Chamazulene-Rich Artemisia arborescens Essential Oils Affect the Cell Growth of Human Melanoma Cells



1
Department of Drug Sciences, University of Catania, Via S. Sofia 64, 95125 Catania, Italy
2
Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, V.le delle Scienze, Parco d’Orleans II, I-90128 Palermo, Italy
3
Centro Interdipartimentale di Ricerca “Riutilizzo Bio-Based Degli Scarti da Matrici Agroalimentari” (RIVIVE), University of Palermo, I-90128 Palermo, Italy
4
Department of Biomedical and Biotechnological Sciences, Section of Physiology, University of Catania, Via S. Sofia, 89, 95123 Catania, Italy
5
Department of Pharmacy, University of Naples Federico II, Via Montesano 49, I-80131 Naples, Italy
*
Author to whom correspondence should be addressed.
Plants 2020, 9(8), 1000; https://doi.org/10.3390/plants9081000
Submission received: 7 July 2020
/
Revised: 30 July 2020
/
Accepted: 1 August 2020
/
Published: 6 August 2020
(This article belongs to the Section Phytochemistry)
Abstract
:Artemisia arborescens is an aromatic shrub whose essential oils are considered a potential source of molecules with industrial and pharmaceutical interest. The chemical profile of A. arborescens essential oils (EOs) was shown to be quite variable and various chemotypes have been identified. In this study, we compared the EOs composition of A. arborescens leaves and flowers collected from four different locations in Sicily. The EOs were assayed for their antiproliferative activity against A375 human malignant melanoma cells, also testing cell viability and cell membrane integrity. The evaluation of DNA fragmentation and caspase-3 activity assay was employed for the detection of apoptosis. The expression of Bcl-2, Bax, cleaved caspase-9, PTEN (Phosphatase and tensin homolog), Hsp70 (Heat Shock Protein 70 kilodaltons) and SOD (superoxide dismutase) proteins was evaluated by Western blot analysis. The levels of ROS and GSH were also analyzed. Results show that EOs presented significant differences in their composition, yield, and cytotoxic activity depending on the collection site. The chamazulene/camphor-rich EOs from plants collected in Acqua Calda (Lipari) resulted particularly active on melanoma cancer cells (IC50 values of 6.7 and 4.5 µg/mL), being able to trigger apoptotic death probably interfering with endogenous defense mechanisms. These oils may be considered as a natural resource of chamazulene, containing this compound up to 63%.
1. Introduction
Artemisia L. (Artemisiinae-Anthemideae-Compositae) comprises more than 500, mostly perennial species [1] and it is considered the largest genus of the tribe Anthemideae. It is a cosmopolitan genus containing wind-pollinated plants with huge ecological plasticity, occurring from high mountains to sea level and from wetlands to arid zones. In Italy, about 20 species are reported and only five are present in Sicilian flora [2]. Besides their several therapeutic applications, Artemisia spp. are used in the preparation of bitter liqueurs (e.g., vermouth), as perfume ingredients and flavoring agents in food, so representing a big potential for industrial and therapeutic use [3]. Due to the large diversity of their chemicals in general (terpenoids, coumarins, flavonoids, caffeoylquinic acids, sterols and acetylenes) and essential oil production, in particular [4,5] a great botanical, industrial and pharmaceutical interest has arisen. Artemisia essential oils (EO), occurring mainly in leaves and flowers, are usually characterized by two-three dominant components at high concentrations (20–70%), with respect to the other compounds present in minor amounts [4]. A significant intraspecific variability regarding EOs composition has been observed among Artemisia taxa; this variability is widely influenced by the geographic region, harvesting season, plant part, growing site, drying conditions, extraction method, but mostly by genotype [4]. Inside each species, some studies have recognized the occurrence of different chemotypes [6].
Artemisia arborescens L. (“great mugwort”, “arborescent mugwort”) is a morphologically variable species with silver to grey-green leaves bearing at flowering time (June–July) small yellow flowers, occurring as a many-branched shrub growing up to one meter in height in various habitats of the Mediterranean region. In fact, it is commonly known as “arboreous absinth”. It is a very bitter herb indigenous in the Middle East listed among traditional medicinal plants, with ethnopharmacological reference dating back to the 16th century: for instance, leaves decoction is used in tea, typically with mint, as tonic and sedative [7]. According to popular folklore, it is used as a remedy for dysfunctions of the gastrointestinal tract and in the treatment of rheumatism, neuralgia, stomach pain, renal colic and lung diseases [8], while the essential oils (wormwood and mugwort) have been used as rodent and mite repellents and for antipyretic, antispasmodic, abortifacient and anti-inflammatory activities. The use of A. arborescens EO is also common in flavoring and fragrance due to its fresh, slightly sweet, minty, woody and earthy scent [9]. Besides, the EO showed in vitro antipyretic, anti-inflammatory, antioxidant, antiangiogenic, antiviral and antimicrobial activities [3,4,6,10,11,12,13,14], with different effects based on the main compounds found in the essential oil. Indeed, the composition of A. arborescens EOs was found to be very variable depending on the environment [5,14]. Basically, the literature describes two chemical profiles for a group of compounds of particular interest such as camphor, chamazulene and thujones: the first one shows high levels of camphor and chamazulene whereas the second one is rich in β-thujone [6]. High thujone type is characterized by deep blue to blue-black color with a characteristic thujone-like odor, minty and woody; high chamazulene type is deep blue-black colored, fruitier, more low-key in impact being sweet, fresh, and slightly minty at first but becoming much mintier after five minutes [15].
Unfortunately, despite the well-established use in traditional medicine of this promising medicinal crop and the wide literature about biological and chemical properties of A. arborescens EO, little information is available about its possible antiproliferative effects.
Based on the above, the aim of the present study was the determination and comparison of essential oils composition of A. arborescens leaves and flowers collected from four different locations in Sicily and the island of Lipari. The plants were analyzed at the same stage of their plant life (full flowering). We also describe the EOs biological activity against A375 human malignant melanoma cells, testing several biochemical parameters, such as cell viability and cell membrane integrity. For the detection of apoptosis, the evaluation of DNA fragmentation and caspase-3 activity assay was employed. The expression of Bcl-2 (B-cell lymphoma 2), Bax (bcl-2-like protein 4), cleaved caspase-9, PTEN (Phosphatase and tensin homolog), Hsp70 (Heat Shock Protein 70 kilodaltons) and SOD (superoxide dismutase) proteins was evaluated by Western blot analysis. The levels of reactive oxygen species (ROS) and GSH (glutathione) were also analyzed. To the authors’ knowledge, this is the first exhaustive study on the anticancer properties of A. arborescens essential oil.
2. Results
2.1. Chemical Analysis
A. arborescens flowers (F1–F4) and leaves (L1–L4) were collected in four different locations in Sicily (Southern Italy). The plants were all in the same phenological stage (full flowering); in fact, previous studies report that the highest content of notable compounds (such as chamazulene) is typically reached in summer (June–July) [13] and the full flowering stage is the more appropriate vegetative stage for harvesting [7]. Collection sites of A. arborescens samples and yields of the essential oils (EOs) obtained are shown in Table 1. Generally, A. arborescens EOs elicited a fresh, slightly sweet, minty, woody, earthy scent.
The GC and GC-MS analyses of our samples resulted in the detection of 78 compounds, making up from 91.7% (F2) to 99.8% (F4) of the total oil. All compounds, divided into eight classes, are listed in Table 2, according to their retention indices Ki on anHP-5MS column for each class.
All EOs were dominated by oxygenated monoterpenes (24.3–71.8%) and hydrocarbons (14.9–63.0%). Particularly, oxygenated monoterpenes were dominant in F1, F2, F3, F4 (42.5, 48.5, 58.3 and 55.6%, respectively) and L3 (71.8%), while hydrocarbons prevailed in L1, L2 and L4 (51.8, 55.6 and 63.0%, respectively). Interestingly, except for the samples collected in Lami (F3 and L3), it seems that A. arborescens flowers tend to accumulate oxygenated monoterpenes while the hydrocarbons can be found more abundantly in the leaves. The oxygenated monoterpene fraction may be considered fundamental in the EO chemical composition, as it gives a substantial contribution towards its aromatic characteristics [9].
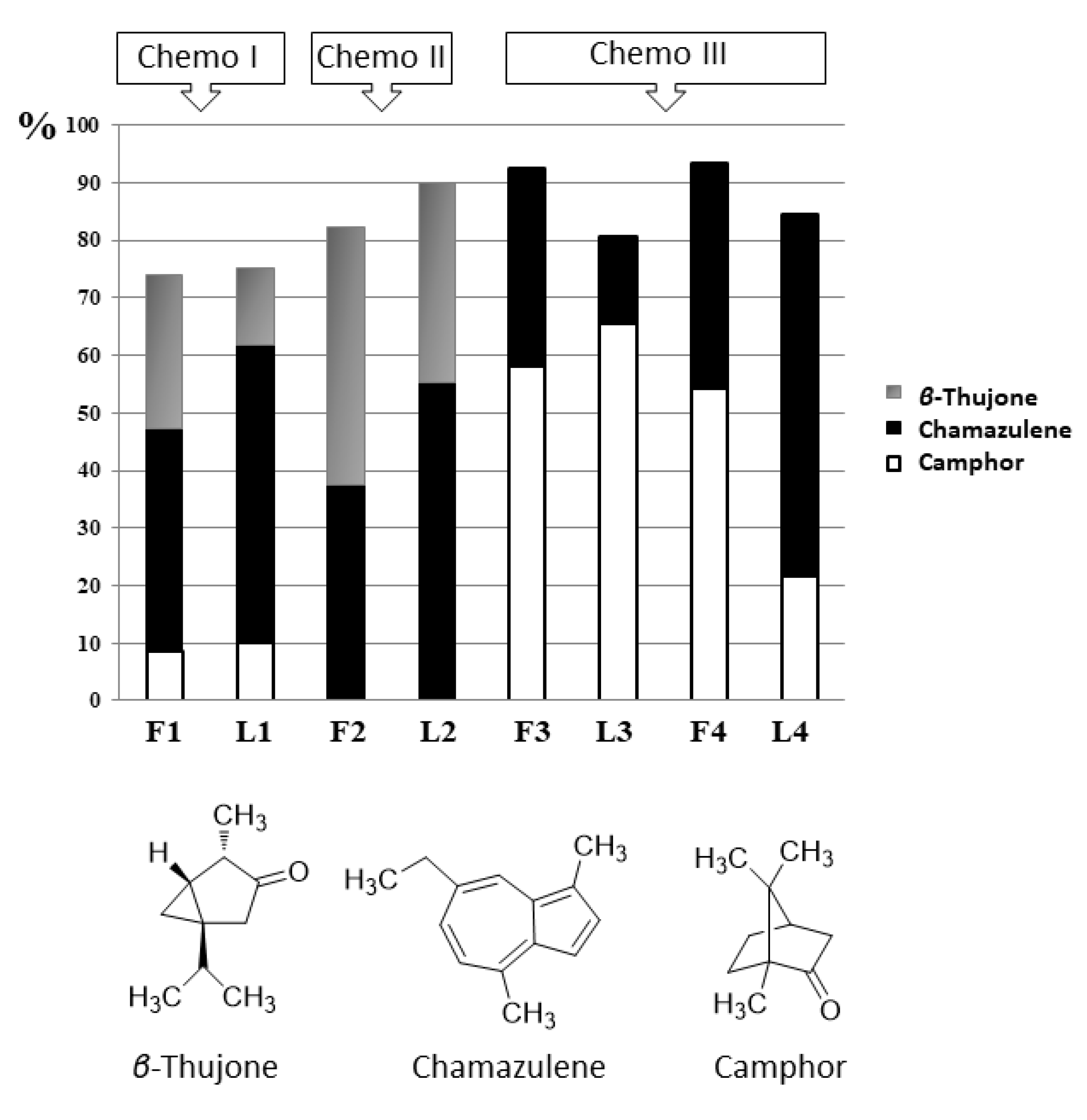
Regarding the main compounds, according to the literature on A. arborescens EOs different chemical profiles can be drawn for three compounds of particular interest such as camphor, chamazulene and thujones: high camphor, chamazulene or β-thujone levels. This is clearly confirmed by the results attained in this research.
