Ten-Year Estimation of Net Primary Productivity in a Mangrove Forest under a Tropical Monsoon Climate in Eastern Thailand: Significance of the Temperature Environment in the Dry Season

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
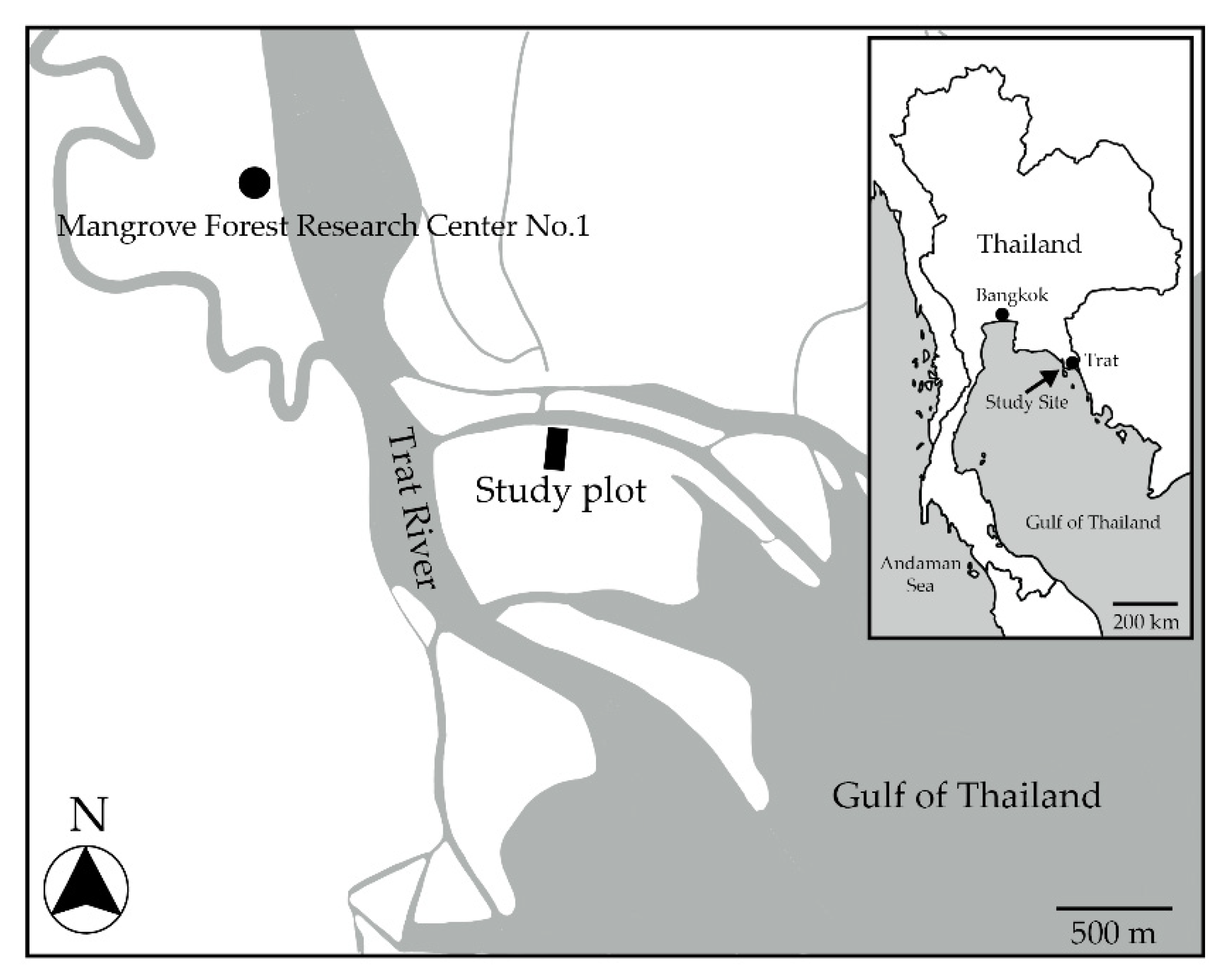
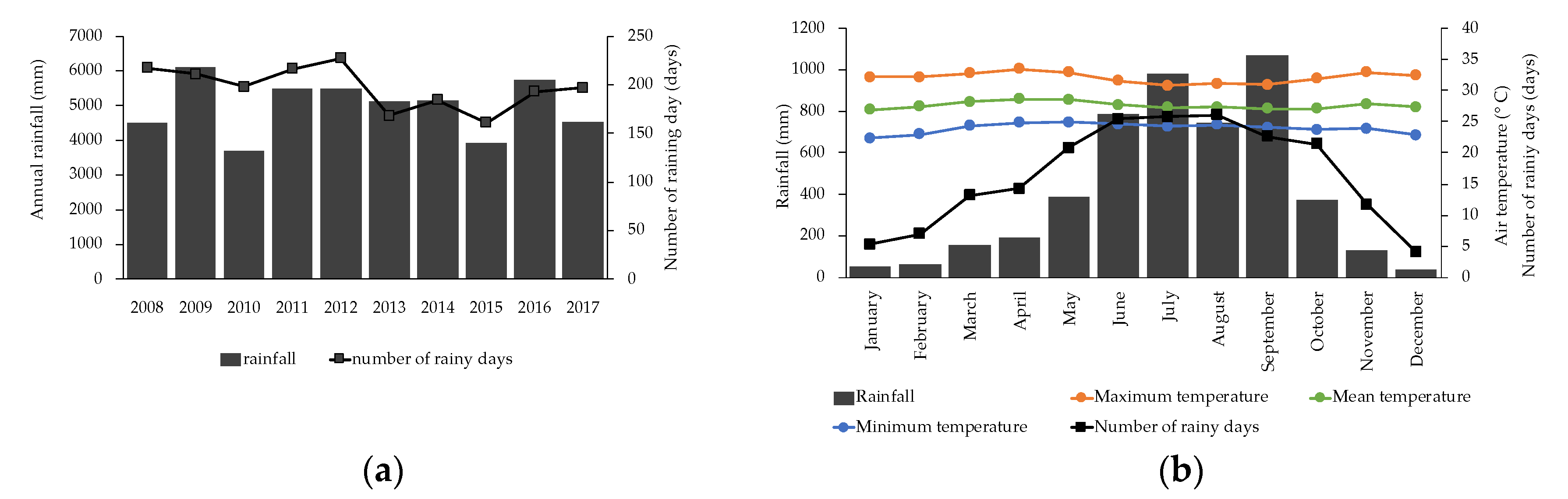
2.1. Study Site
2.2. Tree Census
2.3. Net Primary Productivity
2.4. Relative Elevation and Inundation Periods
2.5. Data Analysis
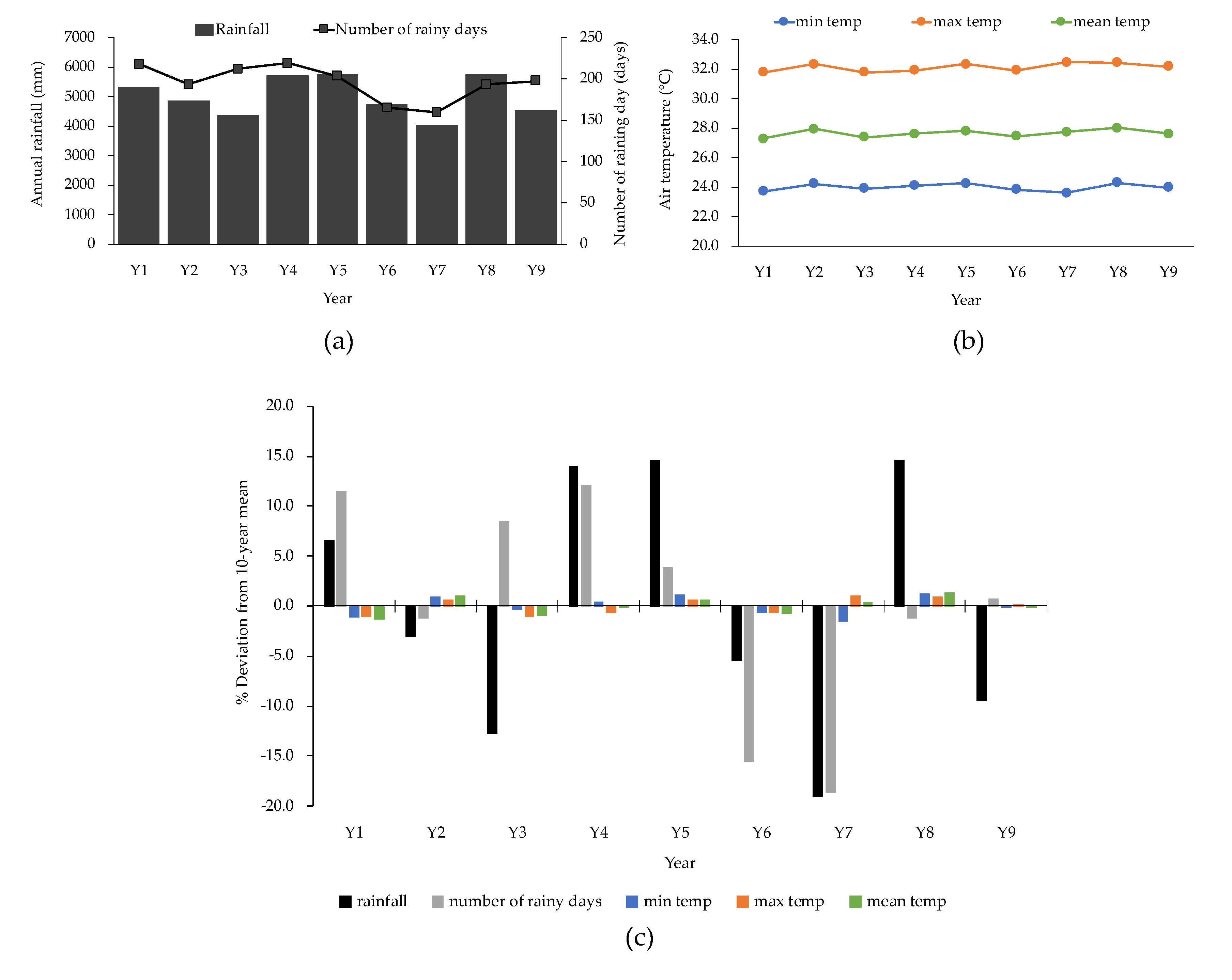
3. Results
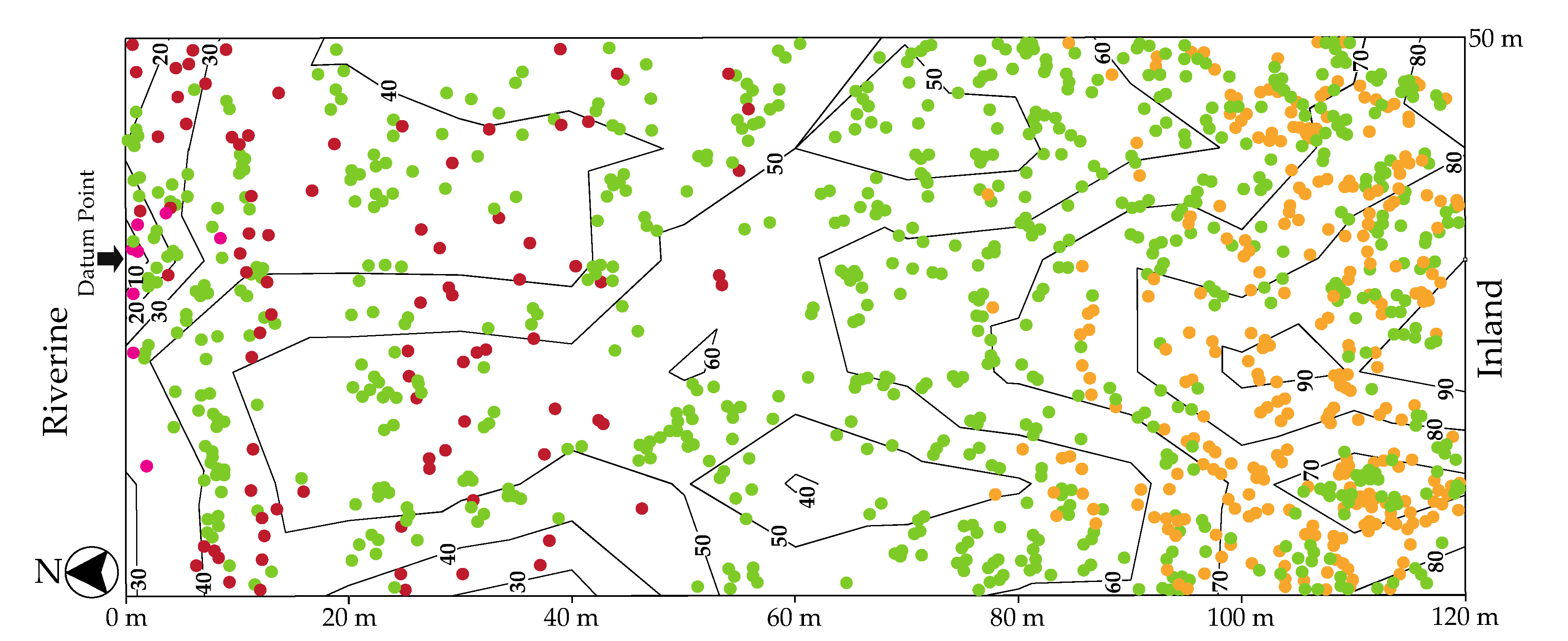
3.1. Species Distribution and Vegetation Structure
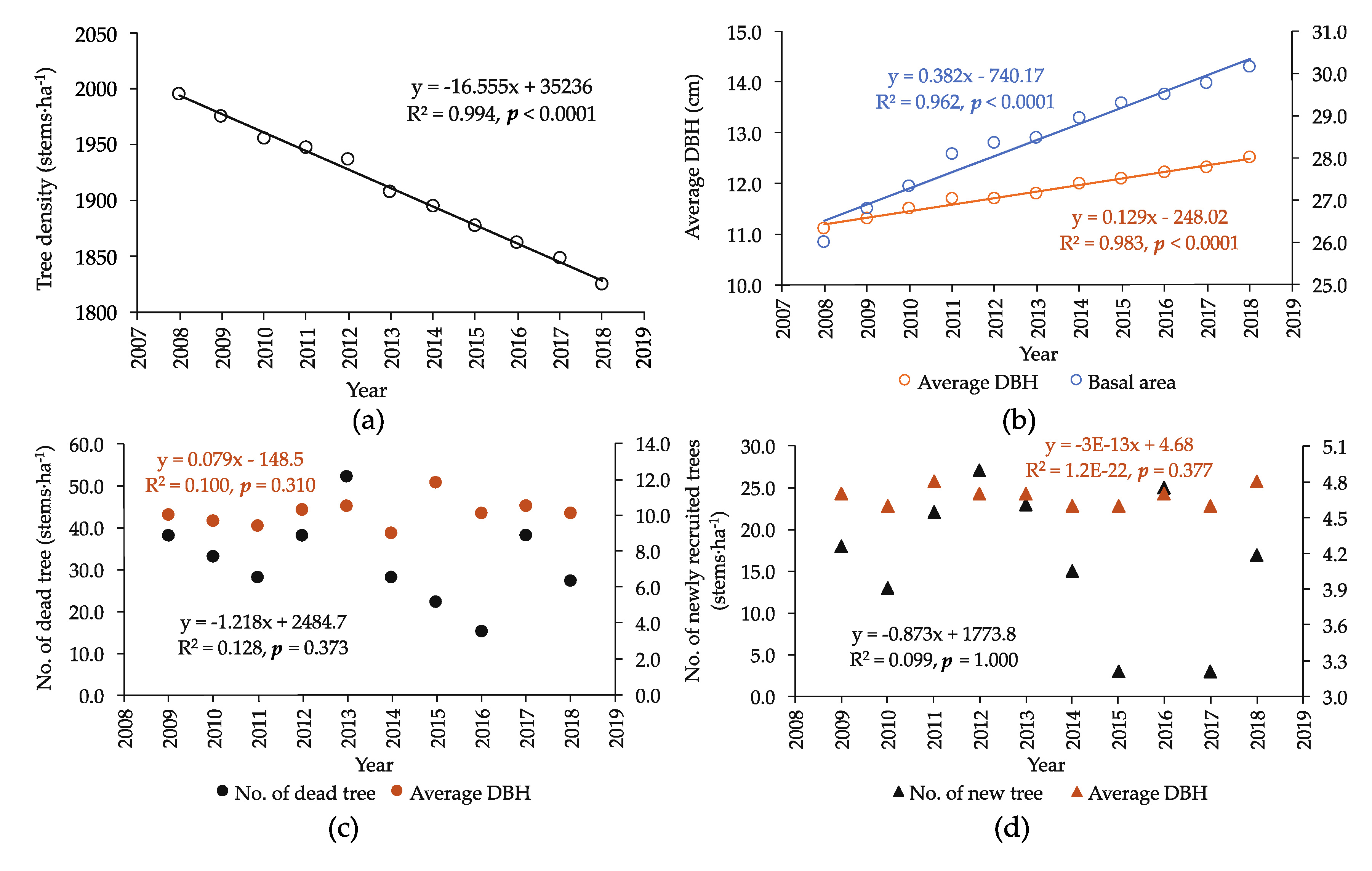
3.2. Growth Increment (ΔY) and Net Primary Productivity (NPP)
3.3. Relative Elevation and Inundation Periods
4. Discussion
4.1. Mangrove Forest Structure and Biomass
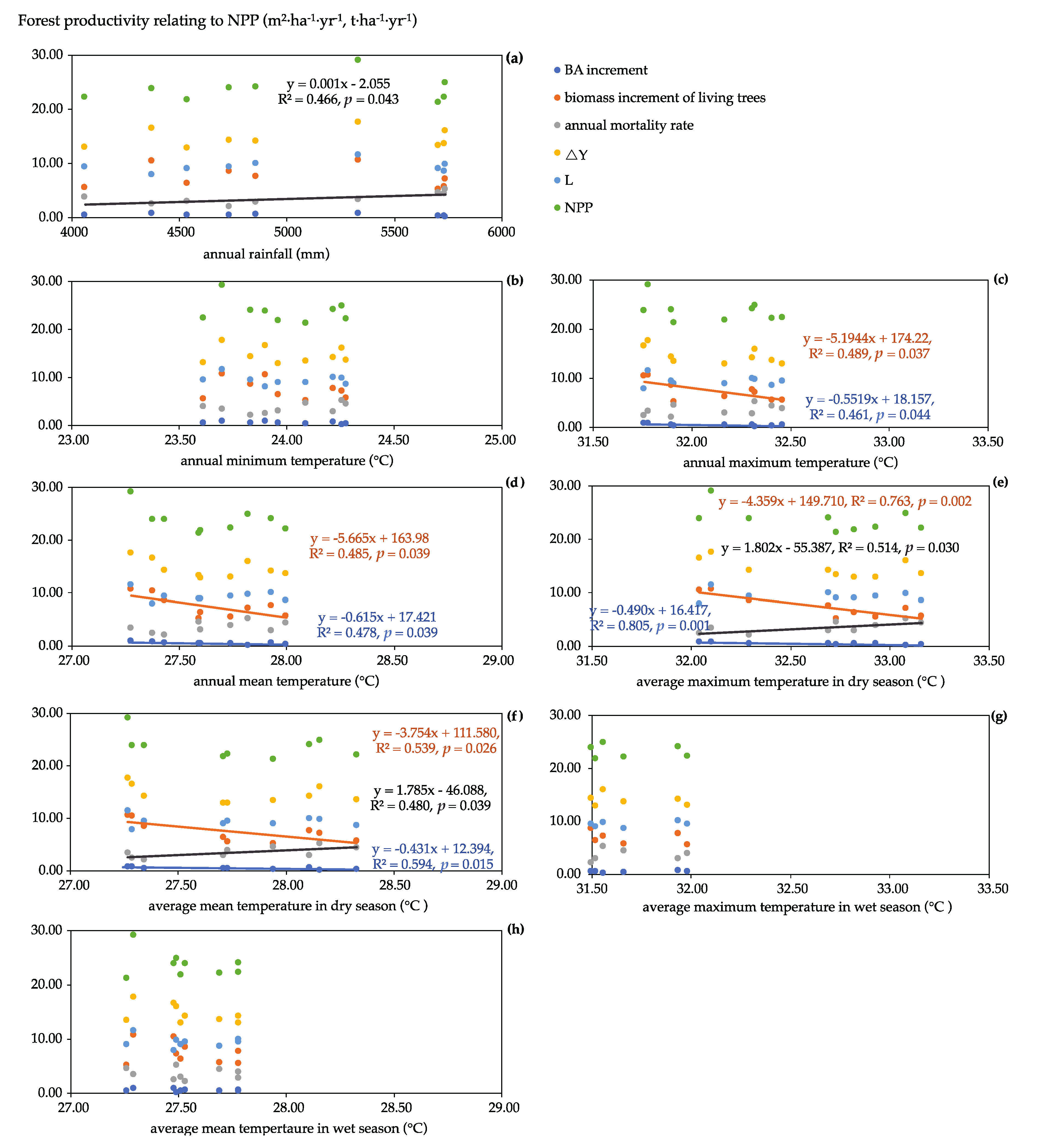
4.2. Growth, NPP Estimation, and Its Components
4.3. Temperature as an Initial Factor Regulating the NPP Magnitude
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nagelkerken, I.; Blaber, S.J.M.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.G.; Meynecke, J.-O.; Pawlik, J.; Penrose, H.M.; Sasekumar, A.; et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef] [Green Version]
