A Comprehensive Study of Cutaneous Fibropapillomatosis in Free-Ranging Roe Deer (Capreolus capreolus) and Red Deer (Cervus elaphus): from Clinical Manifestations to Whole-Genome Sequencing of Papillomaviruses


Abstract
:1. Introduction
2. Materials and Methods
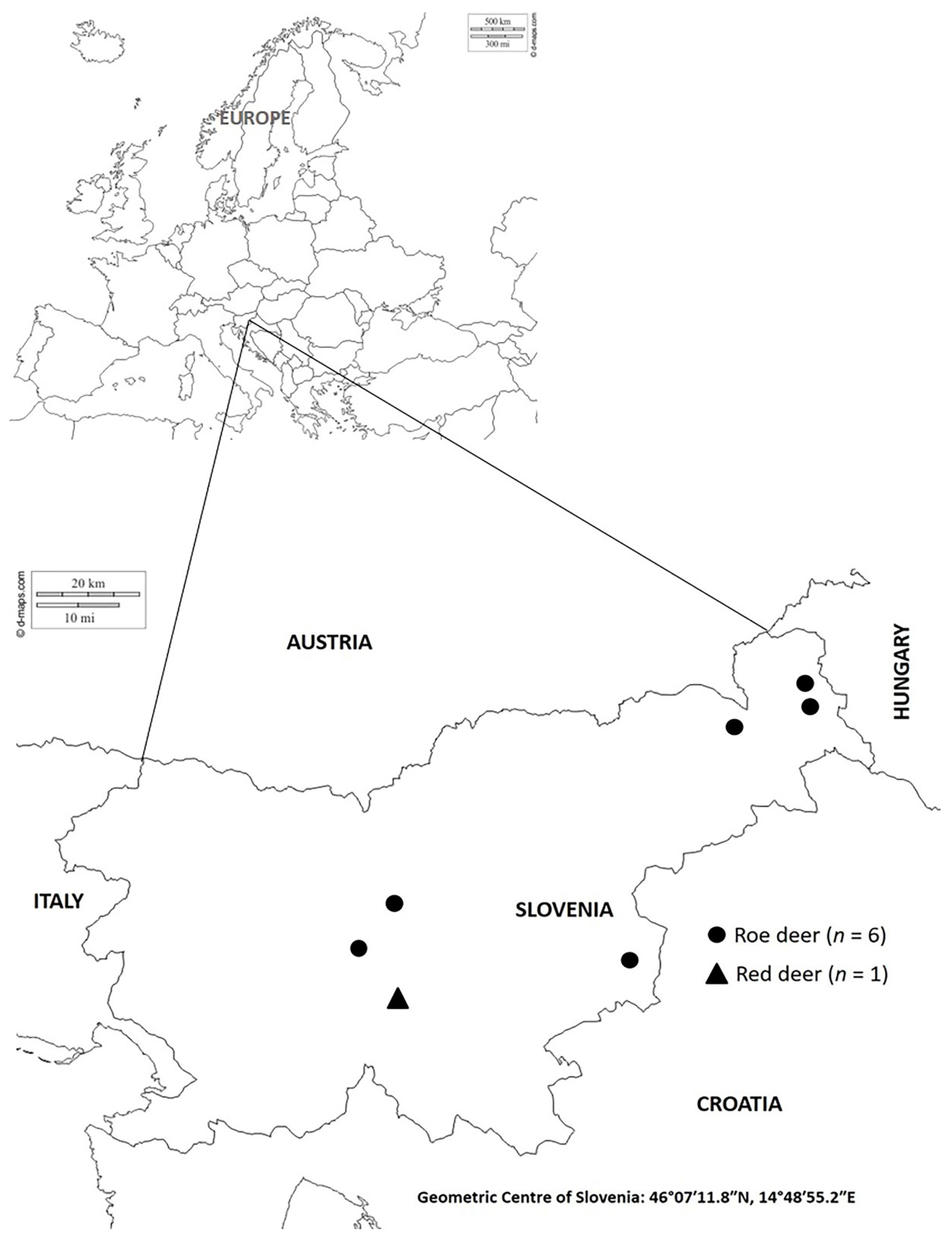
2.1. Samples
2.2. Histopathology
2.3. PCR and Sanger Sequencing of the Partial L1 Gene
2.4. Whole-Genome Sequencing
2.5. Bioinformatic Analysis of NGS Data
