An Overview of Phytophthora Species Inhabiting Declining Quercus suber Stands in Sardinia (Italy)


1
Settore Difesa Delle Piante Forestali, Agris Sardegna, Via Limbara 9, 07029 Tempio Pausania (SS), Italy
2
Dipartimento di Agraria, Sezione di Patologia Vegetale ed Entomologia, Università degli Studi di Sassari, Viale Italia 39, 07100 Sassari, Italy
3
Settore Sughericoltura e Selvicoltura, Agris Sardegna, Via Limbara 9, 07029 Tempio Pausania (SS), Italy
4
Nucleo Ricerca Desertificazione, Università degli Studi di Sassari, Viale Italia 39, 07100 Sassari, Italy
*
Author to whom correspondence should be addressed.
Forests 2020, 11(9), 971; https://doi.org/10.3390/f11090971
Submission received: 13 August 2020
/
Revised: 3 September 2020
/
Accepted: 4 September 2020
/
Published: 8 September 2020
(This article belongs to the Special Issue Phytophthora Infestations in Forest Ecosystems)
Abstract
:Cork oak forests are of immense importance in terms of economic, cultural, and ecological value in the Mediterranean regions. Since the beginning of the 20th century, these forests ecosystems have been threatened by several factors, including human intervention, climate change, wildfires, pathogens, and pests. Several studies have demonstrated the primary role of the oomycete Phytophthora cinnamomi Ronds in the widespread decline of cork oaks in Portugal, Spain, southern France, and Italy, although other congeneric species have also been occasionally associated. Between 2015 and 2019, independent surveys were undertaken to determine the diversity of Phytophthora species in declining cork oak stands in Sardinia (Italy). Rhizosphere soil samples were collected from 39 declining cork oak stands and baited in the laboratory with oak leaflets. In addition, the occurrence of Phytophthora was assayed using an in-situ baiting technique in rivers and streams located throughout ten of the surveyed oak stands. Isolates were identified by means of both morphological characters and sequence analysis of internal transcribed spacer (ITS) regions of ribosomal DNA. In total, 14 different Phytophthora species were detected. Phytophthora cinnamomi was the most frequently isolated species from rhizosphere soil, followed by P. quercina, P. pseudocryptogea, and P. tyrrhenica. In contrast, P. gonapodyides turned out to be the most dominant species in stream water, followed by P. bilorbang, P. pseudocryptogea, P. lacustris, and P. plurivora. Pathogenicity of the most common Phytophthora species detected was tested using both soil infestation and log inoculation methods. This study showed the high diversity of Phytophthora species inhabiting soil and watercourses, including several previously unrecorded species potentially involved in the decline of cork oak forests.
1. Introduction
Cork oak (Quercus suber L.) represents an important component of the Mediterranean forests landscape, covering more than 2 million ha across southern European and northern African countries [1]. This type of forest ecosystem has great socio-economic value, providing a range of non-timber forest products, such as cork, firewood, grazing, honey, and mushrooms, playing a key role in the rural economy in less favorable regions [2,3]. In particular, cork production represents a highly sustainable non-wood product derived from forests in the western Mediterranean countries and an additional source of income for farmers. In Sardinia (Italy), which hosts more than 80% of the Italian distribution area, cork oak is the second most important production chain of the island [4]. Moreover, these forest systems provide a wide range of several un-costed ecosystem services, including biodiversity conservation and desertification control [1,3,5]. For all of these reasons, many cork oak forests are recognized as protected ecosystems under the Pan-European network of protected areas (www.natura.org), Sites of Community Importance and Special Protection Areas for biodiversity conservation (Council Directive 92/43/EEC).
Despite performing these important functions, Mediterranean cork oak forests are currently under large scale reduction due to a wide range of drivers, such as pathogens and pests, climate change, wildfires, overgrazing, degradation, and fragmentation [6,7,8]. Over the last three decades, the role of pathogens in such forest ecosystems has gained increased attention due to the exponential emergence of forest diseases worldwide, particularly in Mediterranean ecosystems [9,10,11,12,13]. Several studies have demonstrated the involvement of the oomycete Phytophthora cinnamomi in the widespread decline of Mediterranean oaks, including cork oak, in Portugal, Spain, southern France, and Italy [14,15,16,17,18,19]. Although P. cinnamomi appears to be the most dominant species, other congeneric species can also be associated with Mediterranean oak decline [12,20,21]. The diversity of Phytophthora species in Mediterranean oak ecosystems has been further explored in recent years using metagenomic approaches based on high-throughput sequencing (HTS), which, through the use of species-specific primers, allow the amplification of a high number of target organisms from environmental DNA [22,23,24]. However, most of these studies are related to holm oak (Quercus ilex L.), while cork oak forests still remain poorly investigated, and to the best of our knowledge, only three Phytophthora species have been formally reported [12]. In the extensive surveys on reforestations and afforestations across Europe between 1998 and 2009 by Jung and collaborators [25], P. cryptogea (now known as P. pseudocryptogea) and P. quercina were reported from only two cork oak plantations in Spain. More recently, three previously unrecorded Phytophthora species have been associated with episodic events of cork oak decline in Italy, including the newly described P. tyrrhenica and the exotic pathogenic P. megasperma and P. multivora [21,26].
The main objective of the present work was to investigate the diversity of Phytophthora species in declining cork oak forest ecosystems in Sardinia. As many Phytophthora species have a specific aquatic lifestyle, their occurrence was also explored along rivers and streams within the surveyed sites. Moreover, the pathogenicity of the most frequently isolated Phytophthora species detected was tested using both soil infestation and log inoculation methods.
2. Materials and Methods
2.1. Soil Sampling and Phytophthora Isolation
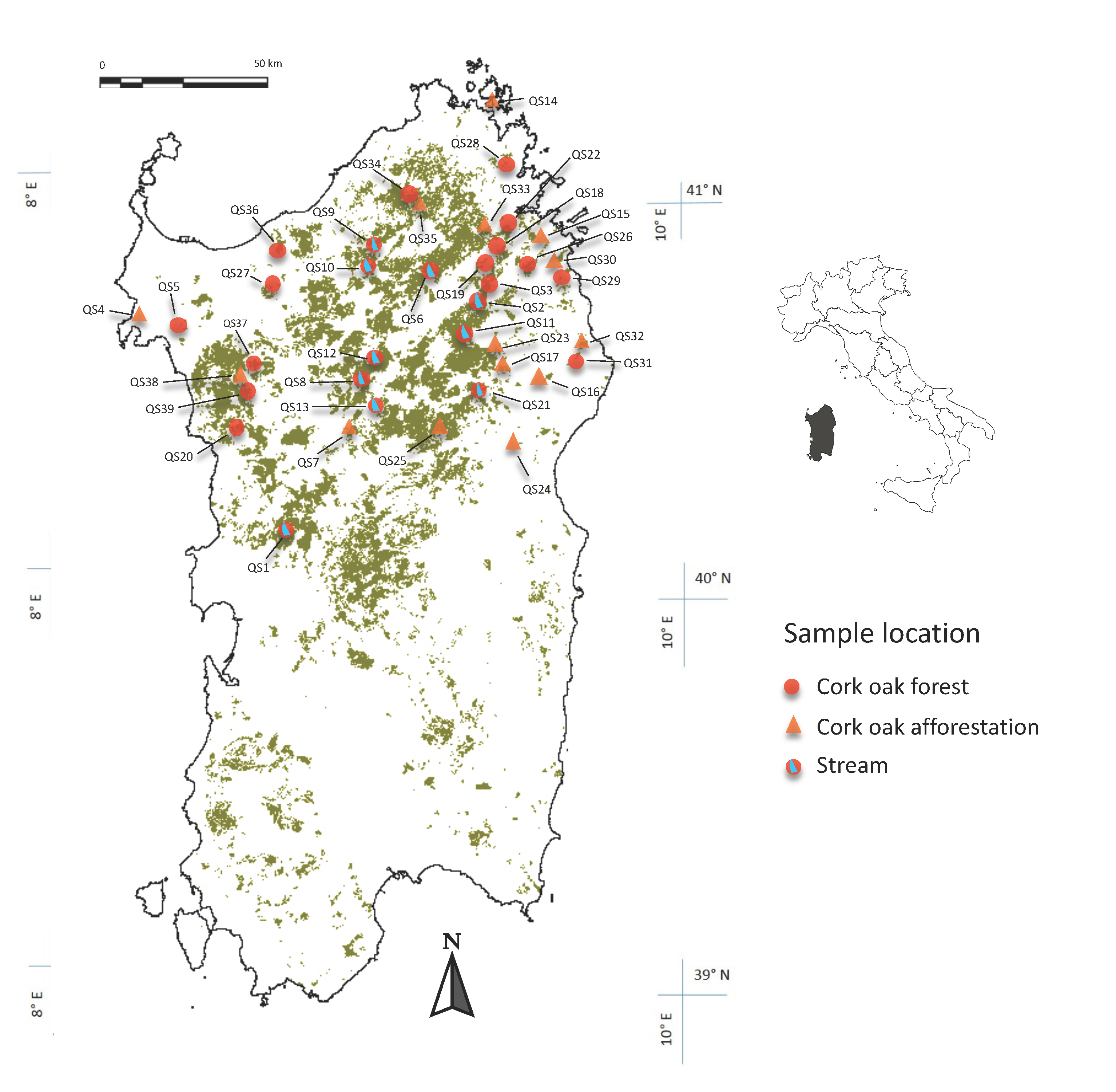
Between 2015 and 2019, independent surveys were undertaken to investigate the diversity of Phytophthora species from 39 declining cork oak forests in Sardinia: 25 natural forests and 14 afforestation sites (Figure 1). These sites were selected over the years on the basis of different reports, submitted by private and public entities, of problems affecting the health of cork oak trees. Tree health monitoring and surveillance work were made by an initial pre-screening in the field based on visible symptoms of decline, such as yellowing leaves, crown transparency, epicormic shoots, branch dieback, bleeding cankers, as well as necrotic lesions at the collar and root levels. Samplings were conducted in the autumn and spring seasons, and in some areas, these were repeated twice. A total of 295 symptomatic cork oak trees were sampled. Rhizosphere soil samples consisted of a mixture of four subsamples taken from around the stem base of selected trees, scraping away the litter and taking about 200–300 g of roots and soil. Phytophthora isolations were made using an adaptation of the baiting methods described by Jung et al. [27]. In the laboratory, roots and soil were flooded in 12 × 10 × 22 cm glass trays with 500 mL of distilled water, then young leaflets taken from 1–2-month-old cork oak seedlings were used as baits floated over the water. After 3–5 days, leaves with black spots were checked under the microscope for the presence of sporangia, dried on filter paper, and plated onto synthetic mucor agar (SMA), a selective medium for Phytophthora [28]. All Petri dishes were incubated in the dark at 20 °C and checked daily for Phytophthora-like hyphae development, which was subsequently transferred to Petri dishes containing carrot-agar (CA; 16 g agar technical no.3, Oxoid Ltd., Basingstoke, UK, 200 g carrots and 1000 mL distilled water) [29].
