Lack of Cascading Effects of Eurasian Lynx Predation on Roe Deer to Soil and Plant Nutrients



1
Norwegian Institute for Nature Research, PO Box 5685 Torgarden, NO-7485 Trondheim, Norway
2
Department of Nature, Environment and Health, Queen Maud University College for Early Childhood Education, Thrond Nergaards veg 7, 7044 Trondheim, Norway
3
Faculty of Applied Ecology, Agricultural Sciences and Biotechnology, Inland Norway University of Applied Sciences, Postboks 400, 2418 Elverum, Norway
*
Author to whom correspondence should be addressed.
†
Present address: Saghaugen gård, Skjomdalveien-Bogholm 21, 8523 Skjomen, Norway.
Diversity 2020, 12(9), 352; https://doi.org/10.3390/d12090352
Submission received: 13 August 2020
/
Revised: 8 September 2020
/
Accepted: 11 September 2020
/
Published: 14 September 2020
(This article belongs to the Special Issue Ecology of Predation and Scavenging and the Interface)
Abstract
:This study examines the extent to which above-ground trophic processes such as large carnivore predation on wild ungulates can cause cascading effects through the provision of carrion resources to below-ground ecosystem processes in the boreal forest of southeastern Norway. We measured the levels of 10 parameters in soil samples and 7 parameters in vegetation (wavy hair-grass, Avenella flexuosa, and bilberry, Vaccinium myrtillus) at 0, 0.5 and 2 m distance from 18 roe deer (Capreolus caprelous) carcasses killed by Eurasian lynx (Lynx lynx). We then compared these values to two control sites 20 m away from each carcass. Sampling was conducted 20–29 months after death. Neither soil nor vegetation samples showed a clear gradient in parameters (CN, NH4+, NO3−, P, PO4−, Ca, K, Mg and Na) from the center of a carcass towards the periphery. Similarly, there was no difference in the effect on soil and vegetation between winter- and summer-killed carcasses. Our results contrast with that of other studies that simulate the effect of predation with whole carcasses and which often exclude scavengers through fencing. The lack of detectable effects after about two years is likely due to the small size of roe deer carcasses and the fact that most tissues are consumed by the predator and scavengers before decomposition.
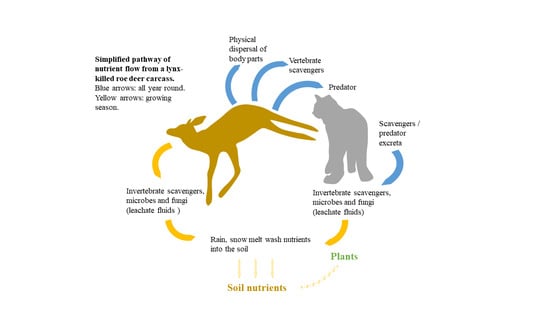
1. Introduction
There has been a focus on the wider ecological effects of large terrestrial carnivores in recent years driven by both the study of ecosystems where they are being reintroduced or are naturally increasing in density [1,2] and where their absence is conspicuous [3]. This focus is largely due to their putative role as ecological keystone species [4] or as strongly interactive species [5]. Although these impacts are not as universal as is often claimed [4,6], there are plenty of examples of large carnivores having cascading effects on the ecosystems of which they are a part [7,8]. These impacts mainly operate through the influence of large terrestrial carnivores on the density, distribution and behavior of their wild ungulate prey, species that have effects on vegetation through diverse mechanisms [3,9,10]. This in turn affects a wide range of other species that respond to the environmental changes [3,11]. Other mechanisms involve the increased availability of carrion to scavengers as a by-product of predation [12,13,14,15,16,17].
Another pathway through which predation can potentially influence diverse ecosystem functions is through the transfer of nutrients from carnivore-killed prey to soil and then into vegetation [9]. Some studies have shown that this nutrient pulse, although local in extent, can be intense [18,19,20,21,22]. For example, a range of studies have focused on the role of bears, Ursus spp., in transferring marine nutrients from anadromous salmon into terrestrial ecosystems [23]. However, the generality of this impact is likely to depend on how much the predator consumes its prey and how much of the remains are consumed or dispersed by scavengers (both micro and macro [24]) as it is only the leachate fluids resulting from invertebrate maggot activity and microbial decomposition that are directly transferred to soil [25]. In addition, nutrients also can originate from dead invertebrates, and excretion from both scavengers and predators feeding on or attracted to the carcass [25,26]. By this mechanism, the impacts of large carnivore predation can cascade through the ecosystem, bridging the divide between above- and below-ground ecosystems [27]. Moreover, carcasses, by locally reducing herbaceous cover, can influence the early stages of tree reproduction [28]. The impacts of carrion on soil and vegetation parameters are also influenced by temperature and the season of death [29,30,31].
The aim of our study was to investigate the presence of these cascading effects in a previously unstudied system. For this purpose, we studied a large carnivore (Eurasian lynx, Lynx lynx)–ungulate (roe deer, Capreolus capreolus) system in the boreal forest of southeastern Norway.
Our predictions were as follows:
- (1)
- Lynx-killed roe deer carcasses will have an effect on nutrient levels in the surrounding soil and vegetation, so that soil and vegetation parameters would be changed to a greater extent closer to the center of the carcass and decline farther away from the carcass [21].
- (2)
2. Materials and Methods
2.1. Study Area
The study was conducted approximately 50 km southeast of Oslo, Norway, (N 59°36′55″, E 11°36′45″) in the municipalities of Spydeberg, Enebakk, Hobøl and Aurskog-Høland in Viken county (formerly Østfold and Akershus counties). The area consists of rolling hills with residential and agricultural areas (25%) surrounded by forest (75%) dominated by Norway spruce (Picea abies) and Scots pine (Pinus sylvestris) mixed with downy birch (Betula pubescens), mainly used for timber production. During the growing season, the agricultural fields are used for the production of cereals and fodder grass. The main ungulate species were roe deer and moose (Alces alces). The main predators of ungulates were lynx, which were present throughout the landscape, and individual transient wolves (Canis lupus). Important scavengers were pine martens (Martes martes), red foxes (Vulpes vulpes), Eurasian badgers (Meles meles), domestic cats (Felis domesticus), ravens (Corvus corax), hooded crows (Corvus cornix), magpies (Pica pica) and Eurasian jays (Garrulus glandarius), as found from video observation (unpublished data).
2.2. Study Design
The ecology of lynx predation on roe deer has been studied in the region since 2000 by (1) following VHF- and GPS-collared lynx to find kills, (2) snow-tracking unmarked lynx to find kills and (3) regularly monitoring marked roe deer to determine the timing and cause of death [32,33]. Field procedures are described in Nilsen et al. 2009 [32]. This work led us to locate and mark the kill-sites of roe deer that had been killed and at least partially consumed by lynx. Permissions were granted by the Norwegian Environment Agency and procedures were approved by the Norwegian Committee for Experimental Animal Welfare (permit numbers 08/127430, 07/81885, 07/7883, 2004/48647, 201/01/641.5/FHB, 127/03/641.5/fhb, 1460/99/641.5/FBe, 1081/97/641.5/FBe, and NINA 1/95).
