Genetic Impairment of Cellulose Biosynthesis Increases Cell Wall Fragility and Improves Lipid Extractability from Oleaginous Alga Nannochloropsis salina

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions
2.2. Generation of cesA KO and Knockdown (KD) Mutants
2.3. Growth Rate Determination, Pigment Content Analysis, and Chlorophyll Fluorescence Measurement
2.4. Light, Fluorescence, and Electron Microscopy
2.5. Cell Breakage, Cell Wall Extraction, and Cellulose and Chrysolaminarin Quantification
2.6. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (RT-qPCR)
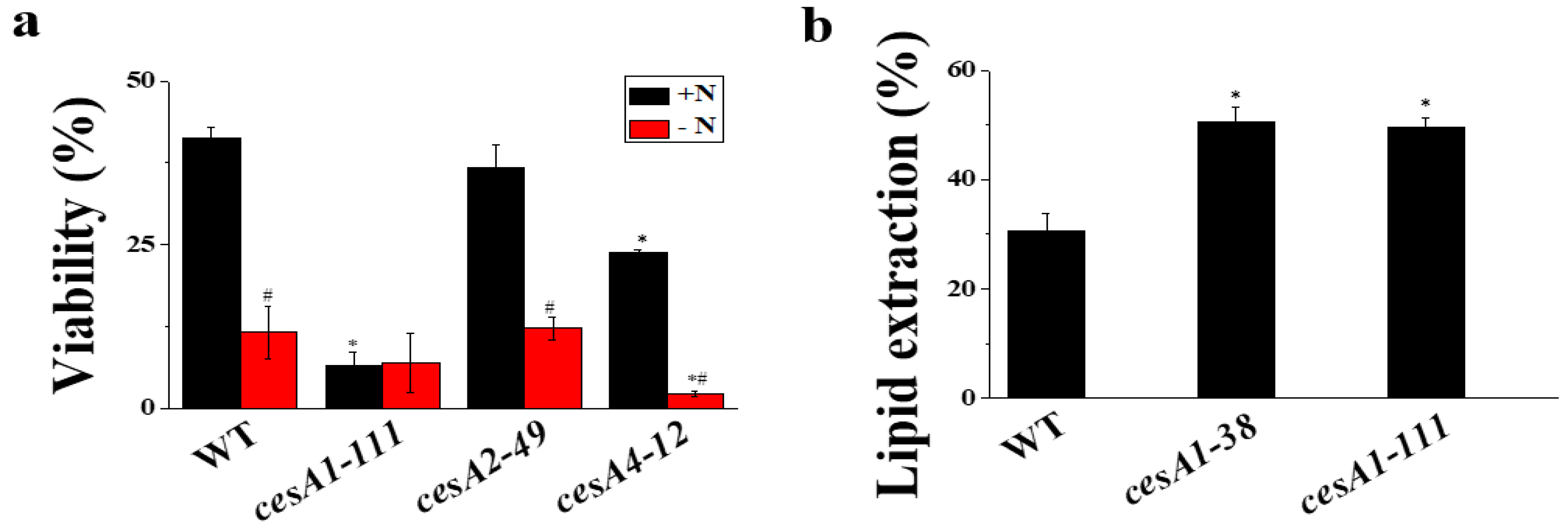
2.7. Cell Breakage, Lipid Extraction Efficiency, Analysis of Fatty Acid Methyl Esters (FAME), and Viability Assays
2.8. Metabolic Profiling
2.9. Statistical Analysis
3. Results
3.1. Generation of cesA KO and KD Mutants
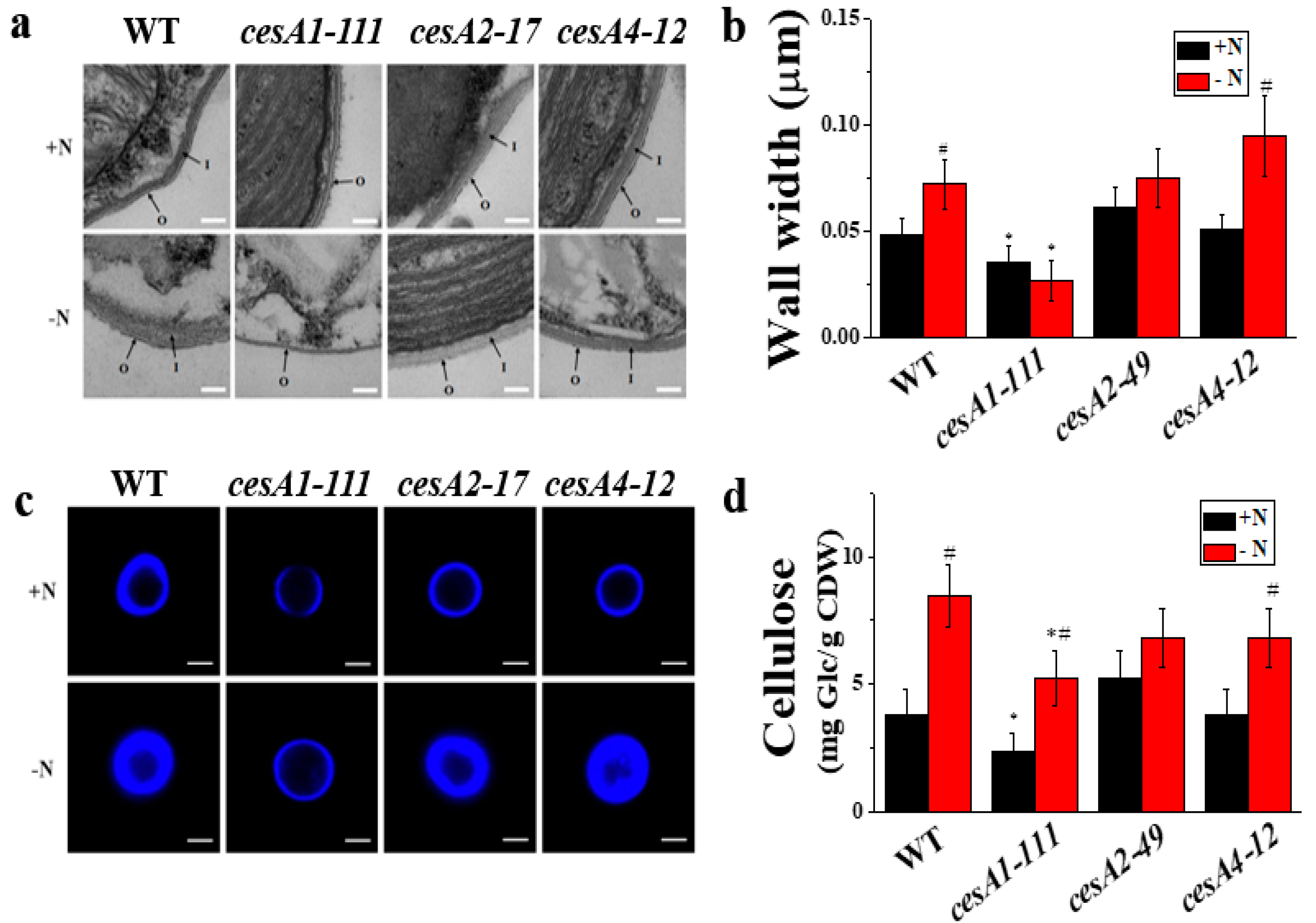
3.2. cesA1 Mutant Exhibits Thinned Cell Wall
3.3. Growth and Photosynthesis of cesA1-111 Mutant Are Comparable to Those of the WT
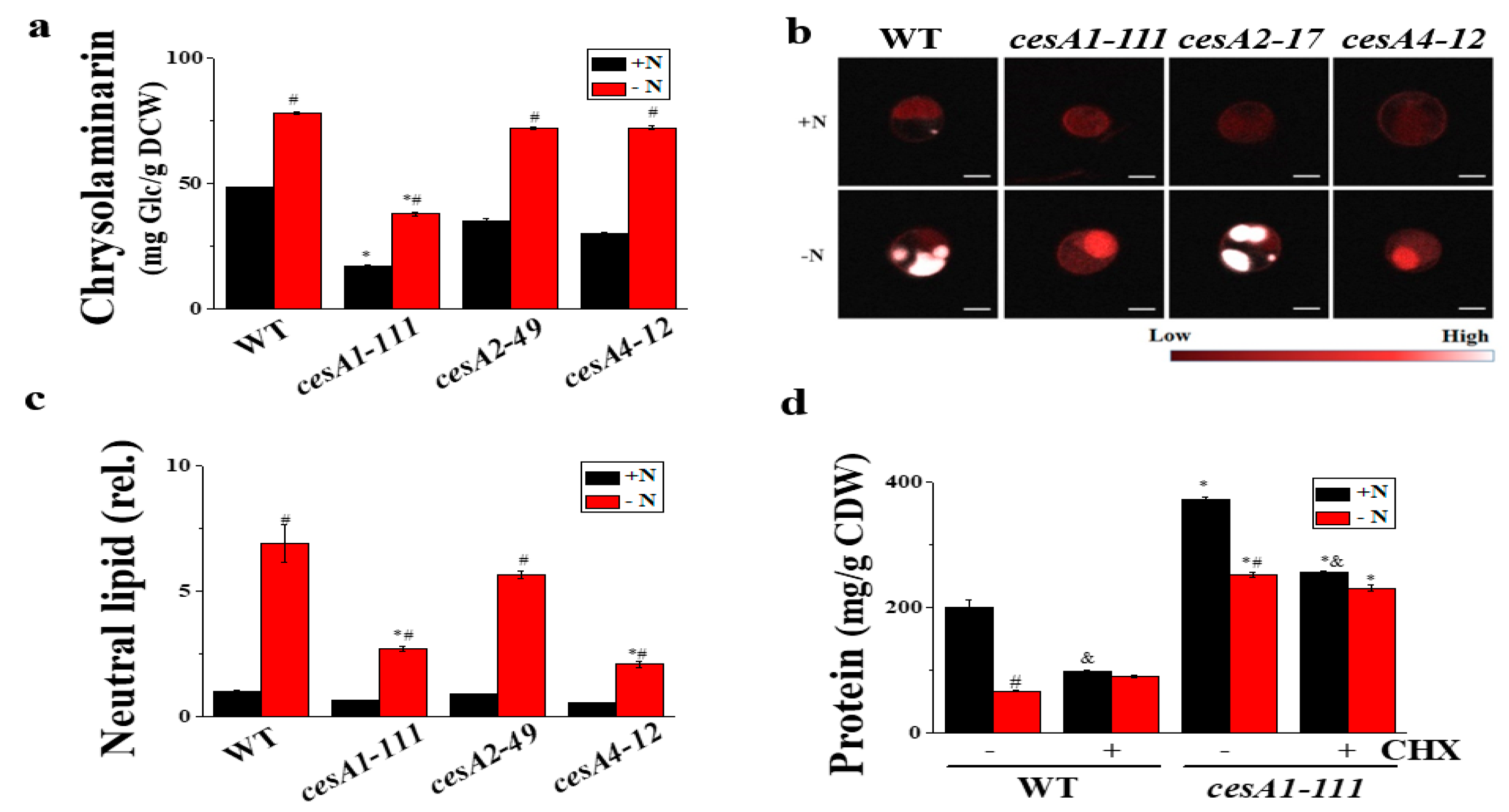
3.4. cesA1-111 Mutant Has Reduced Chrysolaminarin and Lipid Contents, but Enhanced Protein Content
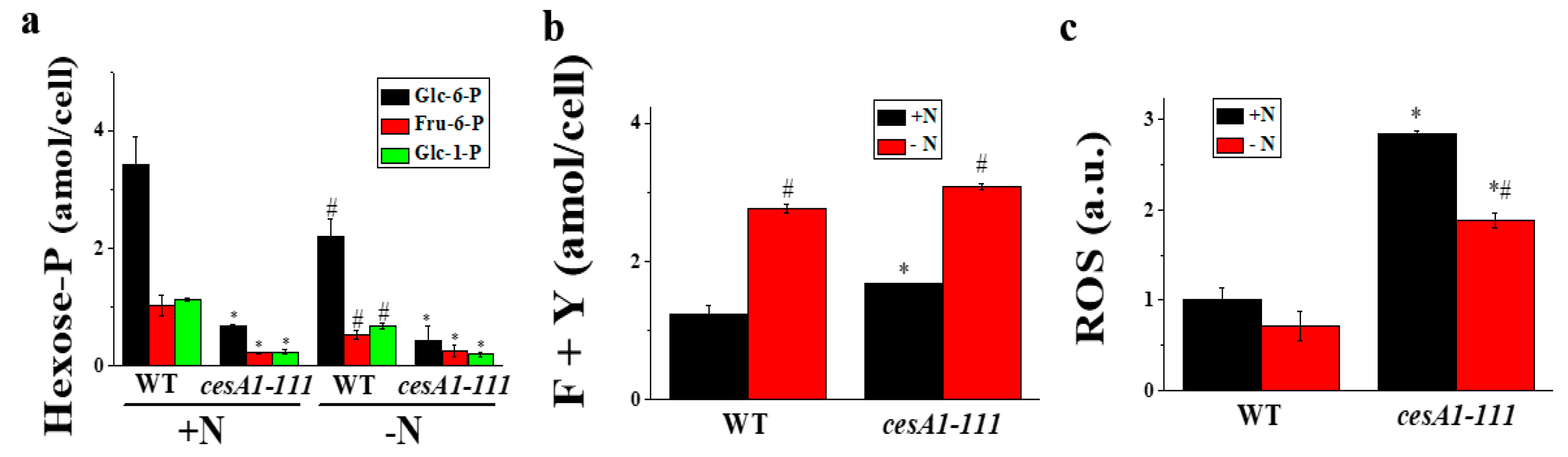
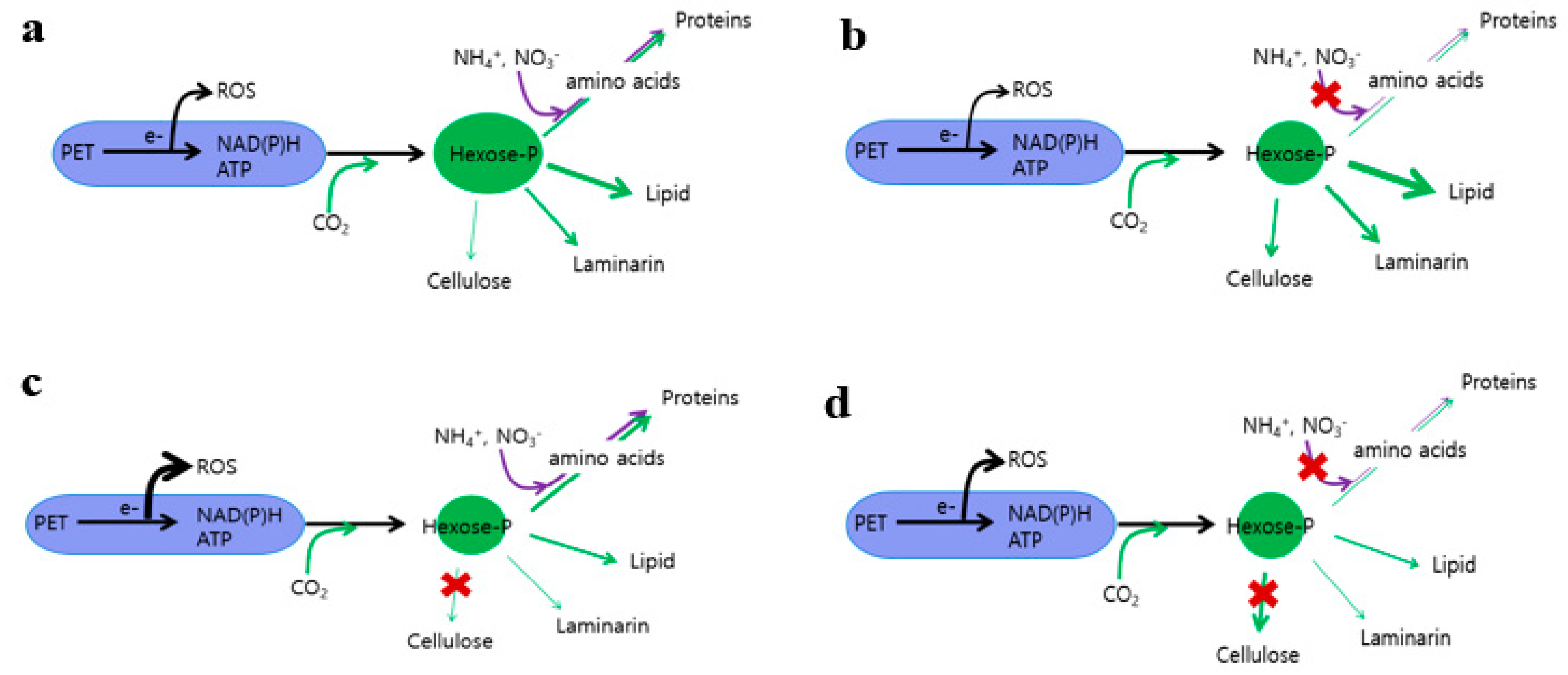
3.5. Hexose-P, Amino Acids, and ROS Contents
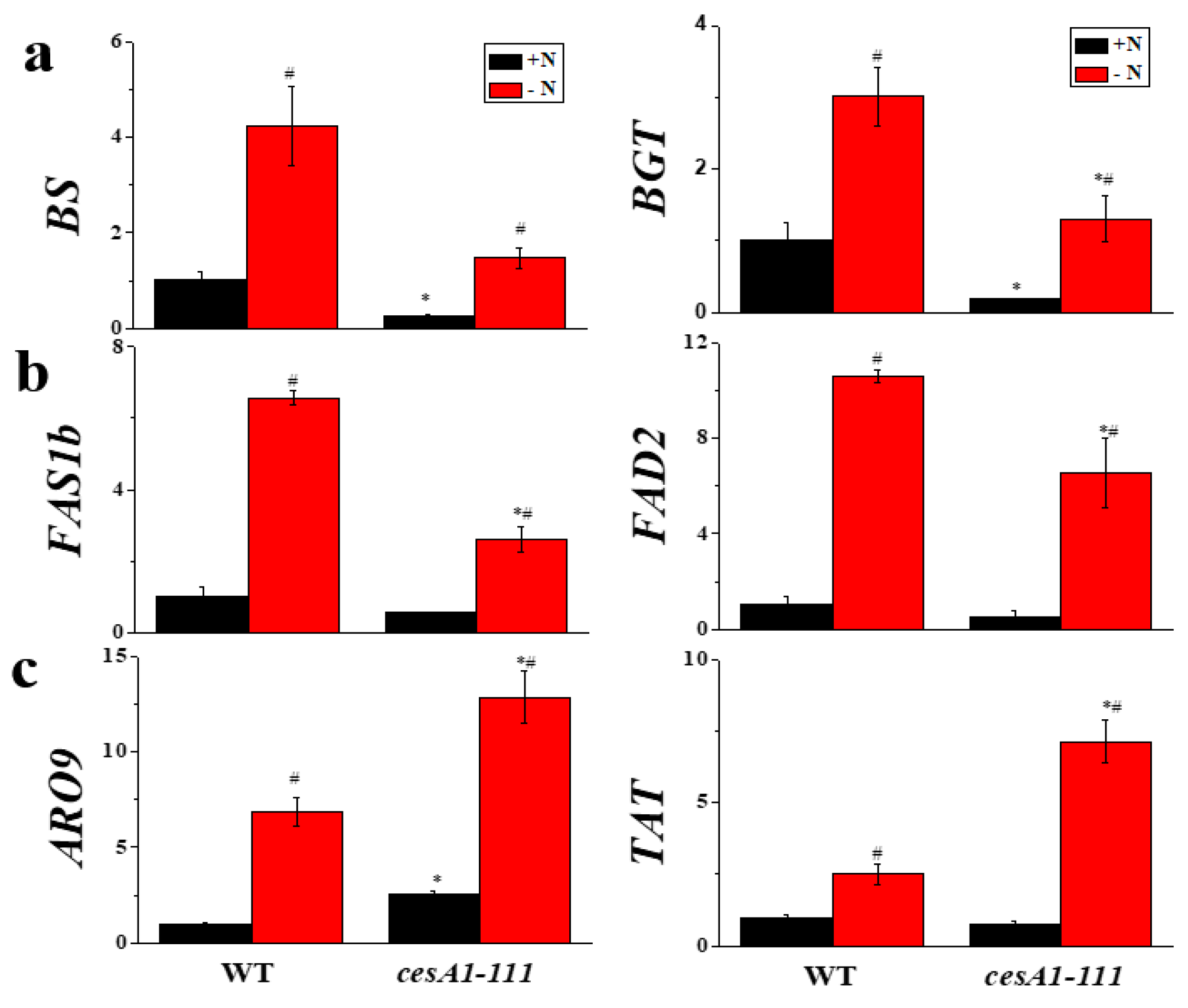
