Infection Dynamics of Swine Influenza Virus in a Danish Pig Herd Reveals Recurrent Infections with Different Variants of the H1N2 Swine Influenza A Virus Subtype

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Design and Herd Description
2.3. Sample Size Calculation, Sample Collection and Clinical Examinations
2.4. Sample Processing and Nucleic Acid Extraction
2.5. cDNA Synthesis and Pre-Amplification
2.6. High-Throughput Real Time PCR
2.7. RNA Extraction and Real Time RT PCR for the Matrix (M) Gene Assay
2.8. One-Tube Full Genome IAV PCR and Purification of PCR Product
2.9. Next Generation Sequencing Data Analysis
2.10. Analysis of N-Linked Glycosylation Sites of the HA Gene Segment
2.11. Statistical Analysis
3. Results
3.1. Prevalence of IAV
3.2. Recurrent Detection of IAV in Pigs
3.3. Association Between IAV Infection and Clinical Signs
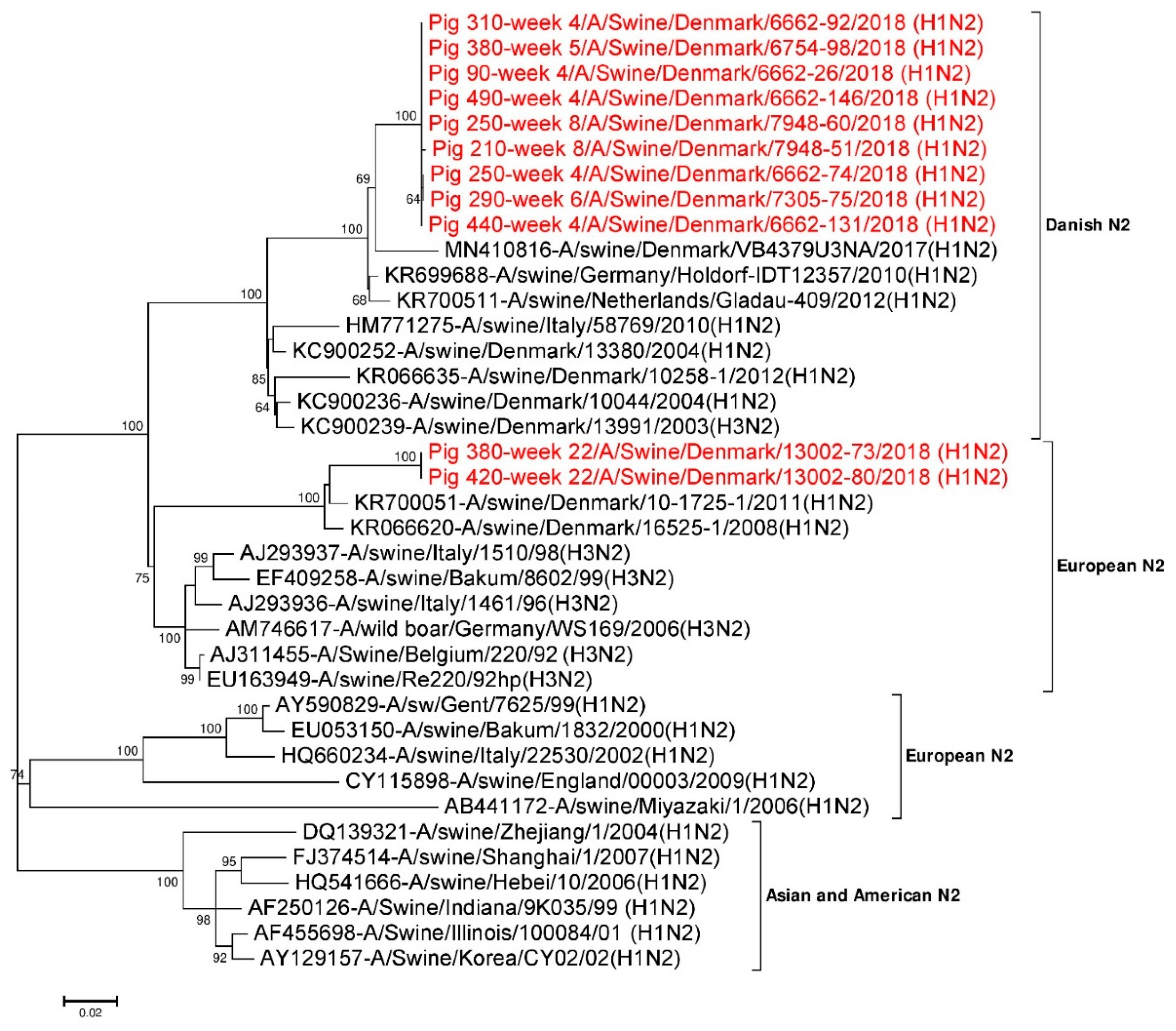
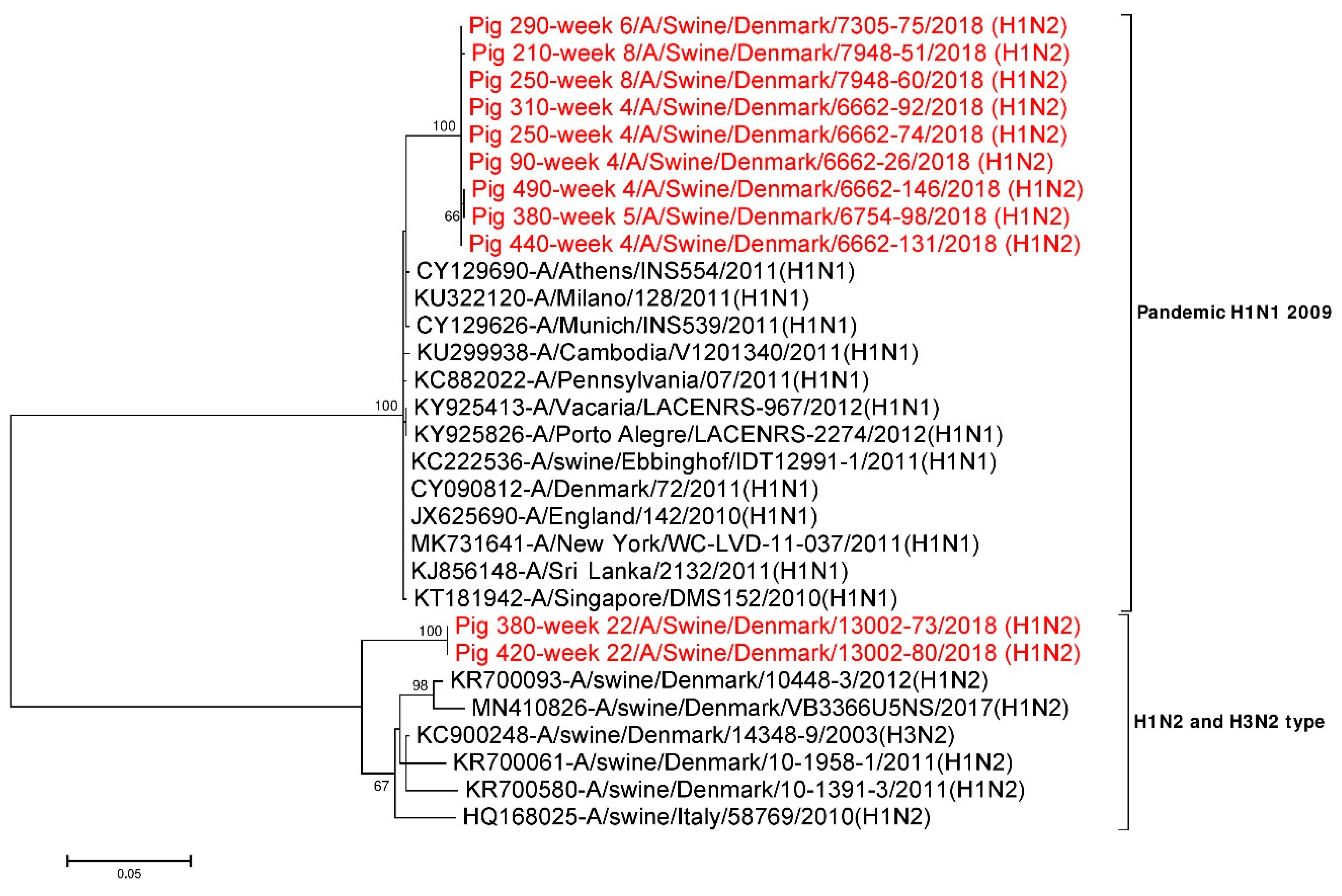
3.4. Subtyping of IAV HA and NA Sequences
3.4.1. Analysis of HA Sequences
3.4.2. Analysis of NA Sequences
3.4.3. Sequence Comparison of the Internal Gene Segments
3.5. Analysis of B- and T-cell Epitopes Present in swIAV Gene Segments
3.6. Analysis of N-Linked Glycosylation Sites (NLG) of the HA Gene Segments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.I.; Holmes, E.C. The evolution of epidemic influenza. Nat. Rev. Genet. 2007, 8, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; Munster, V.J.; Fouchier, R.A. Ecology and evolution of avian influenza viruses. In Genetics and Evolution of Infectious Disease; Elsevier: Amsterdam, The Netherlands, 2011; pp. 729–749. [Google Scholar]
- Kuntz-Simon, G.; Madec, F. Genetic and antigenic evolution of swine influenza viruses in Europe and evaluation of their zoonotic potential. Zoonoses Public Health 2009, 56, 310–325. [Google Scholar] [CrossRef]
- Simon, G.; Larsen, L.E.; Dürrwald, R.; Foni, E.; Harder, T.; Van Reeth, K.; Markowska-Daniel, I.; Reid, S.M.; Dan, A.; Maldonado, J. European surveillance network for influenza in pigs: Surveillance programs, diagnostic tools and Swine influenza virus subtypes identified in 14 European countries from 2010 to 2013. PLoS ONE 2014, 9, e115815. [Google Scholar] [CrossRef]