2.2. Stream Baiting and Phytophthora Isolation
In addition to soil samples, river and stream water within ten surveyed cork oak forests (Figure 1) were assayed using the in-situ baiting technique in spring 2018. Watercourses were chosen depending on their water flow and on the capacity to collect water from the bordering forests. They were subdivided into two different groups, mainly based on the altimetric gradient, including valley floor rivers with permanent water flow, and mountain or hill streams or water catchments into the forests, with water flow strictly correlated with the seasonal rainfall and very often drying up in summer. River baiting was made using an adaptation of the method described by Reeser et al. [30] and Hüberli et al. [31], which consisted of two squared layers of fly mesh or metallic net, sealed together, with young leaves of different plant species, such as Alnus glutinosa Mill., Arbutus unedo L., Buxus sempervirens L., Laurus nobilis L., Fraxinus ornus L., Hedera helix L., Parthenocissus quinquefolia L., Q. ilex L., Q. suber L., and Taxus baccata L. placed between the two mesh layers. Rafts were placed in-situ at the same time as their set up, and each raft contained about 20 leaves. Cork stoppers were used to float the raft over the water surface. Three rafts were placed every 200 m from each other along the watercourse. The rafts were fastened to natural restraints, such as branches and rocks, and left floating over the water for 3–4 days and then brought to the laboratory where the leaves were washed with sterile water and blotted dry on filter paper. Phytophthora isolations were made by placing small fragments cut from necrotic lesions detected on the leaf baits on SMA. Any developing colonies were sub-cultured on CA for further analyses.
2.3. Morphological and Molecular Identification
The isolates obtained from soil samples and stream water were first grouped based on their colony growth patterns after 5–7 days at 20 °C in the dark on CA. In addition, morphological features of sporangia, oogonia, antheridia, chlamydospores, hyphal swellings, and aggregations were examined under the Leitz Diaplan compound microscope (Leitz, Wetzlar, Germany) and compared with species descriptions in the literature [21,32,33,34,35,36,37,38]. A subset of representative morphotypes (88 isolates) was selected for molecular analyses, which consisted of DNA extraction, amplification, sequencing, and analysis of sequences of the entire region of the internal transcribed spacers (ITS1 and ITS2) and the 5.8 S gene of the rDNA. DNA was extracted from mycelium fragments, using the extraction kit InstaGene™ Matrix (BioRad Laboratories, Hercules, CA, USA). The amplification of the ITS region was carried out with a thermocycler (Hybaid PCR Express), using the forward primers ITS1 or ITS6 and the reverse primer ITS4 [39,40]. A total volume of 50 μL, consisting of 18.2 μL of water, 5 μL of BSA, 5 μL of dNTPs, 5 μL of both ITS6 and ITS4 primers, 10 μL of the buffer, 0.3 μL of Go Taq polymerase, and 1.5 μL of DNA from each morphotype, was used for standard PCR (polymerase chain reaction). The cycle used for the amplification of the ITS genes regions was as follows: initial denaturation of 1 min at 94 °C, followed by 35 cycles of 1 min at 94 °C, 1 min at 56 °C, 1 min at 72 °C, followed by a final elongation phase of 7 min at 72 °C. The PCR products were purified using the EUROGOLD gel extraction kit (EuroClone S.p.A., Pero, Italy). After quantification, purified PCR amplicons and the sequencing primers were sent to BMR Genomics sequencing service (https://www.bmr-genomics.it). DNA sequence chromatograms were viewed and edited using BioEdit v. 5.0.6 software [41]. Heterozygous sites observed were labeled according to the IUPAC coding system. Isolates were assigned to a species when sequence identities were above a 99% cut-off with respect to those of ex-type isolates or key isolates. All sequences were deposited at GenBank (http://www.ncbi.nlm.nih.gov/), and the accession numbers are given in Table 1.
2.4. Pathogenicity Tests
Pathogenicity of the most frequent Phytophthora species isolated was assayed using the soil infestation method described by Jung et al. [27], with some modification as reported by Scanu et al. [29]. In particular, two isolates of P. cinnamomi, P. gonapodyides, P. pseudocryptogea, P. psychrophila, P. quercina, and P. tyrrhenica were grown in individual 500 mL Erlenmeyer flasks containing an autoclaved mixture of 250 mL of vermiculite and 150 mL of Lolium italicum seeds thoroughly moistened with 100 mL of carrot juice (200 mL/L carrot juice, 3 g/L CaCO3, and 800 mL/L distilled water). Flasks were incubated at 20 °C for 1 month, then 20 mL of inoculum was collected and inserted inside the soil of 2-year-old cork oak seedlings (provided by the Regional Agency Fo.Re.S.T.A.S.). The substrate in the controls received a sterile mixture of vermiculite/seeds-carrot juice at the same ratio. To stimulate the production of sporangia and pathogen spread and infection via zoospores, pots were flooded immediately after inoculation for 48 h, and flooding was repeated at three-week intervals by immerging pots in 10 L buckets just to 1 cm above the soil surface. There were eight replicates per isolate and controls. After 5 months of incubation at 20 °C (±2 °C), 70% relative humidity with a 12/12 h photoperiod, seedlings were visually assessed for symptoms, and the mortality rate was recorded; then each plant was removed from the pot, and the root system gently washed under tap water. Single roots were cut off at the collar, and after scanning, the total root length of all the plant root systems was measured using the APS Assess 2.0 software (The American Phytopathological Society, St. Paul, MN, USA). The remaining soil was baited following the method described above to determine whether the pathogen was still viable. Re-isolations were also made directly from necrotic roots using SMA.
Pathogenicity of the above isolates (except for those of P. tyrrhenica) was further tested using freshly cut logs of cork oak following the method described by Brasier and Kirk [42]. Four logs (1.4 m long and 20 cm in diameter) were cut from stems of living cork oak trees 24 h before the experiment, and the cut ends were sealed with a liquid waterproofing membrane. In each log, three bands were marked around the log circumference, 30 cm apart from each other, with 5 inoculation points per band, about 15 cm apart. After sterilizing the bark with 70% ethanol, a 7 mm diameter hole was punched through the bark to the wood surface with a steel cork borer. The same-sized plug was taken from the edge of an actively growing colony on CA and used as inoculum by inserting into the hole replacing the bark plug. Three control inoculation points per log were inoculated with a sterile CA plug and covered with the removed piece of bark. Moist cotton wool was placed over the wounds, covered with a 5 × 5 cm piece of aluminum foil, and sealed with an adhesive PVC tape. There were four replicates per isolate. Inoculated logs were covered individually in loose polythene sleeves (sealed at both ends) and incubated at 20 °C (±2 °C) in an air-conditioned laboratory and checked weekly for the appearance of symptoms. After 45 days, the experiment was finished, and logs were destructively sampled by removing the periderm with a drawknife to expose the phloem. Each lesion’s outline was then recorded on tracing paper and scanned on an Epson Perfection V30 photo scanner, and the lesion area calculated using APS Assess software, as described by Scanu and Webber [43]. Re-isolation of all the inoculated Phytophthora species onto SMA was attempted from the lesion margins.
Statistical analyses for both pathogenicity tests were performed using XLSTAT software (Addinsoft). Data were first checked for normality and then subjected to analysis of variance (ANOVA). Statistical differences among mean values of root lengths and lesion areas were determined using Fisher’s protected least significant difference (LSD) test. Differences with p < 0.05 were considered significant.
