Stand Structural Diversity and Species with Leaf Nitrogen Conservation Drive Aboveground Carbon Storage in Tropical Old-Growth Forests

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
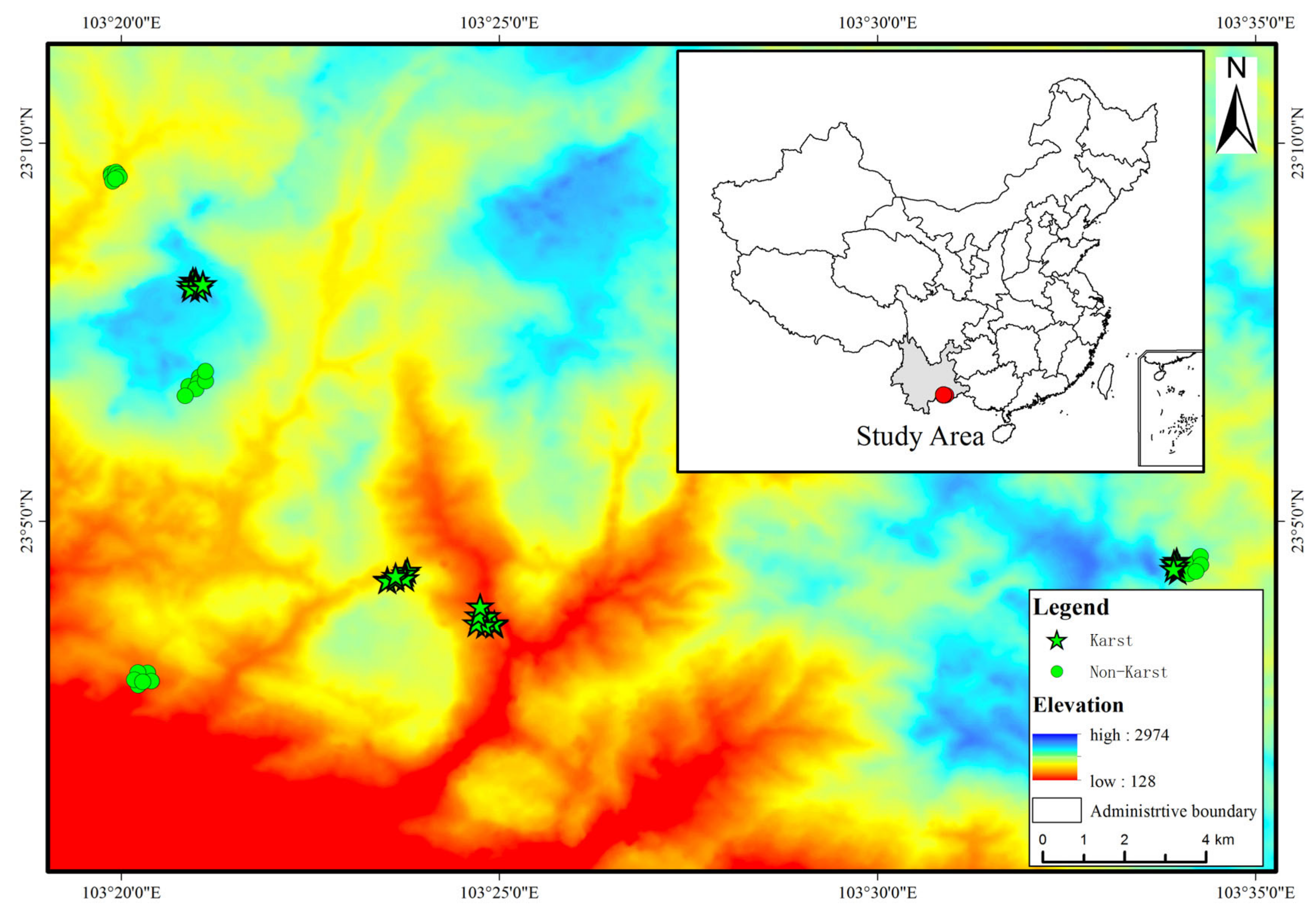
2.1. Site Description and Plots
2.2. Quantification of Aboveground C Storage
2.3. Quantification of Stand Structural and Species Diversity
2.4. Functional Traits
2.5. Statistical Analysis
3. Results
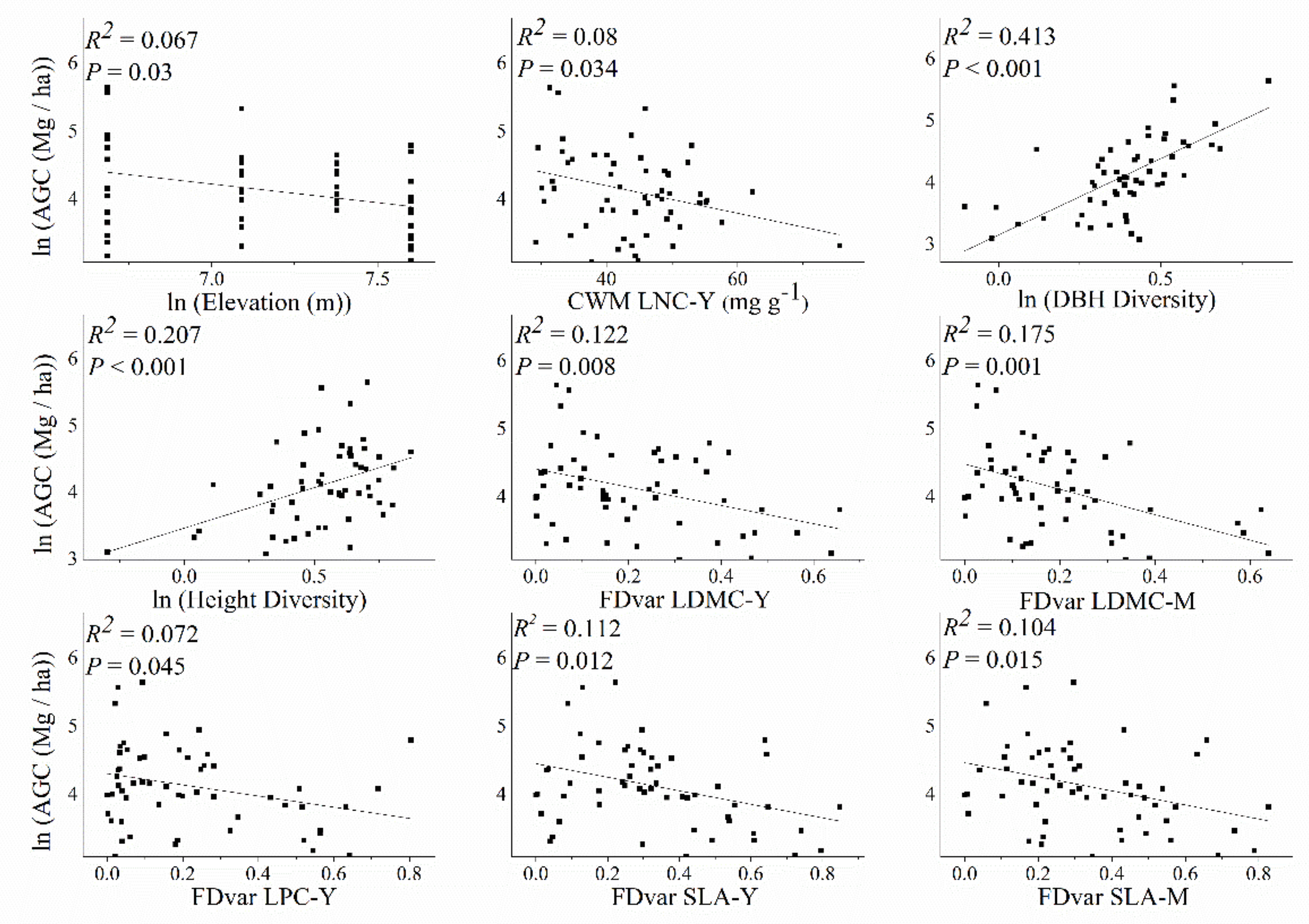
3.1. Effects of Influencing Factors on Aboveground C Storage
3.2. Relative Importance of all Predictor Variables for Aboveground C Storage
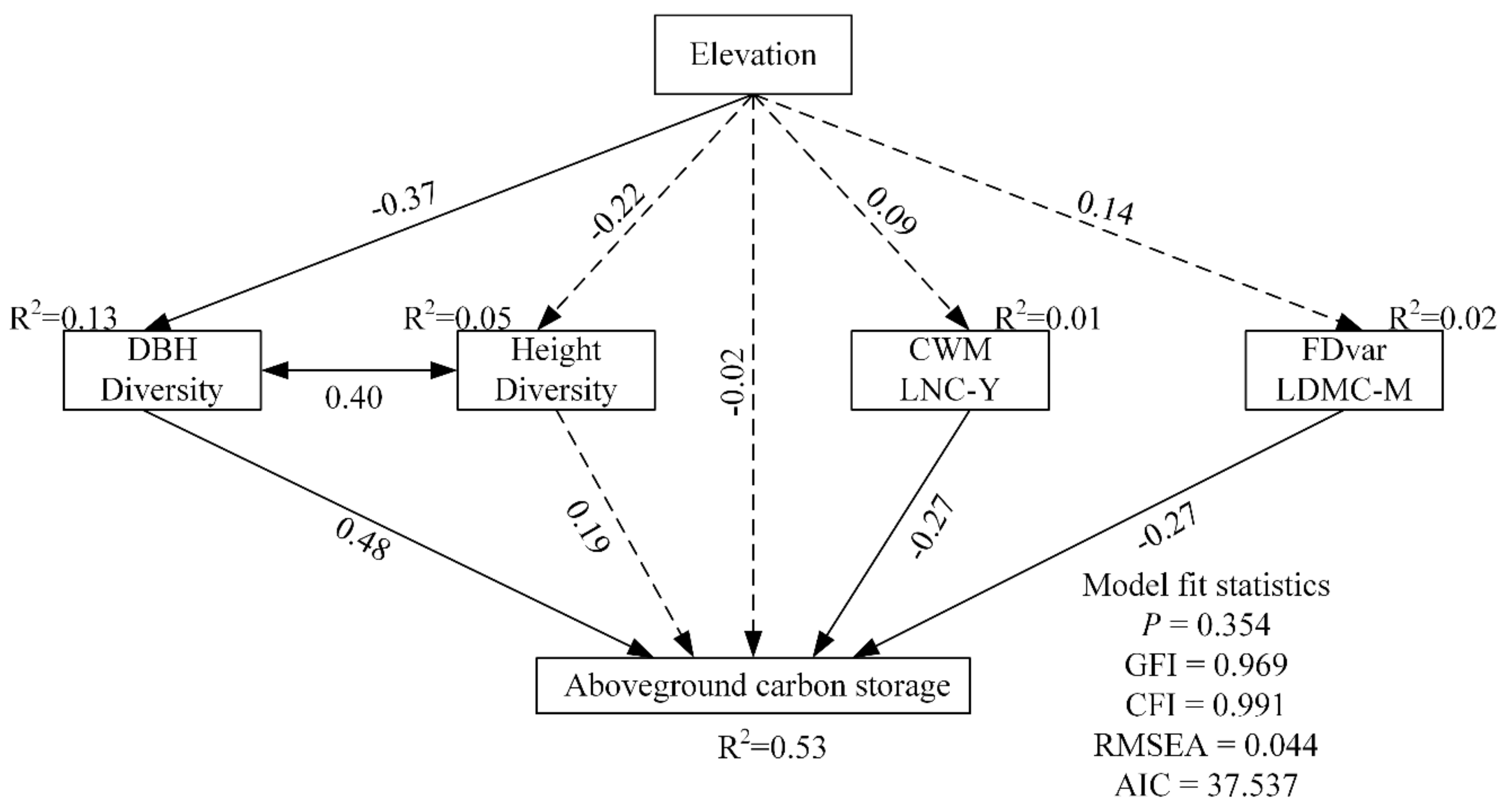
3.3. Direct and Indirect Impacts on Aboveground C Storage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohbeck, M.; Bongers, F.; Martinez-Ramos, M.; Poorter, L. The importance of biodiversity and dominance for multiple ecosystem functions in a human-modified tropical landscape. Ecology 2016, 97, 2772–2779. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Q.; Ali, A.; Wang, S.P.; Gazol, A.; Freckleton, R.; Wang, X.G.; Lin, F.; Zhou, L.; Hao, Z.Q.; Loreau, M. Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests. Sci. Total Environ. 2018, 630, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bao, W.; Bongers, F.; Chen, B.; Chen, G.K.; Guo, K.; Jiang, M.X.; Lai, J.S.; Lin, D.M.; Liu, C.J.; et al. Drivers of tree carbon storage in subtropical forests. Sci. Total Environ. 