Defining New Pathways to Manage the Ongoing Emergence of Bat Rabies in Latin America

 , , , , , ,
, , , , , ,
Abstract
:
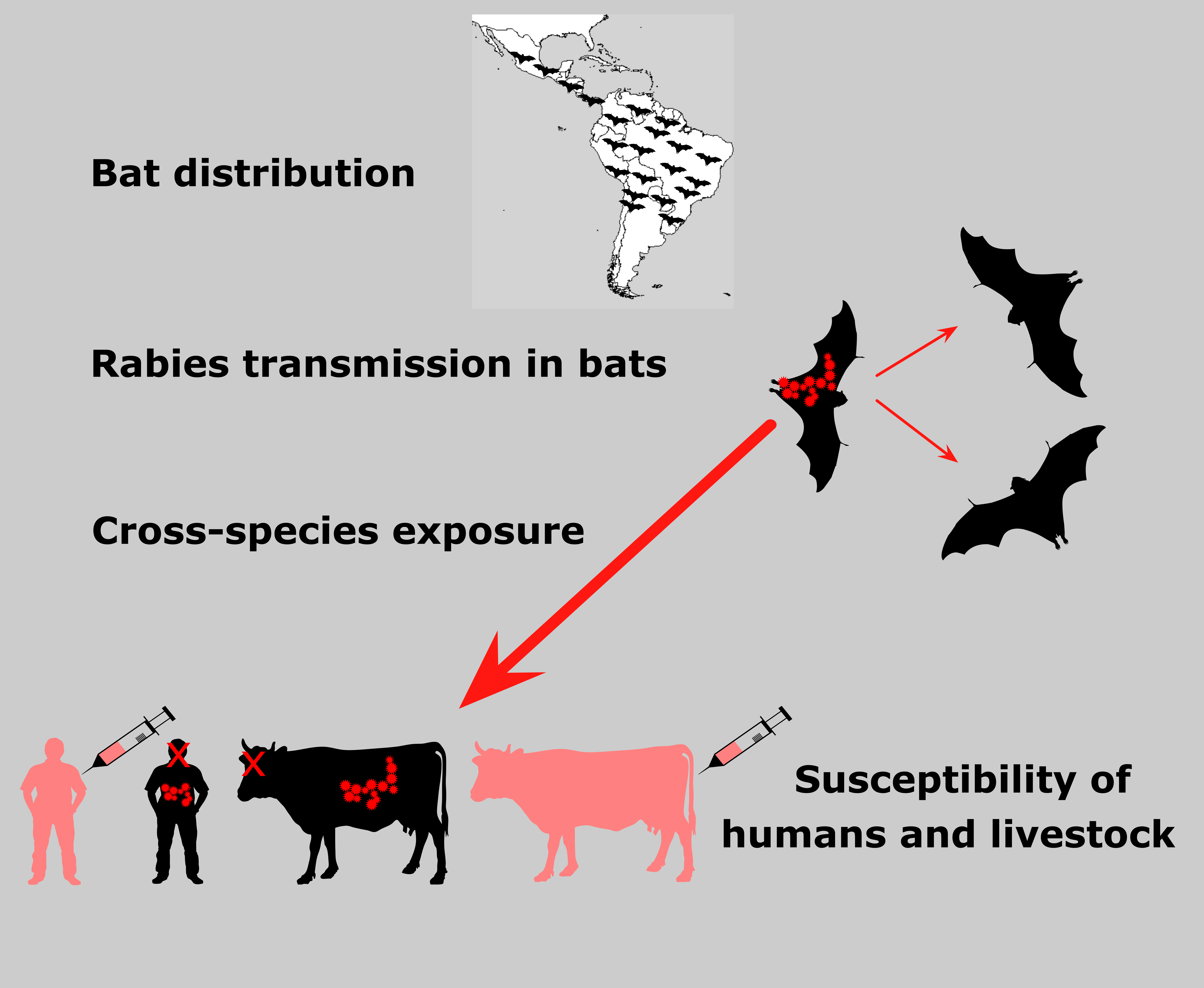{kind=link}
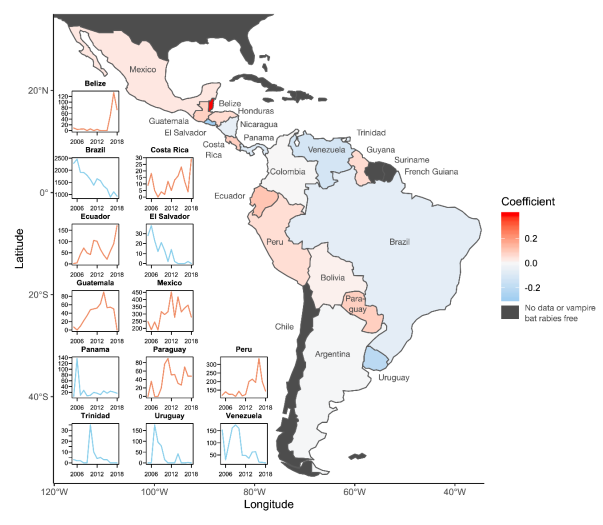{kind=link}
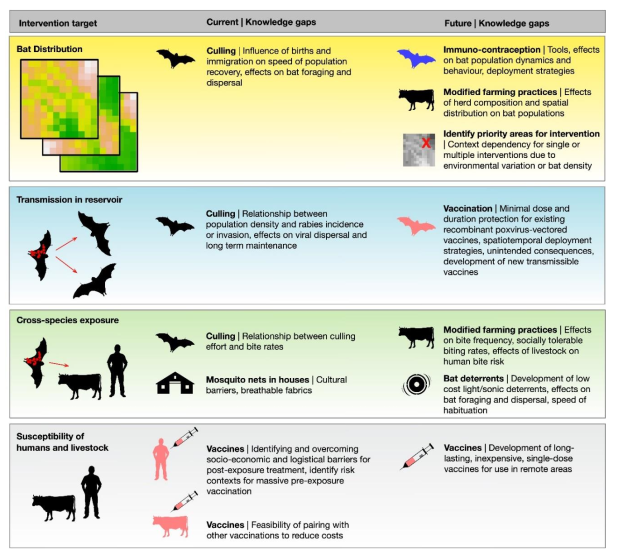{kind=link}
1. Introduction
2. Reservoir Host Distribution
2.1. Current Knowledge
2.2. Current Management Practices
2.3. Improving Management
3. Transmission Dynamics within the Reservoir
3.1. Current Knowledge
3.2. Current Management Practices
3.3. Improving Management
4. Cross-Species Exposures
4.1. Current Knowledge
4.2. Current Management Practices
4.3. Improving Management
5. Susceptibility of Recipient Hosts
5.1. Current Knowledge
5.2. Current Management Practices
5.3. Improving Management
6. Conclusions and Future Steps
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Velasco-Villa, A.; Escobar, L.E.; Sanchez, A.; Shi, M.; Streicker, D.G.; Gallardo-Romero, N.F.; Vargas-Pino, F.; Gutierrez-Cedillo, V.; Damon, I.; Emerson, G. Successful strategies implemented towards the elimination of canine rabies in the Western Hemisphere. Antiviral Res. 2017, 143, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Benavides, J.A.; Rojas Paniagua, E.; Hampson, K.; Valderrama, W.; Streicker, D.G. Quantifying the burden of vampire bat rabies in Peruvian livestock. PLoS Negl. Trop. Dis. 2017, 11, e0006105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigilato, M.A.N.; Clavijo, A.; Knobl, T.; Silva, H.M.T.; Cosivi, O.; Schneider, M.C.; Leanes, L.F.; Belotto, A.J.; Espinal, M.A. Progress towards eliminating canine rabies: policies and perspectives from Latin America and the Caribbean. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2013, 368, 20120143. [Google Scholar] [CrossRef] [PubMed]
- Fenton, B.M.; Streicker, D.G.; Racey, P.A.; Tuttle, M.D.; Medellin, R.A.; Daley, M.J.; Recuenco, S.; Bakker, K.M. Knowledge gaps about rabies transmission from vampire bats to humans. Nat. Ecol. Evol. 2020, 4, 517–518. [Google Scholar] [CrossRef]
- World Health Organization. Expert Consultation on Rabies: Second Report; World Health Organization: Geneva, Switzerland, 2013; Volume 982, pp. 1–139. [Google Scholar]
- Johnson, N.; Aréchiga-Ceballos, N.; Aguilar-Setien, A. Vampire bat rabies: Ecology, epidemiology and control. Viruses 2014, 6, 1911–1928. [Google Scholar] [CrossRef] [Green Version]
- de Paula Silva, N.; Araújo Andrade, E.; Cardoso, D.; de Souza Guimarães, R.; Borges Silva, M.; Gomes Nascimento, K.K.; Xavier, D.; Abel, I. Crab fisherman communities in north Brazil: A new high risk population for vampire bat rabies. bioRxiv 2019, 590083. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, C.; Achkar, S.M.; Kotait, I. First report of rabies in vampire bats (Desmodus rotundus) in an urban area, Ubatuba, São Paulo state, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2007, 49, 389–390. [Google Scholar] [CrossRef] [Green Version]