- Bonin, E.; Quéguiner, S.; Woudstra, C.; Gorin, S.; Barbier, N.; Harder, T.C.; Fach, P.; Hervé, S.; Simon, G. Molecular subtyping of European swine influenza viruses and scaling to high-throughput analysis. Virol J. 2018, 15, 7. [Google Scholar] [CrossRef]
- Pensaert, M.; Ottis, K.; Vandeputte, J.; Kaplan, M.M.; Bachmann, P. Evidence for the natural transmission of influenza A virus from wild ducks to swine and its potential importance for man. Bull. World Health Organ. 1981, 59, 75. [Google Scholar]
- Vincent, A.; Awada, L.; Brown, I.; Chen, H.; Claes, F.; Dauphin, G.; Donis, R.; Culhane, M.; Hamilton, K.; Lewis, N. Review of influenza A virus in swine worldwide: A call for increased surveillance and research. Zoonoses Public Health 2014, 61, 4–17. [Google Scholar] [CrossRef]
- Olsen, C.; Brown, I.; Easterday, B.; Van Reeth, K. Swine influenza. In Diseases of Swine; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J., Eds.; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Simon-Grifé, M.; Martín-Valls, G.E.; Vilar, M.J.; Busquets, N.; Mora-Salvatierra, M.; Bestebroer, T.M.; Fouchier, R.A.; Martín, M.; Mateu, E.; Casal, J. Swine influenza virus infection dynamics in two pig farms; results of a longitudinal assessment. Vet. Res. 2012, 43, 24. [Google Scholar] [CrossRef] [Green Version]
- Madec, F.; Gourreau, J.; Kaiser, C.; Ledantec, J.; Vannier, P.; Aymard, M. Study of the persistence of activity of the H1N1 influenza virus in swine intensive units out of epidemical phases. Comp. Immunol Microb. 1985, 8, 247–258. [Google Scholar] [CrossRef]
- Kyriakis, C.; Rose, N.; Foni, E.; Maldonado, J.; Loeffen, W.; Madec, F.; Simon, G.; Van Reeth, K. Influenza A virus infection dynamics in swine farms in Belgium, France, Italy and Spain, 2006–2008. Vet. Microbiol. 2013, 162, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Ryt-Hansen, P.; Pedersen, A.G.; Larsen, I.; Krog, J.S.; Kristensen, C.S.; Larsen, L.E. Acute Influenza A virus outbreak in an enzootic infected sow herd: Impact on viral dynamics, genetic and antigenic variability and effect of maternally derived antibodies and vaccination. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Ryt-Hansen, P.; Larsen, I.; Kristensen, C.S.; Krog, J.S.; Wacheck, S.; Larsen, L.E. Longitudinal field studies reveal early infection and persistence of influenza A virus in piglets despite the presence of maternally derived antibodies. Vet. Res. 2019, 50, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.I.; Viboud, C.; Vincent, A.L.; Culhane, M.R.; Detmer, S.E.; Wentworth, D.E.; Rambaut, A.; Suchard, M.A.; Holmes, E.C.; Lemey, P. Global migration of influenza A viruses in swine. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pitzer, V.E.; Aguas, R.; Riley, S.; Loeffen, W.L.; Wood, J.L.; Grenfell, B.T. High turnover drives prolonged persistence of influenza in managed pig herds. J. R. Soc. Interface 2016, 13, 20160138. [Google Scholar] [CrossRef]
- Loeffen, W.; Hunneman, W.; Quak, J.; Verheijden, J.; Stegeman, J. Population dynamics of swine influenza virus in farrow-to-finish and specialised finishing herds in the Netherlands. Vet. Microbiol. 2009, 137, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Starick, E.; Lange, E.; Fereidouni, S.; Bunzenthal, C.; Höveler, R.; Kuczka, A.; grosse Beilage, E.; Hamann, H.-P.; Klingelhöfer, I.; Steinhauer, D. Reassorted pandemic (H1N1) 2009 influenza A virus discovered from pigs in Germany. J. Gen. Virol. 2011, 92, 1184–1188. [Google Scholar] [CrossRef]
- Watson, S.J.; Langat, P.; Reid, S.M.; Lam, T.T.-Y.; Cotten, M.; Kelly, M.; Van Reeth, K.; Qiu, Y.; Simon, G.; Bonin, E. Molecular epidemiology and evolution of influenza viruses circulating within European swine between 2009 and 2013. J. Virol. 2015, 89, 9920–9931. [Google Scholar] [CrossRef] [Green Version]
- Howard, W.A.; Essen, S.C.; Strugnell, B.W.; Russell, C.; Barrass, L.; Reid, S.M.; Brown, I.H. Reassortant pandemic (H1N1) 2009 virus in pigs, United Kingdom. Emerg. Infect. Dis. 2011, 17, 1049. [Google Scholar] [CrossRef]
- Krog, J.S.; Hjulsager, C.K.; Larsen, M.A.; Larsen, L.E. Triple-reassortant influenza A virus with H3 of human seasonal origin, NA of swine origin, and internal A (H1N1) pandemic 2009 genes is established in Danish pigs. Influenza Other Respi. Viruses 2017, 11, 298–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebbien, R.; Bragstad, K.; Larsen, L.E.; Nielsen, J.; Bøtner, A.; Heegaard, P.M.; Fomsgaard, A.; Viuff, B.; Hjulsager, C.K. Genetic and biological characterisation of an avian-like H1N2 swine influenza virus generated by reassortment of circulating avian-like H1N1 and H3N2 subtypes in Denmark. Virol. J. 2013, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S. Why are RNA virus mutation rates so damn high? PLoS Biol. 2018, 