Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
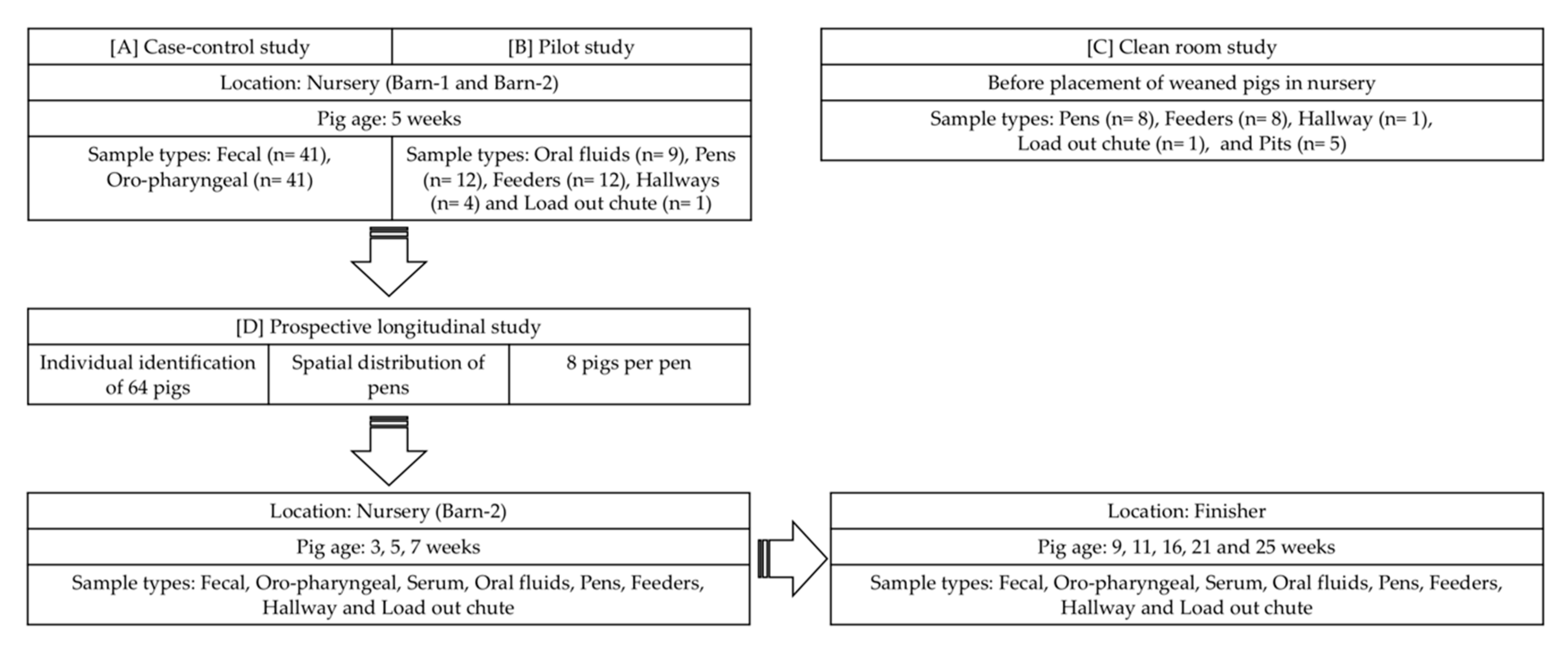
2.2. Study Designs
2.3. Sample Collection and Processing
2.4. Case-Control Study
2.5. Pilot Study
2.6. Clean Room Study
2.7. Prospective Longitudinal Study
2.8. PoAstV3 RT-qPCR
2.9. Statistical Analysis
3. Results
3.1. Case-Control Study
3.2. Pilot Study
3.3. Clean Room Study
3.4. Prospective Longitudinal Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maclachlan, N.J.; Dubovi, E.J.; Barthold, S.W.; Swayne, D.E.; Winton, J.R. Fenner’s Veterinary Virology, 5th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Appleton, H.; Higgins, P.G. Viruses and gastroenteritis in infants. Lancet 1975, 305, 1297. [Google Scholar] [CrossRef]
- Madeley, C.R.; Cosgrove, B.P. Viruses in infantile gastroenteritis. Lancet 1975, 306, 451–452. [Google Scholar] [CrossRef]
- Madeley, C.R.; Cosgrove, B.P. 28 nm particles in faeces in infantile gastroenteritis. Lancet 1975, 7932, 451–452. [Google Scholar] [CrossRef]
- Mendez, E.; Arias, C.F. Astroviruses. In Fields Virology, 5th ed.; Howley, P.M., Knipe, D.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1, pp. 981–1000. [Google Scholar]
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus Taxonomy. In Astrovirus Research; Schultz-Cherry, S., Ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Laurin, M.A.; Dastor, M.; L’Homme, Y. Detection and genetic characterization of a novel pig astrovirus: Relationship to other astroviruses. Arch. Virol. 2011, 156, 2095–2099. [Google Scholar] [CrossRef]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef]
- Shan, T.; Li, L.; Simmonds, P.; Wang, C.; Moeser, A.; Delwart, E. The Fecal Virome of Pigs on a High-Density Farm. J. Virol. 2011, 85, 11697–11708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.T.; Giménez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef]
- Englund, L.; Chriél, M.; Dietz, H.H.; Hedlund, K.O. Astrovirus epidemiologically linked to pre-weaning diarrhoea in mink. Vet. Microbiol. 2002, 85, 1–11. [Google Scholar] [CrossRef]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huilier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M.; et al. Astrovirus MLB2, a New Gastroenteric Virus Associated with Meningitis and Disseminated Infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Wildén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtnenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Gatto, I.R.H.; Matias, F.F.; Gauger, P.; Schwartz, K.; et al. Porcine Astrovirus Type 3 in Central Nervous System of Swine with Polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097–2100. [Google Scholar] [CrossRef] [Green Version]
- Matias, F.F.; Bradner, L.K.; Burrough, E.R.; Cooper, V.L.; Derscheid, R.J.; Gauger, P.C.; Harmon, K.M.; Madson, D.; Piñeyro, P.E.; Schwartz, K.J.; et al. Polioencephalomyelitis in Domestic Swine Associated with Porcine Astrovirus Type 3. Vet. Pathol. 2019, 57, 82–89. [Google Scholar] [CrossRef]