- Castilho, J.G.; de Souza, D.N.; Oliveira, R.N.; Carnieli, P.; Batista, H.B.C.R.; Pereira, P.M.C.; Achkar, S.M.; Macedo, C.I. The Epidemiological Importance of Bats in the Transmission of Rabies to Dogs and Cats in the State of São Paulo, Brazil, Between 2005 and 2014. Zoonoses Public Health 2017, 64, 423–430. [Google Scholar] [CrossRef]
- Benavides, J.A.; Valderrama, W.; Streicker, D.G. Spatial expansions and travelling waves of rabies in vampire bats. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160328. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.N.; Papeş, M.; van Den Bussche, R.A. Present and potential future distribution of common Vampire bats in the Americas and the associated risk to cattle. PLoS ONE 2012, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Streicker, D.G.; Winternitz, J.C.; Satterfield, D.A.; Condori-Condori, R.E.; Broos, A.; Tello, C.; Recuenco, S.; Velasco-Villa, A.; Altizer, S.; Valderrama, W. Host–pathogen evolutionary signatures reveal dynamics and future invasions of vampire bat rabies. Proc. Natl. Acad. Sci. USA 2016, 113, 10926–10931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, A.K.; Olival, K.J.; Morán, D.; Lopez, M.R.; Alvarez, D.; Navarrete-Macias, I.; Liang, E.; Simmons, N.B.; Lipkin, W.I.; Daszak, P.; et al. Viral diversity, prey preference, and bartonella prevalence in Desmodus rotundus in Guatemala. Ecohealth 2016, 13, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Bergner, L.M.; Orton, R.J.; Benavides, J.A.; Becker, D.J.; Tello, C.; Biek, R.; Streicker, D.G. Demographic and environmental drivers of metagenomic viral diversity in vampire bats. Mol. Ecol. 2020, 29, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, R.D.; Elias, D.J.; Mitchell, G.C. Effects of vampire bat control on bovine milk production. J. Wildl. Manage. 1977, 41, 736. [Google Scholar] [CrossRef]
- Schmidt, K.M.; Badger, D.D. Some social and economic aspects in controlling vampire bats. Proc. Okla. Acad. Sci. 1979, 59, 112–114. [Google Scholar]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proc. R. Soc. B Biol. Sci. 2014, 282, 20142124. [Google Scholar] [CrossRef] [Green Version]
- Bobrowiec, P.E.D.; Lemes, M.R.; Gribel, R. Prey preference of the common vampire bat (Desmodus rotundus, Chiroptera) using molecular analysis. J. Mammal. 2015, 96, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Streicker, D.G.; Allgeier, J.E. Foraging choices of vampire bats in diverse landscapes: potential implications for land-use change and disease transmission. J. Appl. Ecol. 2016, 53, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.N.; Monteiro, A.M. V Cattle rabies in the State of São Paulo and its spatial distribution from 1992 to 2003. Arq. Bras. Med. Vet. Zootec. 2011, 63, 279–286. [Google Scholar] [CrossRef] [Green Version]
- De Andrade, F.A.G.; Gomes, M.N.; Uieda, W.; Begot, A.L.; Ramos, O.D.S.; Fernandes, M.E.B. Geographical analysis for detecting high-risk areas for bovine/human rabies transmitted by the common hematophagous bat in the Amazon region, Brazil. PLoS ONE 2016, 11, 1–15. [Google Scholar] [CrossRef]