3.6. Transcript Levels of Genes Involved in Chrysolaminarin, Lipid, and Protein Biosynthesis
3.7. The Cell Wall of cesA1-111 Mutant Is Prone to Mechanical Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wagner, H.; Jakob, T.; Fanesi, A.; Wilhelm, C. Towards an understanding of the molecular regulation of carbon allocation in diatoms: The interaction of energy and carbon allocation. Philos. Trans. R. Soc. B 2017, 372, 20160410. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Radakovits, R.; Jinkerson, R.E.; Fuerstenberg, S.I.; Tae, H.; Settlage, R.E.; Boore, J.L.; Posewitz, M.C. Draft genome sequence and genetic transformation of the oleaginous alga Nannochloropis gaditana. Nat. Commun. 2012, 3, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieler, A.; Wu, G.; Tsai, C.H.; Bullard, B.; Harvey, C.; Reca, I.B.; Thornburg, C.; Achawanantakun, R.; Buehl, C.J.; Campbell, M.S.; et al. Genome, functional gene annotation, and nuclear transformation of the heterokont oleaginous alga Nannochloropsis oceanica CCMP1779. PLoS Genet. 2012, 8, e1003064. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lu, Y.; Xin, Y.; Huang, S.; Xu, J. Genome editing of model microalgae Nannochloropsis spp. by CRISPR/Cas9. Plant. J. 2016, 88, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Beacham, T.A.; Bradley, C.; White, D.A.; Bond, P.; Ali, S.T. Lipid