3. Results
3.1. Symptomatology
A wide range of symptoms of decline was observed on cork oak trees across all the investigated sites. These included rapid dieback of the crown in both mature (Figure 2a) and young oak trees (Figure 2b), which was frequently observed in early autumn, especially after a long summer and drought conditions. In the case of afforestation sites (10 to 20-year-old), the infections could reach epidemic levels and cause extensive mortality of oak trees. Other symptoms included shoot dieback and increased transparency of the whole crown, leaf chlorosis, and abundant proliferation of epicormic shoots on stems and branches (Figure 2c,d). At the collar level, trees showed necrotic bark lesions frequently associated with black exudation and very often girdling the stem (Figure 2e). In the root system of declining oak trees, an extensive loss of both lateral small woody roots and fine roots and callusing or open cankers on suberized roots were observed.
3.2. Soilborne Phytophthora Species
Phytophthora species were recovered from 68.5% of the 295 soil samples tested. The highest level of soil infestation was detected in the afforestation sites (80.4%), while in natural forests, the percentage of positive trees was 61.7%. In total, 224 isolates were obtained from rhizosphere soil samples collected from around symptomatic trees in declining cork oak stands (Table 1). All isolates conformed morphologically to previously known Phytophthora species. ITS sequence analysis of the isolates confirmed the morphological identification of all Phytophthora species. BLAST searches in GenBank showed 99–100% similarity with reference sequences, including those of ex-type cultures or representative isolates (Table 1). In total, eight Phytophthora species belonging to five (clade 3, 6, 7, 8, and 12) out of the twelve known phylogenetic clades were isolated, including P. cinnamomi, P. gonapodyides, P. pseudocryptogea, P. psychrophila, P. quercina, P. syringae, P. tyrrhenica, and P. ×cambivora (Table 1 and Figure 3a,c). Phytophthora cinnamomi from clade 7c was the most frequent species isolated from both natural forests and afforestation stands. It was detected from almost all investigated afforestation sites (from 30 out of 39 sites) with an infection rate of 80.2%, while its incidence was markedly lower in natural cork oak forests (45.7% of 122 investigated trees). At one afforestation site (QS35), P. cinnamomi was recovered from 27 out of 30 cork oak trees sampled (Table 1). It was the only species recovered in 10 investigated stands. Similarly, the second most common species, P. quercina, from clade 12 occurred in both afforestation and natural stands, with an infestation rate ranging from 8.1% to 30.2%, respectively. It was the only species isolated from declining trees in sites QS6, QS13, and QS25. All P. quercina isolates had identical ITS sequences; however, a certain phenotypic variation among the isolates was observed. Both P. pseudocryptogea (clade 8a) and P. tyrrhenica (clade 7a) were recovered from eight declining cork oak stands, with an infestation rate of around 10%. The ITS sequences of all P. pseudocryptogea isolates from rhizosphere soil matched the ex-holotype isolate (GenBank no. KP288376). However, they had a unique polymorphism at position 56 (C instead of Y) and were heterozygous at position 650 (Y instead of T). Phytophthora gonapodyides from clade 6b occurred only in natural contexts, isolated from 12 declining trees in seven sites, and always in association with other Phytophthora species. Among the less frequently isolated species, P. psychrophila (clade 3) and P. ×cambivora (clade 7a) were detected only from natural forests at a very low infestation rate (2.5% and 1%, respectively), while P. syringae was exclusively found at one afforestation site from two symptomatic trees. In 18 soil samples from declining trees and 12 sites, multiple Phytophthora species were detected. Phytophthora cinnamomi was isolated along with P. quercina in four samples and along with both P. quercina and P. tyrrhenica (two samples) or P. pseudocryptogea (one sample). In four cases, P. tyrrhenica was isolated together with P. cinnamomi and, in one case, with P. quercina.
3.3. Waterborne Phytophthora Species
In total, 115 Phytophthora isolates were detected in all watercourses monitored through ten selected declining cork oak stands. Based on morphological analyses and molecular identification, these isolates belonged to five phylogenetic clades (clade 2, 6, 7, 8, and 9) corresponding to nine formally known Phytophthora species, including Phytophthora bilorbang, Phytophthora chlamydospora, P. gonapodyides, P. hydropathica, P. lacustris, P. plurivora, P. pseudocryptogea, P. ×cambivora, and P. ×multiformis (Table 2 and Figure 3b,c). Overall, more than 50% of the isolates obtained were identified as P. gonapodyides, which was recovered from all watercourses surveyed. Interestingly, eight isolates detected from stands QS1, QS8, and QS9 were heterozygous at position 106 (R instead of G). Another isolate from stand QS1 (GON3) shared the heterozygous position 106 and had unique polymorphisms at positions 106 (A instead of R or G) and 517 (T instead of G), respectively. Two isolates, PH255 (QS10) and PH267 (QS9), also differed from the other P. gonapodyides isolates by having unique polymorphisms at positions 145 (T instead of C) and 517 (T instead of G), respectively.
Phytophthora bilorbang was the second most widespread species isolated from five investigated streams, four of which only flow seasonally. All isolates from stand QS21 were heterozygous at position 106 (Y instead of T). Phytophthora pseudocryptogea and P. plurivora were isolated only from permanent water bodies, while P. lacustris occurred from both permanent and intermittent watercourses (Table 2). The ITS sequences of all P. lacustris isolates differed from the ex-type culture (GenBank no. AF266793) having a heterozygous site at position 783 (S instead of C). In addition, one isolate was heterozygous at position 458 (Y instead of C). Almost all isolates identified as P. pseudocryptogea from river water had identical ITS sequences than those isolates obtained from rhizosphere soil, differing from the ex-holotype by 2 bp. Moreover, one isolate (PH269) from stand Q12 differed from the ex-holotype isolate of P. pseudocryptogea and the other isolates obtained in this study by 4–5 bp at positions 56, 601, 650, 728, and 733. Phytophthora chlamydospora and P. ×cambivora were recovered from two streams, while P. ×multiformis and P. hydropathica were exclusively isolated from QS10 and QS13, respectively. The two P. chlamydospora isolates from stand QS9 were heterozygous at position 666 (Y instead of T). All isolates identified as P. hydropathica differed from the ex-type culture (GenBank no. EU583793) by having two heterozygous sites at positions 413 (S instead of C) and 665 (K instead of T) and by a unique polymorphism at position 628 (G instead of C). The ITS sequences of P. ×cambivora often generated overlapping ITS sequences starting at position 396 in both directions. This was a consistent pattern observed in all isolates of P. ×cambivora obtained from river water. The non-overlapping sequences up the indel positions were identical to that of the neotype culture of P. ×cambivora (GenBank no. KU899179).
Looking at the diversity of Phytophthora species across rivers, the two geographically close rivers at sites QS9 and QS10 hosted the highest number of Phytophthora species, followed by a stream in stand QS12 with five species detected. Only two species were isolated from water bodies at stands QS1–3, QS13, and QS21, and they were all from clade 6, except for a clade 9 species at site QS13 detected together with P. gonapodyides.
3.4. Pathogenicity Test
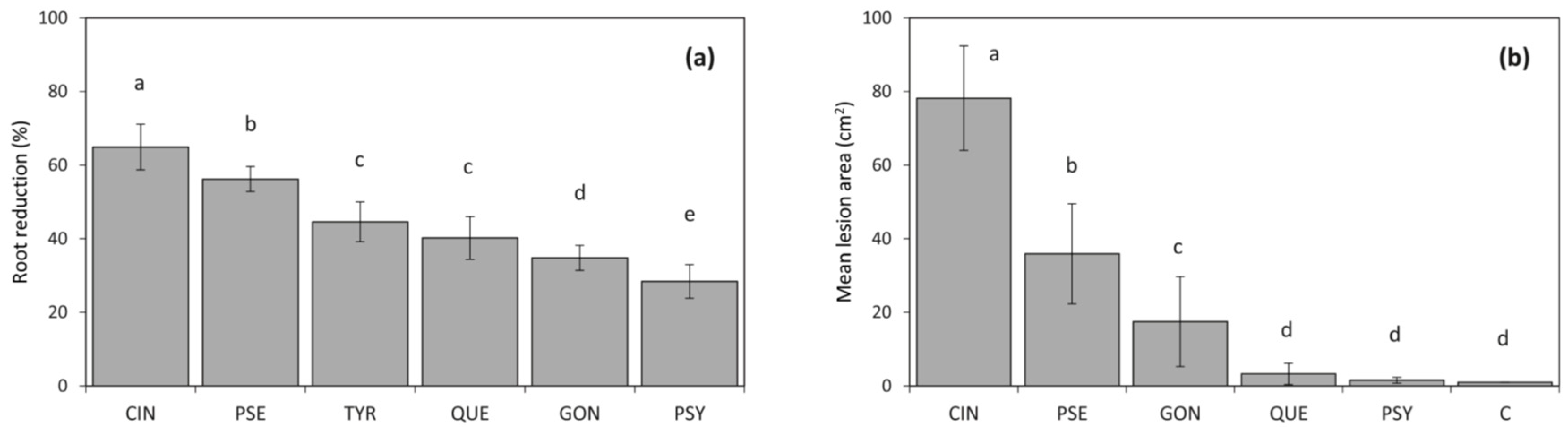
The soil infestation experiment showed that all Phytophthora species tested were able to cause a significant reduction of the root system in 2-year-old cork oak seedlings (Figure 4a). The mean root length was significantly higher in control seedlings (p < 0.05) than in seedlings infected with Phytophthora isolates. Phytophthora cinnamomi was the most aggressive species causing a root length reduction near to 65% compared to the control seedlings, followed by P. pseudocryptogea (56.2%), P. tyrrhenica (44.6%), and P. quercina (40.2%). Phytophthora gonapodyides and P. psychrophila caused a root length reduction below 35%. Phytophthora cinnamomi was the only species associated with extensive lesions on the mother root, with lesions in some cases reaching 15 mm in length. Apart from P. psychrophila, all the other Phytophthora species were re-isolated from limited necrotic lesions on taproot. No symptoms of pathogen infection could be seen on the roots of control seedlings.