2.6. Phylogenetic Analysis
2.7. Data Availability Statement
3. Results
3.1. Gross Pathology and Histopathology
3.2. PCR and Sanger Sequencing of the Partial L1 Gene
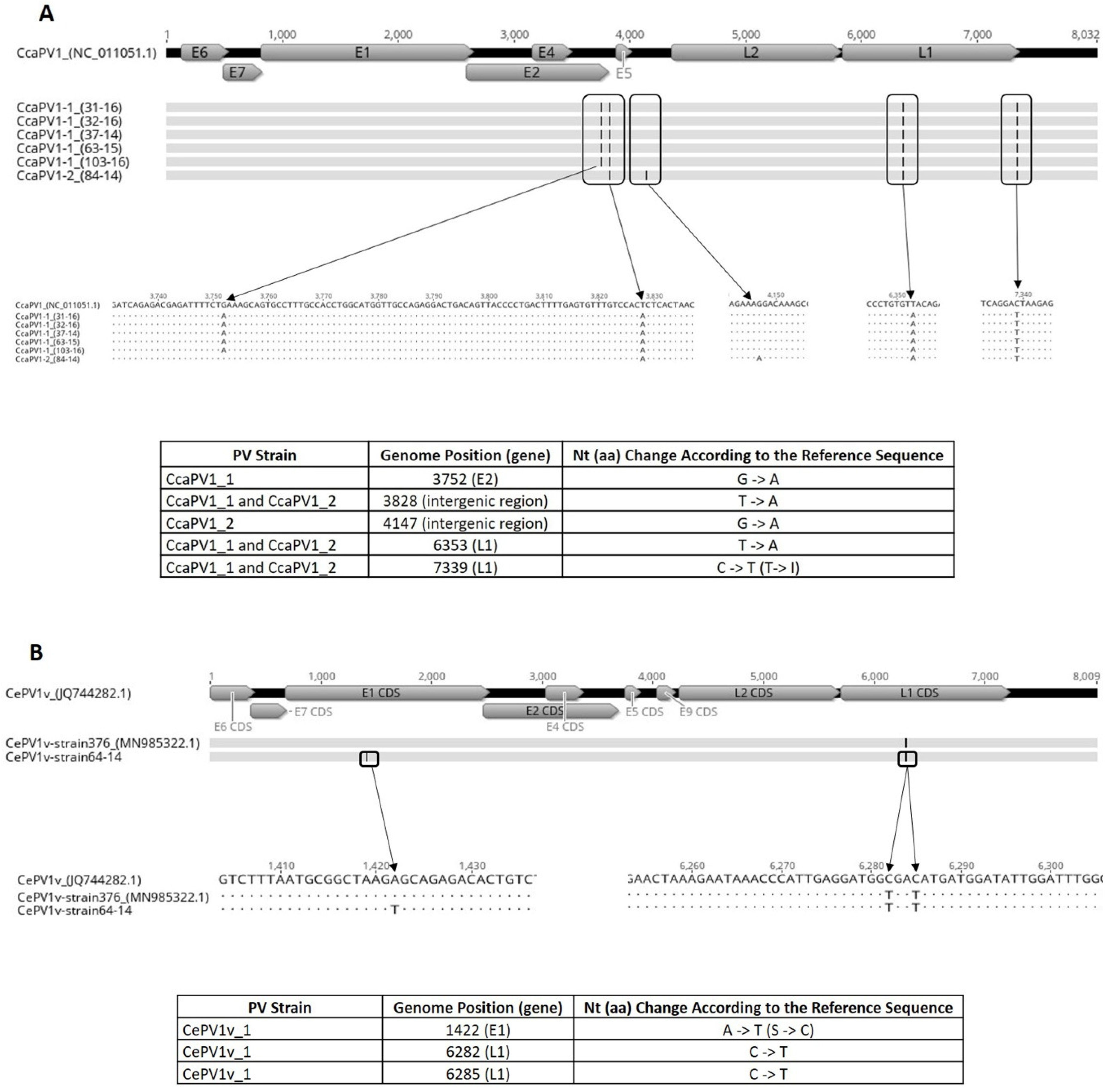
3.3. Whole-Genome Sequencing and Genome Analysis
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ICTV. Papillomaviridae. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/dsdna-viruses/w/papillomaviridae (accessed on 5 February 2020).