- Gonçalves, F.; Fischer, E.; Dirzo, R. Forest conversion to cattle ranching differentially affects taxonomic and functional groups of Neotropical bats. Biol. Conserv. 2017, 210, 343–348. [Google Scholar] [CrossRef]
- Avila-Flores, R.; Bolaina-Badal, A.L.; Gallegos-Ruiz, A.; Sánchez- Gómez, W.S. Use of linear features by the common vampire bat (Desmodus rotundus) in a tropical cattle-ranching landscape. Therya 2019, 10, 229–234. [Google Scholar] [CrossRef]
- Thompson, R.D.; Mitchell, G.C.; Burns, R.J. Vampire bat control by systemic treatment of livestock with an anticoagulant. Science 1972, 177, 806–808. [Google Scholar] [CrossRef]
- Wohlgenant, T.J. Roost interactions between the common vampire bat (Desmodus rotundus) and two frugivorous bats (Phyllostomus discolor and Sturnira lilium) in Guanacaste, Costa Rica. Biotropica 1994, 26, 344–348. [Google Scholar] [CrossRef]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.S.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mamm. Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; de Torrez, E.B.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Fornes, A.; Lord, R.D.; Kuns, M.L.; Larghi, O.P.; Fuenzalida, E.; Lazara, L. Control of bovine rabies through vampire bat control. J. Wildl. Dis. 1974, 10, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Andrade, F.A.G.; França, É.S.; Souza, V.P.; Barreto, M.S.O.D.; Fernandes, M.E.B. Spatial and temporal analysis of attacks by common vampire bats (Desmodus rotundus ) on humans in the rural Brazilian Amazon basin. Acta Chiropterologica 2015, 17, 393–400. [Google Scholar] [CrossRef]
- Delpietro, H.; Konolsaisen, F.; Marchevsky, N.; Russo, G. Domestic cat predation on vampire bats (Desmodus rotundus) while foraging on goats, pigs, cows and human-beings. Appl. Anim. Behav. Sci. 1994, 39, 141–150. [Google Scholar] [CrossRef]
- Pérez-Rivero, J.J.; Pérez-Martínez, M.; Aguilar-Setién, A. Histometric analysis of vampire bat ( Desmodus rotundus ) testicles treated with coumestrol by oral route. J. Appl. Anim. Res. 2013, 42, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Ransom, J.I.; Powers, J.G.; Thompson Hobbs, N.; Baker, D.L. Ecological feedbacks can reduce population-level efficacy of wildlife fertility control. J. Appl. Ecol. 2014, 51, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, J.C.; Streicker, D.G.; Altizer, S.; Rohani, P. Resolving the roles of immunity, pathogenesis, and immigration for rabies persistence in vampire bats. Proc. Natl. Acad. Sci. USA 2013, 110, 20837–20842. [Google Scholar] [CrossRef] [Green Version]
- Rocha, F.; Dias, R.A. The common vampire bat Desmodus rotundus (Chiroptera: Phyllostomidae) and the transmission of the rabies virus to livestock: A contact network approach and recommendations for surveillance and control. Prev. Vet. Med. 2020, 174, 104809. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.R.; Streicker, D.G.; Schnell, M.J. The spread and evolution of rabies virus: conquering new frontiers. Nat. Rev. Microbiol. 2018, 16, 241. [Google Scholar] [CrossRef] [PubMed]