The essential oils of A. arborescens from Capo Zafferano (F1 and L1) and Alimena (F2 and L2) were all characterized by a high percentage of β-thujone, completely absent in the oils from plants collected in Lami (F3 and L3) and Acqua Calda (F4 and L4). Particularly, almost half of F2 was constituted by this compound (44.8%), a percentage that decreases in the oil from the leaves (L2, 34.4%). Despite the abundance of the cis isomer of the ketone thujone (β-thujone), the trans form (α-thujone) was only present in small amounts in F1, F2 and L2 (0.6–1.3%). Among these high β-thujone oils, we can clearly see that F1 and L1 were also characterized by high amounts of camphor (8.5% in F1 and 10.0% in L1), not present at all in F2 and L2. Chamazulene was also a very representative compound in these oils: it accounted for 37.4% in F2 and 38.8% in F1, while it was also more abundant in the oils from leaves (51.7% in L1 and 55.4% in L2).
The essential oils of A. arborescens from Lami (F3 and L3) and Acqua Calda (F4 and L4) showed a different profile and were completely β-thujone-free. The oils from flowers were characterized by a remarkably high percentage of camphor, accounting for 58.2% in F3 and 54.2% in F4. Camphor was even more abundant in L3 (65.6%), while in L4 its percentage decreased (21.5%). The oils from flowers were also characterized by high amounts of chamazulene (34.2% in F3 and 39.0% in F4), while, as regards to the leaves, we can clearly see that its level was low in L3 (14.9%) while we could find an exceptionally high percentage of chamazulene (63.0%) in L4.
The other compounds were all present in very low amounts. We can note that the camphor-rich oils (F3, F4 and L3) contained the monoterpenes α-pinene (1.9–3.7%) and camphene (2.6–3.7%), while the β-thujone-rich oils (F1, F2, L1 and L2) were the only to contain sabinene (0.7–1.2%). The relation between sabinene and thujones was determined by previous research, which confirmed that sabinene is the first bicyclic intermediate to arise in the biosynthesis pathway to the epimeric thujones and related compounds, involving the introduction of oxygen in the cyclization step to the thujane skeleton [9].
Other representative compounds were germacrene D, that we could find only in the β-thujone-rich oils (1.5–2.6%) and in the chamazulene-rich L4 (1.1%), and terpinen-4-ol (0.6–2.5%), present in all the oils except F3. According to some authors, this last compound could be in part an artifact generated from sabinene during distillation [16].
2.2. Biological Assays
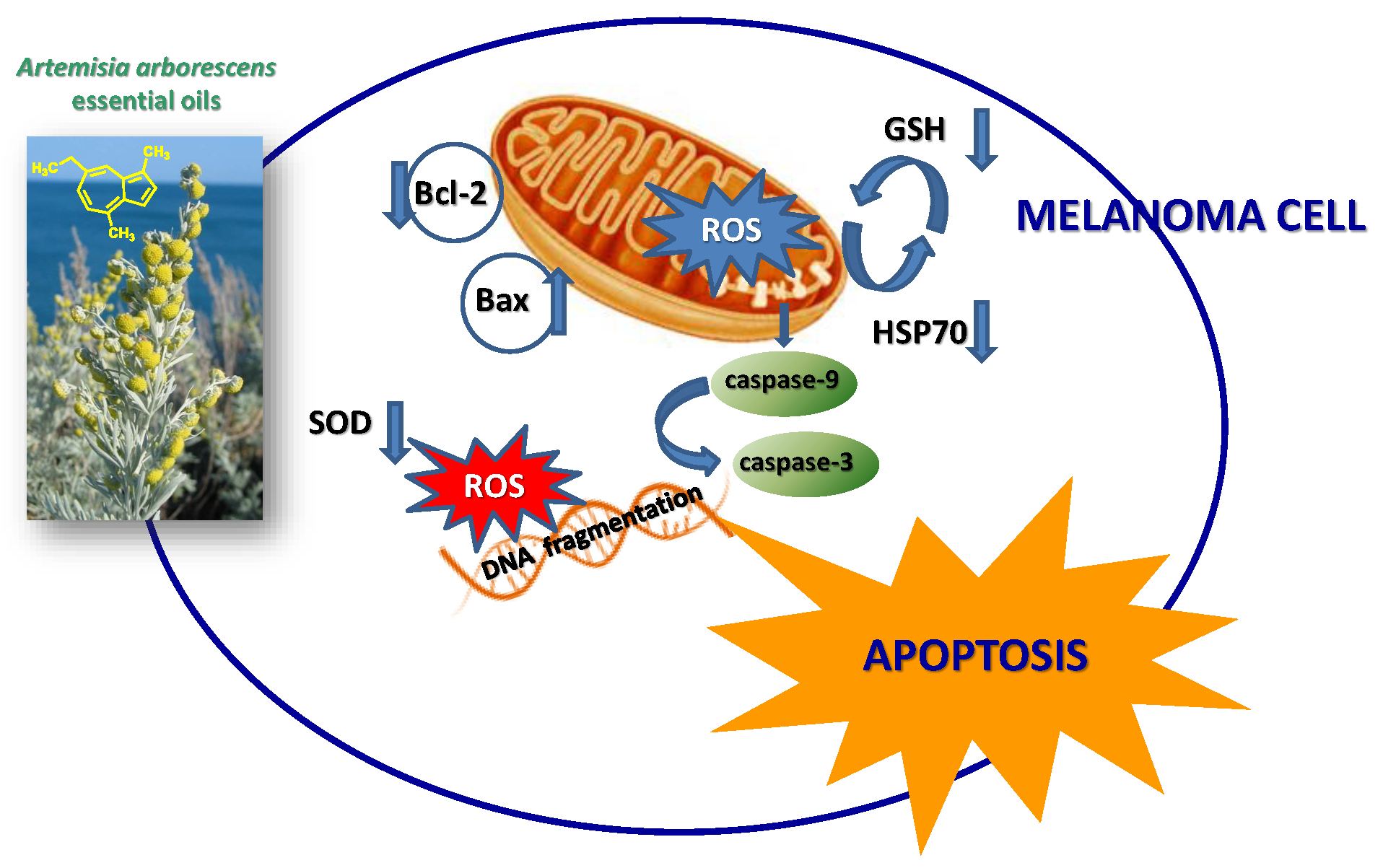
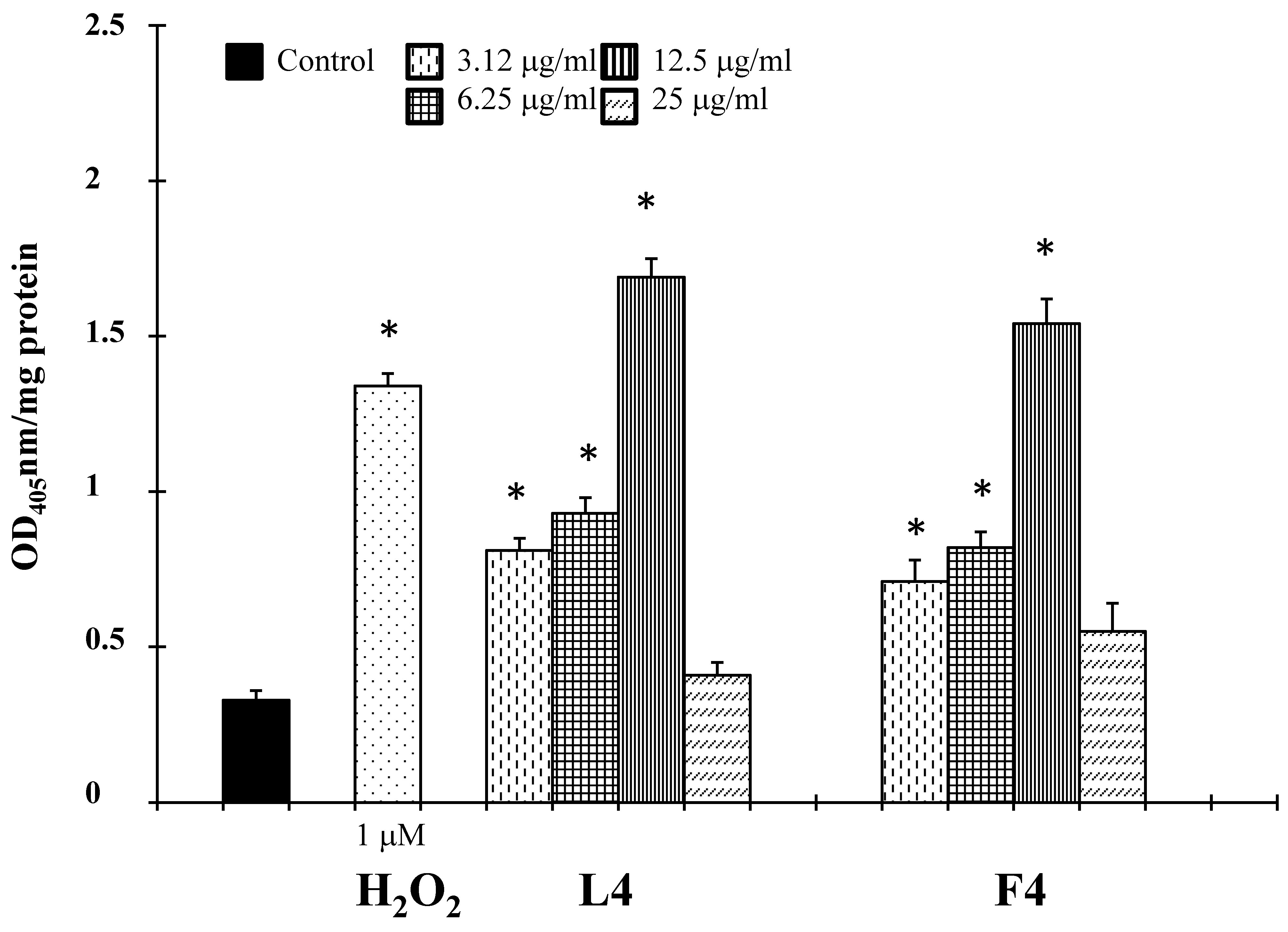
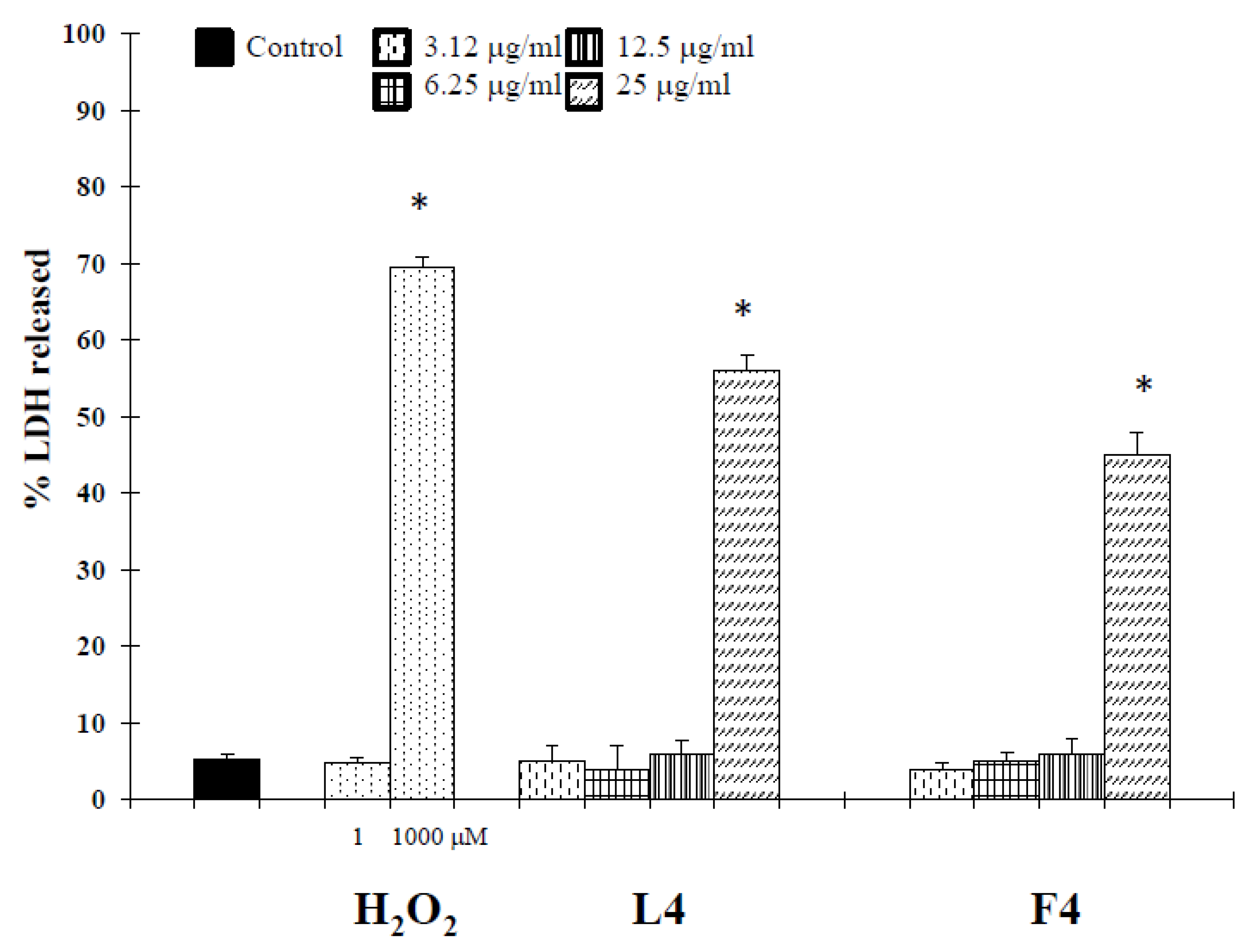
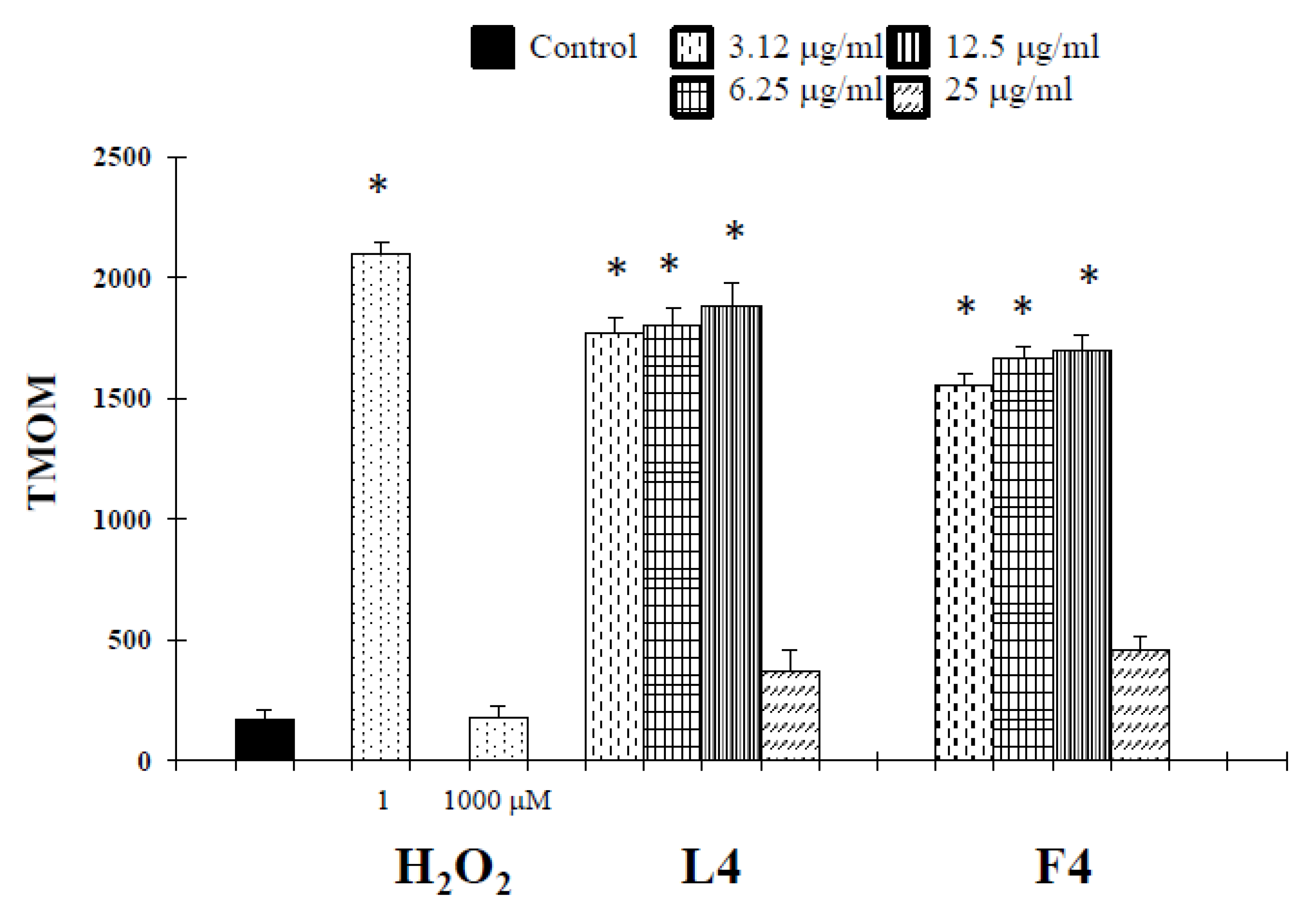
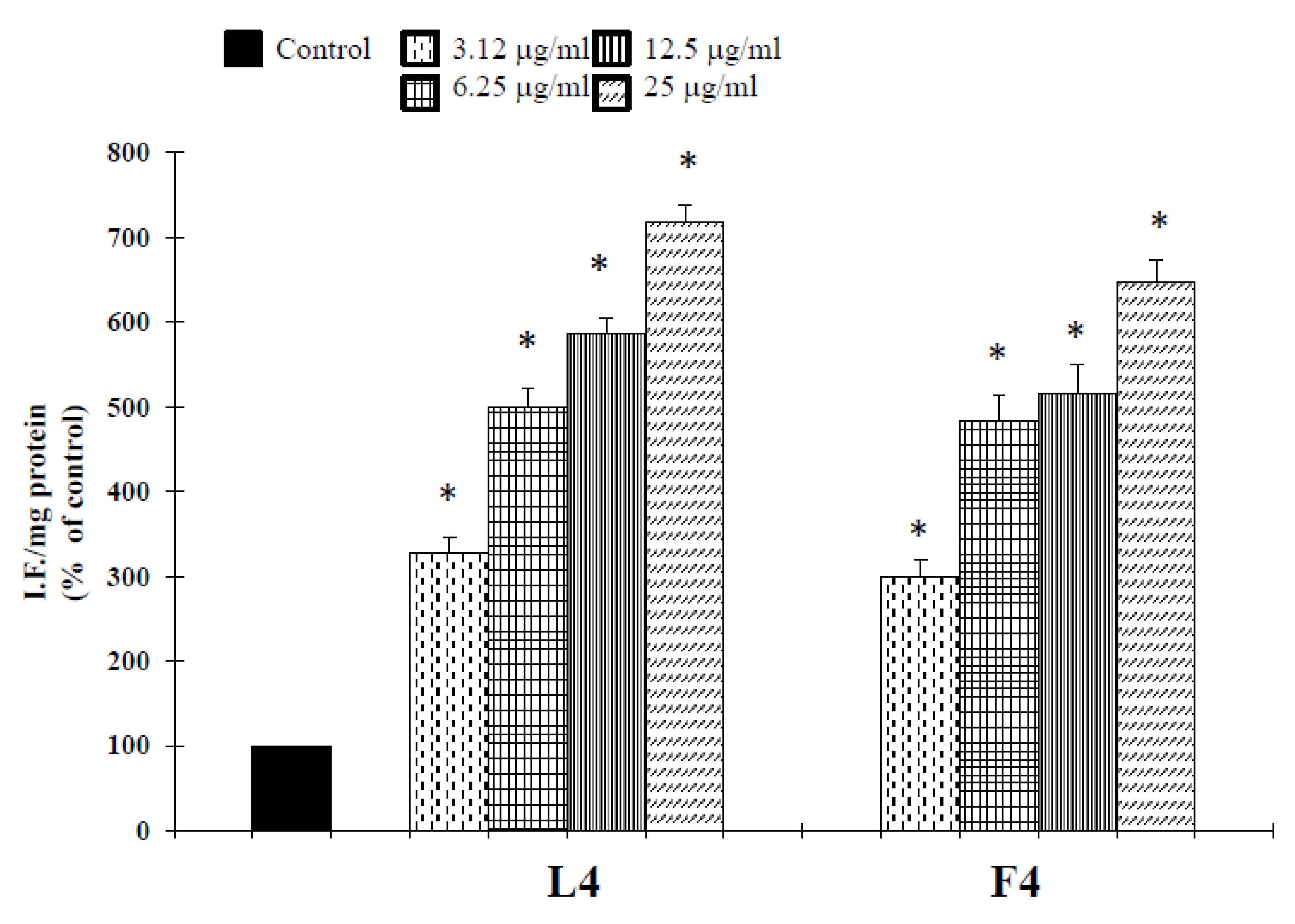
Based on chemical promising results, the essential oils were tested in vitro for their potential anticancer activity on human melanoma cells. Treatment of A375 cells with natural products resulted in a significant reduction in cell viability (Table 3). Interestingly, at 25 µg/mL, concentrations revealed no cytotoxic effect against normal human buccal fibroblast cells (Figure 1). These normal cells were considered a useful model to evaluate the cytotoxic effects of carcinogens and the tumor-specific cytotoxicity of potential antineoplastic agents [17]. Samples F4 and L4 exhibited the major effects, with IC50 values of 6.7 and 4.5 µg/mL, respectively. In the following experiments, we assessed whether cell growth inhibition elicited by samples F4 and