productivity and cell wall ultrastructure of six strains of Nannochloropsis: Implications for biofuel production and downstream processing. Algal Res. 2014, 6, 64–69. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, Z.; Yu, C.; Yin, Y.; Zhou, G. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production. Bioresour. Technol. 2014, 167, 503–509. [Google Scholar] [CrossRef]
- Li, X.; Wang, M.; Liao, X.; Chen, H.; Dai, Y.; Chen, B. Two stages of N-deficient cultivation enhance the lipid content of microalga Scenedesmus sp. J. Am. Oil Chem. Soc. 2015, 92, 503–512. [Google Scholar] [CrossRef]
- Jeong, S.W.; Nam, S.W.; HwangBo, K.; Jeong, W.J.; Chang, Y.K.; Park, Y.-I. Transcriptional regulation of cellulose biosynthesis during the early phase of nitrogen deprivation in Nannochloropsis salina. Sci. Rep. 2017, 7, 524. [Google Scholar] [CrossRef]
- Work, V.H.; Radakvits, R.; Jinkerson, R.E.; Meuser, J.E.; Elliott, L.G.; Vinyard, D.J.; Laurens, L.M.L.; Dismukes, G.C.; Posewitz, M.C. Increased lipid accumulation in the Chlamydomonas reinhardtii sta7-10 strachless isoamylase mutant and increased carbohydrate synthesis in complemented strains. Eukar. Cell 2010, 9, 1251–1261. [Google Scholar] [CrossRef] [Green Version]