- Munday, J.; Pesavento, P. Papillomaviridae and Polyomaviridae. In Fenner’s Veterinary Virology, 5th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 229–243. [Google Scholar]
- Van Doorslaer, K. Evolution of the Papillomaviridae. Virology 2013, 445, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Doorslaer, K.; Chen, Z.; Bernard, H.-U.; Chan, P.K.S.; DeSalle, R.; Dillner, J.; Forslund, O.; Haga, T.; McBride, A.A.; Villa, L.L.; et al. ICTV Virus Taxonomy Profile: Papillomaviridae. J. Gen. Virol. 2018, 99, 989–990. [Google Scholar] [CrossRef] [PubMed]
- Daudt, C.; Da Silva, F.R.C.; Lunardi, M.; Alves, C.B.D.T.; Weber, M.N.; Cibulski, S.P.; Alfieri, A.F.; Alfieri, A.A.; Canal, C.W. Papillomaviruses in ruminants: An update. Transbound. Emerg. Dis. 2018, 65, 1381–1395. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Vaatstra, B.L.; Dunowska, M.; Laurie, R.E.; Hills, S.F.K. Genomic characterization of a novel Epsilonpapillomavirus associated with pigmented papillomas in a red deer (Cervus elaphus). Virus Genes 2016, 52, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Borzacchiello, G.; Roperto, F. Bovine papillomaviruses, papillomas and cancer in cattle. Vet. Res. 2008, 39, 45. [Google Scholar] [CrossRef] [Green Version]
- Campo, M.S. Animal models of papillomavirus pathogenesis. Virus Res. 2002, 89, 249–261. [Google Scholar] [CrossRef]
- Munday, J.S. Bovine and Human Papillomaviruses. Vet. Pathol. 2014, 51, 1063–1075. [Google Scholar] [CrossRef] [Green Version]
- Frias-De-Diego, A.; Jara, M.; Escobar, L.E. Papillomavirus in Wildlife. Front. Ecol. Evol. 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Garcês, A.; Pires, I.; Savini, F.; Scagliarini, A.; Gallina, L. Cutaneous Fibropapilloma in a Red Deer (Cervus elaphus) Associated with Cervus elaphus Papillomavirus in Portugal. J. Wildl. Dis. 2020, 56, 636–639. [Google Scholar] [CrossRef]
- Gil Da Costa, R.M.; Peleteiro, M.C.; Pires, M.; DiMaio, D. An Update on Canine, Feline and Bovine Papillomaviruses. Transbound. Emerg. Dis. 2016, 64, 1371–1379. [Google Scholar] [CrossRef]
- Savini, F.; Molin, E.D.; Gallina, L.; Casà, G.; Scagliarini, A. Papillomavirus in healthy skin and mucosa of wild ruminants in the Italian Alps. J. Wildl. Dis. 2016, 52, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Scagliarini, A.; Gallina, L.; Battilani, M.; Turrini, F.; Savini, F.; Lavazza, A.; Chiari, M.; Coradduzza, E.; Peli, A.; Erdélyi, K.; et al. Cervus elaphus papillomavirus (CePV1): New insights on viral evolution in deer. Vet. Microbiol. 