2019, 654, 684–693. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Cavard, X.; Bergeron, Y.; Chen, H.Y.H.; Pare, D. Mixed-species effect on tree aboveground carbon pools in the east-central boreal forests. Can. J. For. Res. 2010, 40, 37–47. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Fotis, A.T.; Murphy, S.J.; Ricart, R.D.; Krishnadas, M.; Whitacre, J.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Above-ground biomass is driven by mass-ratio effects and stand structural attributes in a temperate deciduous forest. J. Ecol. 2018, 106, 561–571. [Google Scholar] [CrossRef]
- Ali, A.; Yan, E.R.; Chang, S.X.; Cheng, J.Y.; Liu, X.Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Sci. Total Environ. 2017, 574, 654–662. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Quétier, F.; Cáceres, D.M.; Trainor, S.F.; Pérez-Harguindeguy, N.; Bret-Harte, M.S.; Finegan, B.; Peña-Claros, M.; Poorter, L. Linking functional diversity and social actor strategies in a framework for interdisciplinary analysis of nature’s benefits to society. Proc. Natl. Acad. Sci. USA 2011, 108, 895–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, S.; Lavorel, S.; de Bello, F.; Quetier, F.; Grigulis, K.; Robson, M. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. USA 2007, 104, 20684–20689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage—An empirical test in semi-arid forest ecosystems. J. Ecol. 2013, 101, 18–28. [Google Scholar] [CrossRef]