16, e3000003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Sandt, C.E.; Kreijtz, J.H.; Rimmelzwaan, G.F. Evasion of influenza A viruses from innate and adaptive immune responses. Viruses 2012, 4, 1438–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryt-Hansen, P.; Pedersen, A.G.; Larsen, I.; Kristensen, C.S.; Krog, J.S.; Wacheck, S.; Larsen, L.E. Substantial Antigenic Drift in the Hemagglutinin Protein of Swine Influenza A Viruses. Viruses 2020, 12, 248. [Google Scholar] [CrossRef] [Green Version]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of the structure and function of H1 hemagglutinin of influenza virus. Proc. Jpn. Acad. B. 2012, 88, 226–249. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Marasco, W.A. Structural basis of influenza virus neutralization. Ann. N. Y. Acad. Sci. 2011, 1217, 178. [Google Scholar] [CrossRef] [Green Version]
- Tutykhina, I.; Esmagambetov, I.; Bagaev, A.; Pichugin, A.; Lysenko, A.; Shcherbinin, D.; Sedova, E.; Logunov, D.; Shmarov, M.; Ataullakhanov, R. Vaccination potential of B and T epitope-enriched NP and M2 against influenza A viruses from different clades and hosts. PLoS ONE 2018, 13, e0191574. [Google Scholar] [CrossRef]
- Terajima, M.; Babon, J.A.B.; Ennis, F.A. Cross-reactive human B cell and T cell epitopes between influenza A and B viruses. Virol. J. 2013, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Staneková, Z.; Varečková, E. Conserved epitopes of influenza A virus inducing protective immunity and their prospects for universal vaccine development. Virol. J. 2010, 7, 351. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, J.A.; Kotturi, M.F.; Kim, Y.; Oseroff, C.; Vaughan, K.; Salimi, N.; Vita, R.; Ponomarenko, J.; Scheuermann, R.H.; Sette, A. Pre-existing immunity against swine-origin H1N1 influenza viruses in the general human population. Proc. Natl. Acad. Sci. USA 2009, 106, 20365–20370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, P.; Jang, Y.H.; Kwon, S.B.; Lee, C.M.; Han, G.; Seong, B.L. Glycosylation of hemagglutinin and neuraminidase of influenza a virus as signature for ecological spillover and adaptation among influenza reservoirs. Viruses 2018, 10, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danish Pig Meat Industry. Available online: https://agricultureandfood.dk/danish-agriculture-and-food/danish-pig-meat-industry (accessed on 8 June 2020).