Soil and vegetation samples were simultaneously collected at the end of the growing season (between the 29th of June and the 29th of July) in 2004 from 18 carcass sites, where six of the roe deer were killed by lynx in winter (October–April) and 12 in summer (May–September). The samples were taken 20–29 months after death, following the methods described in Melis et al. [20]. The samples were taken along a line with the carcass as the center, at distances of 0, 0.5, 2 and 20 m extending in opposite directions from the center of the carcass. The head to tail length of a roe deer is between 95 and 135 cm, and lynx and other scavengers can move roe deer carcasses slightly while feeding on them, so the distances from the center to 0.5 m were considered as being directly influenced by the carcass. The sample at 20 m was considered not to be affected by the carcass and was used as a control. When we visited the carcass site after the lynx had left (Figure 1a,b), we documented how much of the kill was left, as well as ground vegetation and forest type. We marked the carcass site with a pole and a label (Figure 1a,c,d) at the middle point and took pictures to be able to find again the exact carcass position. In 16 of the 18 carcasses, the lynx had removed 90% of the meat according to our visual estimation several days after the lynx had abandoned the kill. The direction of the sample line was chosen so that it fell parallel to the contour of the landscape, rather than perpendicular to the slope. The samples from both sides were merged for each distance for analysis. The control sample at 20 m was taken within the same vegetation type as the carcass was at. The litter was removed and a 2 × 2 × 5 cm (length × width × depth) sample of soil was taken from the surface. Fresh green leaves were collected (3–5 g dry weight per sample) from the most common plants present at the sites (wavy hair-grass, Avenella flexuosa, and bilberry, Vaccinium myrtillus). The combination of nutrient leaching, pH shock (a rapid increase in pH) and blocking of light and oxygen by the physical presence of the carcass killed the vegetation directly under the carcass (Figure 1c), as described in [25]. This effect could last for more than two years (Figure 1d). Therefore, we were not always able to collect vegetation samples at the center.
2.3. Laboratory Analyses
In total, 70 soil and 52 vegetation (25 A. flexuosa, 27 V. myrtillus) samples were analyzed by the Wageningen University Testing Laboratory of the Resource Ecology Group.
In soil, the concentrations of total nitrogen (N), total phosphorus (P) and calcium (Ca) were determined after digestion with H2SO4−Se-salicylacid-H2O2 [34,35]. Concentrations of inorganic nitrogen (nitrate NO3−, ammonium NH4+), inorganic phosphorus (phosphate PO4−), potassium (K), magnesium (Mg) and sodium (Na) in soil were determined after extraction with 0.01 M CaCl2 by using the method described in [36].