- Kumara, M.P.; Jayatissa, L.P.; Krauss, K.W.; Phillips, D.H.; Huxham, M. High mangrove density enhances surface accretion, surface elevation change, and tree survival in coastal areas susceptible to sea-level rise. Oecologia 2010, 164, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, P.; Chaudhuri, S.; Ghosh, R. The role of mangroves in coastal and estuarine sedimentary accretion in Southeast Asia. In Sedimentary Processes; Aiello, G., Ed.; IntechOpen: London, UK, 2019; Volume 1, pp. 1–23. ISBN 978-1-78984-765-9. [Google Scholar]
- Hamilton, S.E.; Friess, D.A. Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat. Clim. Chang. 2018, 8, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, J.B.; Heider, C.; Norfolk, J.; Payton, F. Carbon stocks of intact mangroves and carbon emissions arising from their conversion in the Dominican Republic. Ecol. Appl. 2014, 24, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Taillardat, P.; Friess, D.A.; Lupascu, M. Mangrove blue carbon strategies for climate change mitigation are most effective at the national scale. Biol. Lett. 2018, 14, 20180251. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Sippo, J.Z.; Lovelock, C.E.; Santos, I.R.; Sanders, C.J.; Maher, D.T. Mangrove mortality in changing climate: An overview. Estuar. Coast. Shelf Sci. 2018, 215, 241–249. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Poungparn, S.; Komiyama, A.; Sangteian, T.; Maknual, C.; Patanaponpaiboon, P.; Suchewaboripont, V. High primary productivity under submerged soil raises the net ecosystem productivity of a secondary mangrove forest in eastern Thailand. J. Trop. Ecol. 2012, 28, 303–306. [Google Scholar] [CrossRef]