In the log inoculation tests, lesions in the phloem tissue of cork oak caused by P. cinnamomi were significantly larger (p < 0.0001) compared with the negative controls, with a mean necrosis area of 80 cm2 (Figure 4b). Phytophthora pseudocryptogea and P. gonapodyides also showed considerable aggressiveness on inoculated logs. The mean lesion area formed by P. cinnamomi was approximately two to three times larger (p < 0.0001) than that developed following inoculation with P. pseudocryptogea and P. gonapodyides, respectively (Figure 4b). Phytophthora psychrophila and P. quercina were not able to colonize phloem tissues producing lesions that did not differ significantly from the negative controls (p > 0.05). Apart from P. psychrophila and P. quercina, all Phytophthora species were readily re-isolated from the necrotic lesions. In contrast, the controls developed only limited discoloration around the inoculation point and never yielded any Phytophthora.
4. Discussion
The extensive surveys made over four years across declining cork oak stands in Sardinia, together with morphological and ITS sequences analyses, have revealed the occurrence of 14 Phytophthora taxa from seven of the 12 known phylogenetic clades [21]. These include species common in Mediterranean oak soil, such as P. cinnamomi, P. gonapodyides, and P. quercina, and the less widespread species P. pseudocryptogea, P. psychrophila, P. syringae, P. tyrrhenica, and P. ×cambivora [12,17,20,44,45]. The detection of nine Phytophthora species from stream and river water represents the first attempt to look at the diversity of aquatic species in such forest ecosystems. Apart from P. gonapodyides and P. pseudocryptogea, all the other species identified from watercourses, P. bilorbang, P. chlamydospora, P. hydropathica, P. lacustris, P. plurivora, P. ×cambivora, and P. ×multiformis, were never reported in cork oak ecosystems.
Most of the previous surveys on Mediterranean oak decline have focused on the association of P. cinnamomi and Q. ilex (ssp. ballota and rotundifolia) [16,17,46,47,48,49], while cork oak has been less studied as it appears to be more tolerant to the disease due to its defense response mechanisms to Phytophthora infection [14,15,50]. Our study represents the first extensive survey on the distribution of P. cinnamomi and other congeneric species in declining cork oak stands.
Among the Phytophthora species detected from soil samples, P. cinnamomi was the most common species encountered. Listed as one of the 100 worst invasive alien species, P. cinnamomi is considered one of the most devastating plant pathogens worldwide [51,52]. It was first associated with the severe dieback and mortality of Mediterranean oaks, including both cork and holm oak, in the Iberian peninsula by Brasier in 1992 [53]; and since then, P. cinnamomi has been reported across the Mediterranean basin, and this is well documented for European countries [14,15,16,17,44,54]. In this study, the pathogen was detected only from rhizosphere soil; however, it was occasionally isolated from bleeding lesions on the stem (data not shown), as reported by Robin et al. [15]. Although cork oak has been shown to be less susceptible than holm oak [46,55], the high ability of P. cinnamomi to colonize phloem tissues, as exhibited in the log inoculation trial in this study and previously [56], together with its widespread occurrence across Sardinian stands, could suggest less tolerance of the Sardinian cork oak population [57]. This hypothesis, however, needs further investigation.
Phytophthora quercina is the second most prevalent species from soil samples. This oak-specific pathogen has been previously reported in central and southern Europe, causing a chronic decline in Quercus cerris, Quercus faginea, Q. ilex, Quercus petrea, Quercus pubescens, and Quercus robur [20,44,58,59]. Although P. quercina was recorded from two cork oak plantations in Spain [25], this appears to be the first widespread occurrence in natural cork oak forests. Interestingly, two distinct phenotypes were observed amongst the isolates detected from cork oak trees, supporting the hypothesis that P. quercina originated from Europe [21]. Multigene sequencing and phylogenetic analyses are currently underway to investigate the genetic population structure of a large number of P. quercina isolates from different oaks and various geographic provenances (Scanu and Jung, unpublished).
Two other slow-growing and homothallic species are detected at low frequency, and these are identified as P. psychorphila and P. syringae. Together with P. quercina, both species were previously reported from Mediterranean oaks in Spain, and their pathogenicity was demonstrated on both Q. ilex and Q. faginea [20]. Pathogenicity tests in the present study showed both species were not able to invade the inner bark of cut logs of cork oak; however, this did not correlate with root susceptibility. Previous results obtained by Perez-Sierra et al. [20] suggest these species are well adapted to the Mediterranean climate and may act as fine root nibblers, the incidence of which varies depending on the occurrence of extreme climatic events, such as recurrent drought and wet seasons [60]. Both P. psychrophila and P. quercina are already reported in Sardinia from declining Q. ilex trees, while P. syringae is associated with dieback and mortality of Juniperus phoenicea on Caprera Island [28]. Due to their low maximum temperature for growth (around 25 °C), typical of Phytophthora species from cool temperate regions, a potential seasonal activity, as suggested for P. cinnamomi and other cool-temperature pathogens, may occur [61,62,63,64]. Of note, P. syringae was detected only from a new plantation, suggesting its possible new introduction through infected plant material [25,65]. This could also happen for the other Phytophthora species considering the massive afforestation effort in Sardinia between 1990 and 2010 through EU programs, like EEC Regulation 2080/92 [4,25].
The finding of the recently described P. tyrrhenica confirmed its original description from declining cork oak trees [21]. Apart from one site (QS39), it was detected at nine stands and always coexisted with other congeneric species. So far, this cryptic species has been recovered from cork and holm oak trees in Sardinia and Sicily (Italy), respectively, and it is considered endemic to the Mediterranean basin [21,66]. Similarly, P. pseudocryptogea from clade 8 was the only species obtained from QS5 and QS38; otherwise, it co-occurred with P. cinnamomi or other species. It is noteworthy that all the previous isolates identified as Phytophthora cryptogea in Sardinia from forest trees, including oaks and Pinus radiate, are indeed P. pseudocryptogea [28,37,48,67]. Phytophthora pseudocryptogea was recently reported as one of the most widespread species in riparian thermo-Mediterranean forest stands and from five rivers in Sicily [66]. The clade 6 species P. gonapodyides had a more scattered distribution (seven stands and 12 trees), and its finding on cork oak confirmed results obtained by Jung et al. [66]. Finally, for the first time, we recorded the hybrid species P. ×cambivora on Q. suber. In Mediterranean regions, this species is frequently associated with “chestnut ink disease” [68], as well as from other Fagaceae [66,69] and Pinaceae trees [70]. In a recent survey in Sicily, P. ×cambivora was isolated from three different Mediterranean oaks, namely Q. cerris, Q. ilex, and Q. pubescens [66].
The detection of nine Phytophthora species from five phylogenetic clades in 10 rivers within declining cork oak stands was unexpected since similar diversity rates are often reported in more diverse forest ecosystems and with higher sampling rates from surveys in Australia, Europe, the USA, and South Africa [31,32,66]. The Phytophthora assemblage from watercourses was different from that detected from soil samples at the same sites, which was consistent with previous surveys [38,66,71,72]. Only three species were shared between terrestrial and aquatic environments, and these were P. pseudocryptogea, P. gonapodyides, and P. ×cambivora. Interestingly, the most common Phytophthora species isolated from rhizosphere soil, P. cinnamomi, was never detected from streams running through declining cork oak stands. As reported by previous similar studies, clade 6 species were the most common inhabitants of streams and rivers, highlighting their specific lifestyle to aquatic environments [73,74]. Phytophthora gonapodyides occurred in all investigated sites, followed by P. bilorbang, P. lacustris, and P. chlamydospora isolated from five, four, and two watercourses, respectively. Both P. bilorbang and P. gonapodyides have been previously reported in Sardinia on Caprera Island, detected from both rhizosphere soil beneath declining Mediterranean maquis vegetation and holm oak trees [28]. In Italy, P. bilorbang has also been reported from riparian ecosystems of Alnus glutinosa, in Sardinia, and on 15-year-old olive trees in Calabria [75,76], while P. gonapodyides is generally encountered in Mediterranean forest ecosystems, including holm oak [20,44,45]. In the pathogenicity tests, P. gonapodyides caused significant lesions in cork oak logs, confirming its ability to colonize bark and xylem tissues in both artificial and natural infections [77], behaving as a weak pathogen able to survive as a saprophyte on twigs and leaves playing a role in the breakdown of trees debris [78,79]. The other two clade 6 species, P. lacustris and P. chlamydospora, are common species previously reported in Italy from water bodies in forest ecosystems [36,66,80]. Phytophthora plurivora and P. pseudocryptogea are cosmopolitan pathogens with a broad host range [34,37,81]. The DNA of both species was recently detected from holm oak stands across different regions in Spain [23,24], and the pathogenicity tests showed they are amongst the most aggressive species on inoculated holm oak seedlings [82]. The detection of the clade 9 P. hydropathica is not surprising since it was recently detected from river water in Sicily and again using metabarcoding in Spain [24,66]. The origin of P. hydropathica is unknown but considering its low frequency (only one river) and occurrence from ornamental plants in a nursery in Italy [83], a recent introduction into wild environments is most likely.