- Prado-Junior, J.A.; Schiavini, I.; Vale, V.S.; Arantes, C.S.; van der Sande, M.T.; Lohbeck, M.; Poorter, L. Conservative species drive biomass productivity in tropical dry forests. J. Ecol. 2016, 104, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; et al. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Glob. Ecol. Biogeogr. 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Lin, D.; Anderson-Teixeira, K.J.; Lai, J.; Mi, X.; Ren, H.; Ma, K. Traits of dominant tree species predict local scale variation in forest aboveground and topsoil carbon stocks. Plant Soil 2016, 409, 435–446. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H. Individual size inequality links forest diversity and aboveground biomass. J. Ecol. 2015, 103, 1245–1252. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities. Ecol. Lett. 2007, 10, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.M.; Spasojevic, M.J.; Muller-Landau, H.C.; Sun, I.F.; Lin, Y.; Sus, S.H.; Chen, Z.; Chen, C.T.; Swenson, N.G.; McEwan, R.W. Functional composition drives ecosystem function through multiple mechanisms in a broadleaved subtropical forest. Oecologia 2016, 182, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Yan, E.R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.T.; Yang, X.D.; Xu, M.S. Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef] [Green Version]
- Dǎnescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 1–15. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; et al. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.J.; Audino, L.D.; Whitacre, J.; Eck, J.L.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Species associations structured by environment and land-use history promote beta-diversity in a temperate forest. Ecology 2015, 96, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Fahey, R.T.; Fotis, A.T.; Woods, K.D. Quantifying canopy complexity and effects on productivity and resilience in late successional hemlock-hardwood forests. Ecol. Appl. 2015, 25, 834–847. [Google Scholar] [CrossRef]
- Jiang, Z.C.; Lian, Y.Q.; Qin, X.Q. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Wang, B.; Yang, S.; Lu, C.; Zhang, J.; Wang, Y. Comparison of net primary productivity in karst and non-karst areas: A case study in Guizhou Province, China. Environ. Earth Sci. 2010, 59, 1337–1347. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Q.Y.; Du, F. Flora diversity characteristics of seed plants in Dawei mountain national nature reserve in Yunnan province, China. Sci. Silvae Sin. 2005, 42, 7–15. [Google Scholar]