- Yap, B.H.J.; Crawford, S.A.; Dagastine, R.R.; Scales, P.J.; Martin, G.J.O. Nitrogen deprivation of microalgae: Effect on cell size, cell wall thickness, cell strength, and resistance to mechanical disruption. J. Ind. Microbiol. Biotechnol. 2016, 43, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Van Donk, E.; Lurling, M.; Hessen, D.O.; Lokhorst, G.M. Altered cell wall morphology in nutrient deficient phytoplankton and its impact on grazers. Limnol. Oceanogr. 1997, 42, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Scholz, M.J.; Weiss, T.L.; Jinkerson, R.E.; Goodenough, U.; Posewitz, M.C.; Gerken, H.G. Ultrastructure and composition of the Nannochloropsis gaditana cell wall. Eukaryot. Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Schwede, S.; Rehman, Z.U.; Gerber, M.; Theiss, C.; Span, R. Effects of thermal pretreatment on anaerobic digestion of Nannochloropsis salina biomass. Bioresour. Technol. 2013, 143, 505–511. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Kwon, S.; Kang, N.K.; Koh, H.G.; Shin, S.J.; Lee, B.; Jeong, B.; Chang, Y.K. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2018, 115, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Kang, N.K.; Suh, W.I.; Koh, H.G.; Lee, B.; Chang, Y.K. Optimization of electroporation-based multiple pulses and further improvement of transformation efficiency using bacterial conditioned medium for Nannochloropsis salina. J. Appl. Phycol. 2019, 31, 1153–1161. [Google Scholar] [CrossRef]