- Alongi, D.M.; Mukhopadhyay, S.K. Contribution of mangroves to coastal carbon cycling in low latitude seas. Agric. For. Meteorol. 2015, 213, 266–272. [Google Scholar] [CrossRef]
- Umnouysin, S.; Sangtiean, T.; Poungparn, S. Zonal distribution of coarse woody debris and its contribution to net primary production in a secondary mangrove forest. Ecol. Res. 2017, 32, 51–60. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Tomotsune, M.; Suchewaboripont, V.; Iimura, Y.; Kida, M.; Yoshitake, S.; Kondo, M.; Kinjo, K. Stand dynamics and aboveground net primary productivity of a mature subtropical mangrove forest on Ishigaki Island, south-western Japan. Reg. Stud. Mar. Sci. 2019, 27, 100516. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling and storage in mangrove forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Poungparn, S.; Komiyama, A.; Tanaka, A.; Sangtiean, T.; Maknual, C.; Kato, S.; Tanapermpool, P.; Patanaponpaiboon, P. Carbon dioxide emission through soil respiration in a secondary mangrove forest of eastern Thailand. J. Trop. Ecol. 2009, 25, 393–400. [Google Scholar] [CrossRef]
- Tomotsune, M.; Yoshitake, S.; Iimura, Y.; Kida, M.; Fujitake, N.; Koizumi, H.; Ohtsuka, T. Effects of soil temperature and tidal condition on variation in carbon dioxide flux from soil sediment in a subtropical mangrove forest. J. Trop. Ecol. 2018, 34, 268–275. [Google Scholar] [CrossRef]