- Chave, J.; Rejou-Mechain, M.; Burquez, A.; Chidumayo, E.; Colgan, M.; Delitti, W.; Duque, A.; Eid, T.; Fearnside, P.; Goodman, R.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Finegan, B.; Peña-Claros, M.; Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreño-Rocabado, G.; Casanoves, F.; Díaz, S.; Eguiguren Velepucha, P.; Fernandez, F.; et al. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. J. Ecol. 2015, 103, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Q.; Wang, S.P.; Ali, A.; Gazol, A.; Ruiz-Benito, P.; Wang, X.G.; Lin, F.; Ye, J.; Hao, Z.Q.; Loreau, M. Aboveground carbon storage is driven by functional trait composition and stand structural attributes rather than biodiversity in temperate mixed forests recovering from disturbances. Ann. For. Sci. 2018, 75. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Díaz, S.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.; Fitter, A.; Raffaelli, D. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Wen, Z.; Zheng, H.R.; Smith, J.; Zhao, H.; Liu, L.; Ouyang, Z.Y. Functional diversity overrides community-weighted mean traits in linking land-use intensity to hydrological ecosystem services. Sci. Total Environ. 2019, 682, 583–590. [Google Scholar] [CrossRef]
- Graham, M. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Calcagno, V. glmulti: Model Selection and Multimodel Inference Made Easy; R Package Version 1.0.7.1; 2013. Available online: http://CRAN.R-project.org/package=glmulti, (accessed on 15 August 2013).