- Rawal, G.; Matias, F.F.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Mueller, A.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Detection and Cellular Tropism of Porcine Astrovirus Type 3 on Breeding Farms. Viruses 2019, 11, 1051. [Google Scholar] [CrossRef] [Green Version]
- Boros, A.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of Neuroinvasive Astrovirus Associated with Encephalomyelitis, Weakness, and Paralysis among Weaned Pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1999–2010. [Google Scholar] [CrossRef]
- Prickett, J.R.; Kim, W.; Simer, R.; Yoon, K.J.; Zimmerman, J. Oral-fluid samples for surveillance of commercial growing pigs for porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 infections. J. Swine Health Prod. 2008, 16, 86–91. [Google Scholar]
- Rotolo, M.L.; Yaxuan, S.; Wang, C.; Giménez-Lirola, L.; Baum, D.H.; Gauger, P.C.; Harmon, K.M.; Hoogland, M.; Main, R.; Zimmerman, J.J. Sampling guidelines for oral fluid-based of surveys of group-housed animals. Vet. Microbiol. 2017, 209, 20–29. [Google Scholar] [CrossRef]
- Suzmilas, M. Explaining odds ratios. J. Can. Acad. Child Adolesc. Psychiatry 2010, 19, 227. [Google Scholar]
- Reuter, G.; Pankovics, P.; Boros, A. Identification of a novel astrovirus in a domestic pig in Hungary. Arch. Virol. 2011, 156, 125–128. [Google Scholar] [CrossRef]
- Kumthip, K.; Khamrin, P.; Saikruang, W.; Kongkaew, A.; Vachirachewin, R.; Ushijima, H.; Maneekarn, N. Detection and genetic characterization of porcine astroviruses in piglets with and without diarrhea in Thailand. Arch. Virol. 2018, 163, 1823–1829. [Google Scholar] [CrossRef]
- Amimo, J.O.; Okoth, E.; Junga, J.O.; Ogara, W.O.; Njahira, M.N.; Wang, Q.; Vlasova, A.N.; Saif, L.J.; Djikeng, A. Molecular detection and genetic characterization of kobuviruses and astroviruses in asymptomatic local pigs in East Africa. Arch. Virol. 2014, 159, 1313–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufkova, L.; Scigalkova, I.; Moutelikova, R.; Malenovska, H.; Prodelaova, J. Genetic diversity of porcine sapoviruses, kobuviruses, and astroviruses in asymptomatic pigs: An emerging new sapovirus GIII genotype. Arch. Virol. 2013, 158, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmet, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.L.; Anderson, G.; Virasami, A.; et al. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Fremond, M.L.; Perot, P.; Muth, E.; Cros, G.; Dumarest, M.; Mahlaoui, N.; Seilhean, D.; Desguerre, I.; Hébert, C.; Corre-Catelin, N.; et al. Next-Generation Sequencing for Diagnosis and Tailored Therapy: A Case Report of Astrovirus-Associated Progressive Encephalitis. J. Pediatric. Infect. Dis. Soc. 2015, 4, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kuroda, M.; Kasai, M.; Matsui, H.; Fukuyama, T.; Katano, H.; Tanaka-Taya, K. Acute Encephalopathy in an Immunocompromised Boy With astrovirus-MLB1 Infection Detected by Next Generation Sequencing. J. Clin. Virol. 2016, 78, 66–70. [Google Scholar] [CrossRef]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of neuroinvasive astrovirus infection in an immunocompromised adult with encephalitis by unbiased next-generation sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef] [Green Version]