- Schuur, E.A.G. Productivity and global climate revisited: The sensitivity of tropical forest growth of precipitation. Ecology 2003, 84, 1165–1170. [Google Scholar] [CrossRef]
- Clark, D.A.; Piper, S.C.; Keeling, C.D.; Clark, D.B. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984–2000. Proc. Natl. Acad. Sci. USA 2003, 100, 5852–5857. [Google Scholar] [CrossRef] [Green Version]
- Brienen, R.J.W.; Lebrija-Trejos, E.; Zuidema, P.A.; Martínez-Ramos, M.M. Climate growth analysis for a Mexican dry forest tree shows strong impact of sea surface temperatures and predicts future growth declines. Glob. Chang. Biol. 2010, 16, 2001–2012. [Google Scholar] [CrossRef]
- Ye, J.S.; Reynolds, J.F.; Sun, G.L.; Li, F.M. Impacts of increased variability in precipitation and air temperature on net primary productivity of the Tibetan Plateau: A modeling analysis. Clim. Chang. 2013, 119, 321–332. [Google Scholar] [CrossRef]
- Schippers, P.; Sterck, F.; Vlam, M.; Zuidema, P.A. Tree growth variation in the tropical forest: Understanding effects of temperature, rainfall and CO2. Glob. Chang. Biol. 2015, 21, 2749–2761. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, G.; Luo, T.; Dan, Y.; Zhou, L.; Lv, X. Variation of net primary productivity and its drivers in China’s forest during 2000–2018. For. Ecosyst. 