Investigating the Association between Motor Function, Neuroinflammation, and Recording Metrics in the Performance of Intracortical Microelectrode Implanted in Motor Cortex


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Neural Probe Implantation Procedure
2.2. Electrophysiological Recordings
2.3. Signal Processing
2.4. Tissue Processing
2.5. Immunohistochemistry Histology
2.6. Quantitative Analysis
2.7. Behavior Training and Testing
2.8. Statistical Analyses
2.8.1. Statistical Analyses of Electrophysiology, Histology, and Motor Behavior Assessment
2.8.2. Network and Regression Analyses
3. Results
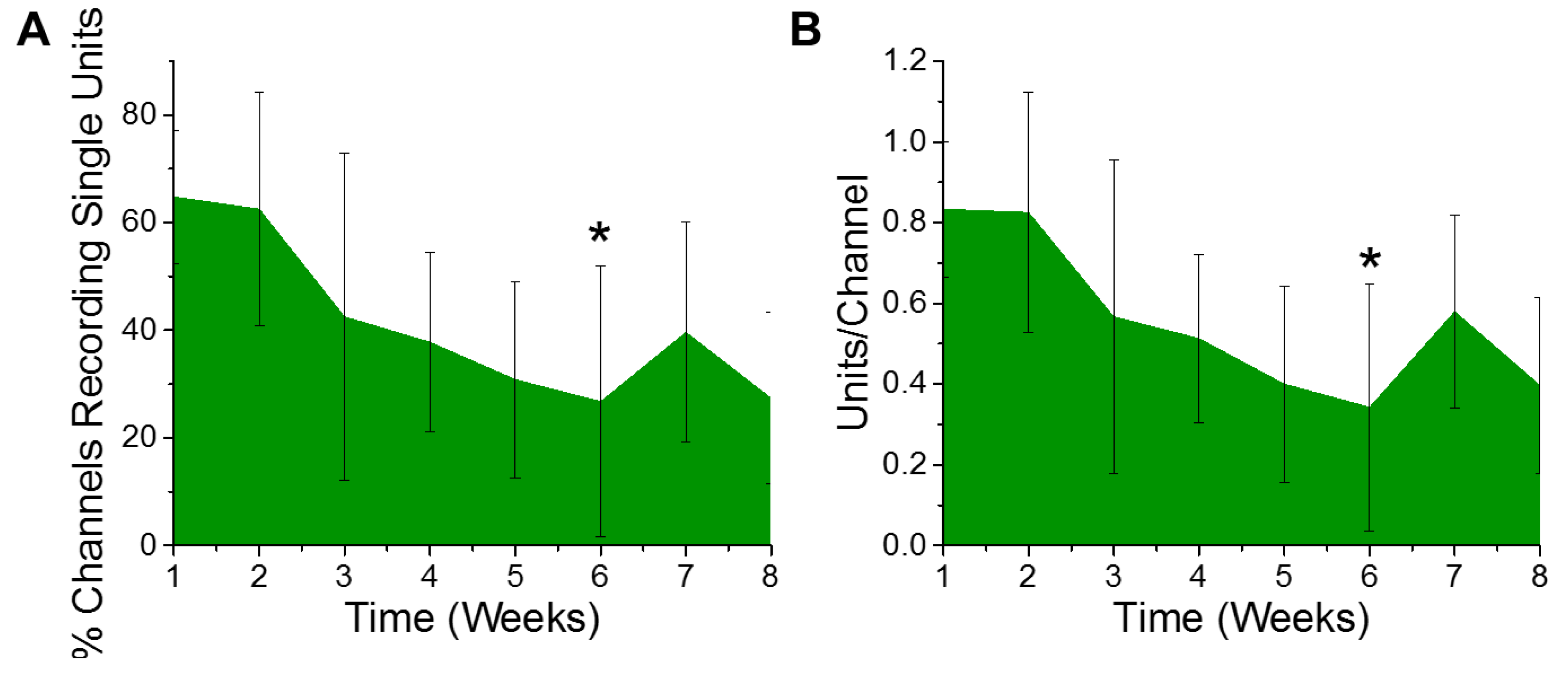
3.1. Electrophysiological Recordings
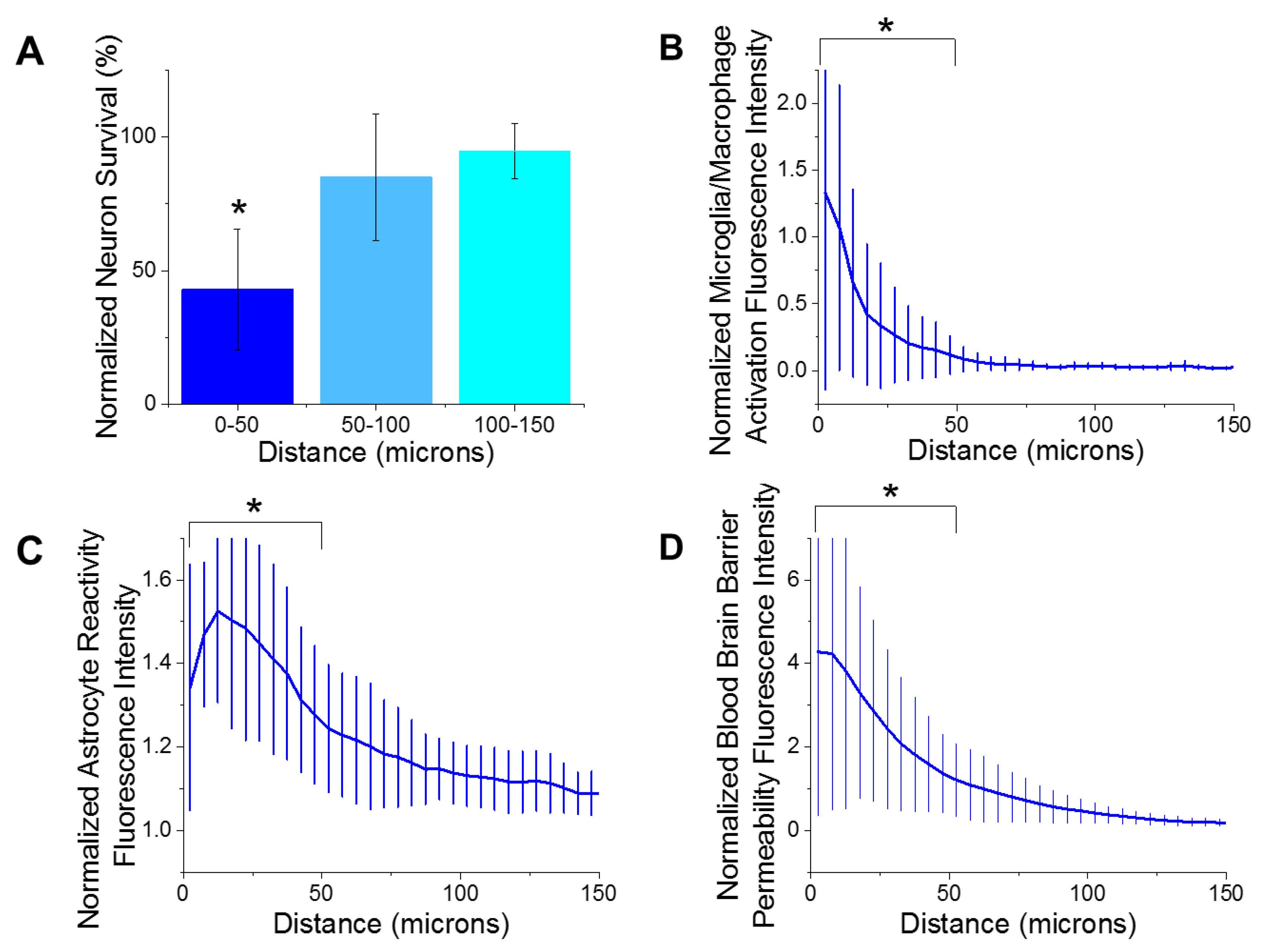
3.2. Histology
3.3. Motor Function Testing
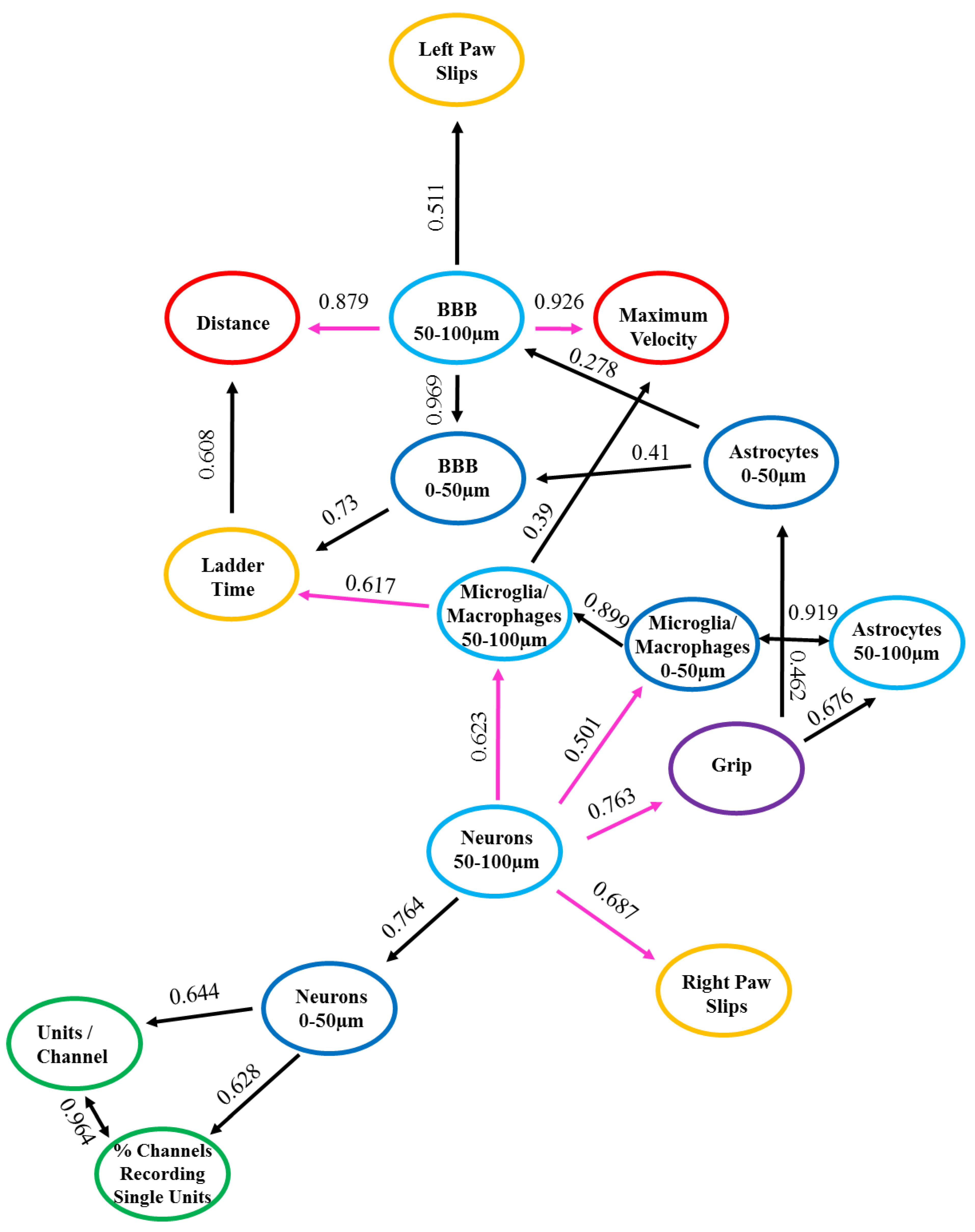
3.4. nFCA Network Relationships between Recording, Motor Behavior and Histology
3.4.1. Positive Relationships
3.4.2. Negative Relationships
3.5. Correlations between Recording, Motor Behavior, and Histology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilja, V.; Pandarinath, C.; Blabe, C.H.; Nuyujukian, P.; Simeral, J.D.; Sarma, A.A.; Sorice, B.L.; Perge, J.A.; Jarosiewicz, B.; Hochberg, L.R. Clinical translation of a high-performance neural prosthesis. Nat. Med. 2015, 21, 1142–1145. [Google Scholar]
- Hochberg, L.R.; Bacher, D.; Jarosiewicz, B.; Masse, N.Y.; Simeral, J.D.; Vogel, J.; Haddadin, S.; Liu, J.; Cash, S.S.; van der Smagt, P.; et al. Reach and grasp by people with tetraplegia using a neurally controlled robotic arm. Nature 2012, 485, 372–375. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, A.B.; Willett, F.R.; Young, D.R.; Memberg, W.D.; Murphy, B.A.; Miller, J.P.; Walter, B.L.; Sweet, J.A.; Hoyen, H.A.; Keith, M.W. Restoration of reaching and grasping movements through brain-controlled muscle stimulation in a person with tetraplegia: A proof-of-concept demonstration. Lancet 2017, 389, 1821–1830. [Google Scholar]
- Chestek, C.A.; Gilja, V.; Nuyujukian, P.; Foster, J.D.; Fan, J.M.; Kaufman, M.T.; Churchland, M.M.; Rivera-Alvidrez, Z.; Cunningham, J.P.; Ryu, S.I.; et al. Long-term stability of neural prosthetic control signals from silicon cortical arrays in rhesus macaque motor cortex. J. Neural Eng. 2011, 8, 045005. [Google Scholar] [CrossRef] [Green Version]
- Gilja, V.; Chestek, C.A.; Diester, I.; Henderson, J.M.; Deisseroth, K.; Shenoy, K.V. Challenges and opportunities for next-generation intracortically based neural prostheses. IEEE Trans. Bio-Med Eng. 2011, 58, 1891–1899. [Google Scholar] [CrossRef]