- Streicker, D.G.; Recuenco, S.; Valderrama, W.; Gomez Benavides, J.; Vargas, I.; Pacheco, V.; Condori Condori, R.E.; Montgomery, J.; Rupprecht, C.E.; Rohani, P.; et al. Ecological and anthropogenic drivers of rabies exposure in vampire bats: implications for transmission and control. Proc. R. Soc. B Biol. Sci. 2012, 279, 3384–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turmelle, A.S.; Jackson, F.R.; Green, D.; McCracken, G.F.; Rupprecht, C.E. Host immunity to repeated rabies virus infection in big brown bats. J. Gen. Virol. 2010, 91, 2360–2366. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Baer, G.M. Experimental rabies in the vampire bat. Am. Soc. Trop. Med. Hyg. 1980, 29, 254–259. [Google Scholar] [CrossRef]
- Cárdenas-Canales, E.M.; Gigante, C.M.; Greenberg, L.; Velasco-Villa, A.; Ellison, J.A.; Satheshkumar, P.S.; Medina-Magües, L.G.; Griesser, R.; Falendysz, E.; Amezcua, I.; et al. Clinical presentation and serologic response during a rabies epizootic in captive common vampire bats (Desmodus rotundus). Trop. Med. Infect. Dis. 2020, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Streicker, D.G.; Fallas González, S.L.; Luconi, G.; Barrientos, R.G.; Leon, B. Phylodynamics reveals extinction–recolonization dynamics underpin apparently endemic vampire bat rabies in Costa Rica. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191527. [Google Scholar] [CrossRef]
- de Thoisy, B.; Bourhy, H.; Delaval, M.; Pontier, D.; Dacheux, L.; Darcissac, E.; Donato, D.; Guidez, A.; Larrous, F.; Lavenir, R.; et al. Bioecological Drivers of Rabies Virus Circulation in a Neotropical Bat Community. PLoS Negl. Trop. Dis. 2016, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.J.; Broos, A.; Bergner, L.M.; Meza, D.K.; Simmons, N.B.; Fenton, M.B.; Altizer, S.; Streicker, D.G. Temporal patterns of vampire bat rabies and host connectivity in Belize. Transbound. Emerg. Dis. 2020, tbed.13754. [Google Scholar] [CrossRef]
- Delpietro, H.A.; Russo, R.G.; Carter, G.G.; Lord, R.D.; Delpietro, G.L. Reproductive seasonality, sex ratio and philopatry in Argentina’s common vampire bats. R. Soc. Open Sci. 2017, 4, 160959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trajano, E. Movements of cave bats in Southeastern Brazil, with emphasis on the population ecology of the common vampire bat, Desmodus rotundus (Chiroptera). Biotropica 1996, 28, 121–129. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; Cross, P.C.; Briggs, C.J.; Daugherty, M.; Getz, W.M.; Latto, J.; Sanchez, M.S.; Smith, A.B.; Swei, A. Should we expect population thresholds for wildlife disease? TREE Ecol. Evol. 2005, 20, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Choisy, M.; Rohani, P. Harvesting can increase severity of wildlife disease epidemics. Proc. R. Soc. B Biol. Sci. 2006, 273, 2025–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dognani, R.; Pierre, E.J.; Silva, M.; do, C.P.; Patrício, M.A.C.; Costa, S.C.; Prado, J.R.; Lisbôa, J.A.N. Epidemiologia descritiva da raiva dos herbívoros notificados no estado do Paraná entre 1977 e 2012. Pesqui. Veterinária Bras. 2016, 36, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Brochier, B.; Kieny, M.P.; Costy, F.; Coppens, P.; Bauduin, B.; Lecocq, J.; Languet, B.; Chappuis, G.; Desmettre, P.; Afiademanyo, K.; et al. Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Lett. to Nat. 1991, 353, 737–740. [Google Scholar] [CrossRef]