- Structural Development in Danish Pig Production 2019. Available online: https://svineproduktion.dk/publikationer/kilder/notater/2019/1915 (accessed on 8 June 2020).
- Jilani, T.N.; Jamil, R.T.; Siddiqui, A.H. H1N1 Influenza (Swine Flu). In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Dual infections of feeder pigs with porcine reproductive and respiratory syndrome virus followed by porcine respiratory coronavirus or swine influenza virus: A clinical and virological study. Vet. Microbiol. 1996, 48, 325–335. [Google Scholar] [CrossRef]
- Thacker, E.L.; Thacker, B.J.; Janke, B.H. Interaction between Mycoplasma hyopneumoniae and swine influenza virus. J. Clin. Microbiol. 2001, 39, 2525–2530. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Janke, B.H.; Richt, J.A. Swine influenza viruses: A North American perspective. Adv. Virus Res. 2008, 72, 127–154. [Google Scholar]
- Goecke, N.B.; Hjulsager, C.K.; Krog, J.S.; Skovgaard, K.; Larsen, L.E. Development of a high-throughput real-time PCR system for detection of enzootic pathogens in pigs. J. Vet. Diagn. Investig. 2020, 32, 51–64. [Google Scholar] [CrossRef]
- Baekbo, P.J. Raised without antibiotics. In Proceedings of the 25th International Pig Veterinary Society Congress and International PRRS Symposium, Chongqing, China, 11–14 June 2018; p. VI-015/946. [Google Scholar]
- Krog, J.S.; Hjulsager, C.K.; Larsen, L.E. Monitoring of Influenza A Virus in Pig Final Report 2016; Veterinary Institute, Technical University of Denmark: Frederiksberg C, Denmark, 2017. [Google Scholar]
- Nagy, A.; Vostinakova, V.; Pirchanova, Z.; Cernikova, L.; Dirbakova, Z.; Mojzis, M.; Jirincova, H.; Havlickova, M.; Dan, A.; Ursu, K. Development and evaluation of a one-step real-time RT-PCR assay for universal detection of influenza A viruses from avian and mammal species. Arch. Virol. 2010, 155, 665–673. [Google Scholar] [CrossRef]
- Ryt-Hansen, P.; Larsen, I.; Kristensen, C.S.; Krog, J.S.; Larsen, L.E. Limited impact of influenza A virus vaccination of piglets in an enzootic infected sow herd. Res. Vet. Sci. 2019, 127, 47–56. [Google Scholar] [CrossRef]
- Zhou, B.; Donnelly, M.E.; Scholes, D.T.; George, K.S.; Hatta, M.; Kawaoka, Y.; Wentworth, D.E. Single-reaction genomic amplification accelerates sequencing and vaccine production for classical and Swine origin human influenza a viruses. J. Virol. 2009, 83, 10309–10313. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Mount, D.W. Using the basic local alignment search tool (BLAST). CSH Protoc. 2007, 2007, pdb–top17. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squires, R.B.; Noronha, J.; Hunt, V.; García-Sastre, A.; Macken, C.; Baumgarth, N.; Suarez, D.; Pickett, B.E.; Zhang, Y.; Larsen, C.N. Influenza research database: An integrated bioinformatics resource for influenza research and surveillance. Influenza Other Respi. Viruses 2012, 6, 404–416. [Google Scholar] [CrossRef]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J. Influenza Research Database: An integrated bioinformatics resource for influenza virus research. Nucleic Acids Res. 2017, 45, D466–D474. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, G.; Fodor, E. The predicted antigenicity of the haemagglutinin of the 1918 Spanish influenza pandemic suggests an avian origin. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1871–1876. [Google Scholar] [CrossRef]