In plant leaves, the concentrations of total N, total P, Ca, K, Mg and Na were determined by digestion with H2SO4−Se-salicylacid-H2O2. The C/N ratio in both soil and vegetation was determined using a C/N analyzer (Fisons EA 1108, CHN-0, Woodstock III, Interscience B.V., Breda, Brabant, The Netherlands). Concentrations of total N and total P in soil and vegetation, and inorganic nitrogen (nitrate NO3−, ammonium NH4+) and inorganic phosphorus (phosphate PO4−) in soil were measured with an auto-analyzer (Skalar San Plus, colorimetric system, Skalar Analytical B.V., Breda, Noord-Brabant, The Netherlands). K, Ca, Mg and Na were measured with an atomic absorption spectrometer (Varian, SpectrAA-600, Varian B.V. Benelux, Middelburg, Zeeland, The Netherlands).
2.4. Statistical Analyses
To account for statistical non-independence caused by having several samples at the same carcass site, the variable “site” was added as a random effect in a generalized linear mixed model (GLMM) framework [37]. We used a model selection procedure based on the Akaike information criterion corrected for small sample sizes [38] to identify which variables were best at explaining the variation in the concentration of a particular nutrient. The full model included the concentration of a nutrient as the dependent variable, and “distance” (distance from the center of the carcass) and “season” (season of kill) as explanatory variables. The minimum adequate model (MAM, hereafter) for each analysis was chosen based on the AICc value, Akaike weight and the number of parameters [38]. If the variable “distance” was not included in the MAM, we assumed that the effect of the carcass was not detectable on that parameter.
Shapiro–Wilk normality tests and Q–Q plots were used to check for normality. Response variables were transformed to improve normality, when not normally distributed. For soil nutrient concentrations, we square root-transformed nitrogen, calcium and magnesium, cube root-transformed phosphorus and sodium and log-transformed (natural logarithm) all other parameters. For vegetation samples, we square root-transformed calcium and magnesium, cube root-transformed sodium and log-transformed all other parameters.
The analyses were conducted separately for soil and for each of the two plant species using R 2.6.1 software [39], where the model selection was done with the R package MuMIn version 1.43.17 [40] and the linear mixed models were run with the add-on package lme4 [41]. The mixed models were fitted with maximum likelihood for the calculation of the AICc values, while the parameter estimates and their standard errors were from models fitted with restricted maximum likelihood [37].
3. Results
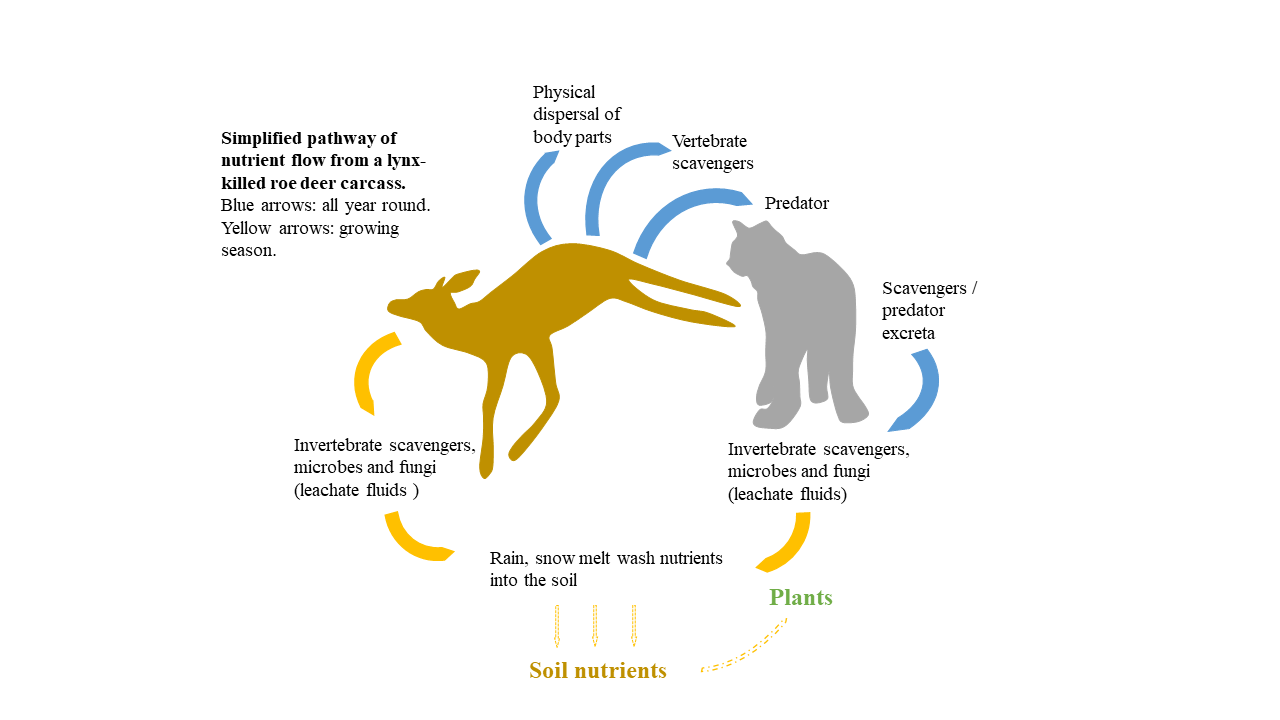
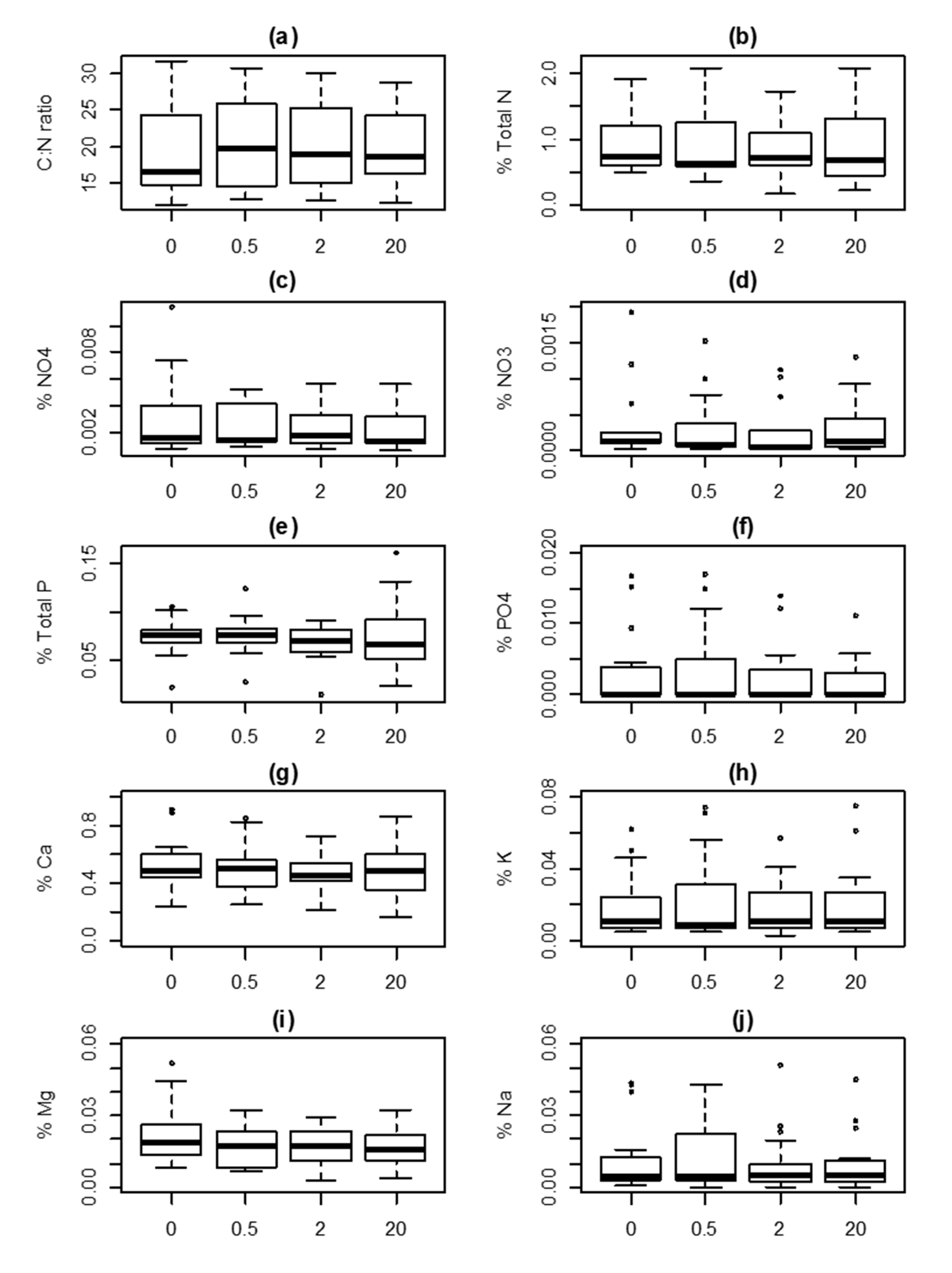
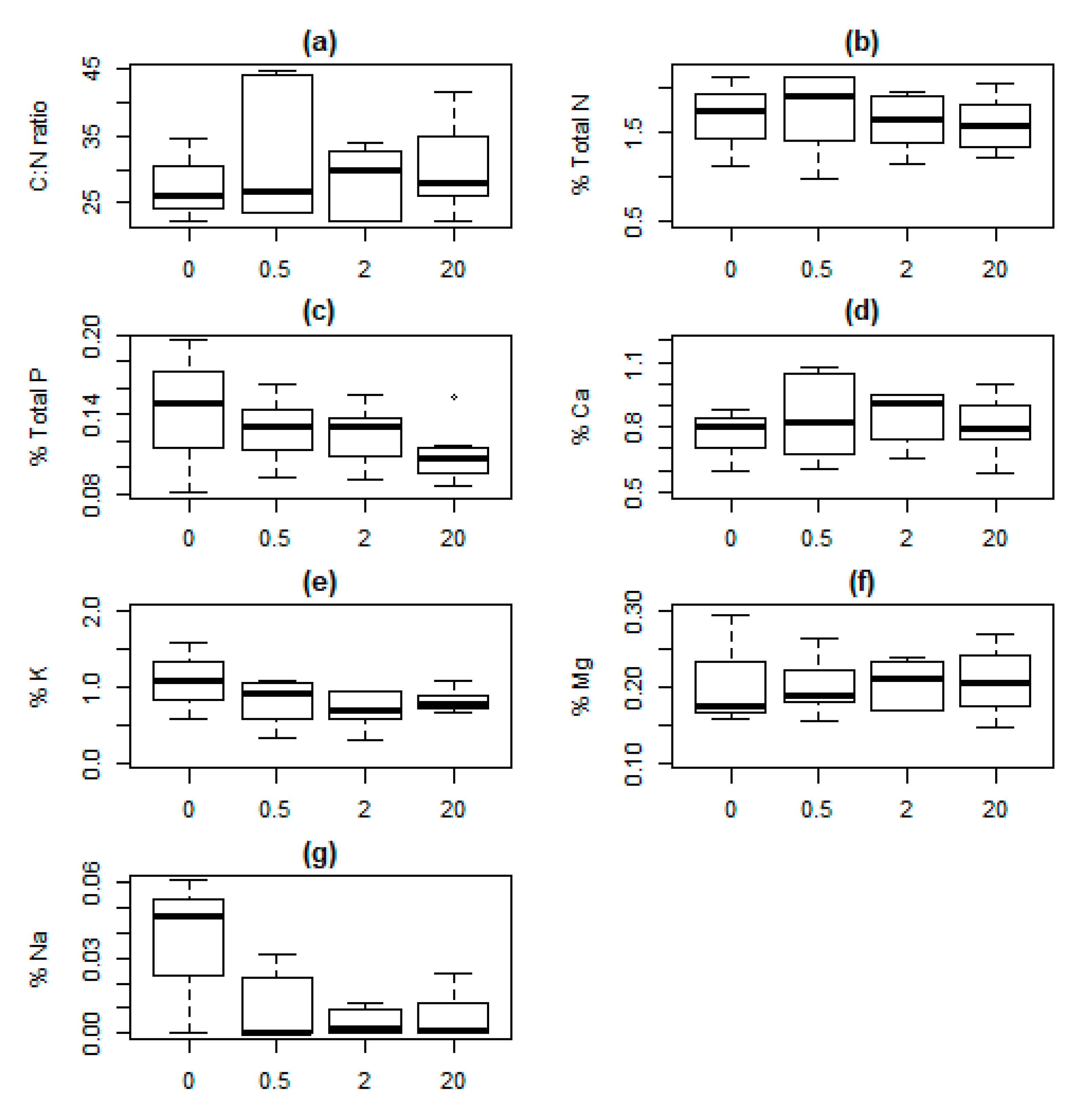
The variable “distance” was not included in the MAM for either soil or vegetation nutrients (data on model selection shown in Table 1 for soil, and in Table 2 and Table 3 for V. myrtillus, and A. flexuosa, respectively). This indicated that the effect of the carcasses on soil and vegetation nutrient concentrations could not be detected (Figure 2, Figure 3 and Figure 4).