- Williams, J.C.; Rennaker, R.L.; Kipke, D.R. Long-term neural recording characteristics of wire microelectrode arrays implanted in cerebral cortex. Brain Res. Brain Res. Protoc. 1999, 4, 303–313. [Google Scholar]
- Rousche, P.J.; Normann, R.A. Chronic recording capability of the Utah Intracortical Electrode Array in cat sensory cortex. J. Neurosci. Methods 1998, 82, 1–15. [Google Scholar]
- Freire, M.A.; Morya, E.; Faber, J.; Santos, J.R.; Guimaraes, J.S.; Lemos, N.A.; Sameshima, K.; Pereira, A.; Ribeiro, S.; Nicolelis, M.A. Comprehensive analysis of tissue preservation and recording quality from chronic multielectrode implants. PLoS ONE 2011, 6, e27554. [Google Scholar] [CrossRef]
- Perge, J.A.; Zhang, S.; Malik, W.Q.; Homer, M.L.; Cash, S.; Friehs, G.; Eskandar, E.N.; Donoghue, J.P.; Hochberg, L.R. Reliability of directional information in unsorted spikes and local field potentials recorded in human motor cortex. J. Neural Eng. 2014, 11, 046007. [Google Scholar]
- Nolta, N.F.; Christensen, M.B.; Crane, P.D.; Skousen, J.L.; Tresco, P.A. BBB leakage, astrogliosis, and tissue loss correlate with silicon microelectrode array recording performance. Biomaterials 2015, 53, 753–762. [Google Scholar] [CrossRef]
- Cody, P.A.; Eles, J.R.; Lagenaur, C.F.; Kozai, T.D.; Cui, X.T. Unique electrophysiological and impedance signatures between encapsulation types: An analysis of biological Utah array failure and benefit of a biomimetic coating in a rat model. Biomaterials 2018, 161, 117–128. [Google Scholar]
- Mahajan, S.; Hermann, J.K.; Bedell, H.W.; Sharkins, J.A.; Chen, L.; Chen, K.; Meade, S.M.; Smith, C.S.; Rayyan, J.; Feng, H.; et al. Toward Standardization of Electrophysiology and Computational Tissue Strain in Rodent Intracortical Microelectrode Models. Front. Bioeng. Biotechnol. 2020, 8, 416. [Google Scholar]
- Jorfi, M.; Skousen, J.L.; Weder, C.; Capadona, J.R. Progress towards biocompatible intracortical microelectrodes for neural interfacing applications. J. Neural. Eng. 2015, 12, 011001. [Google Scholar] [CrossRef]
- Anderson, J.M. Inflammatory response to implants. ASAIO Trans. 1988, 34, 101–107. [Google Scholar]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Bjornsson, C.S.; Oh, S.J.; Al-Kofahi, Y.A.; Lim, Y.J.; Smith, K.L.; Turner, J.N.; De, S.; Roysam, B.; Shain, W.; Kim, S.J. Effects of insertion conditions on tissue strain and vascular damage during neuroprosthetic device insertion. J. Neural. Eng. 2006, 3, 196–207. [Google Scholar]