Of note is the detection of P. ×cambivora and P. ×multiformis, two stable hybrid species from clade 7a that have evolved elsewhere. While P. ×cambivora has been previously reported from Mediterranean oaks in Italy [44], and Q. suber soil in the present study, the isolation of P. ×multiformis most likely occurred due to the presence of Alnus glutinosa trees along the river where the hybrid was isolated. The occurrence of multiple heterozygous sites in the ITS sequences of some isolates, including P. chlamydospora, P. gonapodyides, P. hydropathica, and P. lacustris, together with mixed unreadable ITS sequences generated for some of these isolates could indicate their possible hybrid nature. However, since the ITS region is not a particularly useful locus for studying interspecific hybrids due to the presence of the highly variable non-coding regions ITS1 and ITS2 [83], further molecular analyses, such as cloning, sequencing of other nuclear and mitochondrial genes, estimation of nuclear DNA content by flow cytometry, as well as morphological characterization of the isolates are required [38,84,85].
5. Conclusions
This unexpected high diversity of Phytophthora species in cork oak stands in such a small geographic area underlines how limited our current knowledge of oomycete diversity in Mediterranean oak forests is [86]. Recent molecular studies in Spanish oak forests have revealed the presence of several Phytophthora species besides P. cinnamomi [12,22,24,87], most of which were detected also in this survey. Although the high-throughput amplicon pyrosequencing of environmental DNA represents a very useful tool for assessing Phytophthora diversity in environmental samples, very little is known on the biological status of the detected microorganisms; therefore, specific baiting technique and metagenomic approaches should be carried out in parallel [72,88].
Pathogenicity tests and results obtained on the susceptibility of Mediterranean oaks to Phytophthora taxa from this and other prior studies [15,46,55,56,62] suggest these previously unrecorded species may play a relevant role in the aetiology of cork oak decline either acting as fine root “nibblers” [20,33,58] or shaping recruitment patterns due to their negative effects on seedling establishment [89,90]. Future studies will be based around understanding the ecological role of all Phytophthora species recovered in such ecosystems as well as their possible interactions with hosts and a changing environment that could promote the establishment of invasive Phytophthora in cork oak forests.
Author Contributions
Conceptualization: B.S., S.S.; data curation: B.S., S.S., A.B.; formal analysis: B.S., S.S.; investigation: B.S., S.S., A.B.; methodology: B.S., S.S., A.B.; project administration: B.S., S.S., C.S., P.A.R.; resources: B.S., C.S., P.A.R.; Supervision: B.S.; writing—original draft: S.S., B.S. writing—review and editing: B.S., S.S., A.B., C.S., P.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was in part financially supported by the Sardinian Regional Government (Convenzione fra la Regione Autonoma Della Sardegna, Assessorato della Difesa dell’Ambiente e l’AGRIS Dipartimento della Ricerca per il Sughero e la Silvicoltura per “L’attuazione del programma biennale di lotta ai lepidotteri defogliatori della sughera”). This study was also supported by the “fondo di Ateneo per la ricerca 2019”, an internal funding provided by the University of Sassari.
Acknowledgments
The Regional Agency Fo.Re.S.T.A.S. is gratefully acknowledged for providing cork oak seedlings used in the pathogenicity tests. The authors would like to thank Carolyn Riddel for the English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aronson, J.; Pereira, J.S.; Pausas, J.G. Cork Oak Woodlands on the Edge: Ecology, Adaptive Management, and Restoration; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Merlo, M.; Croitoru, L. Valuing Mediterranean Forests: Towards Total Economic Value; CABI Publishing: Cambridge, MA, USA, 2005; p. 406. [Google Scholar]
- Bugalho, N.M.; Caldeira, C.M.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 5, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Sedda, L.; Delogu, G.; Dettori, S. Forty-four years of land use changes in a Sardinian cork oak agro-silvopastoral system: A qualitative analysis. Open For. Sci. J. 2011, 4, 57–66. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Aparicio, L.; Ibanez, B.; Serrano, M.S.; De Vita, P.; Avila, J.M.; Perez-Ramos, I.M.; Garcia, L.V.; Sanchez, M.E.; Maranon, T. Spatial patterns of soil pathogens in declining Mediterranean forests: Implications for tree species regeneration. New Phytol. 2012, 194, 1014–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaz, C.; Alegria, C.; Abrantes Massano Monteiro, J.A.; Canavarro Teixeira, M.C. Land cover change and afforestation of marginal and abandoned agricultural land: A 10-year analysis in a Mediterranean region. For. Ecol. Manag. 2013, 308, 40–49. [Google Scholar] [CrossRef]
- Kim, M.; Lee, W.K.; Choi, G.M.; Song, C.; Lim, C.H.; Moon, J.; Piao, D.; Kraxner, F.; Shividenko, A.; Forsell, N. Modeling stand-level mortality based on maximum stem number and seasonal temperature. For. Ecol. Manag. 2017, 386, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Garbelotto, M.; Pautasso, M. Impacts of exotic forest pathogens on Mediterranean ecosystems: Four case studies. Eur. J. Plant Pathol. 2012, 133, 101–116. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Gullino, M.L. Emerging and re-emerging fungus and oomycete soil-borne plant diseases in Italy. Phytopathol. Mediterr. 2019, 58, 451–472. [Google Scholar]
- Moricca, S.; Linaldeddu, B.T.; Ginetti, B.; Scanu, B.; Franceschini, A.; Ragazzi, A. Endemic and emerging pathogens threatening cork oak trees: Management options for conserving a unique forest ecosystem. Plant Dis. 2016, 100, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Sena, K.; Crocker, E.; Vincelli, P.; Barton, C. Phytophthora cinnamomi as a driver of forest change: Implications for conservation and management. For. Ecol. Manag. 2018, 409, 799–807. [Google Scholar] [CrossRef]
- Brasier, C.M.; Robredo, F.; Ferraz, J.F.P. Evidence for Phytophthora cinnamomi involvement in Iberian oak decline. Plant Pathol. 1993, 42, 140–145. [Google Scholar] [CrossRef]
- Robin, C.; Desprez-Loustau, M.L.; Capron, G.; Delatour, C. First record of Phytophthora cinnamomi on cork and holm oaks in France and evidence of pathogenicity. Ann. Sci. For. INRA/EDP Sci. 1998, 55, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Gallego, F.J.; de Algaba, A.P.; Fernandez-Escobar, R. Etiology of oak decline in Spain. Eur. J. For. Pathol. 1999, 29, 17–27. [Google Scholar] [CrossRef]
- Sánchez, M.E.; Caetano, P.; Ferraz, J.; Trapero, A. Phytophtora disease of Quercus ilex in south-western Spain. For. Pathol. 2002, 32, 5–18. [Google Scholar] [CrossRef]
- Camilo-Alves, C.S.P.; da Clara, M.I.E.; Ribeiro, N.A. Decline of Mediterranean oak trees and its association with Phytophthora cinnamomi: A review. Eur. J. For. Res. 2013, 132, 411–432. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Franceschini, A.; Anselmi, N.; Vannini, A.; Vettraino, A.M. Occurrence of Phytophthora cinnamomi in cork oak forests in Italy. For. Pathol. 2013, 43, 340–343. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; López-García, C.; León, M.; García-Jiménez, J.; Abad-Campos, P.; Jung, T. Previously unrecorded low temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in Eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Gómez, F.J.; Navarro-Cerrillo, R.M.; de Luque, A.P.; Owald, W.; Vannini, A.; Morales-Rodríguez, C. Assessment of functional and structural changes of soil fungal and oomycete communities in holm oak declined dehesas through metabarcoding analysis. Sci. Rep. 2019, 9, 5315. [Google Scholar] [CrossRef] [Green Version]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The use of genus-specific amplicon pyrosequencing to assess Phytophthora species diversity using eDNA from soil and water in northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Sala, B.; Gramaje, D.; Abad-Campos, P.; Berbegal, M. Diversity of Phytophthora species associated with Quercus ilex L. in Three Spanish Regions Evaluated by NGS. Forests 2019, 10, 979. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Osimani, L.; Morales-Rodríguez, C. Decline and mortality of evergreen oaks in a protected area in Central Italy driven by the pathogenic activity of the invasive Phytophthora cinnamomi and Phytophthora multivora. In Proceedings of the Book of Abstracts Proceedings of the 9th Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09, Phytophthora Diseases in Forests and Natural Ecosystems, La Maddalena, Italy, 17–25 October 2019; p. 88. [Google Scholar]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Scanu, B.; Hunter, G.C.; Linaldeddu, B.T.; Franceschini, A.; Maddau, L.; Jung, T.; Denman, S. A taxonomic re-evaluation reveals that Phytophthora cinnamomi and P. cinnamomi var. parvispora are separate species. For. Pathol. 2014, 44, 1–20. [Google Scholar] [CrossRef]
