Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia tabaci MEAM1

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
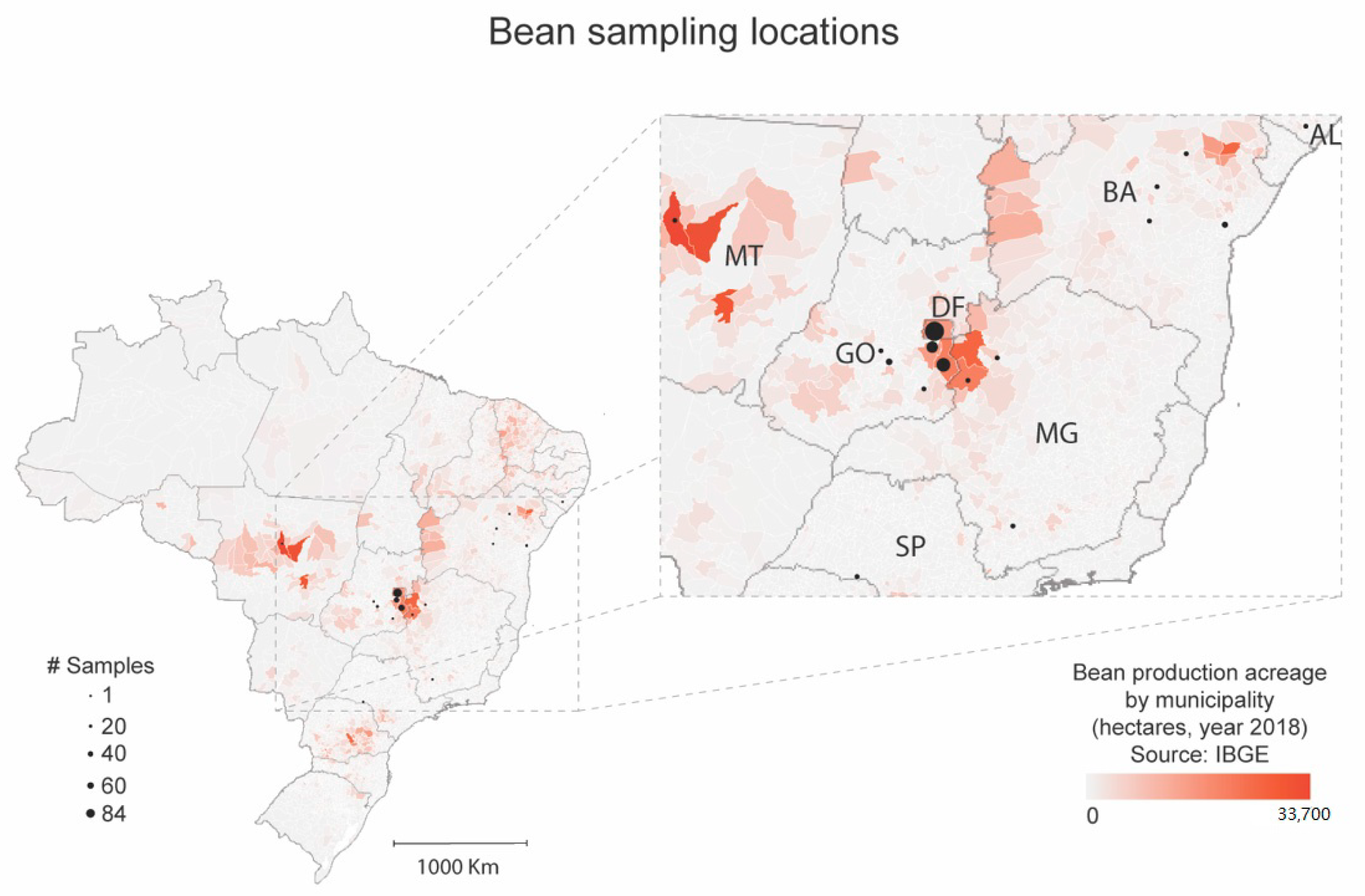
2.2. Distribution of BaCV in Common Beans in Brazil
2.3. RNA and DNA Extraction
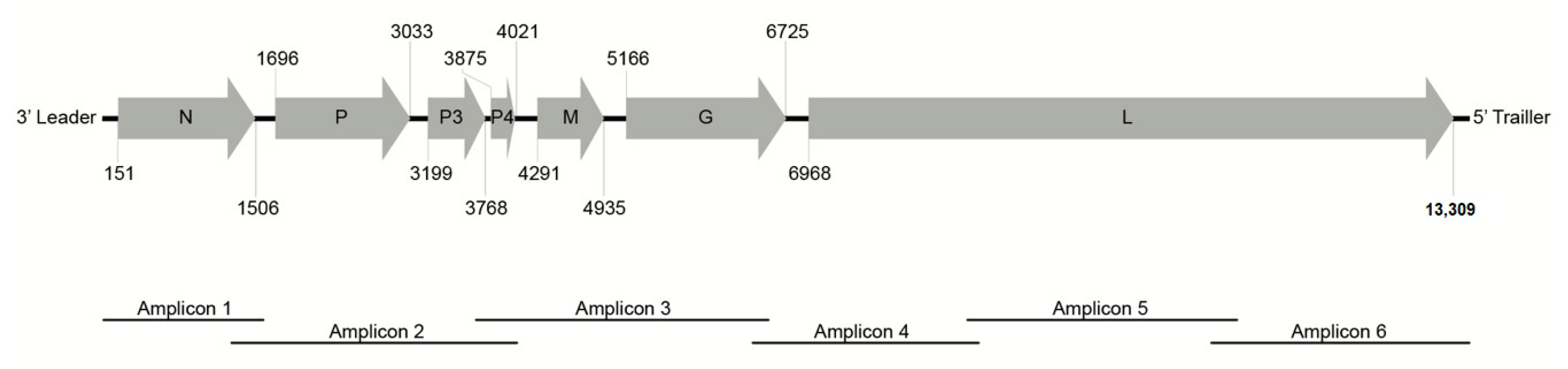
2.4. RT-PCR, PCR, and Cloning
2.5. Sequence Analysis
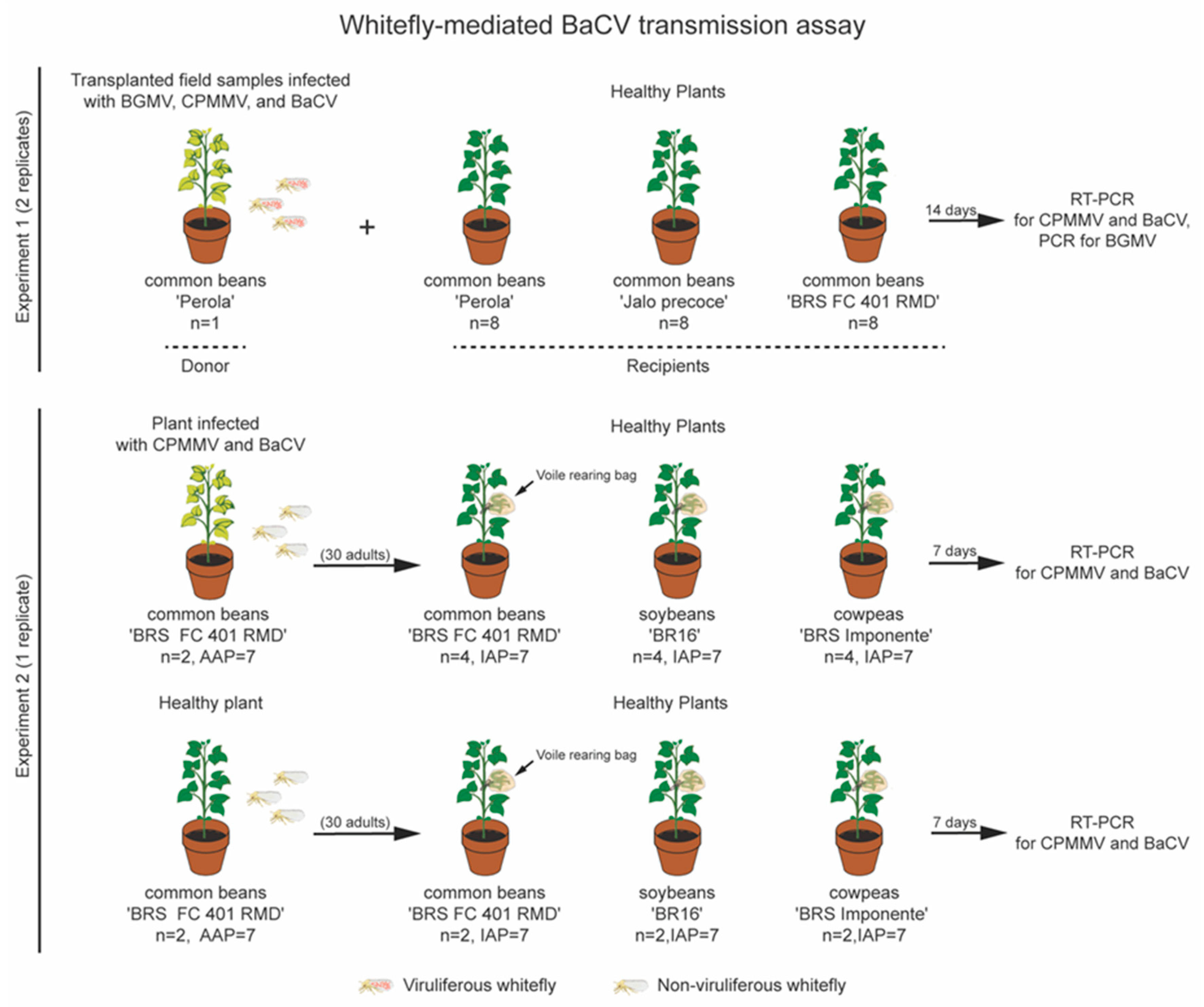