- Ravikumar, M.; Sunil, S.; Black, J.; Barkauskas, D.; Haung, A.Y.; Miller, R.H.; Selkirk, S.M.; Capadona, J.R. The Roles of Blood-derived Macrophages and Resident Microglia in the Neuroinflammatory Response to Implanted Intracortical Microelectrodes. Biomaterials 2014, 35, 8049–8064. [Google Scholar]
- Hermann, J.; Capadona, J. Understanding the Role of Innate Immunity in the Response to Intracortical Microelectrodes. Crit. Rev.™ Biomed. Eng. 2018, 46, 341–367. [Google Scholar]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Ereifej, E.S.; Rial, G.M.; Hermann, J.K.; Smith, C.S.; Meade, S.M.; Rayyan, J.M.; Chen, K.; Feng, H.; Capadona, J.R. Implantation of neural probes in the brain elicits oxidative stress. Front. Bioeng. Biotechnol. 2018, 6, 9. [Google Scholar]
- Bedell, H.W.; Schaub, N.J.; Capadona, J.R.; Ereifej, E.S. Differential expression of genes involved in the acute innate immune response to intracortical microelectrodes. Acta Biomater. 2019, 102, 205–219. [Google Scholar]
- Takmakov, P.; Ruda, K.; Phillips, K.S.; Isayeva, I.S.; Krauthamer, V.; Welle, C.G. Rapid evaluation of the durability of cortical neural implants using accelerated aging with reactive oxygen species. J. Neural Eng. 2015, 12, 026003. [Google Scholar]
- Potter, K.A.; Buck, A.C.; Self, W.K.; Callanan, M.E.; Sunil, S.; Capadona, J.R. The effect of resveratrol on neurodegeneration and blood brain barrier stability surrounding intracortical microelectrodes. Biomaterials 2013, 34, 7001–7015. [Google Scholar] [CrossRef]
- Prasad, A.; Xue, Q.S.; Sankar, V.; Nishida, T.; Shaw, G.; Streit, W.J.; Sanchez, J.C. Comprehensive characterization and failure modes of tungsten microwire arrays in chronic neural implants. J. Neural. Eng. 2012, 9, 056015. [Google Scholar] [CrossRef]
- Prasad, A.; Xue, Q.-S.; Dieme, R.; Sankar, V.; Mayrand, R.; Nishida, T.; Streit, W.J.; Sanchez, J.C. Abiotic-biotic characterization of Pt/Ir microelectrode arrays in chronic implants. Front. Neurosci. 2014, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, G.; Schultze, J.W.; Faßbender, F.; Buß, G.; Lüth, H.; Schöning, M.J. Passivation and corrosion of microelectrode arrays. Electrochim. Acta 1999, 44, 3865–3883. [Google Scholar]
- Barrese, J.C.; Aceros, J.; Donoghue, J.P. Scanning electron microscopy of chronically implanted intracortical microelectrode arrays in non-human primates. J. Neural Eng. 2016, 13, 026003. [Google Scholar]