- Moran, R.; Porath, D. Chlorophyll determination in intact tissues using N, N-dimethylformamide. Plant. Physiol. 1980, 65, 478–479. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA Bioenergy 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Herth, W.; Schnepf, E. The fluorochrome, calcofluor white, binds oriented to structural polysaccharide fibrils. Protoplasma 1980, 105, 129–133. [Google Scholar] [CrossRef]
- Gusbeth, C.A.; Eing, C.; Göttel, M.; Sträßner, R.; Frey, W. Fluorescence diagnostics for lipid status monitoring of microalgae during cultivation. Int. J. Renew. Energy Biofuels 2016, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Hwangbo, K.; Lim, J.M.; Jeong, S.W.; Vikramathithan, J.; Park, Y.-I.; Jeong, W.J. Elevated inorganic carbon concentrating mechanism confer tolerance to high light in an Arctic Chlorella sp. ArM0029B. Front. Plant. Sci. 2018, 9, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Cooksey, K.E.; Guckert, J.B.; Williams, S.A.; Gallis, P.R. Fluorometric determination of the neutral lipid content of microalgal cells using Nile Red. J. Microbiol. Methods 1987, 6, 333–345. [Google Scholar] [CrossRef]
- Domergue, F.; Abbadi, A.; Ott, C.; Zank, T.K.; Zahringer, U.; Heinz, Z. Acyl carriers used as substrates by the desaturases and elongases involved in very long-chain polyunsaturated fatty acids biosynthesis reconstituted in yeast. J. Biol. Chem. 2003, 278, 35115–35126. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Kumaraswamy, G.K.; Vinyard, D.J.; Gu, H.Y.; Ananyev, G.; Posewitz, M.C.; Dismukes, G.C. Metabolic and photosynthetic consequences of blocking starch biosynthesis in the green alga Chlamydomonas reinhardtii sta6 mutant. Plant. J. 2015, 81, 947–960. [Google Scholar] [CrossRef]
- Shi, K.; Gao, Z.; Shi, T.Q.; Song, P.; Ren, L.J.; Huang, H.; Ji, X.J. Reactive oxygen species-mediated cellular stress response and lipid accumulation in oleaginous microorganisms: The state of the art and future perspectives. Front. Microbiol. 2017, 8, 793. [Google Scholar] [CrossRef]
- Jacquier, N.; Choudhary, V.; Mari, M.; Toulmay, A.; Reggiori, F.; Schneiter, R. Lipid droplets are functionally connected to the endoplasmic reticulum in Saccharomyces Cerevisiae. J. Cell Sci. 2011, 124, 2424–2437. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zhang, H.Y.; Chen, H.Q.; Chen, Y.Q.; Chen, W.; Song, Y.D.; Zhang, H.Y.; Chen, H.Q.; Chen, Y.Q.; Chen, W.; et al. Effects of 20 standard amino acids on the growth, total fatty acids production, and gamma-linolenic acid yield in Mucor circinelloides. Curr. Microbiol. 