2013, 165, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Rajský, D.; Rajský, M.; Garaj, P.; Kropil, R.; Ivan, M.; Vodnansky, M.; Hanzal, V.; Erdélyi, K. Emergence and expansion of roe deer (Capreolus capreolus) fibropapillomatosis in Slovakia. Eur. J. Wildl. Res. 2015, 62, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Erdélyi, K.; Dencsö, L.; Lehoczki, R.; Heltai, M.; Sonkoly, K.; Csányi, S.; Solymosi, N. Endemic papillomavirus infection of roe deer (Capreolus capreolus). Vet. Microbiol. 2009, 138, 20–26. [Google Scholar] [CrossRef]
- Erdélyi, K.; Gál, J.; Sugár, L.; Ursu, K.; Forgách, P.; Szeredi, L.; Steineck, T. Papillomavirus-associated fibropapillomas of red deer (Cervus elaphus). Acta Vet. Hung. 2009, 57, 337–344. [Google Scholar] [CrossRef]
- Rector, A.; Van Ranst, M. Animal papillomaviruses. Virology 2013, 445, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Sundberg, J.P.; Nielsen, S.W. Deer fibroma: A review. Can. Vet. J. 1981, 22, 385–388. [Google Scholar]
- Erdélyi, K.; Bálint, Á.; Dencsö, L.; Dán, Á.; Ursu, K. Characterisation of the first complete genome sequence of the roe deer (Capreolus capreolus) papillomavirus. Virus Res. 2008, 135, 307–311. [Google Scholar] [CrossRef]
- Kral, J.; Bandouchova, H.; Brichta, J.; Kovacova, V.; Ondracek, K.; Osickova, J.; Hrubá, H.; Hutařová, Z.; Kominkova, M.; Cernei, N.; et al. Papillomavirus infection of roe deer in the Czech Republic and fibropapilloma-associated levels of metallothionein, zinc, and oxidative stress. Acta Vet. Brno 2015, 84, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Gallina, L.; Savini, F.; Casà, G.; Bertoletti, I.; Bianchi, A.; Gibelli, L.R.; Lelli, D.; Lavazza, A.; Scagliarini, A. Epitheliotropic Infections in Wildlife Ruminants From the Central Alps and Stelvio National Park. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Flajšman, K.; Jerina, K.; Pokorny, B. Age-related effects of body mass on fertility and litter size in roe deer. PLoS ONE 2017, 12, e0175579. [Google Scholar] [CrossRef] [Green Version]
- Lange, C.E.; Tobler, K.; Brandes, K.; Breithardt, K.; Ordeix, L.; Von Bomhard, W.; Favrot, C. Canine inverted papillomas associated with DNA of four different papillomaviruses. Vet. Dermatol. 2009, 21, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forslund, O.; Antonsson, A.; Nordin, P.; Stenquist, B.; Hansson, B.G. A broad range of human papillomavirus types detected with a general PCR method suitable for analysis of cutaneous tumours and normal skin The GenBank accession numbers of the sequences reported in this paper are AF121419 to AF121434. J. Gen. Virol. 