L4 was accompanied by an induction of apoptosis in A375 cells. Active caspases cleave several important intracellular proteins, leading to morphological and biochemical changes associated with apoptosis, such as oligonucleosomal fragmentation of chromosomal DNA [18]. Double-stranded DNA is cleaved in fact by DNA fragmentation factor (DFF), that cleaves double-stranded DNA specifically, with a preference for A/T-rich region. Therefore, the next experiments were performed to characterize the role of activation of caspase-3 in cell growth inhibition mediated by tested products. As shown in Figure 2, the activity of this protein was significantly increased in A375 cells treated for 72 h with the essential oils at a concentration of 3.12–12.5 µg/mL, and hydrogen peroxide (H2O2, 1 μM), an apoptotic inductor in cancer cell lines [17]. Conversely, at 25 µg/mL concentration the activity of this protease was reduced. According to the results of caspase-3 activity (Figure 2), no statistically significant increase in LDH (lactate dehydrogenase) release was observed in cancer cells treated with the products at 3.12–12.5 μg/mL concentrations (Figure 3). Alternatively, as the positive control H2O2 (1000 µM), a necrotic inductor in cancer cell lines, when it is used at high concentrations [17], we found a significant LDH release at more high concentrations of 25 μg/mL (Figure 2). Nuclear DNA was analyzed using single-cell gel electrophoresis (SCGE), known as COMET assay, a sensitive method for the visualization of DNA damage measured at the level of individual cells. The characteristic comet-like image can be observed in cells with DNA strand breaks and nucleus fragmentation. The comet consists of a head and a tail representing different DNA structures. The head contains the nuclear core with macromolecules and unfragmented DNA and the tail consists predominantly of single-stranded DNA. The size of the tail shows the level of DNA damage associated with cell damage. The appearance of the tail can be induced by both necrosis and apoptosis [17], but a characteristic for the shape of a comet in apoptotic cells is that most of the DNA moves into the comet’s tail [17]. Because the evaluation of these comets can be problematic, it is mostly performed using special software for the visual comet scoring. Quantification of the COMET data, in our experimental condition, is reported in Figure 4 as tail moment (TMOM). The tail moment is defined as the product of the percentage of DNA in the tail of the comet and TD value, which is obtained calculating the distance between the center of mass of the comet head and the center of mass of the tail. The percentage of DNA in the comet tail is calculated as the rate of the fluorescence intensity in the comet tail relative to the total fluorescence. As reported for the positive control hydrogen peroxide (H2O2, 1 μM), an apoptotic inductor in cancer cell lines [17], the results clearly evidence an increase in TMOM value after treatment with the essential oils at 3.12–12.5 µg/mL concentrations in cancer cells. These findings on genomic DNA damage, seem to suggest that the tested products trigger apoptotic cell death, because data in the literature indicates that only comets with high values of TMOM and TD can be related to apoptosis [17]. Alternatively, in cells exposed to products at 25 µg/mL concentrations for 72 h, as the positive control H2O2 (1000 µM), COMET assay did not evidence typical comet-like structures that occur during apoptosis.