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, J.; Manica, A.; Swetnam, R.; Balmford, A.; Spalding, M. Predicting global patterns in mangrove forest biomass. Conserv. Lett. 2014, 7, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.J.; Maher, D.T.; Tait, D.R.; Williams, D.; Holloway, C.; Sippo, J.Z.; Santos, I.R. Are global mangrove carbon stocks driven by rainfall? J. Geophys. Res. Biogeosci. 2016, 121, 2600–2609. [Google Scholar] [CrossRef]
- Sannigrahi, S.; Sen, S.; Paul, S. Estimation of mangrove net primary production and carbon sequestration service using light use efficiency model in the Sunderban biosphere region, India. Geophys. Res. Abs. 2016, 18, EGU2016-1884-1. [Google Scholar]
- Poungparn, S.; Charoenphonphakdi, T.; Sangtiean, T.; Patanaponpaiboon, P. Fine root production in three zones of secondary mangrove forest in eastern Thailand. Trees 2016, 30, 467–474. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Umnouysin, S.; Rodtassana, C.; Pravinvongvuthi, T.; Noda, T.; Kato, S. Occurrence of seasonal water replacement in mangrove soil and the trunk growth response of Avicennia alba related to salinity changes in a tropical monsoon climate. Ecol. Res. 2019, 34, 428–439. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Umnouysin, S.; Rodtassana, C.; Kato, S.; Pravinvongvuthi, T.; Sangtiean, T. Daily inundation induced seasonal variation in the vertical distribution of soil water salinity in an estuarine mangrove forest under a tropical monsoon climate. Ecol. Res. 2020. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S.; Kicklighther, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, L. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Leong, R.C.; Friess, D.A.; Crase, B.; Lee, W.K.; Webb, E.L. High-resolution pattern of mangrove species distribution is controlled by surface elevation. Estuar. Coast. Shelf Sci. 2018, 202, 185–192. [Google Scholar] [CrossRef]
- Snedaker, S.C. Mangrove species zonation: Why? In Tasks for Vegetation Science, 2nd ed.; Sen, D.N., Rajpurohit, K.S., Eds.; Dr. W Junk Publishers: The Hague, The Netherlands, 1982; Volume 46, pp. 111–125. ISBN 978-94-009-8037-2. [Google Scholar]
- Ellison, J.C. Wetlands of the Pacific Island region. Wetl. Ecol. Manag. 2009, 17, 169–206. [Google Scholar] [CrossRef]
- Ellison, J.C. Biogeomorphology of Mangroves. In Coastal Wetlands, “An Integrated Ecosystem Approach”, 2nd ed.; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 678–715. ISBN 978-0-444-63893-9. [Google Scholar]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986; p. 419. ISBN 0-521-25567-8. [Google Scholar]
- McIvor, A.; Spencer, T.; Möller, I.; Spalding, M. The Response of Mangrove Soil Surface Elevation to Sea Level Rise; Natural Coastal Protection Series: Report 3. Cambridge Coastal Research Unit Working Paper 42; The Nature Conservancy and Wetlands International: Cambridge, UK, 2013; 59p, Available online: http://coastalresilience.org/science/mangroves/surface-elevation-and-sea-level-rise (accessed on 28 July 2020).