- Brasier, C.M. Physiology of Reproduction in Phytophthora. Ph.D. Thesis, University of Hull, Kingston upon Hull, UK, 1967. [Google Scholar]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef]
- Hüberli, D.; Hardy, G.E.S.J.; White, D.; Williams, N.; Burgess, T.I. Fishing for Phytophthora from Western Australia’s waterways: A distribution and diversity survey. Australas. Plant Path. 2013, 42, 251–260. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of Phytophthora species from declining Mediterranean maquis vegetation, including two new species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Cooke, D.E.L.; Blaschke, H.; Duncan, J.M.; Oßwald, W. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef] [Green Version]
- Nechwatal, J.; Bakonyi, J.; Cacciola, S.O.; Cooke, D.E.L.; Jung, T.; Nagy, Z.A.; Vannini, A.; Vettraino, A.M.; Brasier, C.M. The morphology, behaviour and molecular phylogeny of Phytophthora taxon Salixsoil and its redesignation as Phytophthora lacustris sp. nov. Plant Pathol. 2013, 62, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Safaiefarahani, B.; Mostowfizadeh-Ghalamfarsa, R.; Hardy, G.E.S.J.; Burgess, T.I. Re-evaluation of the Phytophthora cryptogea species complex and the description of a new species, Phytophthora pseudocryptogea sp. nov. Mycol. Prog. 2015, 14, 108. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Scanu, B.; Seress, D.; Kovács, G.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, K.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal DNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Hall, T. Bio Edit Version 5.0.6; Department of Microbiology, North Carolina State University: Raleigh, NC, USA, 2001. [Google Scholar]
- Brasier, C.M.; Kirk, S.A. Designation of the EAN and NAN races of Ophiostoma novo-ulmi as subspecies: Their perithecial size differences and geographical distributions. Mycol. Res. 2001, 105, 547–554. [Google Scholar] [CrossRef]
- Scanu, B.; Webber, J.F. Dieback and mortality of Nothofagus in Britain: Ecology, pathogenicity and sporulation potential of the causal agent Phytophthora pseudosyringae. Plant Pathol. 2016, 65, 26–36. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Luisi, N.; Anselmi, N.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol. 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Corcobado, T.; Cubera, E.; Pérez-Sierra, A.; Jung, T.; Solla, A. First report of Phytophthora gonapodyides involved in the decline of Quercus ilex in xeric conditions in Spain. New Dis. Rep. 2010, 22, 33. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, M.E.; Andicoberry, S.; Trapero, A. Pathogenicity of three Phytophthora spp. causing late seedling rot of Quercus ilex ssp. ballota. For. Pathol. 2005, 35, 115–125. [Google Scholar]
- Corcobado, T.; Cubera, E.; Juárez, E.; Moreno, G.; Solla, A. Drought events determine performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi. Agric. For. Meteorol. 2014, 192–193, 1–8. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera Island (Italy). For. Pathol. 2013, 44, 191–200. [Google Scholar] [CrossRef]
- Frisullo, S.; Lima, G.; Magnano di San Lio, G.; Camele, I.; Melissano, L.; Puglisi, I.; Pane, A.; Agosteo, G.E.; Prudente, L.; Cacciola, S.O. Phytophthora cinnamomi involved in the decline of holm oak (Quercus ilex) stands in southern Italy. For. Sci. 2018, 64, 290–298. [Google Scholar] [CrossRef]
- González, M.; Romero, M.; García, L.; Gomez-Aparicio, L.; Serrano, M.S. Unravelling the role of drought as predisposing factor for Quercus suber decline caused by Phytophthora cinnamomi. Eur. J. Plant. Pathol. 2020, 156, 1015–1021. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poortner, M. One-Hundred of the World’s Worst Invasive Alien Species. A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group, International Union for Conservation of Nature (IUCN): Gland, Switzerland, 2000; Available online: http://www.issg.org (accessed on 28 July 2020).
- Burgess, T.I.; Scott, J.K.; Mcdougall, K.L.; Stukely, M.J.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Glob. Chang. Biol. 2017, 23, 1661–1674. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M. Oak tree mortality in Iberia. Nature 1992, 360, 539. [Google Scholar] [CrossRef]
- Moreira, A.C.; Martins, J.M.S. Influence of site factor on the impact of Phytophthora cinnamomi in cork oak stands in Portugal. For. Pathol. 2005, 35, 145–162. [Google Scholar] [CrossRef]
- Robin, C.; Capron, G.; Desprez-Loustau, M.L. Root infection by Phytophthora cinnamomi in seedlings of three oak species. Plant Pathol. 2001, 50, 708–716. [Google Scholar] [CrossRef]
- Moralejo, E.; García-Muñoz, J.A.; Descals, E. Susceptibility of Iberian trees to Phytophthora ramorum and P. cinnamomi. Plant Pathol. 2009, 58, 271–283. [Google Scholar] [CrossRef]
- Magri, D.; Fineschi, S.; Bellarosa, R.; Buonamici, A.; Sebastiani, F.; Schirone, B.; Simeone, M.C.; Vendramin, G.G. The distribution of Quercus suber chloroplast haplotypes matches the palaeogeographical history of the western Mediterranean. Mol. Ecol. 2007, 16, 5259–5266. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. First report of Phytophthora quercina from oak forests in Austria. Plant Pathol. 2003, 52, 403. [Google Scholar] [CrossRef]
- Ruiz Gómez, F.J.; Pérez-de-Luque, A.; Sánchez-Cuesta, R.; Quero, J.L.; Navarro Cerrillo, R.M. Differences in the response to acute drought and Phytophthora cinnamomi Rands Infection in Quercus ilex L. Seedlings. Forests 2018, 9, 634. [Google Scholar] [CrossRef] [Green Version]
- Robin, C.; Dupuis, F.; Desprez-Loustau, M.L. Seasonal changes in Northern red oak susceptibility to Phytophthora cinnamomi. Plant Dis. 1994, 78, 369–374. [Google Scholar] [CrossRef]
- Luque, J.; Parladé, J.; Pera, J. Seasonal changes in susceptibility of Quercus suber to Botryosphaeria stevensii and Phytophthora cinnamomi. Plant Pathol. 2002, 51, 338–345. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathol. 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Riddell, C.E.; Frederickson-Matika, D.; Armstrong, A.C.; Elliot, M.; Forster, J.; Hedley, P.E.; Morris, J.; Thorpe, P.; Cooke, D.E.L.; Pritchard, L.; et al. Metabarcoding reveals a high diversity of woody host-associated Phytophthora spp. in soils at public gardens and amenity woodlands in Britain. PeerJ 2019, 7, e6931. [Google Scholar] [CrossRef] [Green Version]
- Garbelotto, M.; Frankel, S.; Scanu, B. Soil-and waterborne Phytophthora species linked to recent outbreaks in Northern California restoration sites. Calif. Agric. 2018, 72, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Roberto Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora species in protected natural plots in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Sechi, C.; Seddaiu, S.; Linaldeddu, B.T.; Franceschini, A.; Scanu, B. Dieback and mortality of Pinus radiata trees in Italy associated with Phytophthora cryptogea. Plant Dis. 2014, 98, 159. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Natili, G.; Anselmi, N.; Vannini, A. Recovery and pathogenicity of Phytophthora species associated with resurgence of ink disease on Castanea sativa in Italy. Plant Pathol. 2001, 50, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Belisario, A.; Maccaroni, M.; Vettorazzo, M. First report of Phytophthora cambivora causing bleeding cankers and dieback on beech (Fagus sylvatica) in Italy. Plant Dis. 2006, 90, 1362. [Google Scholar] [CrossRef] [PubMed]
- Talgø, V.; Herrero, M.L.; Toppe, B.; Klemsdal, S.S.; Stensvand, A. First report of root rot and stem canker caused by Phytophthora cambivora on noble fir (Abies procera) for bough production in Norway. Plant Dis. 2006, 90, 682. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J.; Burgess, T.I. Surveys of soil and water reveal a goldmine of Phytophthora diversity in South African natural ecosystems. IMA Fungus 2013, 4, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenkovic, I.; Corcobado, T.; Tomsovsky, M.; Panek, M.; Bakonyi, J.; Maia, C.; et al. A survey in natural forest ecosystems of Vietnam reveals high diversity of both new and described Phytophthora taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Cooke, D.E.L.; Duncan, J.M.; Hansen, E.M. Multiple new phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides–P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef]