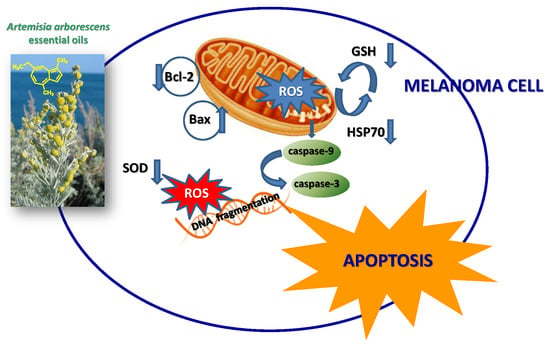
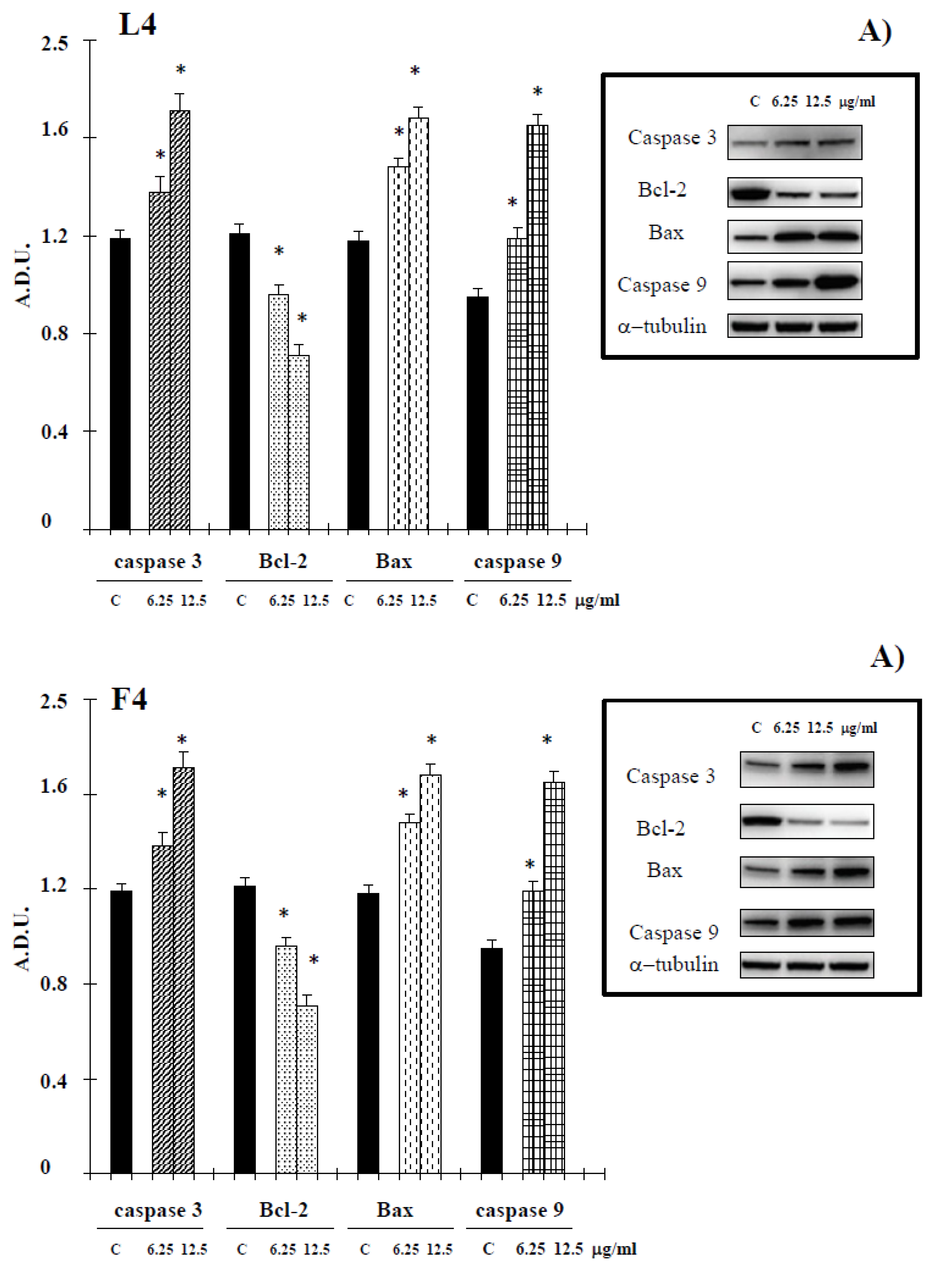
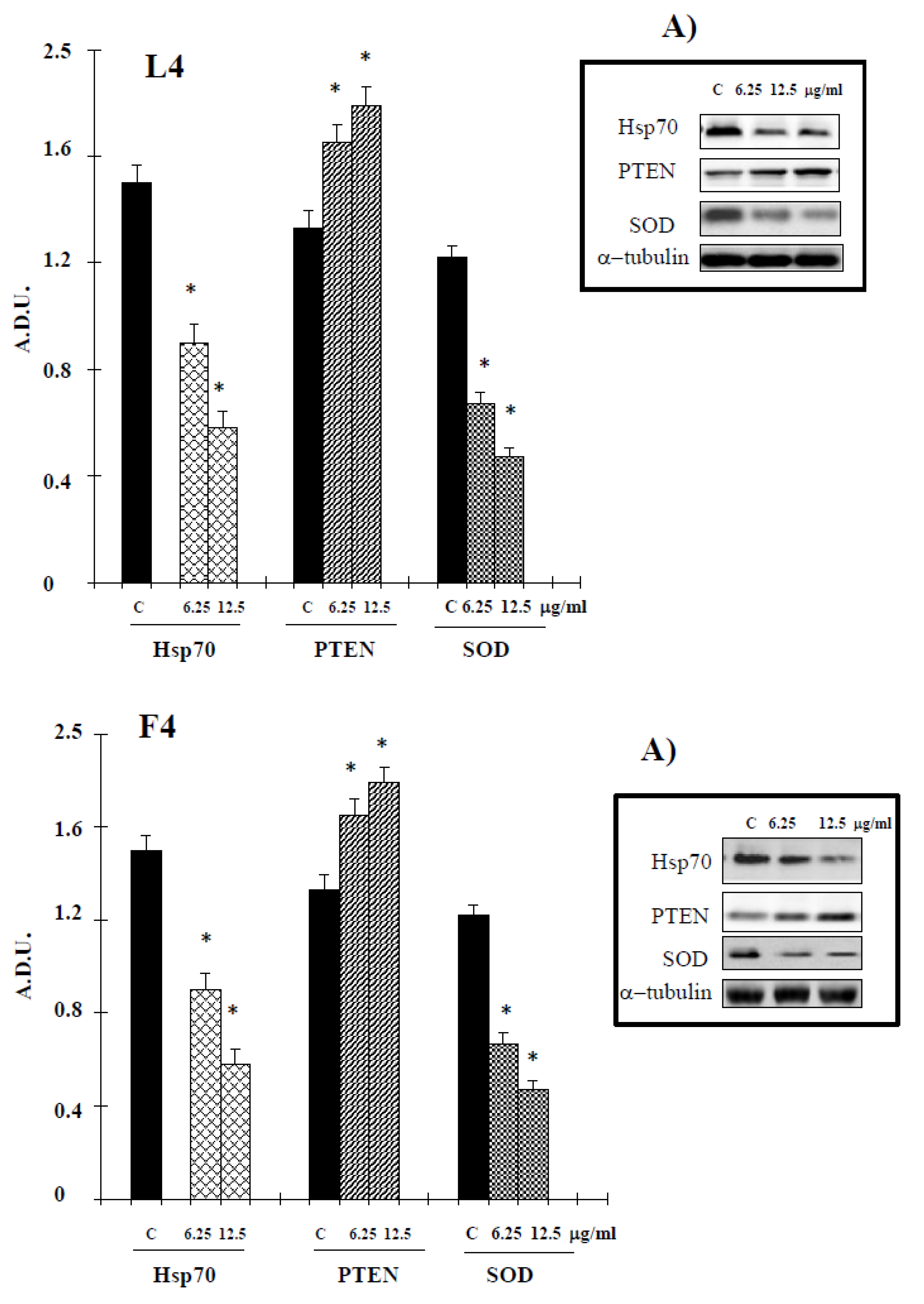
During the intrinsic apoptosis process, apoptosis-related proteins such as Bax, Bcl-2, caspase-9 and caspase-3 are modulated for programmed cell death [18]. To confirm whether the apoptotic effect of essential oils is intrinsic, we conducted Western blotting in A375 cells treated with the tested products for Bcl-2, Bax, caspase 3 and caspase-9. Essential oils suppressed antiapoptotic protein Bcl-2 and increased caspase 3 and caspase-9. Additionally, proapoptotic protein Bax was activated by treatment with tested natural products (Figure 5). In the present study, a possible relationship between Hsp70 expression and essential oil-cell death was investigated. The data, reported in Figure 6, show a reduction of levels of this protein in cancer cells treated with our samples. PTEN plays an important role in melanoma. In PTEN-deficient melanoma cells, the expression of PTEN was able to reduce melanoma progression [19]. The data reported in Figure 6 show an increase in the levels of this tumor suppressor in our experimental condition, supporting the hypothesis that PTEN is one target of F4 and L4 essential oils.
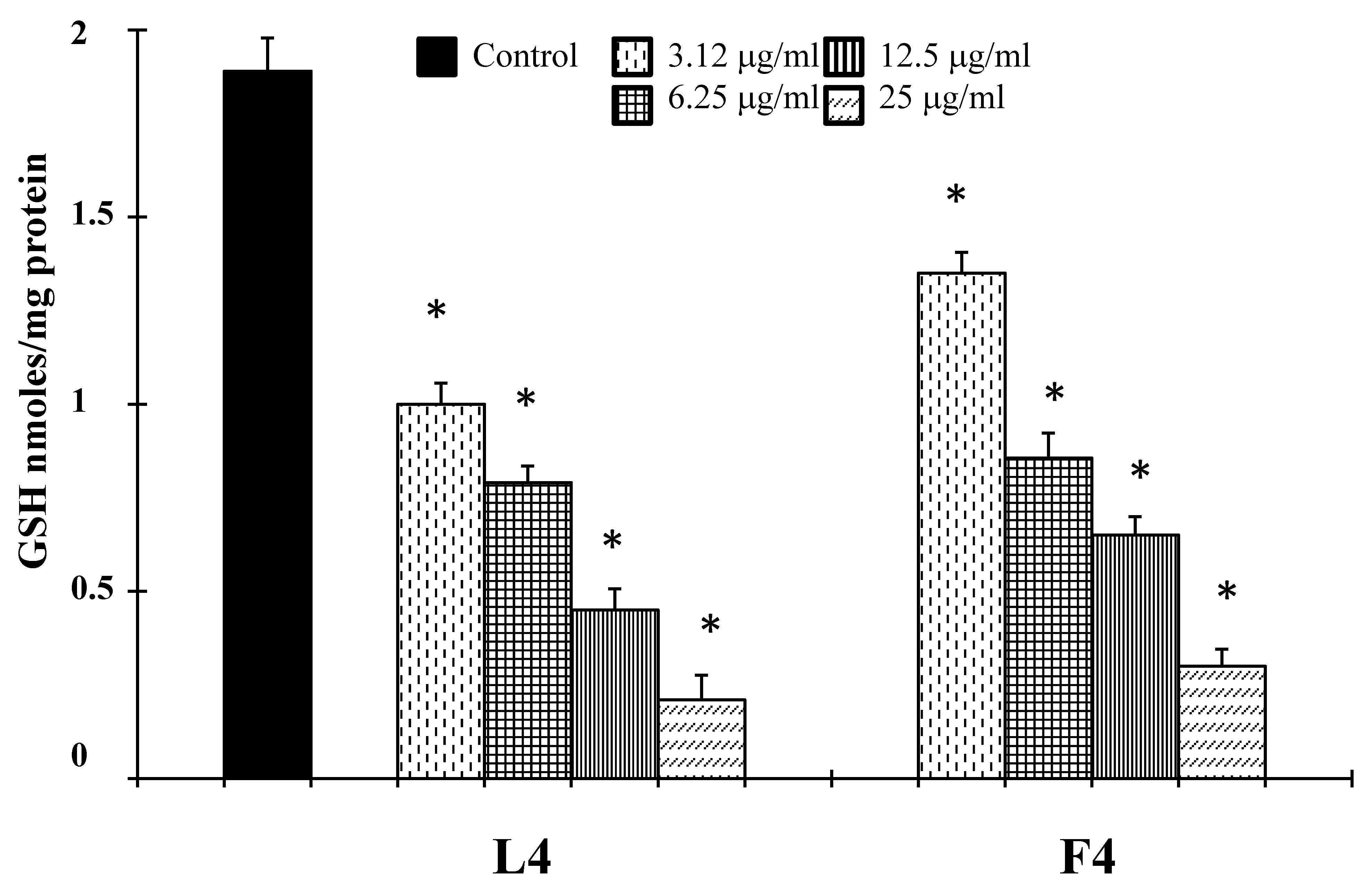
It is well-known that increased generation of ROS, induced by a variety of stimuli, can trigger a series of events resulting in apoptotic occurrence [20]. We, therefore, examined whether essential oil-induced cell death may result from an elevation of ROS. To assess changes in intracellular ROS levels, we employed an oxidation-sensitive fluorescent probe DCFH-DA (2′,7′-dichlorofluorescein diacetate). DCFH-DA can be taken up into cells, and then oxidized by ROS to its fluorescent derivative DCF. The results showed that essential oils were able to increase ROS production in the cells (Figure 7), correlated to a downregulation of the antioxidant enzyme SOD (Figure 8). The GSH content of cancer cells is particularly important, as GSH metabolism, with the aim to promote proliferation and metastasis and alleviate oxidative stress, is accelerated in many types of cancer. High levels of GSH are associated with apoptosis-resistant phenotypes and its depletion is linked to the early stages of cell death initiation [21]. GSH, in fact, serves as a cofactor of several detoxifying enzymes (such as glutathione peroxidase and glutathione transferase) in addition to directly scavenging free radicals. GSH is also involved in recycling other antioxidants by reducing vitamins C and E [22]. We thus evaluated the effect of F4 and L4 on total GSH levels. As different natural compounds [21], these samples, for their active components, induced GSH depletion in melanoma cells (Figure 8).