4. Discussion
In contrast to our predictions, we found no detectable below-ground impacts of lynx-killed roe deer carcasses. Our results differ from other published studies, such as on muskox (Ovibos moschatus) carcasses in the arctic [19], moose in Isle Royale [18], American bison (Bison bison) in prairies [21] and kangaroos (Macropus giganteus) in Eucalyptus woodland [42], that detected an effect of carcass decomposition on soil and vegetation nutrients concentration [18,19,20,21,25], begging the question, what differs in our study system? A number of potential explanations present themselves, but all must be seen in light of the fact that only the liquids resulting from the invertebrate maggot activity and microbial decomposition leach into the soil [25].
First, the effect of roe deer carcasses on soil and vegetation is expected to vary depending on how much of its prey is consumed by the lynx and how much the remnants are consumed or dispersed by scavengers [24,26]. Since roe deer are quite small ungulates, with a mean adult body mass of 20–30 kg [43], and lynx tend to consume most of the consumable meat and organs over the course of 1 to 5 days after a kill [44,45], the amount of nutrients available for leaking into the soil is rather limited. Moreover, vertebrate scavengers tend to arrive within a few hours or days and rapidly remove the rest of the meat/organs [30]. On our sample of kills, more than 90% of the meat was removed from almost all carcasses when we first examined the kill sites several days after the lynx had abandoned the kills.
This issue is enhanced by the fact that a good proportion of the kills (6 out of 18) happened in the winter season, when insects are inactive and decomposition rates are lower—thus giving even more time for scavengers to remove meat and disperse body parts. Combined, these issues point to a need to consider the methodology when comparing studies. Our use of real predator-killed and consumed carcasses contrasts to most other studies that simulate this effect with whole carcasses and which often exclude scavengers through fencing [42]. This difference needs to be considered when generalizing between carrion derived from different sources and between studies with different methodologies.
Second, another reason for not finding an effect could lie with the time interval between roe deer death and carcass sampling. In this study, similarly to a study conducted on much larger European bison (Bison bonasus) carcasses in a temperate forest [20], we collected samples two or more years after death. For European bison, we were able to detect an effect on pH and calcium that lasted up to seven years after the death of the animal. However, the turnover of nitrate in soil was very fast (up to one year) even though bison are significantly larger than roe deer (average body mass across sexes is over 500 kg [46]) and we might expect the impact on soil and vegetation to last longer. In a tallgrass prairie system, the nutrients released from American bison carcasses affected plant species composition for at least five years after bison death, thus increasing grassland heterogeneity [21]. Other studies on kangaroo carcasses (ca 30 kg) have also shown that nutrient effects can still be evident after five years [42].
Thirdly, the lack of elevation in nutrients in the plants that we sampled close to a carcass could be because sampled V. myrtillus belonged to the same clone and therefore to various degree exchange resources underground between ramets or store nutrients in storage organs [47,48,49], thus potentially diluting the spatial impact of the nutrient pulse. It is, however, unlikely that the controls (20 m) should belong to the same clone as the plants close to the carcass. A final consideration concerns our sample sizes. Although not very large, they are very much within the range of sample sizes used in similar studies that have found significant results. Therefore, although we may not be able to reject the existence of any effects of the carcasses, we feel comfortable to say that if they exist, they are much more subtle than other studies have shown for larger carcasses in different ecosystems.
Our findings underline the complexity, diversity and context dependence of ecological processes, as well as illuminating the dangers of over-generalizations from case studies to universal principles. Although there is a growing list of case studies documenting a diversity of large predator-mediated trophic cascades in terrestrial as well as aquatic ecosystems, there are also multiple reviews that underline that there are also many exceptions [6,50] and calls for caution [51]. Genuine scientific progress in this value-laden field will only be possible by building up a solid body of positive and negative results embracing the diversity of contexts (both ecological and anthropogenic) in which predation occurs in the Anthropocene [52,53].
Author Contributions
Conceptualization, I.J.M.T. and J.D.C.L.; methodology, C.M., C.S., I.J.M.T. and J.D.C.L.; formal analysis, C.M. and I.J.M.T.; writing—original draft preparation, C.M., C.S., I.J.M.T. and J.D.C.L.; writing—review and editing, C.M., C.S., I.J.M.T. and J.D.C.L.; funding acquisition, J.D.C.L. and I.J.M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This study was conducted as part of the Scandlynx project which is mainly funded by the Norwegian Directorate for Nature Management, the Research Council of Norway (projects 134242, 165814, 183176, 212919, 251112), the county governor’s office of Viken county, and the Norwegian Institute for Nature Research. IJMT was also funded by two individual grants from the Research Council of Norway’s cultural agreement program with the Netherlands (2002-03 and 2003-04) and two grants from the Schure-Beijerinck-Popping fund. CM was funded by a postdoctoral grant at the Norwegian University for Science and Technology, Trondheim.