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; Verheyden, A.; De Genst, W.; Hettiarachchi, S.; Koedam, K. Four decade vegetation dynamics in Sri Lankan mangroves as detected from sequential aerial photography: A case study in Galle. Bull. Mar. Sci. 2000, 67, 741–759. [Google Scholar]
- Komiyama, A.; Moriya, H.; Prawiroatmodjo, S.; Toma, T.; Ogino, K. Primary productivity of mangrove forest. In Biological System of Mangroves; A Report of East Indonesian Mangrove Expedition 1986; Ogino, K., Chihara, M., Eds.; Ehime University: Ehime, Japan, 1988; pp. 97–117. [Google Scholar]
- Mall, L.P.; Singh, V.P.; Garge, A. Study of biomass, litter fall, litter decomposition and soil respiration in monogeneric mangrove and mixed mangrove forest of Andaman Islands. Trop. Ecol. 1991, 32, 144–152. [Google Scholar]
- Amarasinghe, M.D.; Balasubramaniam, S. Net primary productivity of two mangrove forest stands on the northwestern coast of Sri Lanka. Hydrobiologia 1992, 247, 37–47. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Ahmed, S.; Osawa, A. Biomass and net primary productivity of mangrove communities along the Oliohaline zone of Sundarbans, Bangladesh. For. Ecosyst. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Kamruzzaman, M.; Osawa, A.; Deshar, R.; Sharma, S.; Mouctar, K. Species composition, biomass, and net primary productivity of mangrove forest in Okukubi River, Okinawa Island, Japan. Reg. Stud. Mar. Sci. 2017, 12, 19–27. [Google Scholar] [CrossRef]
- Lovelock, C.E. Soil respiration and belowground carbon allocation in mangrove forests. Ecosystems 2008, 11, 342–354. [Google Scholar] [CrossRef]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado-Molina, C.; Ewe, S.M.L. Patterns of root dynamics in mangrove forests along environmental gradients in Florida Coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, X.; Guan, W.; Liao, B.; Chen, Y.; Li, M.; Zhong, C. Fine root functional group based estimates of fine root production and turnover rate in natural mangrove forests. Plant Soil 2017, 413, 83–95. [Google Scholar] [CrossRef]
- Cormier, N.; Twilley, R.R.; Ewel, K.C.; Krauss, K.W. Fine root productivity varies along nitrogen and phosphorus gradients in high-rainfall mangrove forests of Micronesia. Hydrobiologia 2015, 750, 69–87. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Virgulino-Júnior, P.C.C.; Gardunho, D.C.L.; Silva, D.N.C.; Fernandes, M.E.B. Wood density in mangrove forests on the Brazilian Amazon coast. Trees 2020, 34, 51–60. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove dieback during fluctuating sea levels. Sci. Rep. 2017, 7, 1680. [Google Scholar] [CrossRef]
- Saenger, P.; Snedaker, S.C. Pantropical trends in mangrove above-ground biomass and annual litterfall. Oecologia 1993, 96, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, X.; Wang, M. Propagule dispersal determines mangrove zonation at intertidal and estuarine scales. Forests 2019, 10, 245. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B. Biomass and primary production of Rhizophora apiculata Bl. in a mangrove in southern Thailand. Aquat. Bot. 1978, 4, 43–52. [Google Scholar] [CrossRef]
- Peng, C.J.; Qian, J.W.; Guo, X.D.; Zhao, H.W.; Hu, N.X.; Yang, Q.; Chen, C.P.; Chen, L.Z. Vegetation carbon stocks and net primary productivity of the mangrove forests in Shenzhen, China. J. Appl. Ecol. 2016, 27, 2059–2065. [Google Scholar]
- Ong, J.E.; Gong, W.K.; Clough, B.F. Structure and productivity of a 20-year-old stand of Rhizophora apiculata Bl. mangrove forest. J. Biogeogr. 1995, 22, 417–424. [Google Scholar]
- Day, J.W., Jr.; Conner, W.H.; Ley-Lou, F.; Day, R.H.; Navarro, A.M. The productivity and composition of mangrove forests, Laguna de Terminos, Mexico. Aquat. Bot. 1987, 27, 267–284. [Google Scholar] [CrossRef]
- Xiong, Y.; Cakir, R.; Phan, S.M.; Ola, A.; Krauss, K.W.; Lovelock, C.E. Global patterns of tree stem growth and stand aboveground wood production in mangrove forests. For. Ecol. Manag. 2019, 444, 382–392. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Krauss, K.W.; Osland, M.J.; Reef, R.; Ball, M.C. The physiology of mangrove trees with changing climate. Trop. Tree Physiol. 2016, 6, 149–179. [Google Scholar]
- Moon, T.; Joughin, I. Change in ice front position on Greenland’s outlet glaciers from 1992 to 2007. J. Geophys. Res. 2008, 113, F02022. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Savenije, H.H.G.; Toffolon, M. A new analytical framework for assessing the effect of sea-level rise and dredging on tidal damping in estuaries. J. Geophys. Res. 2012, 117, C09023. [Google Scholar] [CrossRef]