3. Discussion
Based on the previous published data, the EO chemotypes of Artemisia arborescens L. have been defined based on the detected major compounds (camphor, chamazulene and β-thujone) with the highest percentages. The various combinations of these three chemical compounds define the known chemotypes of this species. So, the essential oils of our samples could be classified into three chemotypes: F1 and L1 belong to the first chemotype (Chemo I) defined by camphor-chamazulene-β-thujone, F2 and L2 belong to the chemotype β-thujone/chamazulene (Chemo II), while F3, F4, L3 and L4 are β-thujone-free and belong to the chemotype chamazulene/camphor (Chemo III, Figure 9) [23,24].
The first chemotype (Chemo I), the most common, was also reported for plants from Tunisia, Sardinia and island of Crete, southern Greece, South Italy and in Turkey [7,23,24], the North-western area of Sicily and in Messina [3,11], Algeria and southern Italy (Sicily: Capo Zafferano and Termini Imerese) [14]. The chemotype β-thujone/chamazulene (Chemo II), free of camphor, was found for A. arborescens essential oil recorded in Algeria, Sardinia and Lebanon [13,24]. The chemotype Chemo III chamazulene/camphor (β-thujone-free) characterizes the studied species in Libya [5] and the USA [15]. Based on the above, the first chemotype (Chemo I) appears to be the most recurrent, and in most studies, camphor, β-thujone and chamazulene have been found to account for more than 50% of A. arborescens EO. Surprisingly, an A. arborescens EO with camphor as the only most abundant component (up to 72.2%) was observed in West Northern of Algeria and was considered as the fourth chemotype type of the species worldwide [12].
In our opinion, the oils F3, F4, L3 and L4 belonging to the Chemo III result particularly worthy of attention. First, these oils are characterized by the absence of thujones. Thujone isomers occur in nature as a mixture of α- and β-thujone, and typically the β-thujone isomer has been identified in A. arborescens oils of different geographical origins as the most abundant, ranging from 0.17% to 68.9%, while α-thujone is often contained in traces or below 1% [9]. α-Thujone is a psychotropic and convulsant substance, so thujones have represented, in the past, a controversial topic of discussion relativeto the psychoactive effects of the liquor absinth (obtained from A. absinthium), initially attributed to α-thujone although its content in the liquor is not sufficient to rouse hallucinations. The clinical picture of absinthism, in fact, has been explained as chronic alcohol misuse and dependence [3,24]. However, the use of thujone-containing plants is limited by food regulations. Thujone, expressed as an isomeric mixture, is banned as a food additive in the US and its presence is regulated in foods and beverages in several countries (the EU limit is of 10 mg/L). The chemical differentiation of A. arborescens belonging to Chemo III, such as our samples F3, F4, L3 and L4, from other thujone-containing plants would be certainly useful for their possible exploitation in the food and pharmaceutical industry. In general, it may be asserted that these A. arborescens oils, could be employed in all food additive and flavoring and fragrance fields, without the toxicological disadvantage of the thujone content.
Another characteristic of the Chemo III oils worthy of mention, particularly for the oils L3 and L4 obtained from leaves, was the high amount of two commercially bioactive compounds used for various industrial purposes: camphor and chamazulene. L3 contains the well-known bicyclic monoterpene ketone camphor in really high percentages (65.6%). This molecule was proposed among natural alternatives to synthetic antimicrobials because it can inhibit the growth of various microorganism strains, and different activities were recorded for this compound: local anesthetic, antiseptic, antiviral, anticancer, insecticidal, skin penetration enhancer and preservative in pharmaceuticals and cosmetics [24]. Even if in some A. arborescens EO the content of this compound could reach 32.2–42.0% [5,7], in Sample L3 the content of camphor was the highest ever found up to now (65.6%), with the exception of the unique A. arborescens EO with camphor as the only most abundant component (up to 72.2%) observed in West Northern of Algeria [12].
L4 contains high amounts of chamazulene. Essential oils that contain chamazulene have important therapeutic applications as this compound is an extremely valuable molecule with unequivocally documented potent antioxidant, anti-inflammatory and antispasmodic activities [7,18]. Chamazulene is produced in considerable amounts in A. arborescens which is at the top of the list of known chamazulene containing oils such as Achillea millefolium or the well-known Matricaria chamomilla [25]: it is generally considered as the degradation product that is formed during EO distillation of proazulenic sesquiterpene lactones naturally occurring in the plant, such as the sesquiterpene lactones artabsin, matricin and the 4-epi-matricin [26].
Different chamazulene-rich A. arborescens oils, characterized by the typical deep blue-black color, have been isolated previously, such as the one from Pacific Northwest (USA) plants described by Pappas and Sheppard-Hanger (2000) [15] or the one from Montenegro [5] with a chamazulene content ranging from 30 to 40%, but it must be noted that a distinct chemotype was reported to yield EOs containing more than 50% of chamazulene. This is the case of A. arborescens EO isolated by Ornano et al. (2013) [13] and Costa et al. (2016) [3] from plants collected in Sardinia and Sicily, where the content of chamazulene reaches 51.5–51.8%. However, in Sample L4 the content of chamazulene was the highest ever found up to now (63.0%). Michelakis et al. (2016) [7] could reach a similar chamazulene concentration (63.2–65.1%) in the EOs from plants collected on the island of Crete through a hydrodistillation performed in basic conditions (pH values 8.3 and 9.9).
The environmental conditions, such as drought and altitude, can strongly influence the composition of plant lipophilic fraction due to their influence on the physiological mechanisms linked to the secondary metabolism. We have already demonstrated that the environmental and pedoclimatic conditions influence the yield, the qualitative chemical composition and the antiproliferative activity of S. officinalis L. essential oils [27] and the chamazulene and bisabolol content in chamomile [19]. It is well-known that the quali/quantitative composition of A. arborescens EO is strongly affected by the stage of plant development, environmental factors and geographic origin [9]. For instance, Said et al. [14] found that monoterpenes were higher and sesquiterpenes lower in the EO coming from the relatively arid (Algeria) than the relatively humid (Italy) environments, and their hypothesis was that the accumulation of monoterpenes could be derived from the oxidation of the sesquiterpene fraction as a response of the plant to the oxidative conditions. Militello et al. [6] observed an increase in the monoterpene fraction as plants pass from vegetative to the flowering stage while in the samples studied by Ornano et al. [13] the highest content of chamazulene is reached in summer (July, as confirmed by Michelakis et al. [7]), while that of β-thujone in spring (May), and generally the EO from plant material collected in July showed the highest amount of sesquiterpene hydrocarbons, while that collected in May exhibited the highest content of oxygenated monoterpenes.
The results here described allow us to draw some conclusions. First, except for camphor-rich samples L3/F3, the amount of chamazulene is always major in the leaves with respect to the flowers. Besides, the fact that L3, F3, L4 and F4 belong to the same chemotype and have been collected in the same phenological state can suggest the importance of the growing site. In fact, these plants have been collected all on the island of Lipari but in two different environments: L3/F3 in Lami, in a hilly site, while L4/F4 in Acqua Calda, nearby the sea. This last condition seems to promote the production of chamazulene, as it is suggested also by the observation that all the chamazulene-rich oils have been collected by plants growing in the same condition.
So, this study seems to confirm that, to obtain an Artemisia arborescens chamazulene-rich oil, this should be isolated from leaves of Chemo III plants growing nearby the sea in the full flowering stage, in the summer.
Melanoma is one of the fastest-growing cancers worldwide and is characterized by high mortality. Nowadays, there are many treatment strategies, but generally, the cells do not respond adequately to most chemotherapeutic agents and manifest chemoresistance [28]. In this context, essential oils represent an excellent source of active molecules for the prevention and treatment of this cancer. So, A. arborescens EOs were first tested in vitro for their potential cell growth inhibitory effect on the A375 cell line. We demonstrated that all essential oils selectively affected the survival of human melanoma cancer cells (Table 3).These results are in accordance with previous literature data which show an interesting cytotoxic activity of A. arborescens essential oil against the human cell lines T98G (glioblastoma), MDA-MB 435S (breast adenocarcinoma), A375 (malignant melanoma) and HCT116 (colon carcinoma) [13]. Samples F4 and L4 exhibited the major effects, with IC50 values of 6.7 and 4.5 µg/mL, respectively. No information on the cytotoxicity of chamazulene was found in the literature while camphor has been reported to be active against several cancer cell lines, including human skin fibroblasts (WS1), human lung cancer (A549), human keratinocytes (HaCaT) and colon adenocarcinoma (DLD-1) [29]; however, even if the compounds present in the highest percentage in the oils are likely majorly responsible of their observed biological effects, it is possible that the activity of the main components is also modulated by other molecules present in the oil which may act synergistically, as previously suggested [30]. For instance, α-pinene present in the essential oil F4 was found to be active against hepatoma carcinoma cells [31], and camphene induces programmed cell death (also known as apoptosis) by the mitochondrial pathway in melanoma cells [32].
Programmed cell death is modulated by pro- and antiapoptotic signals that regulate mechanisms of induction or suppression of cell signaling cascades [18]. The evasion of the apoptotic pathway is important to tumor formation and the not controlled the cell growth of melanoma cells. So, the next experiments were performed to determine whether the inhibitory effects of essential oils F4 an L4on cell viability are the result of apoptotic cell death in the melanoma cell line. Interestingly, essential oils F4 and L4 were able to trigger apoptotic death in melanoma cancer cells. In fact, an increase of the caspase-3 enzyme activity (Figure 4) and DNA damage(comet assay, Figure 5), not correlated to LDH release (Figure 3), a marker of membrane breakdown, were found in A375 cells treated with these natural products at lower concentrations (6.25 and 12.5 μg/mL).