Acknowledgments
The authors are grateful to all the landowners who permitted our activity on their land, and to Martin Hauger and Øistein Høgseth for assistance in the field. The Ski municipality assisted by providing carcasses and the Norwegian University for Life Sciences provided drying ovens for vegetation samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, D.W.; Peterson, R.O.; Houston, D.B. Yellowstone after wolves. Bioscience 2003, 53, 330–340. [Google Scholar] [CrossRef]
- Berger, J.; Smith, D.W. Restoring functionality in Yellowstone with recovering carnivores: Gains and uncertainties. In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 100–109. [Google Scholar]
- McShea, W.J. Forest ecosystems without carnivores: When ungulates rule the world. In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 138–153. [Google Scholar]
- Ray, J.C. Large carnivorous animals as tools for conserving biodiversity: Assumptions and uncertainties. In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 34–56. [Google Scholar]
- Soule, M.E.; Estes, J.A.; Miller, B.; Honnold, D.L. Strongly interacting species. conservation policy, management, and ethics. Bioscience 2005, 55, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.C.; Redford, K.H.; Berger, J.; Steneck, R.S. Conclusion: Is large carnivore conservation equivalent to biodiversity conservation and how can we achieve both? In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 400–428. [Google Scholar]
- Pace, M.L.; Cole, J.J.; Carpenter, S.R.; Kitchell, J.F. Trophic cascades revealed in diverse ecosystems. Trends Ecol. Evol. 1999, 14, 483–488. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Hamback, P.A.; Beckerman, A.P. Trophic cascades in terrestrial systems: A review of the effects of carnivore removals on plants. Am. Nat. 2000, 155, 141–153. [Google Scholar] [CrossRef]
- Hobbs, N.T. Large herbivores as sources of disturbance in ecosystems. In Large Herbivore Ecology, Ecosystem Dynamics and Conservation; Pastor, J., Danell, K., Duncan, P., Bergström, R., Eds.; Cambridge University Press: Cambridge, MA, USA, 2006; pp. 261–288. [Google Scholar]
- Ripple, W.J.; Beschta, R.L. Linking wolves and plants: Aldo Leopold on trophic cascades. Bioscience 2005, 55, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.; Stacey, P.B.; Bellis, L.; Johnson, M.P. A mammalian predator-prey imbalance: Grizzly bear and wolf extinction affect avian neotropical migrants. Ecol. Appl. 2001, 11, 947–960. [Google Scholar] [CrossRef]
- DeVault, T.L.; Rhodes, O.E.; Shivik, J.A. Scavenging by vertebrates: Behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 2003, 102, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.S.; Durant, S.M.; Caro, T.M. Patterns of scavenger arrival at cheetah kills in Serengeti National Park Tanzania. Afr. J. Ecol. 2007, 45, 275–281. [Google Scholar] [CrossRef]
- Selva, N.; Fortuna, M.A. The nested structure of a scavenger community. Proc. R. Soc. B Biol. Sci. 2007, 274, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Selva, N.; Jedrzejewska, B.; Jedrzejewski, W.; Wajrak, A. Scavenging on European bison carcasses in Bialowieza Primeval Forest (eastern Poland). Ecoscience 2003, 10, 303–311. [Google Scholar] [CrossRef]
- Selva, N.; Jedrzejewska, B.; Jedrzejewski, W.; Wajrak, A. Factors affecting carcass use by a guild of scavengers in European temperate woodland. Can. J. Zool. 2005, 83, 1590–1601. [Google Scholar] [CrossRef]
- Wilmers, C.C.; Crabtree, R.L.; Smith, D.W.; Murphy, K.M.; Getz, W.M. Trophic facilitation by introduced top predators: Grey wolf subsidies to scavengers in Yellowstone National Park. J. Anim. Ecol. 2003, 72, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Bump, J.K.; Peterson, R.O.; Vucetich, J.A. Wolves modulate soil nutrient heterogeneity and foliar nitrogen by configuring the distribution of ungulate carcasses. Ecology 2009, 90, 3159–3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danell, K.; Berteaux, D.; Brathen, K.A. Effect of muskox carcasses on nitrogen concentration in tundra vegetation. Arctic 2002, 55, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Melis, C.; Selva, N.; Teurlings, I.; Skarpe, C.; Linnell, J.D.C.; Andersen, R. Soil and vegetation nutrient response to bison carcasses in Bialeowieza Primeval Forest, Poland. Ecol. Res. 2007, 22, 807–813. [Google Scholar] [CrossRef]
- Towne, E.G. Prairie vegetation and soil nutrient responses to ungulate carcasses. Oecologia 2000, 122, 232–239. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Hawlena, D.; Trussell, G.C. Predator control of ecosystem nutrient dynamics. Ecol. Lett. 2010, 13, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Naiman, R.J.; Bilby, R.E.; Schindler, D.E.; Helfield, J.M. Pacific salmon, nutrients, and the dynamics of freshwater and riparian ecosystems. Ecosystems 2002, 5, 399–417. [Google Scholar] [CrossRef]
- Putman, R.J. Carrion and Dung: The Decomposition of Animal Waste; Edward Arnold: London, UK, 1983. [Google Scholar]
- Carter, D.O.; Yellowlees, D.; Tibbett, M. Cadaver decomposition in terrestrial ecosystems. Naturwissenschaften 2007, 94, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Barton, P.S.; Cunningham, S.A.; Lindenmayer, D.B.; Manning, A.D. The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 2013, 171, 761–772. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Bump, J.K.; Webster, C.R.; Vucetich, J.A.; Peterson, R.O.; Shields, J.M.; Powers, M.D. Ungulate Carcasses Perforate Ecological Filters and Create Biogeochemical Hotspots in Forest Herbaceous Layers Allowing Trees a Competitive Advantage. Ecosystems 2009, 12, 996–1007. [Google Scholar] [CrossRef]
- DeVault, T.L.; Brisbin, I.L.; Rhodes, O.E. Factors influencing the acquisition of rodent carrion by vertebrate scavengers and decomposers. Can. J. Zool. 2004, 82, 502–509. [Google Scholar] [CrossRef]
- Ray, R.R.; Seibold, H.; Heurich, M. Invertebrates outcompete vertebrate facultative scavengers in simulated lynx kills in the Bavarian Forest National Park, Germany. Anim. Biodiv. Conserv. 2014, 37, 77–88. [Google Scholar]
- Turner, K.L.; Abernethy, E.F.; Conner, L.M.; Olin, E.; Beasley, J.C. Abiotic and biotic factors modulate carrion fate and vertebrate scavenging communities. Ecology 2017, 98, 2413–2424. [Google Scholar] [CrossRef]
- Nilsen, E.B.; Linnell, J.D.C.; Odden, J.; Andersen, R. Climate, season, and social status modulate the functional response of an efficient stalking predator: The Eurasian lynx. J. Anim. Ecol. 2009, 78, 741–751. [Google Scholar] [CrossRef]
- Odden, J.; Linnell, J.D.C.; Andersen, R. Diet of Eurasian lynx, Lynx lynx, in the boreal forest of southeastern Norway: The relative importance of livestock and hares at low roe deer density. Eur. J. Wildlife Res. 2006, 52, 237–244. [Google Scholar] [CrossRef]
- Novozamsky, I.; Houba, V.J.G.; Temminghoff, E.; van der Lee, J.J. Determination of ‘total’ N and ‘total’ P in a single soil digest. Neth. J. Agric. Sci. 1984, 32, 322–324. [Google Scholar]
- Walinga, I.; Van Der Lee, J.J.; Houba, V.J.G.; Van Vark, W.; Novozamsky, I. Digestion in tubes with H2SO4-salicylic acid- H2O2 and selenium and determination of Ca, K, Mg, N, Na, P, Zn. In Plant Analysis Manual; Walinga, I., Van Der Lee, J.J., Houba, V.J.G., Van Vark, W., Novozamsky, I., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 7–45. [Google Scholar]
- Houba, V.J.G.; Temminghoff, E.J.M.; Gaikhorst, G.A.; van Vark, W. Soil analysis procedures using 0.01 M calcium chloride as extraction reagent. Commun. Soil Sci. Plant Anal. 2000, 31, 1299–1396. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effect Models in S and S-Plus; Springer: New York, NY, USA, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.r-project.org/ (accessed on 12 September 2020).
- Bartoń, K. MuMIn: Multi-Model Inference; R Package Version 1.43.17; 2020. Available online: https://www.rdocumentation.org/packages/MuMIn/versions/1.43.17 (accessed on 12 September 2020).
- Bates, D. lme4: Linear Mixed-Effects Models Using S4 Classes; 0.99875-6. 2007. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 12 September 2020).
- Barton, P.S.; McIntyre, S.; Evans, M.J.; Bump, J.K.; Cunningham, S.A.; Manning, A.D. Substantial long-term effects of carcass addition on soil and plants in a grassy eucalypt woodland. Ecosphere 2016, 7, e01537. [Google Scholar] [CrossRef]
- Andersen, R.; Duncan, P.; Linnell, J.D.C. The European Roe Deer: The Biology of Success; Scandinavian University Press: Oslo, Norway, 1998; Volume 376. [Google Scholar]
- Okarma, H.; Jedrzejewski, W.; Schmidt, K.; Kowalczyk, R.; Jedrzejewska, B. Predation of Eurasian lynx on roe deer and red deer in Bialowieza Primeval Forest, Poland. Acta Theriol. 1997, 42, 203–224. [Google Scholar] [CrossRef] [Green Version]
- Breitenmoser, U.; Breitenmoser-Würsten, C. Der Luchs: Ein Grossraubtier in der Kulturlandschaft; Salm Verlag: Bern, Switzerland, 2008. [Google Scholar]
- Krasinska, M.; Krasinski, Z.A. Body mass and measurements of the European bison during postnatal development. Acta Theriol. 2002, 47, 85–106. [Google Scholar] [CrossRef]
- Klimešová, J.; de Bello, F. CLO-PLA: The database of clonal and bud bank traits of Central European flora. J. Veg. Sci. 2009, 20, 511–516. [Google Scholar] [CrossRef]
- Klimešová, J.; Klimeš, L. Clo-Pla3—Database of Clonal Growth of Plants from Central Europe. 2006. Available online: https://clopla.butbn.cas.cz (accessed on 12 September 2020).