- Haley, R.M.; Zuckerman, S.T.; Dakhlallah, H.; Capadona, J.R.; von Recum, H.A.; Ereifej, E.S. Resveratrol Delivery from Implanted Cyclodextrin Polymers Provides Sustained Antioxidant Effect on Implanted Neural Probes. Int. J. Mol. Sci. 2020, 21, 3579. [Google Scholar]
- Polikov, V.S.; Tresco, P.A.; Reichert, W.M. Response of brain tissue to chronically implanted neural electrodes. J. Neurosci. Methods 2005, 148, 1–18. [Google Scholar]
- Biran, R.; Martin, D.; Tresco, P. Neuronal cell loss accompanies the brain tissue response to chronically implanted silicon microelectrode arrays. Exp. Neurol. 2005, 195, 115–126. [Google Scholar]
- Liu, X.; McCreery, D.B.; Carter, R.R.; Bullara, L.A.; Yuen, T.G.; Agnew, W.F. Stability of the interface between neural tissue and chronically implanted intracortical microelectrodes. IEEE Trans. Rehabil. Eng. 1999, 7, 315–326. [Google Scholar]
- Barrese, J.C.; Rao, N.; Paroo, K.; Triebwasser, C.; Vargas-Irwin, C.; Franquemont, L.; Donoghue, J.P. Failure mode analysis of silicon-based intracortical microelectrode arrays in non-human primates. J. Neural. Eng. 2013, 10, 066014. [Google Scholar] [CrossRef]
- Wellman, S.M.; Eles, J.R.; Ludwig, K.A.; Seymour, J.P.; Michelson, N.J.; McFadden, W.E.; Vazquez, A.L.; Kozai, T.D. A materials roadmap to functional neural interface design. Adv. Funct. Mater. 2018, 28, 1701269. [Google Scholar]
- Rennaker, R.L.; Miller, J.; Tang, H.; Wilson, D.A. Minocycline increases quality and longevity of chronic neural recordings. J. Neural. Eng. 2007, 4, L1. [Google Scholar]
- Harris, J.P. The Glia-Neuronal Response to Cortical Electrodes: Interactions with Substrate Stiffness and Electrophysiology. Ph.D. Thesis, Case Western Reserve University, Cleveland, OH, USA, 2012; pp. 1–190. [Google Scholar]
- Saxena, T.; Karumbaiah, L.; Gaupp, E.A.; Patkar, R.; Patil, K.; Betancur, M.; Stanley, G.B.; Bellamkonda, R.V. The impact of chronic blood–brain barrier breach on intracortical electrode function. Biomaterials 2013, 34, 4703–4713. [Google Scholar]
- Golabchi, A.; Wu, B.; Li, X.; Carlisle, D.L.; Kozai, T.D.; Friedlander, R.M.; Cui, X.T. Melatonin improves quality and longevity of chronic neural recording. Biomaterials 2018, 180, 225–239. [Google Scholar]