- Wunderli, W.; Meerbach, A.; Guengoer, T.; Berger, C.; Griener, O.; Caduff, R.; Trkola, A.; Bossart, W.; Gerlach, D.; Schibler, M.; et al. Astrovirus infection in hospitalized infants with severe combined immunodeficiency after allogeneic hematopoietic stem cell transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef]
- Kurtz, J.B.; Lee, T.W.; Parsons, A.J. The action of alcohols on rotavirus, astrovirus and enterovirus. J. Hosp. Infect. 1980, 1, 321–325. [Google Scholar] [CrossRef]
- Schultz-Cherry, S.; King, D.J.; Koci, M.D. Inactivation of an astrovirus associated with poult enteritis mortality syndrome. Avian Dis. 2001, 45, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Abad, F.X.; Pintó, R.M.; Villena, C.; Gajardo, R.; Bosch, A. Astrovirus survival in drinking water. Appl. Environ. Microbiol. 1997, 63, 3119–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011; Volume 9. [Google Scholar]
- Brnić, D.; Prpić, J.; Keros, T.; Roić, B.; Starešina, V.; Jemeršić, L. Porcine astrovirus viremia and high genetic variability in pigs on large holdings in Croatia. Infect. Genet. Evol. 2013, 14, 258–264. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample Types | % | Mean Cq c | Mean gc d/mL of Sample e |
|---|---|---|---|
| (N b) | (Range) | (Range) | |
| Oral fluids | 100% a | 32.96 a | 7.95 × 103 |
| (9 of 9) | (28.02–38.28) | (2.64 × 105–2.39 × 102) | |
| Pens | 75% a | 32.23 a | 1.60 × 104 |
| (9 of 12) | (28.66–33.96) | (1.31 × 105–3.94 × 103) | |
| Feeders | 100% a | 33.86 a | 3.94 × 103 |
| (12 of 12) | (26.60–38.35) | (5.32 × 105–2.39 × 102) | |
| Hallways | 100% | 32.61 | 7.95 × 103 |
| (4 of 4) | (31.87–34.68) | (1.60 × 104–1.96 × 103) | |
| Load-out chute | 100% | 30.34 | 6.51 × 104 |
| (1 of 1) |
| Age (wks a) | Fecal Swabs | Oropharyngeal Swabs | Oral Fluids | |||
|---|---|---|---|---|---|---|
| % | Mean Cq c | % | Mean Cq | % | Mean Cq | |
| (N b) | (Range) | (N) | (Range) | (N) | (Range) | |
| 3 | 95% | 31.06 | 31% | 37.66 | NA d | NA d |
| (61 of 64) | (16.78–38.88) | (20 of 64) | (32.34–39.27) | |||
| 5 | 17% | 35.68 | 2% | 36.22 | 88% | 32.92 |
| (11 of 64) | (27.07–39.59) | (1 of 64) | - | (7 of 8) | (28.31–36.23) | |
| 7 | 47% | 33.63 | 5% | 37.77 | 100% | 31.95 |
| (28 of 60) | (24.07–39.55) | (3 of 60) | (36.90–38.55) | (8 of 8) | (28.63–34.49) | |
| 9 | 28% | 34.85 | 2% | 36.57 | 100% | 31.73 |
| (15 of 53) | (26.50–38.77) | (1 of 53) | - | (8 of 8) | (27.18–34.25) | |
| 11 | 88% | 34.29 | 24% | 37.01 | 100% | 32.20 |
| (44 of 50) | (27.38–38.54) | (12 of 50) | (34.43–39.94) | (8 of 8) | (30.52–32.50) | |