- Tolvanen, A. Differences in Recovery between a Deciduous and an Evergreen Ericaceous Clonal Dwarf Shrub after Simulated Aboveground Herbivory and Belowground Damage. Can. J. Bot. 1994, 72, 853–859. [Google Scholar] [CrossRef]
- Shurin, J.B.; Markel, R.W.; Matthews, B. Comparing trophic cascades across ecosystems. In Trophic Cascades: Predators, Prey and the Changing Dynamics of Nature; Terborgh, J., Estes, J.A., Eds.; Island Press: Washington, DC, USA, 2010; pp. 319–335. [Google Scholar]
- Hayward, M.W.; Edwards, S.; Fancourt, B.A.; Linnell, J.D.C.; Nilsen, E.B. Top-down control of ecosystems and the case for rewilding: Does it all add up? In Rewilding; Pettorelli, N., Durant, S.M., du Toit, J.T., Eds.; Cambridge University Press: Cambridge, MA, USA, 2018; pp. 325–354. [Google Scholar]
- Boitani, L.; Linnell, J.D.C. Bringing large mammals back: Large carnivores in Europe. In Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer: Cham, Switzerland, 2015; pp. 67–84. [Google Scholar]
- Linnell, J.D.C.; Promberger, C.; Boitani, L.; Swenson, J.E.; Breitenmoser, U.; Andersen, R. The linkage between conservation strategies for large carnivores and biodiversity: The view from the “half-full”forests of Europe. In Carnivorous Animals and Biodiversity: Does Conserving One Save the Other? Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 381–398. [Google Scholar]
Figure 1.
Roe deer carcass sites in southeastern Norway. (a,b) Carcass sites visited as soon as the lynx left the kill. Both the carcasses had been fed on by lynx for four nights, and the images were taken two and six days after the lynx had abandoned the kill. (c,d) The same carcass site (c) one month and (d) two years ca. after death. In (a,c,d), it is possible to see the labeled pole marking the center of the carcass.
Figure 1.
Roe deer carcass sites in southeastern Norway. (a,b) Carcass sites visited as soon as the lynx left the kill. Both the carcasses had been fed on by lynx for four nights, and the images were taken two and six days after the lynx had abandoned the kill. (c,d) The same carcass site (c) one month and (d) two years ca. after death. In (a,c,d), it is possible to see the labeled pole marking the center of the carcass.

Figure 2.
Response values in soil with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in southeastern Norway. (a) C/N ratio; (b) nitrogen, N; (c) ammonium, NH4+; (d) nitrate, NO3−; (e) phosphorus, P; (f) phosphate, PO4−; (g) calcium, Ca; (h) potassium, K; (i) magnesium, Mg; (j) sodium, Na. All values are in percentage mass; the boxplots show the median (black line within the box) and quartiles (the boundaries of the box). Whiskers above and below the box indicate the 10 th and 90 th percentiles. Points above and below the whiskers indicate outliers outside the 10 th and 90 th percentiles.
Figure 2.
Response values in soil with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in southeastern Norway. (a) C/N ratio; (b) nitrogen, N; (c) ammonium, NH4+; (d) nitrate, NO3−; (e) phosphorus, P; (f) phosphate, PO4−; (g) calcium, Ca; (h) potassium, K; (i) magnesium, Mg; (j) sodium, Na. All values are in percentage mass; the boxplots show the median (black line within the box) and quartiles (the boundaries of the box). Whiskers above and below the box indicate the 10 th and 90 th percentiles. Points above and below the whiskers indicate outliers outside the 10 th and 90 th percentiles.

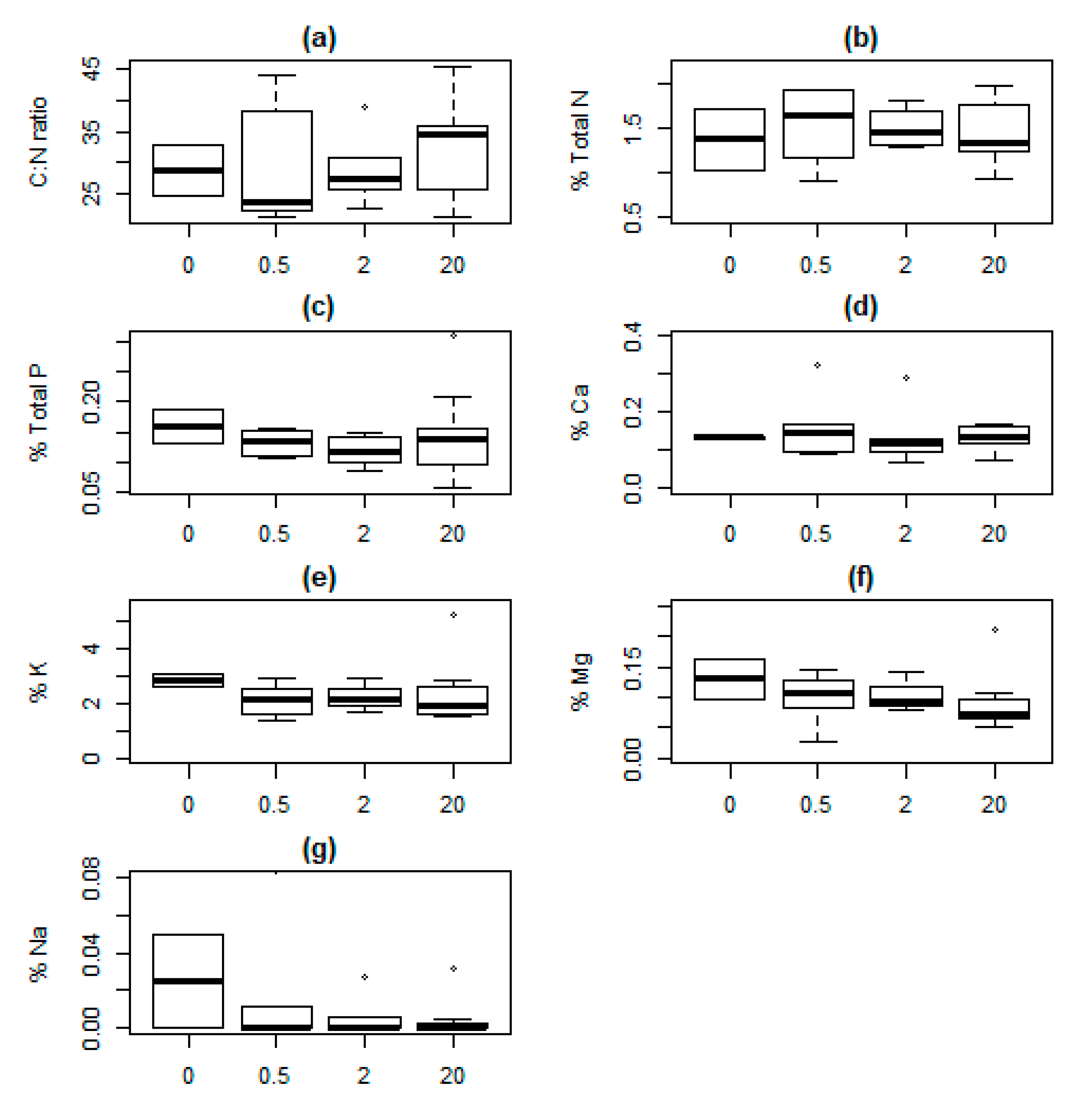
Figure 3.