Hsp70 may participate in oncogenesis and in resistance to chemotherapy [33,34]. The expression of this protein is particularly high in many tumors including melanoma, where it is involved in the different stages of carcinogenesis. In addition, Hsp70 may be associated with drug-resistant melanoma [34]. Elevated Hsp70 levels arrest apoptosis at different stages. Hsp70 has been shown to affect some transcription factors involved in the expression of the Bcl-2 family proteins [33,34,35], that are involved in survival and apoptosis [36]. On the basis of these observations, in this study, a possible relationship between Hsp70 expression and cell death elicited by essential oils (F4 and L4) in melanoma cells was investigated. As supposed, these natural products induced the inhibition of Hsp70 expression (Figure 7), correlated with a significant increase of caspase-3 enzyme activity (Figure 4) and damage at the DNA level (Figure 5), in conjunction with an increase of Bax, caspase-3 and caspase-9 protein expression and the more pronounced decrease in Bcl-2 (Figure 6). On the other hand, it has been reported that Hsp70 may inhibit apoptosis downstream of the release of cytochrome c and upstream of the activation of caspase-3, and directly bind to apoptosis protease-activating factor-1 (Apaf-1), thereby preventing the recruitment of procaspase-9 to the apoptosome [33,34]. In addition, our data (Figure 7) permit us to hypothesize that PTEN is one target of F4 and L4 essential oils.
Tumor cells have a higher level of endogenous ROS than normal cells. The higher levels of endogenous ROS in cancer cells increase their susceptibility to oxidative stress-induced cell death [21]. Hsp70 has been reported to increase tumor cell persistence via inhibition of ROS-mediated apoptosis [37]. It is well-known, in fact, that endogenous antioxidant depletion or ROS addition can induce programmed cell death in many cells [38]. The excess of ROS activates cytoplasmic protein kinase and MAPK cascades and induces programmed cell death in different cells. Since mitochondria are the major source of ROS due to their generation during mitochondria respiration, damage at the level of this organelle also causes ROS to increase. The inhibition of mitochondrial respiratory results in overproduction of ROS, which would act as mediators of cell death signaling pathway. On the other hand, caspase activation also causes mitochondrial damage, and results in the loss of the membrane potential and cytochrome c release, thus inducing apoptosis [39]. Consistent with these literature data, we found that samples F4 and L4, in addition to the reduction of Hsp70 expression (Figure 7), affected endogenous antioxidant defense inhibiting the expression of enzyme superoxide dismutase (SOD, Figure 7), and together they were able to increase ROS generation (Figure 8) and to reduce GSH levels (Figure 9). On the other hand, natural molecules can also start the accumulation of intracellular ROS in cancer cells by the downregulation of different antioxidant enzymes [40] and inducing GSH depletion [21].
4. Materials and Methods
4.1. Plant Materials
The flowers (F1–F4) and leaves (L1–L4) of A. arborescens were randomly collected from different plants (8–10 individuals), in June 2018, in four different locations, as reported in Table 1. Plant materials were identified by Emanuele Schimmenti and the vouchers are deposited at the Herbarium Mediterraneum Panormitanum (PAL), University of Palermo.
4.2. Isolation of Essential Oil
Air-dried aerial parts of all the studied populations were subjected to oil extraction after grinding to a fine powder as previously reported [41]. The essential oils were obtained with different yields (1.32 ± 0.03–0.20 ± 0.02%) on a dry mass (see Table 1), and were blue colored by the presence of chamazulene, with a pleasant and typical smell.
4.3. GC and GC-MS Analysis and Identification of Compounds
GC and GC-MS analysis of the essential oils was carried out as described before [41].
4.4. Identification of Compounds
Most constituents were identified by gas chromatography by comparison of their retention indices (Ki) with those of the literature or with those of authentic compounds available in our laboratories, as described before [41].
4.5. Cell Culture and Treatments
The A375 cell line was obtained from American Type Culture Collection (Rockville, MD, USA) and grown as described in [27]. Normal human buccal fibroblast cells, kindly donated by Institute IGB, CNR (Naples, Italy), were grown in the same way. The cells were plated as described by Russo et al. [27]. In the MTT assay, cancer cells were plated at 6 × 103 cells per well in a 96-well flat-bottomed 200 μL microplate, and at 2 × 104 cells per well for normal human buccal fibroblast cells, in a 96-well flat-bottomed 200 μL microplate. In other tests, cells were plated at 8 × 105 cells (2 mL) per 35 mm culture dish. The cell treatment was carried out as described previously [27].
4.6. MTT Bioassay
MTT assay was performed as described previously [17].
4.7. Lactate Dehydrogenase (LDH) Release
LDH activity was spectrophotometrically measured in the culture medium and in the cellular lysates as previously reported [17]. Hydrogen peroxide (H2O2, 30% w/w, Sigma Aldrich Co., St. Louis, MS, USA) was used as a positive control.
4.8. Activity of Caspase-3
The activity of caspase-3 was determined as previously described [17]. Hydrogen peroxide (H2O2, 30% w/w, Sigma Aldrich Co., St. Louis, MS, USA) was used as a positive control. The total protein content, used to reflect cell number and measured according to Bradford (1976) [42], was evaluated for each sample, and the results are reported as OD405 nm/mg protein and compared to relative control.
4.9. DNA Analysis by COMET Assay
The presence of DNA fragmentation was examined by single-cell gel electrophoresis (COMET assay), as previously reported [17]. Hydrogen peroxide (H2O2, 30% w/w, Sigma Aldrich Co, St. Louis, MS, USA), an apoptotic inductor in cancer cell lines, was used as a positive control.
4.10. Western Blot Analysis
The expression of Bcl-2, Bax, cleaved caspase-9, PTEN, Hsp70 and SOD proteins was evaluated by Western blot analysis, as previously described [17].
4.11. Reactive Oxygen Species Assay
Reactive oxygen species (ROS) determination was performed by using a fluorescent probe 2′,7′-dichlorofluorescein diacetate (DCFH-DA), as previously described [17]. The total protein content, measured according to Bradford (1976) [42] was evaluated for each sample, and the results are reported as fluorescence intensity/mg protein and compared to relative control.
4.12. Measurement of GSH levels
Glutathione (GSH) levels were measured as described previously [43].
4.13. Statistical Analysis
Representative data from three independent experiments, performed in quadruplicate, are shown and quantified, and represented as mean ± SD. Results were analyzed using one-way ANOVA followed by Dunnett’s post-hoc test for multiple comparisons with control. All statistical analyses were performed using the statistical software package SYSTAT, version 9 (Systat Inc., Evanston, IL, USA).
5. Conclusions
From these results, we can conclude that the environment significantly affects the metabolite concentrations and the biological properties of EOs obtained from the same A. arborescens chemotype but collected in different places. The best antiproliferative activity against the human melanoma cell line A375 was displayed by oils belonging to the chamazulene/camphor chemotype (Chemo III) collected in Acqua Calda (Lipari). The EOs exhibited cell growth inhibitory effect against cancer cells through the induction of apoptosis probably interfering with endogenous defense mechanisms. The obtained results demonstrated a possible application to human melanoma treatment, even if further studies are needed to better evaluate the mechanisms of action of the observed activity. Interestingly, the A. arborescens essential oils shown in this study may be considered as a naturally renewable resource of chamazulene and camphor. Being completely thujone-free, they could be possibly employed in the food and pharmaceutical industry without the toxicological disadvantage of the thujone content.
Author Contributions
Conceptualization, A.R., D.R. and M.B.; methodology, A.R., D.R., V.C. and R.A.; formal analysis, A.R., D.R., V.C. and R.A.; writing—original draft preparation, A.R. and D.R.; writing—review and editing, A.R., D.R. and M.B; funding acquisition, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from MIUR-ITALY PRIN 2017 (Project N. 2017A95NCJ).
Conflicts of Interest
The authors declare no financial or other conflict of interest.
References
- Vallès, J.; Garnatje, T. Artemisia and its allies: Genome organization and evolution and their biosystematic, taxonomic and phylogenetic implications in the Artemisiinae and related subtribes (Asteraceae, Anthemideae). Plant Genome Biodivers. Evol. 2005, 1, 255–285. [Google Scholar]
- Tutin, T.G.; Persson, K.; Gutermann, W. Compositae: Artemisia. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: London, UK, 1976; pp. 178–186. [Google Scholar]
- Costa, R.; Ragusa, S.; Russo, M.; Certo, G.; Franchina, F.A.; Zanotto, A.; Grasso, E.; Mondello, L.; Germanò, M.P. Phytochemical screening of Artemisia arborescens L. by means of advanced chromatographic techniques for identification of health-promoting compounds. J. Pharm. Biomed. Anal. 2016, 117, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janaćkovića, P.; Rajčevića, N.; Gavrilovića, M.; Novakovića, J.; Giwelib, A.; Steševićc, D.; Marina, P.D. Essential oil composition of five Artemisia (Compositae) species in regards to chemophenetics. Biochem. Syst. Ecol. 2019, 87, 103960–103968. [Google Scholar] [CrossRef]
- Militello, M.; Carrubba, A.; Blázquez, M.A. Artemisia arborescens L.: Essential oil composition and effects of plant growth stage in some genotypes from Sicily. J. Essent. Oil Res. 2012, 24, 229–235. [Google Scholar] [CrossRef]
- Michelakis, E.C.; Evergetis, E.; Koulocheri, S.D.; Haroutounian, S.A. Exploitation of Artemisia arborescens as a renewable source of chamazulene: Seasonal variation and distillation conditions. Nat. Prod. Commun. 2016, 10, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Ballero, M.; Poli, F.; Sacchetti, G.; Loi, M.C. Ethnobotanical research on the territory of Flumini maggiore (southwestern Sardinia). Fitoterapia 2001, 72, 788–801. [Google Scholar] [CrossRef]
- Lo Presti, M.; Crupi, M.L.; Zellner, B.d’A.; Dugo, G.; Mondello, L.; Dugo, P.; Ragusa, S. Characterization of Artemisia arborescens L. (Asteraceae) leaf-derived essential oil from Southern Italy. J. Essent. Oil Res. 2007, 19, 218–224. [Google Scholar] [CrossRef]
- Saddi, M.; Sanna, A.; Cottiglia, F.; Chisu, L.; Casu, L.; Bonsignore, L.; De Logu, A. Antiherpevirus activity of Artemisia arborescens essential oil and inhibition of lateral diffusion in Vero cells. Ann. Clin. Microbiol. Antimicrob. 2007, 26, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Militello, M.; Settanni, L.; Aleo, A.; Mammina, C.; Moschetti, G.; Giammanco, G.M.; Blàzquez, M.A.; Carrubba, A. Chemical composition and antibacterial potential of Artemisia arborescens L. essential oil. Curr. Microbiol. 2011, 62, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Younes, K.; Merghache, S.; Djabou, N.; Merghache, D.; Muselli, A.; Tabti, B.; Costa, J. Chemical composition, antibacterial and antioxidant activities of a new essential oil chemotype of Algerian Artemisia arborescens L. Afr. J. Pharm. Pharm. 2012, 6, 2912–2921. [Google Scholar] [CrossRef] [Green Version]