2.6. BaCV Transmission by B. tabaci MEAM1
2.7. BaCV Detection in Plants and Whiteflies
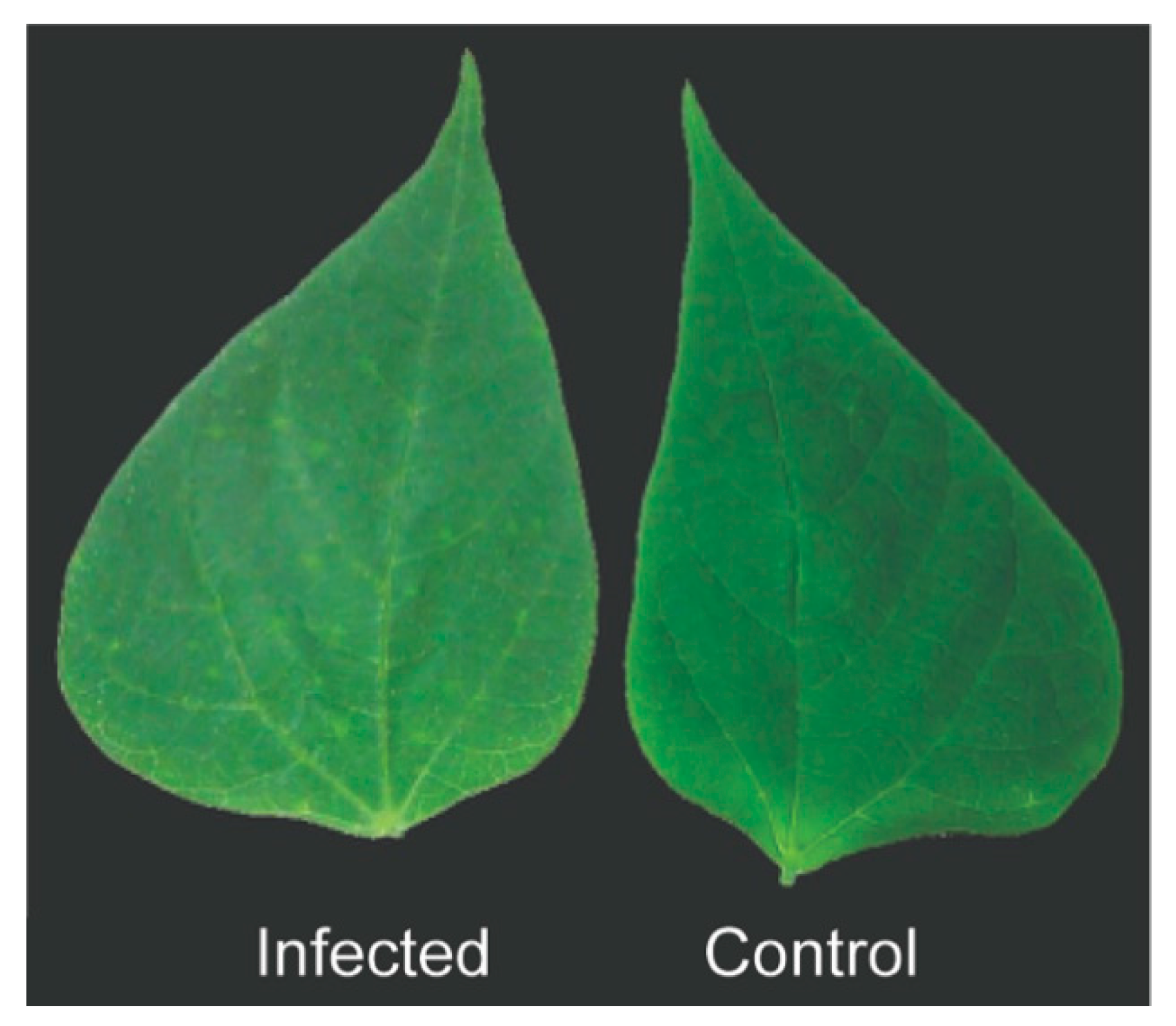
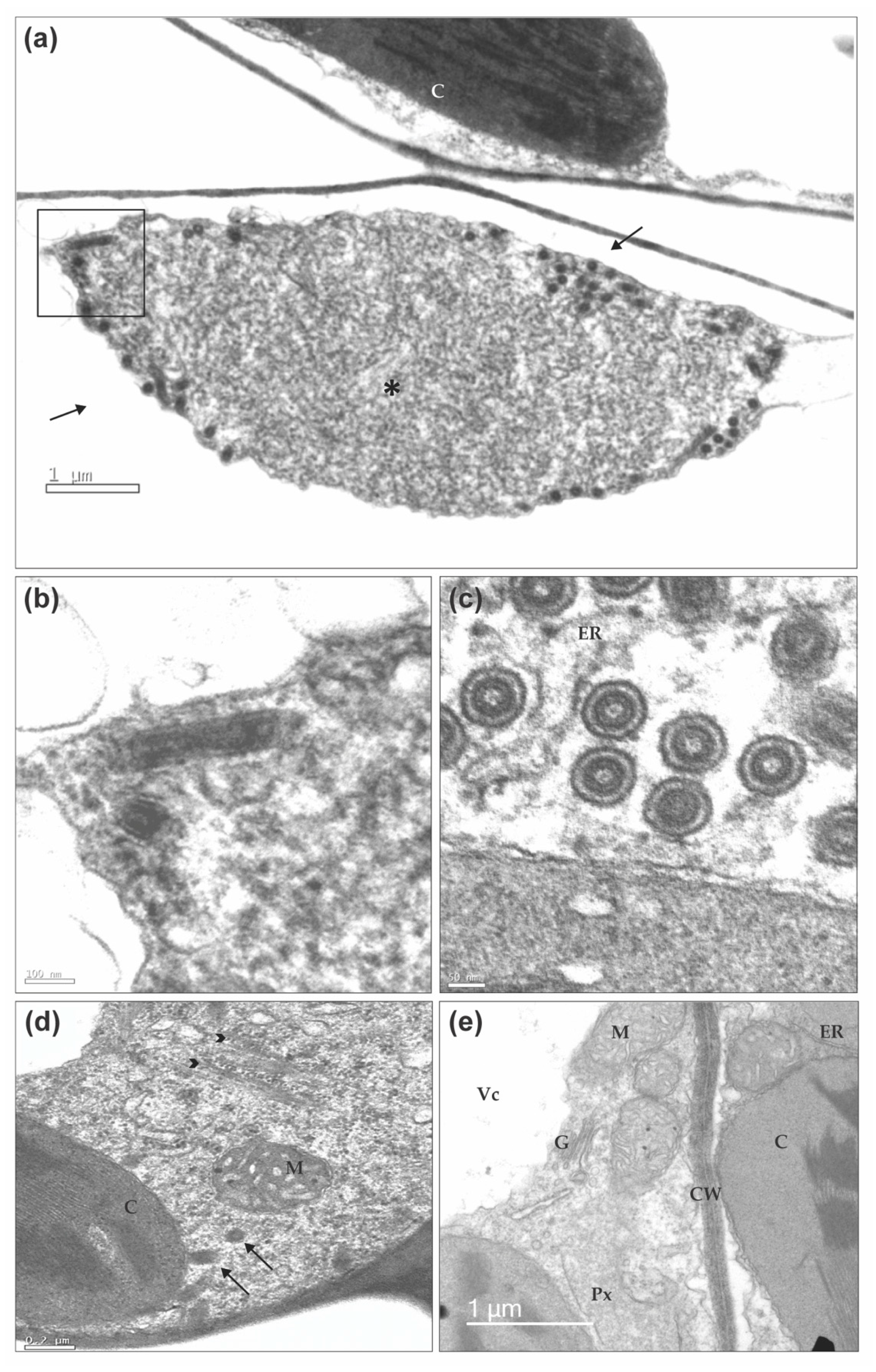
2.8. Transmission Electron Microscopy
3. Results and Discussion
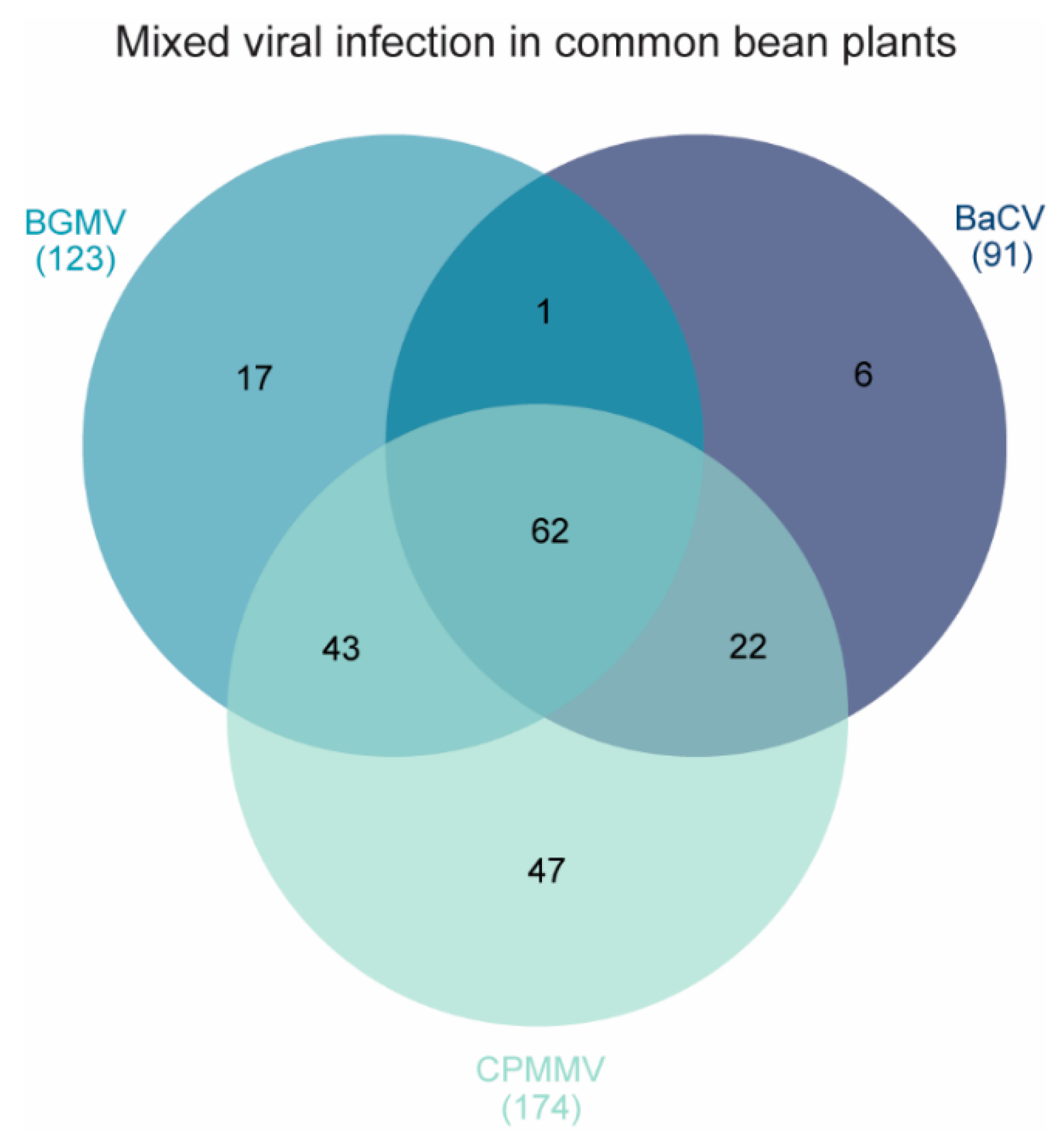
3.1. BaCV Is Widely Distributed throughout Common Bean Producing Areas in Brazil
3.2. Evolutionary Analysis Indicates Whiteflies as the Potential Vector for BaCV
3.3. B. tabaci MEAM1 Transmit BaCV-Luz to Common Beans, Cowpea, and Soybean
3.4. BaCV-Luz Detection in Plants and Whiteflies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Freitas-Astua, J.; Dietzgen, R.G.; Walker, P.J.; Blasdell, K.R.; Breyta, R.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; Stone, D.M.; et al. Split the genus Nucleorhabdovirus, creating three new genera (Alphanucleorhabdovirus, Betanucleorhabdovirus and Gammanucleorhabdovirus) comprising sixteen species, including six new species, in the family Rhabdoviridae. Approved ICTV Proposal. 2019. Available online: https://ictv.global/ICTV/proposals/2019.031M.zip (accessed on 18 May 2020).