- Igarashi, M.; Ito, K.; Yoshida, R.; Tomabechi, D.; Kida, H.; Takada, A. Predicting the antigenic structure of the pandemic (H1N1) 2009 influenza virus hemagglutinin. PLoS ONE 2010, 5, e8553. [Google Scholar] [CrossRef] [Green Version]
- Caton, A.J.; Brownlee, G.G.; Yewdell, J.W.; Gerhard, W. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 1982, 31, 417–427. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, B.; Thomas, M.; Sreenivasan, C.C.; Sheng, Z.; Yu, J.; Hause, B.M.; Wang, D.; Francis, D.H.; Kaushik, R.S. Detailed mapping of the linear B Cell epitopes of the hemagglutinin (HA) protein of swine influenza virus. Virology 2018, 522, 131–137. [Google Scholar] [CrossRef]
- Holzer, B.; Martini, V.; Edmans, M.; Tchilian, E. T and B cell immune responses to influenza viruses in pigs. Front. Immunol. 2019, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Baratelli, M.; Pedersen, L.E.; Trebbien, R.; Larsen, L.E.; Jungersen, G.; Blanco, E.; Nielsen, J.; Montoya, M. Identification of cross-reacting T-cell epitopes in structural and non-structural proteins of swine and pandemic H1N1 influenza A virus strains in pigs. J. Gen. Virol. 2017, 98, 895. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.E.; Breum, S.Ø.; Riber, U.; Larsen, L.E.; Jungersen, G. Identification of swine influenza virus epitopes and analysis of multiple specificities expressed by cytotoxic T cell subsets. Virol. J. 2014, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Colman, P.M.; Varghese, J.; Laver, W. Structure of the catalytic and antigenic sites in influenza virus neuraminidase. Nature 1983, 303, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.; Brown, L.; Laver, W. Antigenic and biological characterization of influenza virus neuraminidase (N2) with monoclonal antibodies. Virology 1984, 135, 30–42. [Google Scholar] [CrossRef]
- Laver, W.; Air, G.; Webster, R.; Markoff, L. Amino acid sequence changes in antigenic variants of type A influenza virus N2 neuraminidase. Virology 1982, 122, 450–460. [Google Scholar] [CrossRef]
- Air, G.; Els, M.; Brown, L.; Laver, W.; Webster, R. Location of antigenic sites on the three-dimensional structure of the influenza N2 virus neuraminidase. Virology 1985, 145, 237–248. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2001, 7, 310–322. [Google Scholar]
- Toft, N.; Agger, J.F.; Brunn, J. Measure of association and effect. In Introduction to Veterinary Epidemiology, 4th ed.; Biofolia: Frederiksberg C, Denmark, 2004; pp. 95–107. [Google Scholar]
- Diaz, A.; Marthaler, D.; Corzo, C.; Muñoz-Zanzi, C.; Sreevatsan, S.; Culhane, M.; Torremorell, M. Multiple genome constellations of similar and distinct influenza A viruses co-circulate in pigs during epidemic events. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y. A phylogeny-based global nomenclature system and automated annotation tool for H1 hemagglutinin genes from swine influenza A viruses. mSphere 2016, 1, e00275-16. [Google Scholar] [CrossRef] [Green Version]
- Pecheur, E.; Sainte-Marie, J.; Bienvenüe, A.; Hoekstra, D. Peptides and membrane fusion: Towards an understanding of the molecular mechanism of protein-induced fusion. J. Membrane. Biol. 1999, 167, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Burmeister, W.; Ruigrok, R.; Cusack, S. The 2.2 A resolution crystal structure of influenza B neuraminidase and its complex with sialic acid. EMBO J. 1992, 11, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Colman, P.M.; Hoyne, P.A.; Lawrence, M.C. Sequence and structure alignment of paramyxovirus hemagglutinin-neuraminidase with influenza virus neuraminidase. J. Virol. 1993, 67, 2972–2980. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Pardo, F.O.C.; Wayne, S.; Culhane, M.R.; Perez, A.; Allerson, M.; Torremorell, M. Effect of strain-specific maternally-derived antibodies on influenza A virus infection dynamics in nursery pigs. PLoS ONE 2019, 14, e0210700. [Google Scholar]
