doi.org/10.3390/agronomy10091428