Response values in Vaccinium myrtillus with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in south-eastern Norway. (a) C:N ratio; (b) Nitrogen, N; (c) Phosphorus, P; (d) Calcium, Ca; (e) Potassium, K; (f) Magnesium, Mg; (g) Sodium, Na. All values are in percentage mass. For additional information, see Figure 2.
Figure 3.
Response values in Vaccinium myrtillus with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in south-eastern Norway. (a) C:N ratio; (b) Nitrogen, N; (c) Phosphorus, P; (d) Calcium, Ca; (e) Potassium, K; (f) Magnesium, Mg; (g) Sodium, Na. All values are in percentage mass. For additional information, see Figure 2.

Figure 4.
Response values in Avenella flexuosa with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in south-eastern Norway. (a) C:N ratio; (b) Nitrogen, N; (c) Phosphorus, P; (d) Calcium, Ca; (e) Potassium, K; (f) Magnesium, Mg; (g) Sodium, Na. All values are in percentage mass. For additional information, see Figure 2.
Figure 4.
Response values in Avenella flexuosa with increasing distance (0, 0.5, 2 and 20 m) from lynx-killed roe deer carcasses in south-eastern Norway. (a) C:N ratio; (b) Nitrogen, N; (c) Phosphorus, P; (d) Calcium, Ca; (e) Potassium, K; (f) Magnesium, Mg; (g) Sodium, Na. All values are in percentage mass. For additional information, see Figure 2.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Set of linear mixed effect models with concentration (g/100 g) of minerals in soil in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) ammonium, NH4+; (d) nitrate, NO3−; (e) phosphorus, P; (f) phosphate, PO4−; (g) calcium, Ca; (h) potassium, K; (i) magnesium, Mg; (j) sodium, Na. The plus sign (+) indicates that the factor was included in the model.
Table 1.
Set of linear mixed effect models with concentration (g/100 g) of minerals in soil in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) ammonium, NH4+; (d) nitrate, NO3−; (e) phosphorus, P; (f) phosphate, PO4−; (g) calcium, Ca; (h) potassium, K; (i) magnesium, Mg; (j) sodium, Na. The plus sign (+) indicates that the factor was included in the model.
| Intercept | Season | Distance a | K b | AICc | Δ AICc c | ωi d | |
|---|---|---|---|---|---|---|---|
| (a) C/N ratio (log) | 2.95 | 3 | −38.70 | 0.00 | 0.634 | ||
| 2.87 | + | 4 | −37.50 | 1.15 | 0.357 | ||
| 2.94 | 0.01 | 4 | −29.20 | 9.50 | 0.006 | ||
| 2.86 | + | 0.01 | 5 | −28.00 | 10.70 | 0.003 | |
| (b) N (square root) | 0.92 | 3 | −35.40 | 0.00 | 0.895 | ||
| 0.92 | + | 4 | −30.60 | 4.80 | 0.081 | ||
| 0.94 | −0.01 | 4 | −27.90 | 7.47 | 0.021 | ||
| 0.95 | + | −0.01 | 5 | −23.00 | 12.32 | 0.002 | |
| (c) NH4+ (log) | −6.25 | 3 | 120.20 | 0.00 | 0.670 | ||
| −6.14 | + | 4 | 121.90 | 1.74 | 0.281 | ||
| −6.18 | −0.04 | 4 | 126.10 | 5.94 | 0.034 | ||
| −6.08 | + | −0.04 | 5 | 127.90 | 7.73 | 0.014 | |
| (d) NO3− (log) | −9.18 | 3 | 231.90 | 0.00 | 0.565 | ||
| −9.11 | + | 4 | 232.90 | 1.01 | 0.341 | ||
| −9.05 | −0.08 | 4 | 236.40 | 4.50 | 0.060 | ||
| −8.98 | + | −0.08 | 5 | 237.40 | 5.58 | 0.035 | |
| (e) P (cube root) | 0.41 | 3 | −241.60 | 0.00 | 0.884 | ||
| 0.40 | + | 4 | −237.50 | 4.13 | 0.112 | ||
| 0.42 | 0.00 | 4 | −230.30 | 11.29 | 0.003 | ||
| 0.40 | + | 0.00 | 5 | −226.00 | 15.56 | 0.000 | |
| (f) PO4− (log) | −7.60 | + | 4 | 261.90 | 0.00 | 0.491 | |
| −7.91 | 3 | 262.40 | 0.44 | 0.394 | |||
| −7.44 | + | −0.09 | 5 | 266.00 | 4.11 | 0.063 | |
| −7.75 | −0.09 | 4 | 266.40 | 4.49 | 0.052 | ||
| (g) Ca (square root) | 0.69 | 3 | −130.50 | 0.00 | 0.945 | ||
| 0.69 | + | 4 | −124.40 | 6.02 | 0.047 | ||
| 0.70 | −0.01 | 4 | −120.90 | 9.59 | 0.008 | ||
| 0.70 | + | −0.01 | 5 | −114.80 | 15.69 | 0.000 | |
| (h) K (log) | −4.25 | 3 | 145.20 | 0.00 | 0.661 | ||
| −4.14 | + | 4 | 146.80 | 1.57 | 0.301 | ||
| −4.20 | −0.03 | 4 | 151.70 | 6.44 | 0.026 | ||
| −4.09 | + | −0.03 | 5 | 153.30 | 8.07 | 0.012 | |
| (i) Mg (square root) | 0.13 | 3 | −285.60 | 0.00 | 0.983 | ||
| 0.13 | + | 4 | −276.60 | 9.01 | 0.011 | ||
| 0.13 | 0.00 | 4 | −275.30 | 10.27 | 0.006 | ||
| 0.14 | + | 0.00 | 5 | −266.30 | 19.34 | 0.000 | |
| (j) Na (cube root) | 0.19 | 3 | −160.00 | 0.00 | 0.865 | ||
| 0.21 | + | 4 | −156.10 | 3.90 | 0.123 | ||
| 0.20 | −0.01 | 4 | −151.00 | 8.95 | 0.010 | ||
| 0.22 | + | −0.01 | 5 | −147.10 | 12.89 | 0.001 |
a The explanatory variable “Distance” was square root-transformed. The models were ranked by the AICc—corrected Akaike information criterion (AIC). The minimum adequate model is on the top of each list; b K, number of parameters; c Δ AICc, difference in Akaike values between the first and the actual model; d ωi, Akaike weights.
Table 2.
Set of linear mixed effect models with concentration of minerals (g/100 g) in Vaccinium myrtillus in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) phosphorus, P; (d) potassium, K; (e) calcium, Ca; (f) magnesium, Mg; (g) sodium, Na. The plus sign (+) indicates that the factor was included in the model. For further abbreviations, see Table 1.
Table 2.