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef]
- Ammar, E.D.; Tsai, C.W.; Whitfield, A.E.; Redinbaugh, M.G.; Hogenhout, S.A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annu. Rev. Entomol. 2009, 54, 447–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietzgen, R.G.; Kondo, H.; Goodin, M.M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: Mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus Res. 2017, 227, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, A.E.; Huot, O.B.; Martin, K.M.; Kondo, H.; Dietzgen, R.G. Plant rhabdoviruses-their origins and vector interactions. Curr. Opin. Virol. 2018, 33, 198–207. [Google Scholar] [CrossRef]
- Alves-Freitas, D.M.T.; Pinheiro-Lima, B.; Faria, J.C.; Lacorte, C.; Ribeiro, S.G.; Melo, F.L. Double-Stranded RNA High-Throughput Sequencing Reveals a New Cytorhabdovirus in a Bean Golden Mosaic Virus-Resistant Common Bean Transgenic Line. Viruses 2019, 11, 90. [Google Scholar] [CrossRef] [Green Version]
- Medina-Salguero, A.X.; Cornejo-Franco, J.F.; Grinstead, S.; Mollov, D.; Mowery, J.D.; Flores, F.; Quito-Avila, D.F. Sequencing, genome analysis and prevalence of a cytorhabdovirus discovered in Carica papaya. PLoS ONE 2019, 14, e0215798. [Google Scholar] [CrossRef]
- Freitas-Astua, J.; Dietzgen, R.G.; Walker, P.J.; Blasdell, K.R.; Breyta, R.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; Stone, D.M.; et al. Create twelve new species in the genus Cytorhabdovirus, family Rhabdoviridae. Approved ICTV Proposal. 2019. Available online: https://ictv.global/ICTV/proposals/2019.030M.zip (accessed on 18 May 2020).
- International Committee on Taxonomy of Viruses—ICTV Online. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/negative-sense-rna-viruses/mononegavirales/w/rhabdoviridae (accessed on 18 May 2020).
- Instituto Brasileiro de Geografia e Estatística—IBGE. Available online: https://www.ibge.gov.br (accessed on 24 July 2020).
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Nicolini, C.; Pio-Ribeiro, G.; Andrade, G.P.; Melo, F.L.; Oliveira, V.C.; Guimaraes, F.C.; Resende, R.O.; Kitajima, E.W.; Rezende, J.A.; Nagata, T. A distinct tymovirus infecting Cassia hoffmannseggii in Brazil. Virus Genes 2012, 45, 190–194. [Google Scholar] [CrossRef]
- Schuster, D.; Buchman, G.; Rashtchian, A. A simple and efficient method for amplification of cDNA ends using 5′ RACE. Focus 1992, 14, 46–52. [Google Scholar]
- Pinheiro-Lima, B.; Alves-Freitas, D.M.T.; Melo, F.L.; Carvalho, R.; de Faria, J.C.; Ribeiro, S.G. Characterization of a new whitefly-transmitted (Bemisia tabaci MEAM 1) cytorhabdovirus infecting common bean. Congr. Nac. Pesqui. Feijão 2017, 12, 52. [Google Scholar]
- Lamas, N.S.; Matos, V.O.R.L.; Alves-Freitas, D.M.T.; Melo, F.L.; Costa, A.F.; Faria, J.C.; Ribeiro, S.G. Occurrence of Cowpea mild mottle virus in Common Bean and Associated Weeds in Northeastern Brazil. Plant Dis. 2017, 101, 1828. [Google Scholar] [CrossRef]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.; Mendes, E.A.; Aragao, F.J. RNAi-mediated resistance to Bean golden mosaic virus in genetically engineered common bean (Phaseolus vulgaris). Mol. Plant Microbe Interact 2007, 20, 717–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco, D.; Loria, A.; Sartor, C.; Cenis, J. PCR-RFLP Identification of Bemisia tabaci biotypes in the Mediterranean Basin. Phytoparasitica 2006, 34, 243–251. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Shimodaira, H. An approximately unbiased test of phylogenetic tree selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Gerlt, J.A.; Bouvier, J.T.; Davidson, D.B.; Imker, H.J.; Sadkhin, B.; Slater, D.R.; Whalen, K.L. Enzyme Function Initiative-Enzyme Similarity Tool (EFI-EST): A web tool for generating protein sequence similarity networks. Biochim. Biophys. Acta 2015, 1854, 1019–1037. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Faria, J.C.; Aragão, F.J.L.; Souza, T.L.P.O.; Quintela, E.D.; Kitajima, E.W.; Ribeiro, S.G. Golden mosaic of common beans in Brazil: Management with a transgenic approach. APS Features 2016, 10, 1–14. [Google Scholar]