- Ornano, L.; Venditti, A.; Ballero, M.; Sanna, C.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Papa, F.; Vittori, S.; Maggi, F.; et al. Chemopreventive and antioxidant activity of the chamazulene-rich essential oil obtained from Artemisia arborescens L. growing on the isle of La Maddalena, Sardinia, Italy. Chem. Biodivers. 2013, 10, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Said, M.E.-A.; Militello, M.; Saia, S.; Settanni, L.; Aleo, A.; Mammina, C.; Bombarda, I.; Vanloot, P.; Roussel, C.; Dupuy, N. Artemisia arborescens essential oil composition, enantiomeric distribution, and antimicrobial activity from different wild populations from the Mediterranean area. Chem. Biodivers. 2016, 13, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Pappas, R.; Sheppard-Hanger, S. Artemisia arborescens essential oil of the Pacific Northwest: A high chamazulene, low thujone essential oil with potential skin-care applications. Aromather. J. 2000, 10, 30–33. [Google Scholar]
- Sacco, T.; Frattini, C.; Bicchi, C. Constituents of essential oil of Artemisia arborescens. Planta Med. 1983, 47, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Cardile, V.; Avola, R.; Graziano, A.C.E.; Piovano, M.; Russo, A. Cytotoxicity of demalonylthyrsiflorin A, a semisynthetic labdane-derived diterpenoid, to melanoma cells. Toxicol. Vitr. 2018, 47, 274–280. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 26, 87. [Google Scholar] [CrossRef] [Green Version]
- Aguissa-Touré, A.H.; Li, G. Genetic alterations of PTEN in human melanoma. Cell. Mol. Life Sci. 2012, 69, 1475–1491. [Google Scholar] [CrossRef]
- Nogueira, V.; Hay, N. Molecular pathways: Reactive oxygen species homeostasis in cancer cells and implications for cancer therapy. Clin. Cancer Res. 2013, 19, 4309–4314. [Google Scholar] [CrossRef] [Green Version]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K. Inhibition of cancer antioxidant defense by natural compounds. Oncotarget 2017, 8, 15996–16016. [Google Scholar] [CrossRef] [Green Version]
- Lam, Y.; Ng, T.B.; Yao, R.M.; Shi, J.; Xu, K.; Wing Sze, S.C.; Zhang, K.Y. Evaluation of chemical constituents and important mechanism of pharmacological biology in Dendrobium Plants. Evid Based Complement. Altern. Med. 2015, 841752, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marongiu, B.; Piras, A.; Porcedda, S. Comparative analysis of the oil and supercritical CO2 extract of Artemisia arborescens L. and Helichrysum splendidum (Thunb.). Less. Nat. Prod. Res. 2010, 20, 421–428. [Google Scholar] [CrossRef]
- Riahi, L.; Chograni, H.; Masmoudi, A.S.; Cherif, A. Genetic resources of Tunisian Artemisia arborescens L. (Asteraceae), pattern of volatile metabolites concentration and bioactivity and implication for conservation. Biochem. Syst. Ecol. 2019, 87, 103952–103963. [Google Scholar] [CrossRef]
- Formisano, C.; Delfine, S.; Oliviero, F.; Tenore, G.C.; Rigano, D.; Senatore, F. Correlation among environmental factors, chemical composition and antioxidative properties of essential oil and extracts of chamomile (Matricaria chamomilla L.) collected in Molise (South-central Italy). Ind. Crop. Prod. 2015, 63, 256–263. [Google Scholar] [CrossRef]
- Appendino, G.; Gariboldi, P. The stereochemistry of matricin and 4-epimatricin, proazulene sesquiterpene lactones from Artemisia arborescens. Phytochemistry 1982, 21, 2555–2557. [Google Scholar] [CrossRef]
- Russo, A.; Formisano, C.; Rigano, D.; Senatore, F.; Delfine, S.; Cardile, V.; Rosselli, S.; Bruno, M. Chemical composition and anticancer activity of essential oils of Mediterranean sage (Salvia officinalis L.) grown in different environmental conditions. Food Chem. Toxicol. 2013, 55, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabro, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA A Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cote, H.; Boucher, M.A.; Pichette, A.; Legault, J. Anti-Inflammatory, antioxidant, antibiotic, and cytotoxic activities of Tanacetum vulgare L. essential oil and its constituents. Medicines 2017, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Formisano, C.; Rigano, D.; Cardile, V.; Arnold, N.A.; Senatore, F. Comparative phytochemical profile and antiproliferative activity on human melanoma cells of essential oils of three Lebanese Salvia species. Ind. Crops Prod. 2016, 83, 492–499. [Google Scholar] [CrossRef]
- Chen, W.Q.; Xu, B.; Mao, J.W.; Wei, F.X.; Li, M.; Liu, T.; Jin, X.B.; Zhang, L.R. Inhibitory effects of α-pinene on hepatoma carcinoma cell proliferation. Asian Pac. J. Cancer Prev. 2014, 7, 3293–3297. [Google Scholar] [CrossRef] [Green Version]
- Girola, N.; Figueiredo, C.R.; Farias, C.F.; Azevedo, R.A.; Ferreira, A.K.; Teixeira, S.F.; Capello, T.M.; Martins, E.G.; Matsuo, A.L.; Travassos, L.R.; et al. Camphene isolated from essential oil of Piper cernuum (Piperaceae) induces intrinsic apoptosis in melanoma cells and displays antitumor activity in vivo. Biochem. Biophys. Res. Commun. 2015, 467, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Goloudina, A.R.; Demidov, O.N.; Garrido, C. Inhibition of HSP70: A challenging anti-cancer strategy. Cancer Lett. 2012, 325, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Budina-Kolomets, A.; Webster, M.R.; Leu, J.I.; Jennis, M.C.; Krepler, C.; Guerrini, A.; Kossenkov, A.V.; Xu, W.; Karakousis, G.C.; Schuchter, L.M.; et al. HSP70 Inhibition Limits FAK-Dependent Invasion and Enhances the Response to Melanoma Treatment with BRAF Inhibitors. Cancer Res. 2016, 76, 2720–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yu, G.; Shen, Y. The naturally occurring xanthone α-mangostin induces ROS mediated cytotoxicity in non-small scale lung cancer cells. Saudi J. Biol. Sci. 2018, 25, 1090–1095. [Google Scholar] [CrossRef]
- Ikwegbue, P.C.; Masamba, P.; Oyinloye, B.E.; Kappo, A.P. Roles of heat shock proteins in apoptosis, oxidative Stress, human inflammatory diseases, and cancer. Pharmaceuticals 2018, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Montero, A.J.; Jassem, J. Cellular redox pathways as a therapeutic target in the treatment of cancer. Drugs 2011, 71, 1385–1396. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and cancer: Have we moved forward? Biochem. J. 2007, 401, 1–11. [Google Scholar] [CrossRef]
- Kim, B.; Srivastava, S.K.; Kim, S.H. Caspase-9 as a therapeutic target for treating cancer. Expert Opin. Targets 2017, 19, 113–127. [Google Scholar] [CrossRef]
- Formisano, C.; Oliviero, F.; Rigano, D.; Saab, A.M.; Senatore, F. Chemical composition of essential oils and in vitro antioxidant properties of extracts and essential oils of Calamintha origanifolia and Micromeria myrtifolia, two Lamiaceae from the Lebanon flora. Ind. Crop. Prod. 2014, 62, 405–411. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cell growth of normal human non-immortalized buccal fibroblast cells (BFC) treated with essential oils (F1, L1, F2, L2, F3, L3, F4, L4) at 25 µg/mL concentration. * Significant vs. control untreated cells (p < 0.001).
Figure 1.
Cell growth of normal human non-immortalized buccal fibroblast cells (BFC) treated with essential oils (F1, L1, F2, L2, F3, L3, F4, L4) at 25 µg/mL concentration. * Significant vs. control untreated cells (p < 0.001).

Figure 2.
Caspase-3 activity in A375 cells treated for 72 h with essential oils F4 and L4. Hydrogen peroxide was used as positive control. * Significant vs. control untreated cells (p < 0.001).
Figure 2.
Caspase-3 activity in A375 cells treated for 72 h with essential oils F4 and L4. Hydrogen peroxide was used as positive control. * Significant vs. control untreated cells (p < 0.001).

Figure 3.
LDH release in A375 cells treated with essential oils F4 and L4 for 72 h. Hydrogen peroxide was used as a positive control. * Significant vs. control untreated cells (p < 0.001).
Figure 3.
LDH release in A375 cells treated with essential oils F4 and L4 for 72 h. Hydrogen peroxide was used as a positive control. * Significant vs. control untreated cells (p < 0.001).

Figure 4.
COMET assay of genomic DNA in A375 cancer cells treated with essential oils F4 and L4 for 72 h. Hydrogen peroxide was used as positive control. TMOM= tail moment expressed as the product of TD (distance between head and tail) and TDNA. * Significant vs. control untreated cells (p < 0.001).
Figure 4.
COMET assay of genomic DNA in A375 cancer cells treated with essential oils F4 and L4 for 72 h. Hydrogen peroxide was used as positive control. TMOM= tail moment expressed as the product of TD (distance between head and tail) and TDNA. * Significant vs. control untreated cells (p < 0.001).

Figure 5.
Levels of caspase 3, Bcl-2, Bax, cleaved caspase-9 proteins in A375 cells untreated and treated with essential oils F4 and L4 for 72 h. Values are expressed as arbitrary densitometric units (A.D.U.) corresponding to signal intensity present on the autoradiography of Western blots. A) Representative blots of control and treated cells are reported. * Significant vs. control untreated cells (p < 0.001).
Figure 5.
Levels of caspase 3, Bcl-2, Bax, cleaved caspase-9 proteins in A375 cells untreated and treated with essential oils F4 and L4 for 72 h. Values are expressed as arbitrary densitometric units (A.D.U.) corresponding to signal intensity present on the autoradiography of Western blots. A) Representative blots of control and treated cells are reported. * Significant vs. control untreated cells (p < 0.001).

Figure 6.
Levels of Hsp70 (Heat Shock Protein 70 kilodaltons), PTEN (Phosphatase and tensin homolog) and SOD (superoxide dismutase)proteins in A375 cells treated with essential oils F4 and L4 for 72 h. Values are expressed as arbitrary densitometric units (A.D.U.) corresponding to signal intensity present on the autoradiography of Western blots. A) Representative blots of control and treated cells are reported. * Significant vs. control untreated cells (p < 0.001).
Figure 6.
Levels of Hsp70 (Heat Shock Protein 70 kilodaltons), PTEN (Phosphatase and tensin homolog) and SOD (superoxide dismutase)proteins in A375 cells treated with essential oils F4 and L4 for 72 h. Values are expressed as arbitrary densitometric units (A.D.U.) corresponding to signal intensity present on the autoradiography of Western blots. A) Representative blots of control and treated cells are reported. * Significant vs. control untreated cells (p < 0.001).

Figure 7.
Reactive oxygen species (ROS) determination in A375 cells treated with essential oils F4 and L4 for 72 h. * Significant vs. control untreated cells (p < 0.001).
Figure 7.
Reactive oxygen species (ROS) determination in A375 cells treated with essential oils F4 and L4 for 72 h. * Significant vs. control untreated cells (p < 0.001).

Figure 8.
Levels of GSH in A375 cells treated with essential oils F4 and L4 for 72 h. * Significant vs. control untreated cells (p < 0.001).
Figure 8.
Levels of GSH in A375 cells treated with essential oils F4 and L4 for 72 h. * Significant vs. control untreated cells (p < 0.001).

Figure 9.
Representation of the A. arborescens EOs according to their chemotypes based on the major compounds camphor, chamazulene and β-thujone. Chemo I: camphor-chamazulene-β-thujone, Chemo II: β-thujone/chamazulene, Chemo III: chamazulene/camphor.
Figure 9.
Representation of the A. arborescens EOs according to their chemotypes based on the major compounds camphor, chamazulene and β-thujone. Chemo I: camphor-chamazulene-β-thujone, Chemo II: β-thujone/chamazulene, Chemo III: chamazulene/camphor.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Collection sites of A. arborescens samples and yields of the essential oils (EOs).
| No | Place of Collection | Coordinate | Altitude s/l | Date | Essential Oil Yield % |
|---|---|---|---|---|---|
| F1 | Capo Zafferano, Sicily | 38°06′38” N; 13°31′47” E | 22 m | 13 June 2018 | 1.25 ± 0.04 |
| L1 | Capo Zafferano, Sicily | 38°06′38” N; 13°31′47” E | 22 m | 13 June 2018 | 1.32 ± 0.03 |
| F2 | Alimena, Sicily | 37°40′50” N; 14°05′01” E | 645 m | 19 June 2018 | 0.93 ± 0.03 |
| L2 | Alimena, Sicily | 37°40′50” N; 14°05′01” E | 645 m | 19 June 2018 | 0.20 ± 0.02 |
| F3 | Lami, Lipari Island | 38°29′55” N, 14°56′47” E | 282 m | 16 June 2018 | 1.08 ± 0.03 |
| L3 | Lami, Lipari Island | 38°29′55” N, 14°56′47” E | 282 m | 16 June 2018 | 1.53 ± 0.04 |
| F4 | Acqua Calda, Lipari Island | 38°31′11” N, 14°56′00” E | 2 m | 17 June 2018 | 1.11 ± 0.03 |
| L4 | Acqua Calda, Lipari Island | 38°31′11” N, 14°56′00” E | 2 m | 17 June 2018 | 1.12 ± 0.03 |
Table 2.
Chemical composition of essential oils of the Artemisia arborescens samples F1–F4 and L1–L4.
Table 2.