Set of linear mixed effect models with concentration of minerals (g/100 g) in Vaccinium myrtillus in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) phosphorus, P; (d) potassium, K; (e) calcium, Ca; (f) magnesium, Mg; (g) sodium, Na. The plus sign (+) indicates that the factor was included in the model. For further abbreviations, see Table 1.
| Intercept | Season | Distance | K | AICc | Δ AICc | ωi | |
|---|---|---|---|---|---|---|---|
| (a) C:N ratio (log) | 3.37 | 3 | −5.00 | 0.000 | 0.818 | ||
| 3.33 | + | 4 | −1.90 | 3.140 | 0.170 | ||
| 3.34 | 0.01 | 4 | 3.70 | 8.720 | 0.010 | ||
| 3.29 | + | 0.01 | 5 | 7.00 | 12.090 | 0.002 | |
| (b) N (square root) | 1.29 | 3 | −8.50 | 0.000 | 0.896 | ||
| 1.27 | + | 4 | −3.70 | 4.770 | 0.082 | ||
| 1.35 | −0.02 | 4 | −0.90 | 7.640 | 0.020 | ||
| 1.33 | + | −0.02 | 5 | 4.20 | 12.720 | 0.002 | |
| (c) P (cube root) | 0.49 | 3 | −90.80 | 0.000 | 0.939 | ||
| 0.51 | −0.01 | 4 | −84.90 | 5.940 | 0.048 | ||
| 0.49 | + | 4 | −82.20 | 8.650 | 0.012 | ||
| 0.51 | + | −0.01 | 5 | −75.80 | 15.000 | 0.001 | |
| (d) Ca (square root) | 0.91 | 3 | −59.00 | 0.000 | 0.958 | ||
| 0.90 | + | 4 | −52.40 | 6.590 | 0.035 | ||
| 0.93 | −0.01 | 4 | −49.10 | 9.890 | 0.007 | ||
| 0.92 | + | −0.01 | 5 | −42.20 | 16.790 | 0.000 | |
| (e) K (log) | −0.26 | 3 | 29.50 | 0.000 | 0.868 | ||
| −0.28 | + | 4 | 33.60 | 4.130 | 0.110 | ||
| −0.28 | 0.01 | 4 | 37.00 | 7.570 | 0.020 | ||
| −0.30 | + | 0.01 | 5 | 41.40 | 11.950 | 0.002 | |
| (f) Mg (square root) | 0.46 | 3 | −87.30 | 0.000 | 0.968 | ||
| 0.45 | + | 4 | −80.20 | 7.050 | 0.029 | ||
| 0.47 | 0.00 | 4 | −75.90 | 11.370 | 0.003 | ||
| 0.46 | + | 0.00 | 5 | −68.50 | 18.820 | 0.000 | |
| (g) Na (cube root) | 0.13 | 3 | −27.90 | 0.000 | 0.931 | ||
| 0.15 | + | 4 | −22.40 | 5.470 | 0.060 | ||
| 0.15 | −0.01 | 4 | −18.40 | 9.490 | 0.008 | ||
| 0.17 | + | −0.01 | 5 | −12.70 | 15.200 | 0.000 |
Table 3.
Set of linear mixed effect models with concentration of minerals (g/100 g) in Avenella flexuosa in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) phosphorus, P; (d) potassium, K; (e) calcium, Ca; (f) magnesium, Mg; (g) sodium, Na. The plus sign (+) indicates that the factor was included in the model. For further abbreviations, see Table 1.
Table 3.
Set of linear mixed effect models with concentration of minerals (g/100 g) in Avenella flexuosa in southeastern Norway in 2004 as the dependent variable and winter vs. summer (Season) and distance from the center of the carcass (Distance) as the explanatory variables. (a) C/N ratio; (b) nitrogen, N; (c) phosphorus, P; (d) potassium, K; (e) calcium, Ca; (f) magnesium, Mg; (g) sodium, Na. The plus sign (+) indicates that the factor was included in the model. For further abbreviations, see Table 1.
| Intercept | Season | Distance | K | AICc | Δ AICc | ωi | |
|---|---|---|---|---|---|---|---|
| (a) C/N ratio (log) | 3.42 | 3 | 6.30 | 0.000 | 0.903 | ||
| 3.40 | + | 4 | 11.30 | 4.940 | 0.076 | ||
| 3.36 | 0.02 | 4 | 14.10 | 7.730 | 0.019 | ||
| 3.34 | + | 0.02 | 5 | 19.40 | 13.060 | 0.001 | |
| (b) N (square root) | 1.20 | 3 | −4.50 | 0.000 | 0.935 | ||
| 1.20 | + | 4 | 1.30 | 5.800 | 0.051 | ||
| 1.17 | 0.01 | 4 | 4.00 | 8.530 | 0.013 | ||
| 1.16 | + | 0.01 | 5 | 10.10 | 14.570 | 0.001 | |
| (c) P (cube root) | 0.50 | 3 | −59.20 | 0.000 | 0.938 | ||
| 0.49 | + | 4 | −53.60 | 5.570 | 0.058 | ||
| 0.49 | 0.00 | 4 | −48.20 | 10.920 | 0.004 | ||
| 0.48 | + | 0.00 | 5 | −42.20 | 16.970 | 0.000 | |
| (d) Ca (square root) | 0.36 | 3 | −53.80 | 0.000 | 0.948 | ||
| 0.34 | + | 4 | −47.80 | 5.960 | 0.048 | ||
| 0.36 | 0.00 | 4 | −42.70 | 11.080 | 0.004 | ||
| 0.35 | + | 0.00 | 5 | −36.50 | 17.240 | 0.000 | |
| (e) K (log) | 0.73 | 3 | 16.20 | 0.000 | 0.894 | ||
| 0.74 | + | 4 | 20.80 | 4.620 | 0.089 | ||
| 0.69 | 0.01 | 4 | 24.30 | 8.100 | 0.016 | ||
| 0.71 | + | 0.01 | 5 | 29.10 | 12.960 | 0.001 | |
| (f) Mg (square root) | 0.30 | 3 | −56.80 | 0.000 | 0.976 | ||
| 0.30 | + | 4 | −48.80 | 7.970 | 0.018 | ||
| 0.32 | −0.01 | 4 | −46.50 | 10.280 | 0.006 | ||
| 0.32 | + | −0.01 | 5 | −38.20 | 18.640 | 0.000 | |
| (g) Na (cube root) | 0.11 | 3 | 23.00 | 0.000 | 0.923 | ||
| 0.14 | + | 4 | 17.60 | 5.380 | 0.063 | ||
| 0.15 | −0.01 | 4 | 14.50 | 8.460 | 0.013 | ||
| 0.17 | + | −0.01 | 5 | −8.70 | 14.300 | 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Teurlings, I.J.M.; Melis, C.; Skarpe, C.; Linnell, J.D.C. Lack of Cascading Effects of Eurasian Lynx Predation on Roe Deer to Soil and Plant Nutrients. Diversity 2020, 12, 352. https://doi.org/10.3390/d12090352
AMA Style
Teurlings IJM, Melis C, Skarpe C, Linnell JDC. Lack of Cascading Effects of Eurasian Lynx Predation on Roe Deer to Soil and Plant Nutrients. Diversity. 2020; 12(9):352. https://doi.org/10.3390/d12090352
Chicago/Turabian StyleTeurlings, Ivonne J. M., Claudia Melis, Christina Skarpe, and John D. C. Linnell. 2020. "Lack of Cascading Effects of Eurasian Lynx Predation on Roe Deer to Soil and Plant Nutrients" Diversity 12, no. 9: 352. https://doi.org/10.3390/d12090352
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.