2014, 69, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Juppner, J.; Mubeen, U.; Leisse, A.; Caldana, C.; Wiszniewski, A.; Steinhauser, D.; Giavalisco, P. The target of rapamycin kinase affects biomass accumulation and cell cycle progression by altering carbon/nitrogen balance in synchronized Chlamydomonas reinhardtii cells. Plant. J. 2018, 93, 355–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, L.T.; Levitan, O.; Frada, M.J.; Sun, J.S.; Falkowski, P.G.; Dismukes, G.C.; Levitan, O.; Frada, M.J.; Sun, J.S.; Falkowski, P.G.; et al. Regulatory branch points affecting protein and lipid biosynthesis in the diatom Phaeodactylum tricornutum. Biomass Bioenergy 2013, 59, 306–315. [Google Scholar] [CrossRef]
- Abbriano, R.; Vardar, N.; Yee, D.; Hildebrand, M. Manipulation of a glycolytic regulator alters growth and carbon partitioning in the marine diatom Thalassiosira pseudonana. Algal Res. 2018, 32, 250–258. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | SGR | Chl | Car | ФPSII | NPQ | O2 Evolution |
|---|---|---|---|---|---|---|
| WT | ||||||
| +N | 0.14 ± 0.01 | 203.18 ± 2.14 | 30.23 ± 1.61 | 0.58 ± 0.03 | 1.58 ± 0.32 | 0.34 ± 0.04 |
| −N | 0.05 ± 0.02 # | 112.67 ± 3.28 # | 19.38 ± 0.79 # | 0.49 ± 0.01 # | 3.09 ± 0.02 # | 0.41 ± 0.02 |