- Jung, T.; Stukely, M.J.C.; Hardy, G.E.S.J.; White, D.; Paap, T.; Dunstan, W.A.; Burgess, T.I. Multiple new Phytophthora species from ITS Clade 6 associated with natural ecosystems in Australia: Evolutionary and ecological implications. Persoonia 2011, 26, 13–39. [Google Scholar] [CrossRef] [Green Version]
- Scanu, B.; Linaldeddu, B.T.; Peréz-Sierra, A.; Deidda, A.; Franceschini, A. Phytophthora ilicis as a leaf and stem pathogen of Ilex aquifolium in Mediterranean islands. Phytopathol. Mediterr. 2014, 53, 480–490. [Google Scholar]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First Report of Root Rot Caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef]
- Brown, A.V.; Brasier, C.M. Colonization of tree xylem by Phytophthora ramorum, P. kernoviae and other Phytophthora species. Plant Pathol. 2007, 56, 227–241. [Google Scholar] [CrossRef]
- Brasier, C.M.; Sanchez-Hernandez, E.; Kirk, S.A. Phytophthora inundata sp. nov., a part heterothallic pathogen of trees and shrubs in wet or flooded soils. Mycol. Res. 2003, 107, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Cline, E.T.; Farr, D.F.; Rossman, A.Y. Synopsis of Phytophthora with accurate scientific names, host range, and geographic distribution. Plant Health Prog. 2008, 9. [Google Scholar] [CrossRef]
- Ginetti, B.; Moricca, S.; Squires, J.N.; Cooke, D.E.L.; Ragazzi, A.; Jung, T. Phytophthora acerina sp. nov., a new species causing bleeding cankers and dieback of Acer pseudoplatanus trees in planted forests in northern Italy. Plant Pathol. 2014, 63, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Seddaiu, S.; Linaldeddu, B.T. First Report of Phytophthora acerina, P. plurivora, and P. pseudocryptogea associated with declining common alder trees in Italy. Plant Dis. 2020, 104, 1874. [Google Scholar] [CrossRef]
- Mora-Sala, B.; Abad-Campos, P.; Berbegal, M. Response of Quercus ilex seedlings to Phytophthora spp. root infection in a soil infestation test. Eur. J. Plant Pathol. 2018, 154, 2115–2225. [Google Scholar] [CrossRef]
- Vitale, S.; Luongo, L.; Galli, M.; Belisario, A. First report of Phytophthora hydropathica causing wilting and shoot dieback on viburnum in Italy. Plant Dis. 2014, 98, 1582. [Google Scholar] [CrossRef]
- Burgess, T.I. Molecular characterization of natural hybrids formed between five related indigenous clade 6 Phytophthora species. PLoS ONE 2015, 10, e0134225. [Google Scholar] [CrossRef] [Green Version]
- Nagel, J.H.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J.; Hardy, G.E.S.J.; Stukely, M.J.C.; Burgess, T.I. Characterization of Phytophthora hybrids from ITS clade 6 associated with riparian ecosystems in South Africa and Australia. Fungal Biol. 2013, 117, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Dickie, I.A.; Wakelin, A.M.; Martínez-García, L.B.; Richardson, S.J.; Makiola, A.; Tylianakis, J.M. Oomycetes along a 120,000-year temperate rainforest ecosystem development chrono sequence. Fungal Ecol. 2019, 39, 192–200. [Google Scholar] [CrossRef]
- Català, S.; Berbegal, M.; Pérez-Sierra, A.; Abad-Campos, P. Metabarcoding and development of new real-time specific assays reveal Phytophthora species diversity in holm oak forests in eastern Spain. Plant Pathol. 2017, 66, 115–123. [Google Scholar] [CrossRef]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Phytophthora species associated with roots of native and non-native trees in natural and managed forests. Microb. Ecol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Corcobado, T.; Miranda-Torres, J.J.; Martín-García, J.; Jung, T.; Solla, A. Early survival of Quercus ilex subspecies from different populations after infections and coinfections by multiple Phytophthora species. Plant Pathol. 2017, 66, 792–804. [Google Scholar] [CrossRef]
- Domìnguez-Begines, J.; Avila, J.M.; Garcìa, L.V.; Gòmez-Aparicio, L. Soil-borne pathogens as determinants of regeneration patterns at community level in Mediterranean forests. N. Phytol. 2020, 227, 588–600. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The geographic location of the 39 Quercus suber stands investigated in this study (QS1–QS39). The green area represents the geographic distribution of cork oak in Sardinia. A map of Italy is inserted into the top right corner showing the location of Sardinia.
Figure 1.
The geographic location of the 39 Quercus suber stands investigated in this study (QS1–QS39). The green area represents the geographic distribution of cork oak in Sardinia. A map of Italy is inserted into the top right corner showing the location of Sardinia.

Figure 2.
Symptoms caused by Phytophthora species on Quercus suber. (a) The sudden death of mature trees; (b) severe dieback and mortality of trees in 10-year-old afforestation; (c) trees showing a chronic decline with increasing transparency and wilting on the crown; (d) widespread dead and declining trees in a natural stand; (e) bleeding cankers at the stem base of a young tree.
Figure 2.
Symptoms caused by Phytophthora species on Quercus suber. (a) The sudden death of mature trees; (b) severe dieback and mortality of trees in 10-year-old afforestation; (c) trees showing a chronic decline with increasing transparency and wilting on the crown; (d) widespread dead and declining trees in a natural stand; (e) bleeding cankers at the stem base of a young tree.

Figure 3.
Diversity and frequency of the 8 soilborne (a) and 9 waterborne (b) Phytophthora species detected in this study; the horizontal axis is the count of sites. (c) Mosaic plot showing the distribution of soilborne (brown bars) and waterborne (blue bars) Phytophthora species grouped for phylogenetic clades. The bar width is proportional to the number of species, while bar heights show the relative proportion of soilborne and waterborne Phytophthora species per clade. CIN = P. cinnamomi, QUE = P. quercina, PSE = P. pseudocryptogea, TYR = P. tyrrhenica, GON = P. gonapodyides, PSY = P. psychrophila, CAM = P. ×cambivora, SYR = P. syringae, BIL = P. bilorbang, LAC = P. lacustris, PLU = P. plurivora, CHL = P. chlamydospora, HYD = P. hydropathica, MUL = P. ×multiformis.
Figure 3.
Diversity and frequency of the 8 soilborne (a) and 9 waterborne (b) Phytophthora species detected in this study; the horizontal axis is the count of sites. (c) Mosaic plot showing the distribution of soilborne (brown bars) and waterborne (blue bars) Phytophthora species grouped for phylogenetic clades. The bar width is proportional to the number of species, while bar heights show the relative proportion of soilborne and waterborne Phytophthora species per clade. CIN = P. cinnamomi, QUE = P. quercina, PSE = P. pseudocryptogea, TYR = P. tyrrhenica, GON = P. gonapodyides, PSY = P. psychrophila, CAM = P. ×cambivora, SYR = P. syringae, BIL = P. bilorbang, LAC = P. lacustris, PLU = P. plurivora, CHL = P. chlamydospora, HYD = P. hydropathica, MUL = P. ×multiformis.

Figure 4.
Root length reduction (%) compared to control seedling roots of 2-year-old seedlings of Quercus suber after 5 months of growth in soil infested with Phytophthora spp. obtained in this study (a). Mean lesion sizes, caused by isolates of Phytophthora following inoculation and incubation of logs for 4 weeks (b). Different letters above bars indicate significant differences according to Fisher’s protected least significant difference (LSD) test (p = 0.05). Bars represent standard errors. CIN = P. cinnamomi, PSE = P. pseudocryptogea, TYR = P. tyrrhenica, QUE = P. quercina, GON = P. gonapodyides, PSY = P. psychrophila, C = control.
Figure 4.
Root length reduction (%) compared to control seedling roots of 2-year-old seedlings of Quercus suber after 5 months of growth in soil infested with Phytophthora spp. obtained in this study (a). Mean lesion sizes, caused by isolates of Phytophthora following inoculation and incubation of logs for 4 weeks (b). Different letters above bars indicate significant differences according to Fisher’s protected least significant difference (LSD) test (p = 0.05). Bars represent standard errors. CIN = P. cinnamomi, PSE = P. pseudocryptogea, TYR = P. tyrrhenica, QUE = P. quercina, GON = P. gonapodyides, PSY = P. psychrophila, C = control.


{kind=link}
{kind=link}
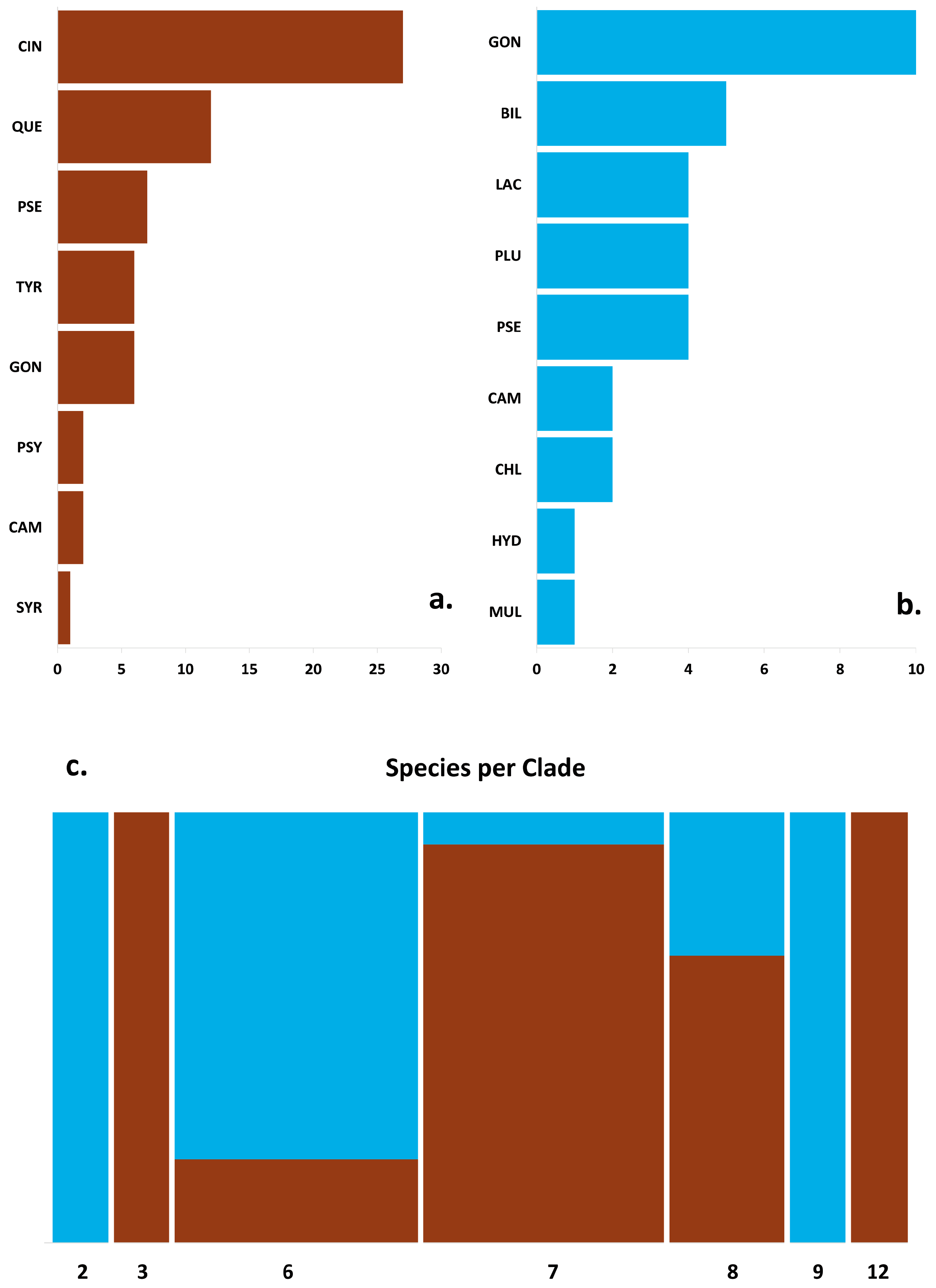{kind=link}
{kind=link}
Table 1.