- Orfanidou, C.G.; Pappi, P.G.; Efthimiou, K.E.; Katis, N.I.; Maliogka, V.I. Transmission of Tomato chlorosis virus (ToCV) by Bemisia tabaci Biotype Q and Evaluation of Four Weed Species as Viral Sources. Plant Dis. 2016, 100, 2043–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.; Xu, W.Y.; Gao, Q.; Jiang, Z.H.; Liu, S.Y.; Fang, X.D.; Gao, D.M.; Wang, Y.; Wang, X.B. Transmission Characteristics of Barley Yellow Striate Mosaic Virus in Its Planthopper Vector Laodelphax striatellus. Front. Microbiol. 2018, 9, 1419. [Google Scholar] [CrossRef]
- Borges, A.; Tsai, S.M.; Caldas, D.G. Validation of reference genes for RT-qPCR normalization in common bean during biotic and abiotic stresses. Plant Cell Rep. 2012, 31, 827–838. [Google Scholar] [CrossRef]
- Knight, M.R.; Jenkins, G.I. Genes encoding the small subunit of ribulose 1,5-bisphosphate carboxylase/oxygenase in Phaseolus vulgaris L.: Nucleotide sequence of cDNA clones and initial studies of expression. Plant Mol. Biol. 1992, 18, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Chandrashekar, K.; Thakur, N.; Verma, P.C.; Borgio, J.F.; Singh, P.K.; Tuli, R. RNA interference for the control of whiteflies (Bemisia tabaci) by oral route. J. Biosci. 2011, 36, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Alves-Freitas, D.M.T.; Melo, F.L.; Faria, J.C.; Ribeiro, S.G. dsRNA deep sequencing reveals five viral species in common beans. J. Braz. Soc. Virol. 2016, 20, 43–44. [Google Scholar]
- Moreno, A.B.; Lopez-Moya, J.J. When Viruses Play Team Sports: Mixed Infections in Plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-virus interactions in a plant host and in a hemipteran vector: Implications for vector fitness and virus epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef] [PubMed]
- Francki, R.I.B.; Randles, J.W.; Dietzgen, R.G. Lettuce necrotic yellows virus. AAB Descr. Plant Viruses 1989, 343, 5. [Google Scholar]
- Jones, S.; McGavin, W.; MacFarlane, S. The complete sequences of two divergent variants of the rhabdovirus raspberry vein chlorosis virus and the design of improved primers for virus detection. Virus Res. 2019, 265, 162–165. [Google Scholar] [CrossRef]
- Sylvester, E.S.; Richardson, J.; Frazier, N.W. Serial passage of strawberry crinkle virus in the aphid Chaetosiphon jacobi. Virology 1974, 59, 301–306. [Google Scholar] [CrossRef]
- Franova, J.; Pribylova, J.; Koloniuk, I. Molecular and Biological Characterization of a New Strawberry Cytorhabdovirus. Viruses 2019, 11, 982. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Appiano, A. Barley yellow striate mosaic virus and associated viroplasms in barley cells. J. Gen. Virol. 1973, 21, 315–322. [Google Scholar] [CrossRef]
- Gollifer, D.E.; Jackson, G.V.H.; Dabek, A.J.; Plumb, R.T.; May, Y.Y. The occurrence and transmission of viruses of edible aroids in the Solomon Islands and the Southwest Pacific. Pestic. Artic. News Summ. 1977, 23, 171–177. [Google Scholar] [CrossRef]
- Maurino, M.F.; Laguna, G.; Giolitti, F.; Nome, C.; Pecci, M.P.G. First Ocurrence of a Rhabdovirus Infecting Maize in Argentina. Plant Dis. 2012, 96, 1383. [Google Scholar] [CrossRef] [PubMed]
- Maurino, F.; Dumon, A.D.; Llauger, G.; Alemandri, V.; de Haro, L.A.; Mattio, M.F.; Del Vas, M.; Laguna, I.G.; Gimenez Pecci, M.P. Complete genome sequence of maize yellow striate virus, a new cytorhabdovirus infecting maize and wheat crops in Argentina. Arch. Virol. 2018, 163, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Toriyama, S. Purification and some properties of northern cereal mosaic virus. Uirusu 1972, 22, 114–124. [Google Scholar] [CrossRef]
- Toriyama, S. Northern cereal mosaic. In Viruses and Virus Diseases of Poaceae (Gramineae); Lapierre, H., Signoret, P.-A., Eds.; INRA: Paris, France, 2004; pp. 578–580. [Google Scholar]
- Yang, X.; Huang, J.; Liu, C.; Chen, B.; Zhang, T.; Zhou, G. Rice Stripe Mosaic Virus, a Novel Cytorhabdovirus Infecting Rice via Leafhopper Transmission. Front. Microbiol. 2016, 7, 2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejerman, N.; Dietzgen, R.G. Letter to the Editor: Bean-Associated Cytorhabdovirus and Papaya Cytorhabdovirus are Strains of the Same Virus. Viruses 2019, 11, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietzgen, R.G.; Bejerman, N.E.; Goodin, M.M.; Higgins, C.M.; Huot, O.B.; Kondo, H.; Martin, K.M.; Whitfield, A.E. Diversity and epidemiology of plant rhabdoviruses. Virus Res. 2020, 281, 197942. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.S.; Dietzgen, R.G. Plant rhabdoviruses: New insights and research needs in the interplay of negative-strand RNA viruses with plant and insect hosts. Arch. Virol. 2014, 159, 1889–1900. [Google Scholar] [CrossRef]