- Widlansky, M.J.; Timmermann, A.; Cai, W. Future extreme sea level seesaws in the tropical Pacific. Clim. Chang. 2015, 1, e1500560. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.K.; Mitra, A. Salinity: A primary growth driver of mangrove flora. Curr. Trends For. Res. CTFR 2018, 114. [Google Scholar] [CrossRef]
- Jintana, V.; Komiyama, A.; Moriya, H.; Ogino, K. Forest ecological studies of mangrove ecosystem in Ranong, Southern Thailand: (4) Diameter growth measurement by dendrometry. In Study on the Mangrove Ecosystem; Sugi, J., Ed.; Nodai Research Institute, Tokyo University of Agriculture: Tokyo, Japan, 1985; pp. 227–233. [Google Scholar]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The next generation of scenarios for climate change research and assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2013; 1525p. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Y1 | Y2 | Y3 | Y4 | Y5 | Y6 | Y7 | Y8 | Y9 | |
|---|---|---|---|---|---|---|---|---|---|
| Basal area increment (m2·ha−1·yr−1) | |||||||||
| 0.79 | 0.51 | 0.76 | 0.29 | 0.1 | 0.46 | 0.36 | 0.28 | 0.36 | |
| (1) Biomass increment in living trees (t·ha−1·yr−1) | |||||||||
| Aboveground | 7.816 | 5.659 | 7.651 | 3.835 | 5.366 | 6.275 | 4.08 | 4.152 | 4.659 |
| Root | 2.803 | 1.947 | 2.735 | 1.318 | 1.747 | 2.227 | 1.40 | 1.421 | 1.60 |
| Total | 10.619 | 7.606 | 10.386 | 5.153 | 7.113 | 8.502 | 5.479 | 5.573 | 6.259 |
| (2) Increment of newly recruited trees mass (t·ha−1·yr−1) | |||||||||
| Aboveground | 0.014 | 0.041 | 0.029 | 0.025 | 0.024 | 0.005 | 0.000 | 0.018 | 0.001 |
| Root | 0.007 | 0.023 | 0.015 | 0.012 | 0.013 | 0.003 | 0.001 | 0.009 | 0.000 |
| Total | 0.021 | 0.064 | 0.044 | 0.037 | 0.037 | 0.008 | 0.001 | 0.027 | 0.001 |
| (3) Annual mortality rate (t·ha−1·yr−1) | |||||||||
| Aboveground | 2.27 | 1.94 | 1.66 | 3.19 | 3.56 | 1.39 | 2.70 | 3.03 | 2.03 |
| Root | 1.02 | 0.86 | 0.74 | 1.31 | 1.57 | 0.64 | 1.08 | 1.28 | 0.88 |
| Total | 3.29 | 2.80 | 2.40 | 4.50 | 5.13 | 2.03 | 3.78 | 4.31 | 2.91 |
| (4) Fine root production (t·ha−1·yr−1) | |||||||||
| 3.64 | 3.64 | 3.64 | 3.64 | 3.64 | 3.64 | 3.64 | 3.64 | 3.64 | |
| (1+2+3+4) Growth increment;△Y (t·ha−1·yr−1) | |||||||||
| Aboveground | 10.10 | 7.64 | 9.34 | 7.05 | 8.95 | 7.67 | 6.78 | 7.20 | 6.69 |
| Root | 7.47 | 6.47 | 7.13 | 6.28 | 6.97 | 6.51 | 6.12 | 6.35 | 6.12 |
| Total | 17.57 | 14.11 | 16.47 | 13.33 | 15.92 | 14.18 | 12.90 | 13.55 | 12.81 |
| Litter production (t·ha−1·yr−1) | |||||||||
| Leaf | 8.12 | 7.50 | 6.09 | 6.41 | 6.69 | 6.84 | 6.89 | - | - |
| Wood | 1.21 | 1.04 | 0.56 | 0.55 | 0.69 | 1.11 | 0.89 | - | - |
| Reproductive organ | 2.14 | 1.40 | 0.67 | 0.90 | 1.54 | 1.76 | 1.56 | - | - |
| Total | 11.47 | 9.94 | 7.32 | 7.86 | 8.92 | 9.71 | 9.34 | 8.56 * | 8.91 * |
| NPP (t·ha−1·yr−1) | |||||||||
| Aboveground | 21.57 | 17.58 | 16.66 | 14.91 | 17.87 | 17.38 | 16.12 | 15.76 | 15.60 |
| Root | 7.47 | 6.47 | 7.13 | 6.28 | 6.97 | 6.51 | 6.12 | 6.35 | 6.12 |
| Total | 29.04 | 24.05 | 23.79 | 21.19 | 24.84 | 23.89 | 22.24 | 22.11 | 21.72 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poungparn, S.; Komiyama, A.; Umnouysin, S.; Rodtassana, C.; Sangtiean, T.; Maknual, C.; Pravinvongvuthi, T.; Suchewaboripont, V.; Kato, S. Ten-Year Estimation of Net Primary Productivity in a Mangrove Forest under a Tropical Monsoon Climate in Eastern Thailand: Significance of the Temperature Environment in the Dry Season. Forests 2020, 11, 987. https://doi.org/10.3390/f11090987
Poungparn S, Komiyama A, Umnouysin S, Rodtassana C, Sangtiean T, Maknual C, Pravinvongvuthi T, Suchewaboripont V, Kato S. Ten-Year Estimation of Net Primary Productivity in a Mangrove Forest under a Tropical Monsoon Climate in Eastern Thailand: Significance of the Temperature Environment in the Dry Season. Forests. 2020; 11(9):987. https://doi.org/10.3390/f11090987
Chicago/Turabian StylePoungparn, Sasitorn, Akira Komiyama, Suthathip Umnouysin, Chadtip Rodtassana, Tanuwong Sangtiean, Chatree Maknual, Tamanai Pravinvongvuthi, Vilanee Suchewaboripont, and Shogo Kato. 2020. "Ten-Year Estimation of Net Primary Productivity in a Mangrove Forest under a Tropical Monsoon Climate in Eastern Thailand: Significance of the Temperature Environment in the Dry Season" Forests 11, no. 9: 987. https://doi.org/10.3390/f11090987