- Van der Sande, M.T.; Peña-Claros, M.; Ascarrunz, N.; Arets, E.J.M.M.; Licona, J.C.; Toledo, M.; Poorter, L. Abiotic and biotic drivers of biomass change in a Neotropical forest. J. Ecol. 2017, 105, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O′Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [Green Version]
- Tobner, C.M.; Paquette, A.; Gravel, D.; Reich, P.B.; Williams, L.J.; Messier, C. Functional identity is the main driver of diversity effects in young tree communities. Ecol. Lett. 2016, 19, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Competitive exclusion in herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Seabloom, E.W.; Borer, E.T.; Adler, P.B.; Harpole, W.S.; Hautier, Y.; Hillebrand, H.; Lind, E.M.; Partel, M.; et al. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 2016, 529, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Brown, S.; Lugo, A.E. Effects of changes in biodiversity on ecosystem function in tropical forests. Conserv. Biol. 1996, 10, 17–24. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Keren, S.; Medarević, M.; Obradović, S.; Zlokapa, B. Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe. Forests 2018, 9, 479. [Google Scholar] [CrossRef] [Green Version]
- Keren, S.; Svoboda, M.; Janda, P.; Nagel, T.A. Relationships between Structural Indices and Conventional Stand Attributes in an Old-Growth Forest in Southeast Europe. Forests 2020, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Van Gelder, H.A.; Poorter, L.; Sterck, F.J. Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytol. 2006, 171, 367–378. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billes, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Quesada, C.A.; Phillips, O.L.; Schwarz, M.; Czimczik, C.I.; Baker, T.R.; Pati o, S.; Fylla, N.M.; Hodnett, M.G.; Herrera, R.; Almeida, S.; et al. Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate. Biogeosciences 2012, 9, 2203–2246. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [PubMed]
- Brant, A.N.; Chen, H.Y.H. Patterns and Mechanisms of Nutrient Resorption in Plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, W.H.; Yu, M.K.; Wang, G.G.; Wu, T.G. Latitudinal patterns of leaf N, P stoichiometry and nutrient resorption of Metasequoia glyptostroboides along the eastern coastline of China. Sci. Total Environ. 2018, 618, 1–6. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Read, Q.D.; Moorhead, L.C.; Swenson, N.G.; Bailey, J.K.; Sanders, N.J. Convergent effects of elevation on functional leaf traits within and among species. Funct. Ecol. 2014. [Google Scholar] [CrossRef]
- He, N.P.; Liu, C.C.; Tian, M.; Li, M.L.; Yang, H.; Yu, G.R.; Guo, D.L.; Smith, M.D.; Yu, Q.; Hou, J.H. Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions. Funct. Ecol. 2017, 00, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, R.A.; Muller-Landau, H.C.; Abdul Rahman, K.; Bebber, D.P.; Bin, Y.; Bohlman, S.A.; Bourg, N.A.; Brinks, J.; Bunyavejchewin, S.; Butt, N.; et al. Scale-dependent relationships between tree species richness and ecosystem function in forests. J. Ecol. 2013. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation | 800 m | 1200 m | 1600 m | 2000 m | ||||
|---|---|---|---|---|---|---|---|---|
| Region Types | Karst | Non-Karst | Karst | Non-Karst | Karst | Non-Karst | Karst | Non-Karst |
| Soil type | Red soils | Brown soils | Red soils | Brown soils | Red soils | Brown soils | Red soils | Brown soils |
| Slope (°) | 25 | 27 | 42 | 35 | 27 | 30 | 36 | 40 |