Location, forest type, and altitude of the 39 declining cork oak stands sampled in Sardinia and Phytophthora taxa isolated from the rhizosphere soil samples collected in this study.
Table 1.
Location, forest type, and altitude of the 39 declining cork oak stands sampled in Sardinia and Phytophthora taxa isolated from the rhizosphere soil samples collected in this study.
| Site | Location (Municipality) | Forest Type a | Altitude (m a.s.l) | Trees Sampled (No.) | Positive Trees (No.) | Phytophthora spp. b | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAM | CIN | GON | PSE | PSY | QUE | SYR | TYR | ||||||
| QS1 | Abbasanta | For | 352 | 4 | 2 | 1 | 1 | ||||||
| QS2 | Alà dei Sardi | For | 620 | 4 | 2 | 1 | |||||||
| QS3 | Alà dei Sardi | For | 580 | 5 | 3 | 1 | 2 | ||||||
| QS4 | Alghero | Aff | 80 | 6 | 5 | 4 | 1 | 1 | |||||
| QS5 | Alghero | For | 122 | 4 | 2 | 2 | |||||||
| QS6 | Berchidda | For | 201 | 8 | 2 | 2 | |||||||
| QS7 | Bolotana | Aff | 340 | 4 | 4 | 4 | |||||||
| QS8 | Bono | For | 841 | 6 | 4 | 2 | 1 | 1 | |||||
| QS9 | Bortigiadas | For | 158 | 14 | 10 | 5 | 2 | 4 | |||||
| QS10 | Bortigiadas | For | 157 | 12 | 11 | 1 | 4 | 4 | 2 | 2 | 5 | 3 | |
| QS11 | Buddusò | For | 762 | 11 | 3 | 3 | |||||||
| QS12 | Bultei | For | 503 | 6 | 4 | 1 | 2 | 1 | |||||
| QS13 | Illorai | For | 214 | 4 | 2 | 2 | |||||||
| QS14 | La Maddalena | Aff | 118 | 6 | 4 | 3 | 2 | ||||||
| QS15 | Loiri P.S.P. | Aff | 240 | 16 | 12 | 8 | |||||||
| QS16 | Lula | Aff | 560 | 4 | 3 | 2 | 3 | ||||||
| QS17 | Lula | Aff | 380 | 4 | 4 | 2 | 4 | ||||||
| QS18 | Monti | For | 268 | 8 | 7 | 4 | 1 | 2 | |||||
| QS19 | Monti | For | 260 | 4 | 2 | 1 | 2 | ||||||
| QS20 | Montresta | For | 480 | 10 | 8 | 3 | 2 | 2 | 4 | ||||
| QS21 | Nuoro | For | 490 | 10 | 3 | 3 | |||||||
| QS22 | Olbia | For | 60 | 4 | 4 | 4 | |||||||
| QS23 | Onanì | Aff | 455 | 4 | 1 | 1 | |||||||
| QS24 | Orgosolo | Aff | 568 | 6 | 6 | 4 | 2 | ||||||
| QS25 | Orotelli | Aff | 410 | 4 | 4 | 4 | 3 | ||||||
| QS26 | Padru | For | 363 | 8 | 6 | 4 | 2 | ||||||
| QS27 | Ploaghe | For | 320 | 11 | 5 | 3 | 3 | ||||||
| QS28 | San Pantaleo | For | 180 | 4 | 4 | 4 | |||||||
| QS29 | San Teodoro | For | 13 | 8 | 5 | 4 | 2 | ||||||
| QS30 | San Teodoro | Aff | 118 | 6 | 4 | 4 | |||||||
| QS31 | Siniscola | For | 125 | 12 | 6 | 6 | |||||||
| QS32 | Siniscola | Aff | 248 | 8 | 8 | 6 | 2 | ||||||
| QS33 | Telti | Aff | 210 | 6 | 2 | 2 | |||||||
| QS34 | Tempio P. | For | 438 | 4 | 3 | 2 | 1 | ||||||
| QS35 | Tempio P. | Aff | 465 | 30 | 27 | 27 | |||||||
| QS36 | Tergu | For | 346 | 12 | 12 | 1 | 1 | 2 | 5 | 4 | |||
| QS37 | Villanova M. | For | 538 | 10 | 4 | 4 | |||||||
| QS38 | Villanova M. | Aff | 440 | 3 | 2 | 2 | |||||||
| QS39 | Villanova M. | For | 428 | 5 | 2 | 2 | |||||||
| GenBank accession numbers | MT823269 | MT328694 MT328695 MT328696 | MT823270 | MT328706 | MT328708 | MT328709 MT328710 | MT328711 | MT328712 | |||||
a For = forest, Aff = afforestation. b CAM = P. ×cambivora, CIN = P. cinnamomi, GON = P. gonapodyides, PSE = P. pseudocryptogea, PSY = P. psychrophila, QUE = P. quercina, SYR = P. syringae, TYR = P. tyrrhenica.
Table 2.
Location, name, and typology of the 10 watercourses sampled across declining cork oak stands in Sardinia and Phytophthora taxa identified.
Table 2.
Location, name, and typology of the 10 watercourses sampled across declining cork oak stands in Sardinia and Phytophthora taxa identified.
| Site No. | River/Stream | Description | Phytophthora spp. a | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| BIL | CAM | CHL | GON | HYD | LAC | MUL | PLU | PSE | |||
| QS1 | Pizziu | Permanent river | + | + | |||||||
| QS2 | Sa Labia | Permanent river | + | + | |||||||
| QS6 | Berchidda | Permanent river | + | + | |||||||
| QS8 | Monte Pisanu | Intermittent stream | + | + | + | + | |||||
| QS9 | Santu Brancazzu | Intermittent stream | + | + | + | + | + | + | |||
| QS11 | Sos Canales | Intermittent stream | + | + | |||||||
| QS12 | Olletto | Intermittent stream | + | + | + | + | + | ||||
| QS13 | Tirso | Permanent river | + | + | |||||||
| QS21 | Errede | Intermittent stream | + | + | |||||||
| QS10 | Puddina | Intermittent stream | + | + | + | + | + | + | |||
| GenBank accessions | MT328690 MT328691 MT328692 | MT328713 | MT328693 | MT328697 MT328698 MT328699 | MT822885 | MT328700 MT328701 | MT822886 | MT328704 MT328705 | MT328707 | ||
a BIL = P. bilorbang, CAM = P. ×cambivora, CHL = P. chlamydospora, GON = P. gonapodyides, HYD = P. hydropathica, LAC = P. lacustris, MUL = P. ×multiformis, PLU = P. plurivora, PSE = P. pseudocryptogea.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Seddaiu, S.; Brandano, A.; Ruiu, P.A.; Sechi, C.; Scanu, B. An Overview of Phytophthora Species Inhabiting Declining Quercus suber Stands in Sardinia (Italy). Forests 2020, 11, 971. https://doi.org/10.3390/f11090971
AMA Style
Seddaiu S, Brandano A, Ruiu PA, Sechi C, Scanu B. An Overview of Phytophthora Species Inhabiting Declining Quercus suber Stands in Sardinia (Italy). Forests. 2020; 11(9):971. https://doi.org/10.3390/f11090971
Chicago/Turabian StyleSeddaiu, Salvatore, Andrea Brandano, Pino Angelo Ruiu, Clizia Sechi, and Bruno Scanu. 2020. "An Overview of Phytophthora Species Inhabiting Declining Quercus suber Stands in Sardinia (Italy)" Forests 11, no. 9: 971. https://doi.org/10.3390/f11090971
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.