- Benavides, J.A.; Megid, J.; Campos, A.; Rocha, S.; Vigilato, M.A.N.; Hampson, K. An evaluation of Brazil’s surveillance and prophylaxis of canine rabies between 2008 and 2017. PLoS Negl. Trop. Dis. 2019, 13, e0007564. [Google Scholar] [CrossRef] [Green Version]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the Global Burden of Endemic Canine Rabies. PLoS Negl. Trop. Dis. 2015, 9, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Tschapka, M.; Wilkinson, G.S. Free-ranging vampire bats (Desmodus rotundus, Phyllostomidae) survive 15 years in the wild. Int. J. Mamm. Biol. 1999, 64, 239–240. [Google Scholar]
- Mulder, J.L. Longevity records in the red fox. Lutra 2004, 47, 51–52. [Google Scholar]
- Wilkinson, G.S.; Adams, D.M. Recurrent evolution of extreme longevity in bats. Biol. Lett. 2019, 15, 20180860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, G.S.; South, J.M. Life history, ecology and longevity in bats. Aging Cell 2002, 1, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Gold, S.; Donnelly, C.A.; Nouvellet, P.; Woodroffe, R. Rabies virus-neutralising antibodies in healthy, unvaccinated individuals: What do they mean for rabies epidemiology? PLoS Negl. Trop. Dis. 2020, 14, e0007933. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.F.; Martorelli, L.F.A.; Aires, C.C.; Sallum, P.C.; Massad, E. Indirect oral immunization of captive vampires, Desmodus rotundus. Virus Res. 2005, 111, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Setién, A.; Brochier, B.; Tordo, N.; De Paz, O.; Desmettre, P.; Peharpre, D.; Pastoret, P.P. Experimental rabies infection and oral vaccination in vampire bats (Desmodus rotundus). Vaccine 1998, 16, 1122–1126. [Google Scholar] [CrossRef]
- Stading, B.; Ellison, J.A.; Carson, W.C.; Satheshkumar, S.; Rocke, T.E.; Osorio, J.E. Protection of bats ( Eptesicus fuscus ) against rabies following topical or oronasal exposure to a recombinant raccoon poxvirus vaccine. PLoS Negl. Tropic. Dis. 2017, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Stading, B.R.; Osorio, J.E.; Velasco-Villa, A.; Smotherman, M.; Kingstad-Bakke, B.; Rocke, T.E. Infectivity of attenuated poxvirus vaccine vectors and immunogenicity of a raccoonpox vectored rabies vaccine in the Brazilian Free-tailed bat (Tadarida brasiliensis). Vaccine 2016, 34, 5352–5358. [Google Scholar] [CrossRef] [Green Version]