1999, 80, 2437–2443. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Boil. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubble, M. Capreolus capreolus. In The Atlas of European Mammals (Poyser Natural History); Mitchell-Jone, A.J., Bogdanowicz, W., Krystufek, B., Reijnders, P.J.H., Spitzenberger, F., Stubbe, C., Thissen, J.B.M., Vohralík, V., Zima, J., Eds.; Academic Press: Cambridge, MA, USA, 1999; Volume 1. [Google Scholar]
- Wilson, D.E.; Ruff, S. The Smithsonian Book of North American Mammals. J. Mammal. 2000, 81, 621–622. [Google Scholar] [CrossRef]
- Hoffmann, R.S.; Corbet, G.B. Mammals of the Palaeartic Region: A Taxonomic review. J. Mammal. 1979, 60, 656–657. [Google Scholar] [CrossRef]
- Koubek, P.; Zima, J. Cervus elaphus. In The Atlas of European Mammals (Poyser Natural History); Mitchell-Jone, A.J., Bogdanowicz, W., Krystufek, B., Reijnders, P.J.H., Spitzenberger, F., Stubbe, C., Thissen, J.B.M., Vohralík, V., Zima, J., Eds.; Academic Press: Cambridge, MA, USA, 1999; Volume 1. [Google Scholar]
- Livet, J. Hoofed Mammals. Barcelona. In Handbook of the Mammals of the World; Wilson, D.E., Mittermeier, R.A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; Volume 31, pp. 621–623. [Google Scholar]
- Kocsner, T. Skin Fibromatosis of Roe Deer. Master’s Thesis, University of Veterinary Sciences, Budapest, Hungary, 2001. [Google Scholar]
- Erdélyi, K. Fibropapillomatosis of Roe Deer (Capreolus capreolus) in Hungary. Ph.D. Thesis, Szent István University, Faculty of Agricultural and Environmental Sciences, Gödöllő, Hungary, 2010. [Google Scholar]
- Kureljušić, B.; Savić, B.; Pavlović, I.; Jezdimirović, N.; Kureljušić, J.; Stanojević, S.; Minić, S.; Ivetić, V. Fibromatosis of the roe deer (Capreolus capreolus). In Proceedings of the International Symposium on Hunting, Modern Aspects of Sustainable Management of Game Population, Zemun-Belgrade, Serbia, 22–24 June 2012; pp. 110–112. [Google Scholar]
- Farkaš, F.; Konjević, D.; Grabarević, Ž.; Janicki, Z.; Slavica, A.; Sabočanec, R. Roe deer (Capreolus capreolus) warts—Fibromas, papillomas or fibropapillomas. Acta Clin. Croat. 2012, 51, 179. [Google Scholar]
- Stergar, M.; Jonozovič, M.; Jerina, K. Območja razširjenosti in relativne gostote avtohtonih vrst parkljarjev v Sloveniji. Gozd. Vestn. 2009, 67, 367–380. [Google Scholar]
- Jerina, K. Prostorska razširjenost, vitalnost in populacijska dinamika prostoživečih vrst parkljarjev v Sloveniji. Digit. Knj. Slov. 2010. Available online: http://www.dlib.si/?URN=URN:NBN:SI:DOC-KQTSA7BQ (accessed on 7 September 2020).