Chemical composition of essential oils of the Artemisia arborescens samples F1–F4 and L1–L4.
| Ki a | Ki b | Compound | F1 % d | F2 % d | F3 % d | F4 % d | L1 % d | L2 % d | L3 % d | L4 % d | Id. c |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Monoterpenes | 7.4 | 2.6 | 5.9 | 5.2 | 6.2 | 0.9 | 9.9 | 0.5 | |||
| 925 | 1014 | Tricyclene | t | 1, 2 | |||||||
| 930 | 1014 | α-Thujene | 0.2 | t | t | 0.1 | 1,2 | ||||
| 938 | 1032 | α-Pinene | 0.7 | 0.2 | 1.9 | 2.3 | 0.6 | 3.7 | 0.2 | 1, 2, 3 | |
| 938 | 1117 | Verbenene | t | 1, 2 | |||||||
| 953 | 1076 | Camphene | 0.7 | 2.6 | 2.7 | 0.7 | 3.7 | 0.2 | 1, 2 | ||
| 973 | 1132 | Sabinene | 1.2 | 0.8 | 1.2 | 0.7 | 1, 2 | ||||
| 993 | 1174 | Myrcene | 1.5 | 0.4 | 1.1 | 0.1 | 1, 2, 3 | ||||
| 1005 | 1150 | α-Phellandrene | 0.1 | 1, 2 | |||||||
| 1012 | 1157 | δ3-Carene | 0.2 | 0.3 | 0.1 | ||||||
| 1013 | 1189 | α-Terpinene | 0.7 | 0.4 | t | 0.6 | 0.1 | t | 1, 2, 3 | ||
| 1025 | 1278 | p-Cymene | 0.2 | 0.1 | 0.3 | t | 1, 2, 3 | ||||
| 1029 | 1218 | β-Phellandrene | 0.4 | t | 1, 2 | ||||||
| 1030 | 1203 | Limonene | 0.3 | 1, 2, 3 | |||||||
| 1049 | 1265 | (E)-β-Ocimene | 2.2 | ||||||||
| 1057 | 1256 | γ-Terpinene | 1.4 | 0.6 | 0.1 | 1.1 | t | 1, 2, 3 | |||
| 1086 | 1265 | Terpinolene | 0.3 | 0.1 | 1.3 | 0.2 | 1, 2, 3 | ||||
| Oxygenated monoterpenes | 42.5 | 48.5 | 58.3 | 55.6 | 31.5 | 37.9 | 71.8 | 24.3 | |||
| 1034 | 1213 | 1,8-Cineole | 0.2 | 0.2 | 0.1 | 1, 2, 3 | |||||
| 1063 | 1555 | (Z)-Sabinene hydrate | 0.8 | 0.1 | 0.8 | 1, 2 | |||||
| 1098 | 1457 | (E)-Sabinene hydrate | 0.2 | 1, 2 | |||||||
| 1098 | 1553 | Linalool | 1.2 | 1, 2, 3 | |||||||
| 1105 | 1431 | α-Thujone | 0.8 | 1.3 | 0.6 | 1, 2 | |||||
| 1115 | 1451 | β-Thujone | 26.7 | 44.8 | 13.5 | 34.4 | 1, 2 | ||||
| 1125 | 1540 | Chrysanthenone | 0.1 | 1, 2 | |||||||
| 1125 | 1623 | Myrcenol | 0.2 | 1, 2 | |||||||
| 1126 | 1637 | cis-p-Menth-2-en-1-ol | 0.1 | 1, 2 | |||||||
| 1145 | 1532 | Camphor | 8.5 | 58.2 | 54.2 | 10.0 | 65.6 | 21.5 | 1, 2, 3 | ||
| 1167 | 1718 | Borneol | 0.2 | t | 0.2 | 1.7 | 0.2 | 1, 2, 3 | |||
| 1176 | 1611 | Terpinen-4-ol | 2.2 | 1.4 | 1.4 | 2.4 | 0.6 | 4.5 | 2.5 | 1, 2, 3 | |
| 1189 | 1706 | α-Terpineol | 0.3 | 0.2 | 0.3 | 0.1 | 1, 2, 3 | ||||
| 1192 | 1805 | Myrtenol | t | 1, 2 | |||||||
| 1233 | 1662 | Pulegone | 1.3 | 1, 2 | |||||||
| 1259 | 1665 | Linalyl acetate | 0.1 | ||||||||
| 1259 | 1891 | Myrtanol | 0.5 | 0.3 | 1, 2 | ||||||
| 1261 | 1809 | Perilla aldehyde | 0.4 | 0.2 | 0.2 | 1, 2 | |||||
| 1284 | 1597 | Bornyl acetate | 1.6 | 1.9 | 1, 2, 3 | ||||||
| 1289 | 1812 | p-Mentha-1,3-dien-7-al (α-Terpinen-7-al) | 0.1 | 1, 2 | |||||||
| 1329 | 1949 | Piperitenone | 1.0 | 1, 2 | |||||||
| 1602 | 1893 | Geranyl isovalerate | 0.2 | 0.3 | 1, 2 | ||||||
| Sesquiterpenes | 5.6 | 2.2 | - | - | 4.8 | 1.5 | 1.7 | 3.2 | |||
| 1373 | 1493 | α-Ylangene | 0.3 | 1, 2 | |||||||
| 1377 | 1497 | α-Copaene | 0.3 | 0.2 | 0.1 | 1, 2 | |||||
| 1385 | 1535 | β-Bourbonene | 0.1 | 0.1 | 1, 2 | ||||||
| 1418 | 1612 | (E)-Caryophyllene | 0.8 | 0.3 | 1.2 | 1.7 | 1.3 | 1, 2, 3 | |||
| 1455 | 1689 | α-Humulene | 0.1 | 0.1 | 0.2 | 1, 2 | |||||
| 1463 | 1667 | allo-Aromadendrene | 0.1 | ||||||||
| 1477 | 1726 | Germacrene D | 2.6 | 1.7 | 2.4 | 1.5 | 1.1 | 1, 2 | |||
| 1491 | 1756 | Bicyclogermacrene | 0.2 | 0.1 | 1, 2 | ||||||
| 1498 | 1744 | α-Selinene | 0.2 | ||||||||
| 1503 | 1740 | α-Muurolene | 0.1 | 1, 2 | |||||||
| 1506 | 1760 | (E,E)-α-Farnesene | 0.4 | 0.2 | 1, 2 | ||||||
| 1509 | 1746 | cis-(Z)-α-Bisabolene | 0.2 | t | 1,2 | ||||||
| 1510 | 1743 | β-Bisabolene | 0.3 | 1, 2 | |||||||
| 1526 | 1773 | δ-Cadinene | 0.2 | 0.2 | 0.1 | 1, 2 | |||||
| 1541 | 1918 | α-Calacorene | 0.4 | 0.2 | 1, 2 | ||||||
| 1554 | 1856 | Germacrene B | t | ||||||||
| Oxygenated sesquiterpenes | 1.6 | - | - | - | 2.4 | - | - | 4.0 | |||
| 1578 | 2150 | Spathulenol | 0.2 | 0.1 | 1, 2 | ||||||
| 1579 | 2208 | Caryophyllene oxide | 0.1 | 0.4 | 0.8 | 1, 2, 3 | |||||
| 1605 | (2R,5E)-Caryophyll-5-en-12-al | t | 1, 2 | ||||||||
| 1627 | (2S,5E)-Caryophyll-5-en-12-al | t | 0.5 | 1, 2 | |||||||
| 1632 | 2371 | Caryophylla-3,8(13)-dien-5α-ol | 0.4 | 0.7 | 0.4 | 1, 2 | |||||
| 1640 | 2361 | Caryophylla-4(12),8(13)-dien-5β-ol | 0.3 | 0.5 | 1, 2 | ||||||
| 1645 | 2209 | Torreyol | 0.4 | ||||||||
| 1650 | 2258 | β-Eudesmol | 0.3 | 0.5 | 1.3 | 1, 2 | |||||
| 1652 | 2255 | α-Cadinol | 0.4 | 0.3 | 1, 2 | ||||||
| 1665 | 2371 | Caryophylla-3,8(13)-dien-5β-ol | 0.3 | 1, 2 | |||||||
| 1765 | 2518 | cis-Lanceol | 0.1 | 1, 2 | |||||||
| 2075 | Caryophylla-2(12),6(13)-dien-5-one | t | 1, 2 | ||||||||
| Carbonilic compounds | 0.5 | 0.7 | - | - | 0.2 | 0.2 | - | - | |||
| 963 | 1543 | Benzaldehyde | t | 1, 2, 3 | |||||||
| 989 | 1349 | 6-Methyl-5-hepten-2-one | 0.5 | 0.4 | 0.1 | 1, 2 | |||||
| 1621 | 1(2H)-Naphthalenone, 2,2,3-trimethyl | 0.1 | 0.2 | 1, 2 | |||||||
| 1702 | 2416 | 4-Isopropyl-6-methyl-1-tetralone | 0.3 | 1, 2 | |||||||
| Phenols | 0.6 | 0.2 | - | - | - | - | - | - | |||
| 1206 | 1805 | 4-Methyl veratrole | 0.5 | 0.2 | 1, 2 | ||||||
| 1400 | 2016 | Methyl eugenol | 0.1 | 1, 2, 3 | |||||||
| Hydrocarbons | 38.9 | 37.4 | 34.2 | 39.0 | 51.8 | 55.6 | 14.9 | 63.0 | |||
| 1338 | Naphthalene, 1,4-dihydro-2,5,8-trimethyl- | 0.1 | 1, 2 | ||||||||
| 1358 | Naphthalene, 1,2-dihydro-2,5,8-trimethyl | 0.1 | 0.2 | 1, 2 | |||||||
| 1735 | 2434 | Chamazulene | 38.8 | 37.4 | 34.2 | 39.0 | 51.7 | 55.4 | 14.9 | 63.0 | 1, 2 |
| Others | 0.2 | 0.1 | - | - | 0.3 | - | - | - | |||
| 1005 | 1174 | 2-Methylbutyl isobutanoate | 0.1 | 1, 2 | |||||||
| 1398 | 1904 | Benzyl isovalerate | 0.2 | 1, 2 | |||||||
| 1189 | 1789 | Methyl salicylate | 0.2 | 1, 2, 3 | |||||||
| 1233 | 1484 | cis-3-Hexenyl-2-Methylbutanoate | 0.1 | 1, 2 | |||||||
| TOTAL | 97.3 | 91.7 | 98.4 | 99.8 | 97.2 | 96.1 | 98.3 | 95.0 |
a Ki: Retention index on a HP-5MS column; b Ki: Retention index on a HP-Innowax column; c Id: Identification: 1 = comparison of retention index; 2 = comparison of mass spectra with MS libraries; 3 = comparison with authentic compounds; d t: trace, less than 0.05%.
Table 3.
IC50 values of melanoma cells A375 treated with the essential oils after 72 h.
| Treatments | A375 Cells |
|---|---|
| Essential Oils | IC50 a (µg/mL) |
| F1 | 21.1 ± 0.17 |
| L1 | 23.4 ± 0.15 |
| F2 | 15.3 ± 0.13 |
| L2 | 16.2 ± 0.17 |
| F3 | 16.3 ± 0.14 |
| L3 | 19.7 ± 0.18 |
| F4 | 6.7 ± 0.22 |
| L4 | 4.5 ± 0.18 |
a Results are expressed as IC50 values (mg/mL) ± SD. The IC50 value, relative to untreated control, represents the concentration that inhibited cell vitality by 50%. Each value represents the mean ± SD of three experiments, performed in quadruplicate.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Russo, A.; Bruno, M.; Avola, R.; Cardile, V.; Rigano, D. Chamazulene-Rich Artemisia arborescens Essential Oils Affect the Cell Growth of Human Melanoma Cells. Plants 2020, 9, 1000. https://doi.org/10.3390/plants9081000
AMA Style
Russo A, Bruno M, Avola R, Cardile V, Rigano D. Chamazulene-Rich Artemisia arborescens Essential Oils Affect the Cell Growth of Human Melanoma Cells. Plants. 2020; 9(8):1000. https://doi.org/10.3390/plants9081000
Chicago/Turabian StyleRusso, Alessandra, Maurizio Bruno, Rosanna Avola, Venera Cardile, and Daniela Rigano. 2020. "Chamazulene-Rich Artemisia arborescens Essential Oils Affect the Cell Growth of Human Melanoma Cells" Plants 9, no. 8: 1000. https://doi.org/10.3390/plants9081000
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.