- Loeffen, W.; Heinen, P.; Bianchi, A.; Hunneman, W.; Verheijden, J. Effect of maternally derived antibodies on the clinical signs and immune response in pigs after primary and secondary infection with an influenza H1N1 virus. Vet. Immunol. Immunopathol. 2003, 92, 23–35. [Google Scholar] [CrossRef]
- Van Reeth, K.; Gregory, V.; Hay, A.; Pensaert, M. Protection against a European H1N2 swine influenza virus in pigs previously infected with H1N1 and/or H3N2 subtypes. Vaccine 2003, 21, 1375–1381. [Google Scholar] [CrossRef]
- Lu, X.; Liu, F.; Zeng, H.; Sheu, T.; Achenbach, J.E.; Veguilla, V.; Gubareva, L.V.; Garten, R.; Smith, C.; Yang, H. Evaluation of the antigenic relatedness and cross-protective immunity of the neuraminidase between human influenza A (H1N1) virus and highly pathogenic avian influenza A (H5N1) virus. Virology 2014, 454, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; van der Meulen, K.; Van Reeth, K. Prior infection of pigs with a recent human H 3 N 2 influenza virus confers minimal cross-protection against a E uropean swine H 3 N 2 virus. Influenza Other Respi. Viruses 2013, 7, 1260–1268. [Google Scholar] [CrossRef] [Green Version]
- Padilla-Quirarte, H.O.; Guerrero, D.V.L.; Gutierrez-Xicotencatl, M.L.; Esquivel-Guadarrama, F.R. Protective antibodies against influenza proteins. Front. Immunol. 2019, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Rose, N.; Hervé, S.; Eveno, E.; Barbier, N.; Eono, F.; Dorenlor, V.; Andraud, M.; Camsusou, C.; Madec, F.; Simon, G. Dynamics of influenza A virus infections in permanently infected pig farms: Evidence of recurrent infections, circulation of several swine influenza viruses and reassortment events. Vet. Res. 2013, 44, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Wang, Q.; Zhao, F.; Chen, W.; Li, Z. Prediction of biological functions on glycosylation site migrations in human influenza H1N1 viruses. PLoS ONE 2012, 7, e32119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.C.; Wilson, I.A. A perspective on the structural and functional constraints for immune evasion: Insights from influenza virus. J. Mol. Biol. 2017, 429, 2694–2709. [Google Scholar] [CrossRef] [PubMed]
- Skehel, J.; Stevens, D.; Daniels, R.; Douglas, A.; Knossow, M.; Wilson, I.; Wiley, D. A carbohydrate side chain on hemagglutinins of Hong Kong influenza viruses inhibits recognition by a monoclonal antibody. Proc. Natl. Acad. Sci. USA 1984, 81, 1779–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, Y.; Takashita, E.; Sugawara, K.; Matsuzaki, Y.; Muraki, Y.; Hongo, S. Effect of the addition of oligosaccharides on the biological activities and antigenicity of influenza A/H3N2 virus hemagglutinin. J. Virol. 2004, 78, 9605–9611. [Google Scholar] [CrossRef] [Green Version]
- Van Reeth, K.; Labarque, G.; Nauwynck, H.; Pensaert, M. Differential production of proinflammatory cytokines in the pig lung during different respiratory virus infections: Correlations with pathogenicity. Res. Vet. Sci. 1999, 67, 47–52. [Google Scholar] [CrossRef]