- Bernard, H.-U.; Burk, R.D.; Chen, Z.; Van Doorslaer, K.; Hausen, H.Z.; De Villiers, E.-M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Bravo, I.G.; De Sanjose, S.; Gottschling, M. The clinical importance of understanding the evolution of papillomaviruses. Trends Microbiol. 2010, 18, 432–438. [Google Scholar] [CrossRef] [PubMed]
- De Villiers, E.-M.; Fauquet, C.; Broker, T.R.; Bernard, H.-U.; Hausen, H.Z. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschling, M.; Göker, M.; Stamatakis, A.; Bininda-Emonds, O.R.P.; Nindl, I.; Bravo, I.G. Quantifying the Phylodynamic Forces Driving Papillomavirus Evolution. Mol. Boil. Evol. 2011, 28, 2101–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murahwa, A.T.; Nindo, F.; Onywera, H.; Meiring, T.L.; Martin, D.P.; Williamson, A.-L. Evolutionary dynamics of ten novel Gamma-PVs: Insights from phylogenetic incongruence, recombination and phylodynamic analyses. BMC Genom. 2019, 20, 368. [Google Scholar] [CrossRef]
- Murahwa, A.T.; Tshabalala, M.; Williamson, A.-L. Recombination Between High-Risk Human Papillomaviruses and Non-Human Primate Papillomaviruses: Evidence of Ancient Host Switching Among Alphapapillomaviruses. J. Mol. Evol. 2020, 88, 453–462. [Google Scholar] [CrossRef]
- Gottschling, M.; Stamatakis, A.; Nindl, I.; Stockfleth, E.; Alonso, Á.; Bravo, I.G. Multiple Evolutionary Mechanisms Drive Papillomavirus Diversification. Mol. Biol. Evol. 2007, 24, 1242–1258. [Google Scholar] [CrossRef] [Green Version]
- Bocaneti, F.; Altamura, G.; Corteggio, A.; Velescu, E.; Roperto, F.; Borzacchiello, G. Bovine Papillomavirus: New Insights into an Old Disease. Transbound. Emerg. Dis. 2014, 63, 14–23. [Google Scholar] [CrossRef]
- Hafner, M.; Pičulin, I.; Artnak, M.; Leskovic, B. Območja Razširjenosti, Habitatne Potrebe in Prehranske Značilnosti Divjadi. In Varovanje in Urejanje Življenskega Okolja Divjadi; Lovska Zveza Slovenije: Ljubljana, Slovenia, 2014; pp. 149–181. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Species/Year/Month of Harvesting | Sex | Age (in Years) |
|---|---|---|
| Roe deer/2014/August | Male | 2 |
| Roe deer/2014/December | Male | 3 |
| Roe deer/2015/October | Female | 6 |
| Roe deer/2016/May | Male | 6 |
| Roe deer/2016/May | Male | 3 |
| Roe deer/2016/December | Female | 6 |
| Red deer/2016/September | Female | 1 |
| PV Strain | Total Reads | Mapped Reads | Mean Read Length |
|---|---|---|---|
| CcaPV1-1 (31-16) | 197,354 | 312 | 326 |
| CcaPV1-1 (32-16) | 229,894 | 737 | 302 |
| CcaPV1-1 (103-16) | 206,940 | 1519 | 279 |
| CcaPV1-1 (63-15) | 213,479 | 241 | 309 |
| CcaPV1-1 (37-14) | 254,725 | 380 | 292 |
| CcaPV1-2 (84-14) | 317,345 | 636 | 278 |
| CePV1v-1 (64-16) | 2,867,278 | 10,010 | 282 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kmetec, J.; Kuhar, U.; Fajfar, A.G.; Vengušt, D.Ž.; Vengušt, G. A Comprehensive Study of Cutaneous Fibropapillomatosis in Free-Ranging Roe Deer (Capreolus capreolus) and Red Deer (Cervus elaphus): from Clinical Manifestations to Whole-Genome Sequencing of Papillomaviruses. Viruses 2020, 12, 1001. https://doi.org/10.3390/v12091001
Kmetec J, Kuhar U, Fajfar AG, Vengušt DŽ, Vengušt G. A Comprehensive Study of Cutaneous Fibropapillomatosis in Free-Ranging Roe Deer (Capreolus capreolus) and Red Deer (Cervus elaphus): from Clinical Manifestations to Whole-Genome Sequencing of Papillomaviruses. Viruses. 2020; 12(9):1001. https://doi.org/10.3390/v12091001
Chicago/Turabian StyleKmetec, Jernej, Urška Kuhar, Aleksandra Grilc Fajfar, Diana Žele Vengušt, and Gorazd Vengušt. 2020. "A Comprehensive Study of Cutaneous Fibropapillomatosis in Free-Ranging Roe Deer (Capreolus capreolus) and Red Deer (Cervus elaphus): from Clinical Manifestations to Whole-Genome Sequencing of Papillomaviruses" Viruses 12, no. 9: 1001. https://doi.org/10.3390/v12091001