- de Moraes, L.A.; Muller, C.; Bueno, R.; Santos, A.; Bello, V.H.; De Marchi, B.R.; Watanabe, L.F.M.; Marubayashi, J.M.; Santos, B.R.; Yuki, V.A.; et al. Distribution and phylogenetics of whiteflies and their endosymbiont relationships after the Mediterranean species invasion in Brazil. Sci. Rep. 2018, 8, 14589. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.L.; Nagata, T.; Junqueira, B.R.; Zanardo, L.G.; Paiva, A.C.; Carvalho, C.M. Construction of a full-length infectious cDNA clone of Cowpea mild mottle virus. Virus Genes 2017, 53, 137–140. [Google Scholar] [CrossRef]
- Zanardo, L.G.; Carvalho, C.M. Cowpea mild mottle virus (Carlavirus, Betaflexiviridae): A review. Trop. Plant Pathol. 2017, 42, 417–430. [Google Scholar] [CrossRef]
- Fernandes, F.R.; Cruz, A.R.; Faria, J.C.; Zerbini, F.M.; Aragao, F.J. Three distinct begomoviruses associated with soybean in central Brazil. Arch. Virol. 2009, 154, 1567–1570. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Lee, W.; Park, J.; Lee, G.S.; Lee, S.; Akimoto, S. Taxonomic status of the Bemisia tabaci complex (Hemiptera: Aleyrodidae) and reassessment of the number of its constituent species. PLoS ONE 2013, 8, e63817. [Google Scholar] [CrossRef] [Green Version]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the Insect Supervectors Bemisia tabaci and Frankliniella occidentalis in the Emergence and Global Spread of Plant Viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olive, E.; Pan, L.L.; Liu, S.S.; Navas-Castillo, J. Transmission of Begomoviruses and Other Whitefly-Borne Viruses: Dependence on the Vector Species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Kanakala, S.; Lebedev, G.; Kontsedalov, S.; Silverman, D.; Alon, T.; Mor, N.; Sela, N.; Luria, N.; Dombrovsky, A.; et al. Transmission of a New Polerovirus Infecting Pepper by the Whitefly Bemisia tabaci. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, T.M.; Inoue-Nagata, A.K.; Vidal, A.H.; Ribeiro, S.G.; Nagata, T. The recombinant isolate of cucurbit aphid-borne yellows virus from Brazil is a polerovirus transmitted by whiteflies. Plant Pathol. 2020, 69, 1042–1050. [Google Scholar] [CrossRef]
- Cornejo-Franco, J.F.; Alvarez-Quinto, R.A.; Quito-Avila, D.F. Transmission of the umbra-like Papaya virus Q in Ecuador and its association with meleira-related viruses from Brazil. Crop Prot. 2018, 110, 99–102. [Google Scholar] [CrossRef]
- Martins, D.S.; Lima, A.F.; Fornazier, M.J.; Barcellos, B.D.; Queiroz, R.B.; Fanton, C.J.; Zanuncio-Junior, J.S.; Fornazier, D.L. Whiteflies (Hemiptera: Aleyrodidae) Associated with papaya (Carica papaya L.). Rev. Científica Intelletto 2016, 2, 78–86. [Google Scholar]
- Sinisterra, X.H.; McKenzie, C.L.; Hunter, W.B.; Powell, C.A.; Shatters, R.G. Differential transcriptional activity of plant-pathogenic begomoviruses in their whitefly vector (Bemisia tabaci, Gennadius: Hemiptera Aleyrodidae). J. Gen. Virol. 2005, 86, 1525–1532. [Google Scholar] [CrossRef]
- Pakkianathan, B.C.; Kontsedalov, S.; Lebedev, G.; Mahadav, A.; Zeidan, M.; Czosnek, H.; Ghanim, M. Replication of Tomato Yellow Leaf Curl Virus in Its Whitefly Vector, Bemisia tabaci. J. Virol. 2015, 89, 9791–9803. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.L.; Wang, X.R.; Wei, X.M.; Huang, H.; Wu, J.X.; Chen, X.X.; Liu, S.S.; Wang, X.W. The autophagy pathway participates in resistance to tomato yellow leaf curl virus infection in whiteflies. Autophagy 2016, 12, 1560–1574. [Google Scholar] [CrossRef] [Green Version]
- Czosnek, H.; Ghanim, M.; Morin, S.; Rubinstein, G.; Fridman, V.; Zeidan, M. Whiteflies: Vectors, and victims (?), of geminiviruses. Adv. Virus Res. 2001, 57, 291–322. [Google Scholar] [CrossRef]
- He, Y.Z.; Wang, Y.M.; Yin, T.Y.; Fiallo-Olive, E.; Liu, Y.Q.; Hanley-Bowdoin, L.; Wang, X.W. A plant DNA virus replicates in the salivary glands of its insect vector via recruitment of host DNA synthesis machinery. Proc. Natl. Acad. Sci. USA 2020, 117, 16928–16937. [Google Scholar] [CrossRef]