- Van Reeth, K. Cytokines in the pathogenesis of influenza. Vet. Microbiol. 2000, 74, 109–116. [Google Scholar] [CrossRef]
- Van Reeth, K. Avian and swine influenza viruses: Our current understanding of the zoonotic risk. Vet. Res. 2007, 38, 243–260. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Age of Piglets/Pigs | Prevalence of IAV (with 95% Confidence Interval) |
|---|---|
| Week 2 | 4.3% (2/46, 0.5–14.8%) |
| Week 3 | 10.5% (4/38, 2.9–24.8%) |
| Week 4 | 33.3% (13/39, 19.1–50.2%) |
| Week 5 | 31.1% (14/45, 18.2–46.7%) |
| Week 6 | 27.3% (12/44, 14.9–42.8%) |
| Week 8 | 14.3% (6/42, 5.4–28.5%) |
| Week 12 | 2.4% (1/41, 0.06–12.9%) |
| Week 22 | 30.8% (4/13, 9.1–61.4%) |
| Total | 76.6% (36/47, 61.9–87.7%) |
| Antigenic Sites (H1 HA Gene Segment) | Amino Acid Changes |
|---|---|
| Cb | S82N |
| Sa | S156K, R158G, G159N, L164I, S165N, G166N |
| Sb | Y188D, R189S |
| Ca1 | G173E, S206T |
| Ca2 | F140S, H141Y, A144S, N145K, E225T |
| Antigenic sites (N2 NA Gene Segment) | Amino Acid Change |
|---|---|
| 1 | - |
| 2a | N199K |
| 2b | N329D, R331G |
| 2c | K344R, S346D |
| 2d | S367N, N368K, L370S |
| 3 | S400R, D402N |
| 4 | Q432L, T434D |
| Pig ID 380 | Gene Segments | Nucleotide Diversity | Amino Acid Diversity |
|---|---|---|---|
| Week 5–Week 22 | PB2 | 5% (114/2280) | 2.5% (19/760) |
| Week 5–Week 22 | PB1 | 4.6% (104 /2274) | 1.8% (14/758) |
| Week 5–Week 22 | PA | 4.7% (101/2151) | 2.5% (18/717) |
| Week 5–Week 22 | NP | 3.7% (55/1497) | 1.8% (9/499) |
| Week 5–Week 22 | M1-M2 | 6.5% (64/982) | 2.8% (10/351) |
| Week 5–Week 22 | M1 | 6.9% (52/759) | 1.2% (3/253) |
| Week 5–Week 22 | M2 | 5.4% (16/294) | 7.1% (7/98) |
| Week 5–Week 22 | NEP-NS1 | 20.5% (173/844) | 19.8% (68/344) |
| Week 5–Week 22 | NS1 | 22.1% (146/660) | 23.6% (52/220) |
| Week 5–Week 22 | NEP | 15.9% (59/372) | 12.9% (16/124) |
| NLG Sites (Amino Acid Position) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HA Sequence | NN ST (20) | NS TD (21) | NV TV (33) | NG TC (94) | NE TS* (Between 125–126) | NN SY (165) | NH TY (198) | NC TT (276) | NG TY (483) | NG SL (541) | Total NLG Sites |
| All pig IDs (week 4 to 8) (Score) | - | +++ | +++ | ++ | - | - | - | + | + | ++ | 6 |
| All pig IDs at week 22 (Score) | - | +++ | +++ | - | ++ | + | - | + | + | ++ | 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatta, T.R.; Ryt-Hansen, P.; Nielsen, J.P.; Larsen, L.E.; Larsen, I.; Chamings, A.; Goecke, N.B.; Alexandersen, S. Infection Dynamics of Swine Influenza Virus in a Danish Pig Herd Reveals Recurrent Infections with Different Variants of the H1N2 Swine Influenza A Virus Subtype. Viruses 2020, 12, 1013. https://doi.org/10.3390/v12091013
Bhatta TR, Ryt-Hansen P, Nielsen JP, Larsen LE, Larsen I, Chamings A, Goecke NB, Alexandersen S. Infection Dynamics of Swine Influenza Virus in a Danish Pig Herd Reveals Recurrent Infections with Different Variants of the H1N2 Swine Influenza A Virus Subtype. Viruses. 2020; 12(9):1013. https://doi.org/10.3390/v12091013
Chicago/Turabian StyleBhatta, Tarka Raj, Pia Ryt-Hansen, Jens Peter Nielsen, Lars Erik Larsen, Inge Larsen, Anthony Chamings, Nicole B. Goecke, and Soren Alexandersen. 2020. "Infection Dynamics of Swine Influenza Virus in a Danish Pig Herd Reveals Recurrent Infections with Different Variants of the H1N2 Swine Influenza A Virus Subtype" Viruses 12, no. 9: 1013. https://doi.org/10.3390/v12091013