- Goss-Varley, M.; Dona, K.R.; McMahon, J.A.; Shoffstall, A.J.; Ereifej, E.S.; Lindner, S.C.; Capadona, J.R. Microelectrode implantation in motor cortex causes fine motor deficit: Implications on potential considerations to Brain Computer Interfacing and Human Augmentation. Sci. Rep. 2017, 7, 15254. [Google Scholar] [CrossRef] [Green Version]
- Ravikumar, M.; Hageman, D.J.; Tomaszewski, W.H.; Chandra, G.M.; Skousen, J.L.; Capadona, J.R. The effect of residual endotoxin contamination on the neuroinflammatory response to sterilized intracortical microelectrodes. J. Mater. Chem. B 2014, 2, 2517–2529. [Google Scholar] [CrossRef] [Green Version]
- Ereifej, E.S.; Smith, C.S.; Meade, S.M.; Chen, K.; Feng, H.; Capadona, J.R. The neuroinflammatory response to nanopatterning parallel grooves into the surface structure of intracortical microelectrodes. Adv. Funct. Mater. 2018, 28, 1704420. [Google Scholar]
- Mahajan, S.; Sharkins, J.A.; Hunter, A.H.; Avishai, A.; Ereifej, E.S. Focused Ion Beam Lithography to Etch Nano-architectures into Microelectrodes. JoVE 2019, 155, e60004. [Google Scholar]
- Bedell, H.W.; Hermann, J.K.; Ravikumar, M.; Lin, S.; Rein, A.; Li, X.; Molinich, E.; Smith, P.D.; Selkirk, S.M.; Miller, R.H.; et al. Targeting CD14 on blood derived cells improves intracortical microelectrode performance. Biomaterials 2018, 163, 163–173. [Google Scholar] [CrossRef]
- Hermann, J.K.; Ravikumar, M.; Shoffstall, A.; Ereifej, E.; Kovach, K.; Chang, J.; Soffer, A.; Wong, C.; Srivastava, V.; Smith, P.; et al. Inhibition of the Cluster of Differentiation 14 Innate Immunity Pathway with IAXO-101 Improves Chronic Microelectrode Performance. J. Neural Eng. 2018, 15, 025002. [Google Scholar]
- Quiroga, R.Q.; Nadasdy, Z.; Ben-Shaul, Y. Unsupervised spike detection and sorting with wavelets and superparamagnetic clustering. Neural Comput. 2004, 16, 1661–1687. [Google Scholar]
- Nguyen, J.K.; Jorfi, M.; Buchanan, K.L.; Park, D.J.; Foster, E.J.; Tyler, D.J.; Rowan, S.J.; Weder, C.; Capadona, J.R. Influence of resveratrol release on the tissue response to mechanically adaptive cortical implants. Acta Biomater. 2016, 29, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Potter, K.A.; Simon, J.S.; Velagapudi, B.; Capadona, J.R. Reduction of autofluorescence at the microelectrode-cortical tissue interface improves antibody detection. J. Neurosci. Methods. 2012, 203, 96–105. [Google Scholar] [CrossRef]
- nfca: Numerical Formal Concept Analysis for Systematic Clustering. 2018. Available online: https://rdrr.io/cran/nFCA/man/nfca.html (accessed on 30 August 2020).