- Jackson, A.O.; Dietzgen, R.G.; Goodin, M.M.; Bragg, J.N.; Deng, M. Biology of plant rhabdoviruses. Annu. Rev. Phytopathol. 2005, 43, 623–660. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.O.; Dietzgen, R.G.; Goodin, M.M.; Li, Z. Development of Model Systems for Plant Rhabdovirus Research. Adv. Virus Res. 2018, 102, 23–57. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.D.; Hogenhout, S.A. A neurotropic route for Maize mosaic virus (Rhabdoviridae) in its planthopper vector Peregrinus maidis. Virus Res. 2008, 131, 77–85. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Sharma, S.; Singh, H.; Dixit, S.; Kumar, J.; Verma, P.C.; Chandrashekar, K. Whitefly genome expression reveals host-symbiont interaction in amino acid biosynthesis. PLoS ONE 2015, 10, e0126751. [Google Scholar] [CrossRef] [Green Version]
- Mwaipopo, B.; Nchimbi-Msolla, S.; Njau, P.J.R.; Mark, D.; Mbanzibwa, D.R. Comprehensive Surveys of Bean common mosaic virus and Bean common mosaic necrosis virus and Molecular Evidence for Occurrence of Other Phaseolus vulgaris Viruses in Tanzania. Plant Dis. 2018, 102, 2361–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Huang, A.; Zhou, X.; Li, Z.; Dietzgen, R.G.; Zhou, C.Y.; Cao, M. Natural defect of a plant rhabdovirus glycoprotein gene: A case study of virus-plant co-evolution. Phytopathology 2020. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State/Total Number of Samples | City | Sampled Areas | Year | Virus Incidence (Infected/Tested) | ||
|---|---|---|---|---|---|---|
| BaCV | CPMMV | BGMV | ||||
| Bahia (BA) (n = 17) | Cruz das Almas | 1 | 2015 | 2/8 | 8/8 | 4/8 |
| Morro do Chapéu | 1 | 2015 | 0/4 | 3/4 | 0/4 | |
| Piritiba | 1 | 2015 | 0/3 | 3/3 | 0/3 | |
| Antônio Gonçalves | 1 | 2015 | 0/2 | 2/2 | 0/2 | |
| Total in Bahia | 2/17 | 16/17 | 4/17 | |||
| Alagoas (AL) (n = 5) | Arapiraca | 1 | 2018 | 0/5 | 4/5 | 1/5 |
| Total in Alagoas | 0/5 | 4/5 | 1/5 | |||
| Minas Gerais (MG) (n = 9) | Bonfinópolis de Minas | 1 | 2016 | 0/3 | 3/3 | 0/3 |
| Paracatu | 1 | 2016 | 1/1 | 1/1 | 1/1 | |
| Três Pontas | 1 | 2018 | 0/5 | 1/5 | 2/5 | |
| Total in Minas Gerais | 1/9 | 5/9 | 3/9 | |||
| São Paulo (SP) (n = 1) | Palmital | 1 | 2015 | 1/1 | 1/1 | 1/1 |
| Total in São Paulo | 1/1 | 1/1 | 1/1 | |||
| Goiás (GO) (n = 101) | Luziânia | 1 | 2016 | 15/15 | 15/15 | 15/15 |
| 2 | 2016 | 7/7 | 7/7 | 7/7 | ||
| 3 | 2016 | 3/4 | 4/4 | 4/4 | ||
| 4 | 2016 | 4/4 | 4/4 | 4/4 | ||
| Cristalina | 1 | 2016 | 10/14 | 14/14 | 14/14 | |
| 2 | 2016 | 0/14 | 12/14 | 13/14 | ||
| 3 | 2016 | 11/11 | 10/11 | 11/11 | ||
| 4 | 2016 | 4/4 | 4/4 | 4/4 | ||
| Santo Antônio de Goiás | 1 | 2016 | 0/4 | 4/4 | 4/4 | |
| 2 | 2016 | 2/17 | 17/17 | 8/8 | ||
| 1 | 2018 | 0/5 | 4/5 | 4/5 | ||
| Urutaí | 1 | 2018 | 0/1 | 0/1 | 0/1 | |
| Araçu | 1 | 2018 | 0/1 | 1/1 | 1/1 | |
| Total in Goiás | 56/101 | 96/101 | 89/92 | |||
| Distrito Federal (DF) (n = 84) | Brasília | 1 | 2017 | 8/30 | 19/30 | 14/30 |
| 1 | 2007 | 0/2 | 0/2 | 2/2 | ||
| 1 | 2012 | 0/11 | 1/11 | 7/11 | ||
| 1 | 2015 | 8/12 | 6/12 | - | ||
| 2 | 2015 | 15/29 | 25/29 | - | ||
| Total in Distrito Federal | 31/84 | 51/84 | 23/43 | |||
| Mato Grosso (MT) (n = 2) | Sorriso | 1 | 2018 | 0/2 | 1/2 | 2/2 |
| Total in Mato Grosso | 0/2 | 1/2 | 2/2 | |||
| TOTAL | 91/219 | 174/219 | 123/169 | |||
| Plant | Cultivar | Positive/Total | Transmission Rate (%) |
|---|---|---|---|
| P. vulgaris (common bean) | ‘BRSFC 401 RMD’ | 3/4 | 75 |
| V. unguiculata (cowpea) | ‘BRS Imponente’ | 2/4 | 50 |
| G. max (soybean) | ‘BR16′ | 1/4 | 25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro-Lima, B.; Pereira-Carvalho, R.C.; Alves-Freitas, D.M.T.; Kitajima, E.W.; Vidal, A.H.; Lacorte, C.; Godinho, M.T.; Fontenele, R.S.; Faria, J.C.; Abreu, E.F.M.; et al. Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia tabaci MEAM1. Viruses 2020, 12, 1028. https://doi.org/10.3390/v12091028
Pinheiro-Lima B, Pereira-Carvalho RC, Alves-Freitas DMT, Kitajima EW, Vidal AH, Lacorte C, Godinho MT, Fontenele RS, Faria JC, Abreu EFM, et al. Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia tabaci MEAM1. Viruses. 2020; 12(9):1028. https://doi.org/10.3390/v12091028
Chicago/Turabian StylePinheiro-Lima, Bruna, Rita C. Pereira-Carvalho, Dione M. T. Alves-Freitas, Elliot W. Kitajima, Andreza H. Vidal, Cristiano Lacorte, Marcio T. Godinho, Rafaela S. Fontenele, Josias C. Faria, Emanuel F. M. Abreu, and et al. 2020. "Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia tabaci MEAM1" Viruses 12, no. 9: 1028. https://doi.org/10.3390/v12091028