- Rocke, T.E.; Tripp, D.W.; Russell, R.E.; Abbott, R.C.; Richgels, K.L.D.; Matchett, M.R.; Biggins, D.E.; Griebel, R.; Schroeder, G.; Grassel, S.M.; et al. Sylvatic Plague Vaccine Partially Protects Prairie Dogs (Cynomys spp.) in Field Trials. Ecohealth 2017, 14, 438–450. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.N.; Uieda, W.; Do Rosário Dias De Oliveira Latorre, M. Influence of sex differences in the same colony for chemical control of vampire Desmodus rotundus (Phyllostomidae) populations in the state of Sao Paulo, Brazil. Pesqui. Vet. Bras. 2006, 26, 38–43. [Google Scholar] [CrossRef]
- Nuismer, S.L.; Althouse, B.M.; May, R.; Bull, J.J.; Stromberg, S.P.; Antia, R. Eradicating infectious disease using weakly transmissible vaccines. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161903. [Google Scholar] [CrossRef]
- Freitas, M.B.; Welker, A.F.; Millan, S.F.; Pinheiro, E.C. Metabolic responses induced by fasting in the common vampire bat Desmodus rotundus. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2003, 173, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.S. Reciprocal food sharing in the vampire bat. Nature 1984, 311, 276–279. [Google Scholar] [CrossRef]
- Voigt, C.C.; Kelm, D.H. Host Preference of the Common Vampire Bat (Desmodus Rotundus; Chiroptera) Assessed By Stable Isotopes. J. Mammal. 2006, 87, 1–6. [Google Scholar] [CrossRef]
- Bohmann, K.; Gopalakrishnan, S.; Nielsen, M.; Nielsen, L.D.S.; Jones, G.; Streicker, D.G.; Gilbert, M.T.P. Using DNA metabarcoding for simultaneous inference of common vampire bat diet and population structure. Mol. Ecol. Resour. 2018, 18, 1050–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya, M.I.; Pacheco, L.F.; Aguirre, L.F. Relationships between vampire bat (Desmodus rotundus) attacks to goats, livestock management, and some habitat characteristics in the Bolivian prepuna. Mastozool. Neotrop. 2015, 22, 73–84. [Google Scholar]
- Gilbert, A.T.; Petersen, B.W.; Recuenco, S.; Niezgoda, M.; Gómez, J.; Laguna-Torres, V.A.; Rupprecht, C. Evidence of rabies virus exposure among humans in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2012, 87, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.U.; Silva-Nunes, M. Da Evidence-based public health and prospects for malaria control in Brazil. J. Infect. Dev. Ctries. 2010, 4, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.C.; Aron, J.; Santos-Burgoa, C.; Uieda, W.; Ruiz-Velazco, S. Common vampire bat attacks on humans in a village of the Amazon region of Brazil. Cad. Saude Publica 2001, 17, 1531–1536. [Google Scholar] [CrossRef]
- McGuire, L.P.; Fenton, M.B. Hitting the Wall: Light Affects the Obstacle Avoidance Ability of Free-Flying Little Brown Bats (Myotis lucifugus). Acta Chiropterologica 2010, 12, 247–250. [Google Scholar] [CrossRef]
- Arnett, E.B.; Hein, C.D.; Schirmacher, M.R.; Huso, M.M.P.; Szewczak, J.M. Evaluating the Effectiveness of an Ultrasonic Acoustic Deterrent for Reducing Bat Fatalities at Wind Turbines. PLoS ONE 2013, 8, e65794. [Google Scholar] [CrossRef]