- Carpineto, C.; Romano, G. Concept Data Analysis: Theory and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Choi, V.; Huang, Y.; Lam, V.; Potter, D.; Laubenbacher, R.; Duca, K. Using formal concept analysis for microarray data comparison. J. Bioinform. Comput. Biol. 2008, 6, 65–75. [Google Scholar]
- Ma, J. Contributions to Numerical Formal Concept Analysis, Bayesian Predictive Inference and Sample Size Determination. Ph.D. Thesis, Case Western Reserve University, Cleveland, OH, USA, 2011. [Google Scholar]
- Ma, J.; Sun, J.; Zhang, G.-Q. Numerical Formal Concept Analysis (nFCA): A New Systematic Clustering Technique, with a Network Structure. 2018. Under review. [Google Scholar]
- Buzsáki, G. Large-scale recording of neuronal ensembles. Nat. Neurosci. 2004, 7, 446–451. [Google Scholar]
- McConnell, G.C.; Rees, H.D.; Levey, A.I.; Gutekunst, C.-A.; Gross, R.E.; Bellamkonda, R.V. Implanted neural electrodes cause chronic, local inflammation that is correlated with local neurodegeneration. J. Neural. Eng. 2009, 6, 056003. [Google Scholar] [CrossRef]
- Potter, K.A.; Buck, A.C.; Self, W.K.; Capadona, J.R. Stab injury and device implantation within the brain results in inversely multiphasic neuroinflammatory and neurodegenerative responses. J. Neural. Eng. 2012, 9, 046020. [Google Scholar]
- Kozai, T.D.; Jaquins-Gerstl, A.S.; Vazquez, A.L.; Michael, A.C.; Cui, X.T. Brain tissue responses to neural implants impact signal sensitivity and intervention strategies. ACS Chem. Neurosci. 2015, 6, 48–67. [Google Scholar] [CrossRef] [Green Version]
- Karumbaiah, L.; Saxena, T.; Carlson, D.; Patil, K.; Patkar, R.; Gaupp, E.A.; Betancur, M.; Stanley, G.B.; Carin, L.; Bellamkonda, R.V. Relationship between intracortical electrode design and chronic recording function. Biomaterials 2013, 34, 8061–8074. [Google Scholar] [CrossRef]
- Michelson, N.J.; Vazquez, A.L.; Eles, J.R.; Salatino, J.W.; Purcell, E.K.; Williams, J.J.; Cui, X.T.; Kozai, T.D. Multi-scale, multi-modal analysis uncovers complex relationship at the brain tissue-implant neural interface: New emphasis on the biological interface. J. Neural Eng. 2018, 15, 033001. [Google Scholar]
- Lau, L.W.; Cua, R.; Keough, M.B.; Haylock-Jacobs, S.; Yong, V.W. Pathophysiology of the brain extracellular matrix: A new target for remyelination. Nat. Rev. Neurosci. 2013, 14, 722–729. [Google Scholar]
- Bailey, K.R.; Crawley, J.N. Anxiety-related behaviors in mice. In Frontiers in Neuroscience. Methods of Behavioral Analysis in Neuroscience; Buccafusco, J.J., Ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 77–101. [Google Scholar]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar]
- Metz, G.A.; Whishaw, I.Q. Cortical and subcortical lesions impair skilled walking in the ladder rung walking test: A new task to evaluate fore- and hindlimb stepping, placing, and co-ordination. J. Neurosci. Methods 2002, 115, 169–179. [Google Scholar]
- Metz, G.A.; Whishaw, I.Q. The ladder rung walking task: A scoring system and its practical application. J. Vis. Exp. 2009, 28, e1204. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, H.; Yamamoto, K.; Nozato, S.; Inagaki, T.; Tsuchimochi, H.; Shirai, M.; Yamamoto, R.; Imaizumi, Y.; Hongyo, K.; Yokoyama, S.; et al. Modified forelimb grip strength test detects aging-associated physiological decline in skeletal muscle function in male mice. Sci. Rep. 2017, 7, 42323. [Google Scholar] [CrossRef]
- Adolph, K.E.; Franchak, J.M. The development of motor behavior. Wiley Interdiscip. Rev. Cogn. Sci. 2017, 8, e1430. [Google Scholar]
- Ward, M.P.; Rajdev, P.; Ellison, C.; Irazoqui, P.P. Toward a comparison of microelectrodes for acute and chronic recordings. Brain Res. 2009, 1282, 183–200. [Google Scholar]