| Rock bare rate (%) | 70 | 0 | 70 | 0 | 45 | 0 | 55 | 0 |
| Height (m) | 8.77 (3.5, 22.8) | 9.78 (4.1, 23.8) | 9.08 (3.6, 18.2) | 14.57 (3.4, 25.5) | 8.34 (3.1, 29) | 8.18 (3.2, 25) | 6.38 (4.2, 18.9) | 11.08 (4, 20) |
| DBH (cm) | 13.25 (5, 116.1) | 15.42 (5, 220) | 22.03 (4.8, 82.1) | 24.19 (5, 166.3) | 10.48 (5, 76.2) | 9.11 (4.9, 163.8) | 8.56 (4.9, 111.5) | 21.79 (4.9, 170) |
| Density (plants/hm2) | 989 (765, 1083) | 833 (698, 936) | 533 (324, 725) | 322 (278, 365) | 1500 (1263, 1786) | 1622 (1359, 1827) | 811 (524, 1254) | 411 (316, 625) |
| DBH Class (cm) | AIC | R2 | p-Value | Height Class (m) | AIC | R2 | p-Value |
|---|---|---|---|---|---|---|---|
| 2 | 105.84 | 0.168 | 0.002 | 2 | 105.84 | 0.207 | <0.001 |
| 4 | 89.37 | 0.38 | <0.001 | 3 | 107.16 | 0.175 | 0.001 |
| 6 | 87.27 | 0.402 | <0.001 | 4 | 108.14 | 0.133 | 0.006 |
| 8 | 85.04 | 0.413 | <0.001 | 5 | 106.13 | 0.195 | <0.001 |
| Model and Predictor | Coeff. | Beta | t | p | R2 | AICc |
|---|---|---|---|---|---|---|
| Effects of community-weighted mean (CWM) | ||||||
| Model1 | <0.001 | 0.28 | 105.52 | |||
| Constant | 9.20 | 0 | 7.21 | <0.001 | ||
| CWM SLA-Y | −0.01 | −0.39 | −2.33 | 0.024 | ||
| CWM LT-Y | −2.66 | −0.46 | −3.21 | 0.002 | ||
| CWM LNC-Y | −0.02 | −0.34 | −2.70 | 0.009 | ||
| CWM LDMC-M | −5.35 | −0.44 | −0.28 | 0.007 | ||
| Effects of functional diversity (FDvar), species diversity, and stand structural diversity | ||||||
| Model2 | <0.001 | 0.51 | 80.80 | |||
| Constant | 3.35 | 0 | 14.36 | <0.001 | ||
| FDvar LDMC-M | −1.17 | −0.27 | −2.59 | 0.012 | ||
| DBH Diversity | 1.85 | 0.49 | 4.24 | <0.001 | ||
| Height Diversity | 0.53 | 0.17 | 1.60 | 0.116 | ||
| Joint effect of all predicted variables | ||||||
| Model3 | <0.001 | 0.57 | 76.18 | |||
| Constant | 4.14 | 0 | 11.15 | <0.001 | ||
| CWM LNC-Y | −0.02 | −0.25 | −2.65 | 0.011 | ||
| FDvar LDMC-M | −1.16 | −0.27 | −2.73 | 0.009 | ||
| DBH Diversity | 1.78 | 0.47 | 4.30 | <0.001 | ||
| Height Diversity | 0.56 | 0.18 | 1.78 | 0.081 | ||
| Predictor | Pathway to Aboveground Carbon Storage | Effect | p-Value |
|---|---|---|---|
| Elevation | Direct effect | −0.02 | 0.906 |
| Indirect effect via DBH diversity | −0.18 | 0.002 | |
| Indirect effect via height diversity | −0.04 | 0.057 | |
| Indirect effect via CWM LNC-Y | −0.02 | 0.412 | |
| Indirect effect via FDvar LDMC-M | −0.04 | 0.245 | |
| Total effect | −0.30 | 0.044 | |
| DBH Diversity | Direct effect | 0.48 | <0.001 |
| Height Diversity | Direct effect | 0.19 | 0.057 |
| CWM LNC-Y | Direct effect | −0.27 | 0.003 |
| FDvar LDMC-M | Direct effect | −0.27 | 0.004 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Liu, Y.; Wu, X.; Pang, D.; Yang, X.; Hussain, A.; Zhou, J. Stand Structural Diversity and Species with Leaf Nitrogen Conservation Drive Aboveground Carbon Storage in Tropical Old-Growth Forests. Forests 2020, 11, 994. https://doi.org/10.3390/f11090994
Wang G, Liu Y, Wu X, Pang D, Yang X, Hussain A, Zhou J. Stand Structural Diversity and Species with Leaf Nitrogen Conservation Drive Aboveground Carbon Storage in Tropical Old-Growth Forests. Forests. 2020; 11(9):994. https://doi.org/10.3390/f11090994
Chicago/Turabian StyleWang, Genzhu, Yuguo Liu, Xiuqin Wu, Danbo Pang, Xiao Yang, Azfar Hussain, and Jinxing Zhou. 2020. "Stand Structural Diversity and Species with Leaf Nitrogen Conservation Drive Aboveground Carbon Storage in Tropical Old-Growth Forests" Forests 11, no. 9: 994. https://doi.org/10.3390/f11090994