| 16 | 20% | 36.37 | 2% | 36.54 | 75% | 36.14 |
| (10 of 49) | (30.33–39.57) | (1 of 49) | - | (6 of 8) | (33.36–38.50) | |
| 21 | 4% | 37.37 | 0% | 0 | 75% | 35.73 |
| (2 of 47) | (36.29–38.45) | (0 of 47) | - | (6 of 8) | (33.13–38.28) | |
| 25 | 41% | 34.57 | 4% | 37.70 | 100% | 34.23 |
| (19 of 46) | (30.27–37.85) | (2 of 46) | (37.57–37.84) | (8 of 8) | (33.22–35.48) | |
| Age (wks a) | Pens | Feeders | Hallway | Load-Out Chute | ||||
|---|---|---|---|---|---|---|---|---|
| % | Mean Cq c | % | Mean Cq | % | Mean Cq | % | Mean Cq | |
| (N b) | (Range) | (N) | (Range) | (N) | (N) | |||
| 3 | 100% | 31.01 | 100% | 30.25 | 100% | 31.07 | 100% | 29.10 |
| (8 of 8) | (29.33–32.43) | (8 of 8) | (27.19–34.70) | (1 of 1) | (1 of 1) | |||
| 5 | 63% | 32.83 | 88% | 32.17 | 100% | 29.89 | 100% | 30.28 |
| (5 of 8) | (30.12–34.77) | (7 of 8) | (27.63–37.83) | (1 of 1) | (1 of 1) | |||
| 7 | 100% | 32.13 | 100% | 33.33 | 100% | 29.16 | 100% | 28.49 |
| (8 of 8) | (27.11–36.57) | (8 of 8) | (29.50–35.34) | (1 of 1) | (1 of 1) | |||
| 9 | 88% | 34.14 | 88% | 33.23 | 100% | 33.78 | 100% | 35.37 |
| (7 of 8) | (29.79–39.40) | (7 of 8) | (27.64–36.46) | (1 of 1) | (1 of 1) | |||
| 11 | 100% | 36.31 | 88% | 34.02 | 100% | 32.90 | 100% | 32.90 |
| (8 of 8) | (33.80–39.97) | (7 of 8) | (33.01–37.02) | (1 of 1) | (1 of 1) | |||
| 16 | 88% | 34.69 | 75% | 35.35 | 100% | 35.79 | 100% | 33.41 |
| (7 of 8) | (32.81–39.42) | (6 of 8) | (33.64–37.59) | (1 of 1) | (1 of 1) | |||
| 21 | 88% | 36.49 | 25% | 38.53 | 100% | 32.66 | 100% | 35.75 |
| (7 of 8) | (34.67–38.20) | (2 of 8) | (38.08–38.97) | (1 of 1) | (1 of 1) | |||
| 25 | 75% | 35.63 | 100% | 35.99 | 100% | 35.89 | 100% | 34.00 |
| (6 of 8) | (34.30–37.77) | (8 of 8) | (33.03–39.74) | (1 of 1) | (1 of 1) | |||
| Sample Types | % | Mean Cq c | Mean gc d/mL of Samplee |
|---|---|---|---|
| (N b) | (Range) | (Range) | |
| Pens | 88% a | 33.98 a | 3.94 × 103 |
| (56 of 64) | (27.11–39.97) | (5.32 × 105–5.90 × 101) | |
| Feeders | 83% a | 33.75 a | 3.94 × 103 |
| (53 of 64) | (27.19–39.74) | (5.32 × 105–5.90 × 101) | |
| Hallway | 100% a | 32.63 a | 7.95 × 103 |
| (8 of 8) | (29.16–35.89) | (1.31 × 105–9.72 × 102) | |
| Load-out chute | 100% a | 32.18 a | 1.60 × 104 |
| (8 of 8) | (28.49–35.75) | (1.31 × 105–9.72 × 102) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease. Viruses 2020, 12, 992. https://doi.org/10.3390/v12090992
Rawal G, Ferreyra FM, Macedo NR, Bradner LK, Harmon KM, Allison G, Linhares DCL, Arruda BL. Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease. Viruses. 2020; 12(9):992. https://doi.org/10.3390/v12090992
Chicago/Turabian StyleRawal, Gaurav, Franco Matias Ferreyra, Nubia R. Macedo, Laura K. Bradner, Karen M. Harmon, Grant Allison, Daniel C. L. Linhares, and Bailey L. Arruda. 2020. "Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease" Viruses 12, no. 9: 992. https://doi.org/10.3390/v12090992