- McCreery, D.; Cogan, S.; Kane, S.; Pikov, V. Correlations between histology and neuronal activity recorded by microelectrodes implanted chronically in the cerebral cortex. J. Neural. Eng. 2016, 13, 036012. [Google Scholar]
- Bennett, C.; Mohammed, F.; Álvarez-Ciara, A.; Nguyen, M.A.; Dietrich, W.D.; Rajguru, S.M.; Streit, W.J.; Prasad, A. Neuroinflammation, oxidative stress, and blood-brain barrier (BBB) disruption in acute Utah electrode array implants and the effect of deferoxamine as an iron chelator on acute foreign body response. Biomaterials 2019, 188, 144–159. [Google Scholar]
- Bennett, C.; Samikkannu, M.; Mohammed, F.; Dietrich, W.D.; Rajguru, S.M.; Prasad, A. Blood brain barrier (BBB)-disruption in intracortical silicon microelectrode implants. Biomaterials 2018, 164, 1–10. [Google Scholar]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte–endothelial interactions at the blood–brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar]
- Ereifej, E.S.; Shell, C.; Schofield, J.S.; Charkhkar, H.; Cuberovic, I.; Dorval, A.C.; Graczyk, E.L.; Kozai, T.D.Y.; Otto, K.J.; Tyler, D.J. Neural engineering: The process, applications, and its role in the future of medicine. J. Neural Eng. 2019, 16, 063002. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Outcome Variable: Percent Channels Recording Single Units | Outcome Variable: Units/Channel | ||
|---|---|---|---|
| AIC Value: 1756.784 LME model with random intercept and slope | AIC Value: 619.05 GLM model | ||
| 0–50 µm Histology Distance | logit(y (t)) ~ b × x + a1i + a2i × time, where “i” is the ith subject, b = (b0, b1, …, bq) and ai is the random effects y = % channels recording single units x = (1, time, Right Paw Slip(t), Left Paw Slip(t), Maximum Velocity(t), BBB Permeability, Microglia/Macrophage Activation, Astrocyte Reactivity) | log(y (t)) ~ b × x, where “i” is the ith subject, b = (b0, b1, …, bq) y = units/channel x = (1, Right Paw Slip(t), Maximum Velocity(t), BBB Permeability, Microglia/Macrophage Activation, Astrocyte Reactivity) | |
| 50–100 µm Histology Distance | AIC Value: 1768.454 LME model with random intercept | AIC Value: 1136.184 LME model with random intercept | |
| logit(y (t)) ~ b × x + ai, where “i” is the ith subject, b = (b0, b1, …, bq) and ai is the random effect y = % channels recording single units x = (1, Ladder Time(t), Left Paw Slip(t), Maximum Velocity(t), BBB Permeability, Neuron Density) | log(y (t)) ~ b × x + ai, where “i” is the ith subject, b = (b0, b1, …, bq) and ai is the random effect y = units/channel x = (1, Ladder Time(t), Right Paw Slip(t), Left Paw Slip(t), Maximum Velocity(t), BBB Permeability, Neuron Density) |
| nFCA Relationships | ||||||
|---|---|---|---|---|---|---|
| Variable | Relationship | # Networks | Network Variable | Strength | Direction | |
| Recording Metrics | % Channels Recording Single Units | Positive | 1 | Units/Channel | 0.964 | BI |
| Units/Channel | Positive | 1 | % Channels Recording Single Units | 0.964 | BI | |
| Behavior Metrics | Maximum Velocity (Grid) | N/A | 0 | N/A | N/A | N/A |
| Distance (Grid) | N/A | 0 | N/A | N/A | N/A | |
| Ladder Time | Positive | 1 | Distance (Grid) | 0.608 | UNI | |
| Left Paw Slip (Ladder) | N/A | 0 | N/A | N/A | N/A | |
| Right Paw Slip (Ladder) | N/A | 0 | N/A | N/A | N/A | |
| Grip | Positive | 1 | Astrocyte Reactivity (0-50 µm) | 0.462 | UNI | |
| Positive | Astrocyte Reactivity (50-100 µm) | 0.676 | UNI | |||
| Histology Metrics | Neurons (0–50 µm) | Positive | 2 | % Channels Recording Single Units | 0.628 | UNI |
| Positive | Units/Channel | 0.644 | UNI | |||
| Neurons (50–100 µm) | Positive | 5 | Neurons (0–50 µm) | 0.764 | UNI | |
| Negative | Right Paw Slip (Ladder) | 0.683 | UNI | |||
| Negative | Grip | 0.763 | UNI | |||
| Negative | Microglia/Macrophages Activation (0–50 µm) | 0.501 | UNI | |||