- Delpietro, H.A. Case reports on defensive behaviour in equine and bovine subjects in response to vocalization of the common vampire bat (Desmodus rotundus). Appl. Anim. Behav. Sci. 1989, 22, 377–380. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Hanlon, C.A.; Hemachudha, T. Rabies re-examined. Lancet Infect. Dis. 2002, 2, 327–343. [Google Scholar] [CrossRef]
- Benavides, J.A.; Velasco-Villa, A.; Godino, L.C.; Satheshkumar, P.S.; Nino, R.; Rojas-Paniagua, E.; Shiva, C.; Falcon, N.; Streicker, D.G. Abortive vampire bat rabies infections in Peruvian peridomestic livestock. PLoS Negl. Trop. Dis. 2020, 14, e0008194. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.B.; Martorelli, L.A.; Kataoka, A.P.G.A.; Campos, A.C.A.; Rodrigues, C.S.; Sanfilippo, L.F.; Cunha, E.S.; Durigon, E.L.; Favoretto, S.R. Antibodies ro rabies virus in terrestrial wild mammals in native rainforest on the North coast of São Paulo State, Brazil. J. Wildl. Dis. 2014, 50, 469–477. [Google Scholar] [CrossRef]
- World Health Organization. WHO Expert Consultation on Rabies. Third Report; WHO Technical Report Series, No. 1012; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Gómez-Benavides, J.; Laguna-Torres, V.A.; Recuenco, S. The real significance of being bitten by a hematophagous bat in indigenous communities in the remote Peruvian Amazon. Rev. Peru. Med. Exp. Salud Publica 2010, 27, 657–658. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.; Shwiff, S.; Gebhardt, K.; Ramírez, A.J.; Kohler, D.; Lecuona, L. Economic evaluation of vampire bat (Desmodus rotundus) rabies prevention in Mexico. Transbound. Emerg. Dis. 2014, 61, 140–146. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Rabies Vaccines and Immunoglobulins; WHO Position, Weekly Epidemiological Record; WHO: Geneva, Switzerland, 2018; Volume 16, pp. 201–220. [Google Scholar]
- Wang, C.; Dulal, P.; Zhou, X.; Xiang, Z.; Goharriz, H.; Banyard, A.; Green, N.; Brunner, L.; Ventura, R.; Collin, N.; et al. A simian-adenovirus-vectored rabies vaccine suitable for thermostabilisation and clinical development for low-cost single-dose pre-exposure prophylaxis. PLoS Negl. Trop. Dis. 2018, 12, e0006870. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benavides, J.A.; Valderrama, W.; Recuenco, S.; Uieda, W.; Suzán, G.; Avila-Flores, R.; Velasco-Villa, A.; Almeida, M.; Andrade, F.A.G.d.; Molina-Flores, B.; et al. Defining New Pathways to Manage the Ongoing Emergence of Bat Rabies in Latin America. Viruses 2020, 12, 1002. https://doi.org/10.3390/v12091002
Benavides JA, Valderrama W, Recuenco S, Uieda W, Suzán G, Avila-Flores R, Velasco-Villa A, Almeida M, Andrade FAGd, Molina-Flores B, et al. Defining New Pathways to Manage the Ongoing Emergence of Bat Rabies in Latin America. Viruses. 2020; 12(9):1002. https://doi.org/10.3390/v12091002
Chicago/Turabian StyleBenavides, Julio A., William Valderrama, Sergio Recuenco, Wilson Uieda, Gerardo Suzán, Rafael Avila-Flores, Andres Velasco-Villa, Marilene Almeida, Fernanda A.G. de Andrade, Baldomero Molina-Flores, and et al. 2020. "Defining New Pathways to Manage the Ongoing Emergence of Bat Rabies in Latin America" Viruses 12, no. 9: 1002. https://doi.org/10.3390/v12091002