| cesA1-111 | ||||||
| +N | 0.11 ± 0.01 * | 172.36 ± 8.46 * | 28.86 ± 2.70 | 0.59 ± 0.02 | 1.70 ± 0.30 | 0.37 ± 0.02 |
| −N | 0.03 ± 0.01 # | 90.16 ± 2.5 *,# | 17.15 ± 0.35 # | 0.52 ± 0.02 | 3.38 ± 0.0 # | 0.41 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.W.; HwangBo, K.; Lim, J.M.; Nam, S.W.; Lee, B.S.; Jeong, B.-r.; Chang, Y.K.; Jeong, W.-J.; Park, Y.-I. Genetic Impairment of Cellulose Biosynthesis Increases Cell Wall Fragility and Improves Lipid Extractability from Oleaginous Alga Nannochloropsis salina. Microorganisms 2020, 8, 1195. https://doi.org/10.3390/microorganisms8081195
Jeong SW, HwangBo K, Lim JM, Nam SW, Lee BS, Jeong B-r, Chang YK, Jeong W-J, Park Y-I. Genetic Impairment of Cellulose Biosynthesis Increases Cell Wall Fragility and Improves Lipid Extractability from Oleaginous Alga Nannochloropsis salina. Microorganisms. 2020; 8(8):1195. https://doi.org/10.3390/microorganisms8081195
Chicago/Turabian StyleJeong, Seok Won, Kwon HwangBo, Jong Min Lim, Seung Won Nam, Bong Soo Lee, Byeong-ryool Jeong, Yong Keun Chang, Won-Joong Jeong, and Youn-Il Park. 2020. "Genetic Impairment of Cellulose Biosynthesis Increases Cell Wall Fragility and Improves Lipid Extractability from Oleaginous Alga Nannochloropsis salina" Microorganisms 8, no. 8: 1195. https://doi.org/10.3390/microorganisms8081195