| Negative | Microglia/Macrophages Activation (50–100 µm) | 0.623 | UNI | |||
| Blood–Brain Barrier (BBB) Permeability (0–50 µm) | Positive | 1 | Ladder Time | 0.73 | UNI | |
| Blood–Brain Barrier (BBB) Permeability (50–100 µm) | Positive | 4 | Blood–Brain Barrier (BBB) Permeability (0–50 µm) | 0.969 | UNI | |
| Positive | Left Paw Slip (Ladder) | 0.511 | UNI | |||
| Negative | Maximum Velocity (Grid) | 0.926 | UNI | |||
| Negative | Distance (Grid) | 0.879 | UNI | |||
| Microglia/Macrophages Activation (0–50 µm) | Positive | 3 | Microglia/Macrophages Activation (50–100 µm) | 0.899 | UNI | |
| Positive | Astrocyte Reactivity (0–50 µm) | 0.873 | UNI | |||
| Positive | Astrocyte Reactivity (50–100 µm) | 0.919 | BI | |||
| Microglia/Macrophages Activation (50–100 µm) | Positive | 2 | Maximum Velocity (Grid) | 0.39 | UNI | |
| Negative | Ladder Time | 0.617 | UNI | |||
| Astrocyte Reactivity (0–50 µm) | Positive | 2 | Blood–Brain Barrier (BBB) Permeability (0–50 µm) | 0.41 | UNI | |
| Positive | Blood–Brain Barrier (BBB) Permeability (50–100 µm) | 0.278 | UNI | |||
| Astrocyte Reactivity (50–100 µm) | Positive | 1 | Microglia/Macrophages Activation (0–50 µm) | 0.919 | BI | |
| Outcome Variable: Percent Channels Recording Single Units | Outcome Variable: Units/Channel | ||||||
|---|---|---|---|---|---|---|---|
| Exploratory Variable | Estimate | p-value | Exploratory Variable | Estimate | p-value | ||
| 0–50 µm Histology | Right Paw Slip (Ladder) | −1.955 | 0.146 | Right Paw Slip (Ladder) | −1.390 | 0.015 | |
| Left Paw Slip (Ladder) | −4.348 | 0.124 | Maximum Velocity (Grid) | −3.580 | 0.006 | ||
| Maximum Velocity (Grid) | −11.760 | 0.003 | BBB Permeability | −0.029 | 2.51 × 10−6 | ||
| BBB Permeability | −0.082 | 3.910 × 10−8 | Microglia/Macrophage Activation | −0.205 | 4.58 × 10−6 | ||
| Microglia/Macrophage Activation | −0.550 | 6.310 × 10−6 | Astrocyte Reactivity | 0.520 | 9.57 × 10−7 | ||
| Astrocyte Reactivity | 1.260 | 9.480 × 10−6 | Intercept | 0.413 | 0.510 | ||
| Intercept | 4.724 | 0.008 | |||||
| Time | −0.215 | 0.240 | |||||
| 50–100 µm Histology | Ladder Time | 0.019 | 0.020 | Ladder Time | 0.007 | 0.054 | |
| Ladder_Left_Front_Slips | −0.127 | 0.967 | Right Paw Slip (Ladder) | −0.960 | 0.100 | ||
| Maximum Velocity (Grid) | −12.072 | 0.001 | Left Paw Slip (Ladder) | −0.833 | 0.500 | ||
| BBB Permeability | −0.303 | 0.027 | Maximum Velocity (Grid) | −3.525 | 0.013 | ||
| Neuron Density | 0.026 | 0.245 | BBB Permeability | −0.091 | 0.088 | ||
| Intercept | 3.448 | 0.149 | Neuron Density | 0.006 | 0.528 | ||
| Intercept | 0.446 | 0.661 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ereifej, E.S.; Li, Y.; Goss-Varley, M.; Kim, Y.; Meade, S.M.; Chen, K.; Rayyan, J.; Feng, H.; Dona, K.; McMahon, J.; et al. Investigating the Association between Motor Function, Neuroinflammation, and Recording Metrics in the Performance of Intracortical Microelectrode Implanted in Motor Cortex. Micromachines 2020, 11, 838. https://doi.org/10.3390/mi11090838
Ereifej ES, Li Y, Goss-Varley M, Kim Y, Meade SM, Chen K, Rayyan J, Feng H, Dona K, McMahon J, et al. Investigating the Association between Motor Function, Neuroinflammation, and Recording Metrics in the Performance of Intracortical Microelectrode Implanted in Motor Cortex. Micromachines. 2020; 11(9):838. https://doi.org/10.3390/mi11090838
Chicago/Turabian StyleEreifej, Evon S., Youjun Li, Monika Goss-Varley, Youjoung Kim, Seth M. Meade, Keying Chen, Jacob Rayyan, He Feng, Keith Dona, Justin McMahon, and et al. 2020. "Investigating the Association between Motor Function, Neuroinflammation, and Recording Metrics in the Performance of Intracortical Microelectrode Implanted in Motor Cortex" Micromachines 11, no. 9: 838. https://doi.org/10.3390/mi11090838