Reproductive Outcomes and Endocrine Profile in Artificially Inseminated versus Embryo Transferred Cows

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Oocyte Collection and In Vitro Maturation
2.3. In Vitro Fertilization
2.4. Embryo Culture
2.5. Embryo Vitrification and Warming
2.6. Recipient Synchronization and Embryo Transfer
2.7. Artificial Insemination
2.8. Pregnancy Detection and Follow-up until Parturition
2.9. Blood Collection and Analysis
2.10. Statistical Analysis
3. Results
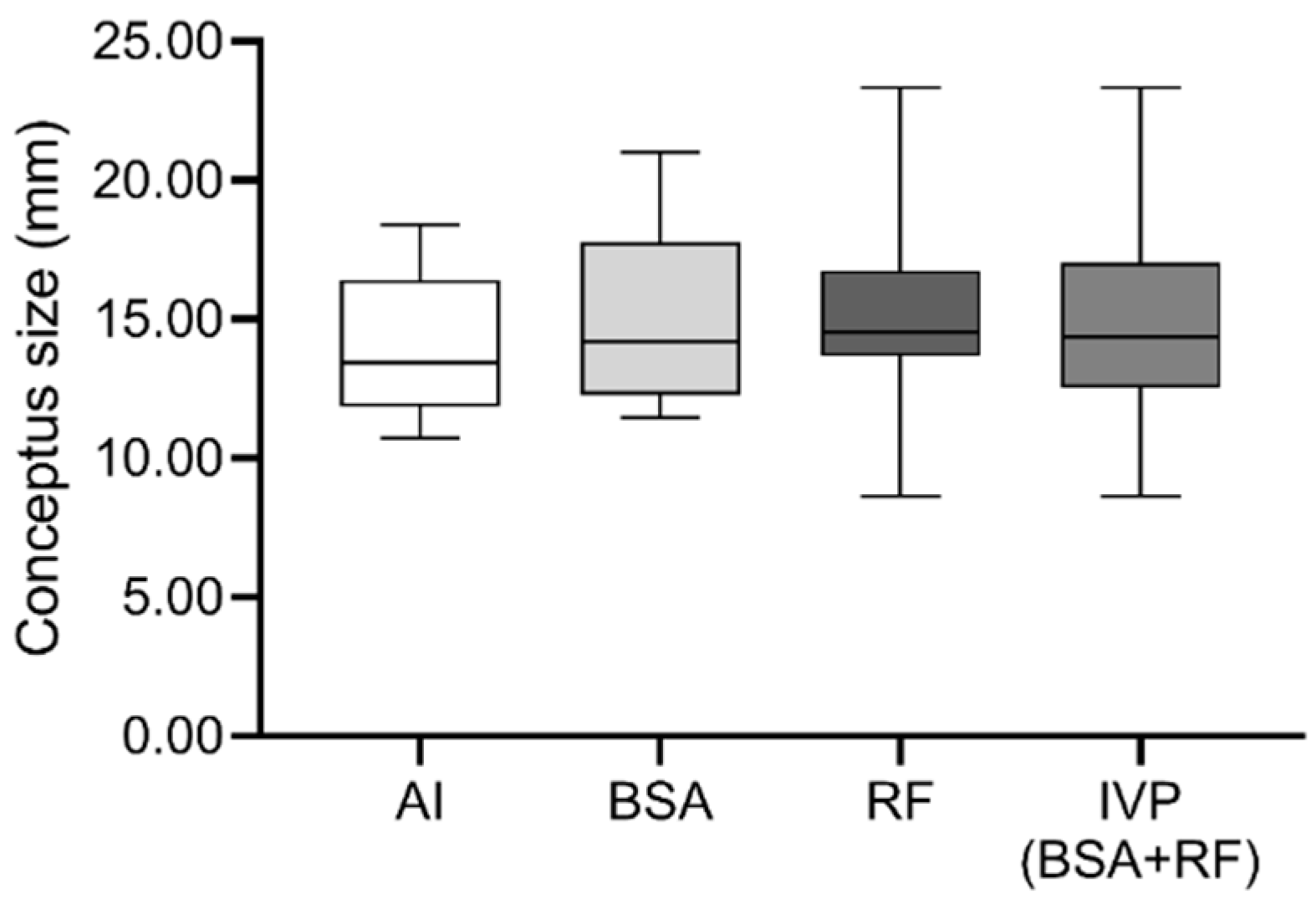
3.1. Rate of In Vitro Produced Embryos, Pregnancy Maintenance and Conceptus Size after Embryo Transfer/Artificial Insemination, Were Similar between Groups
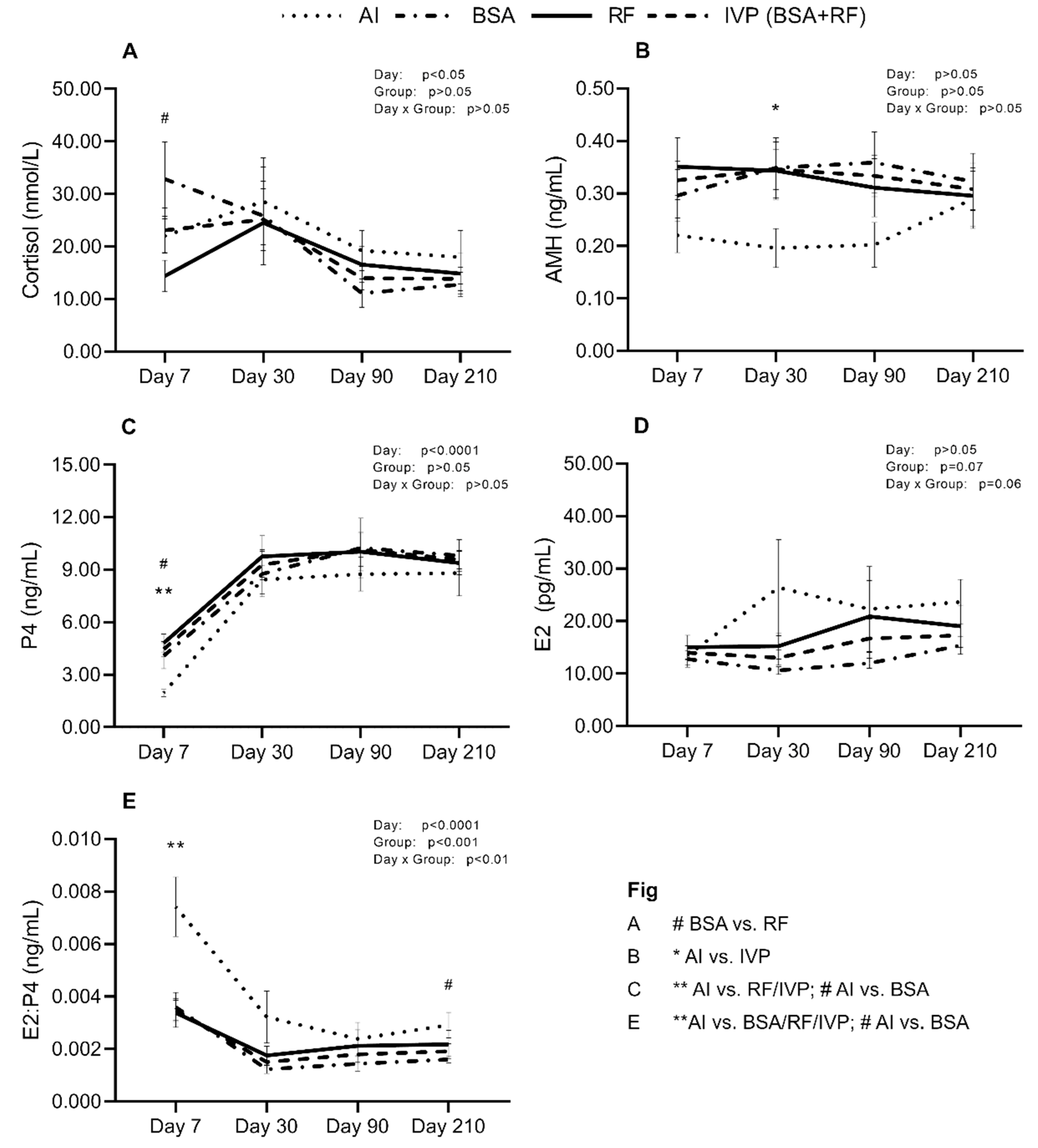
3.2. Recipient’s Hormonal Levels at Day 7, 30, 90, and 210 of Gestation Showed Differences at Specific Time-Points
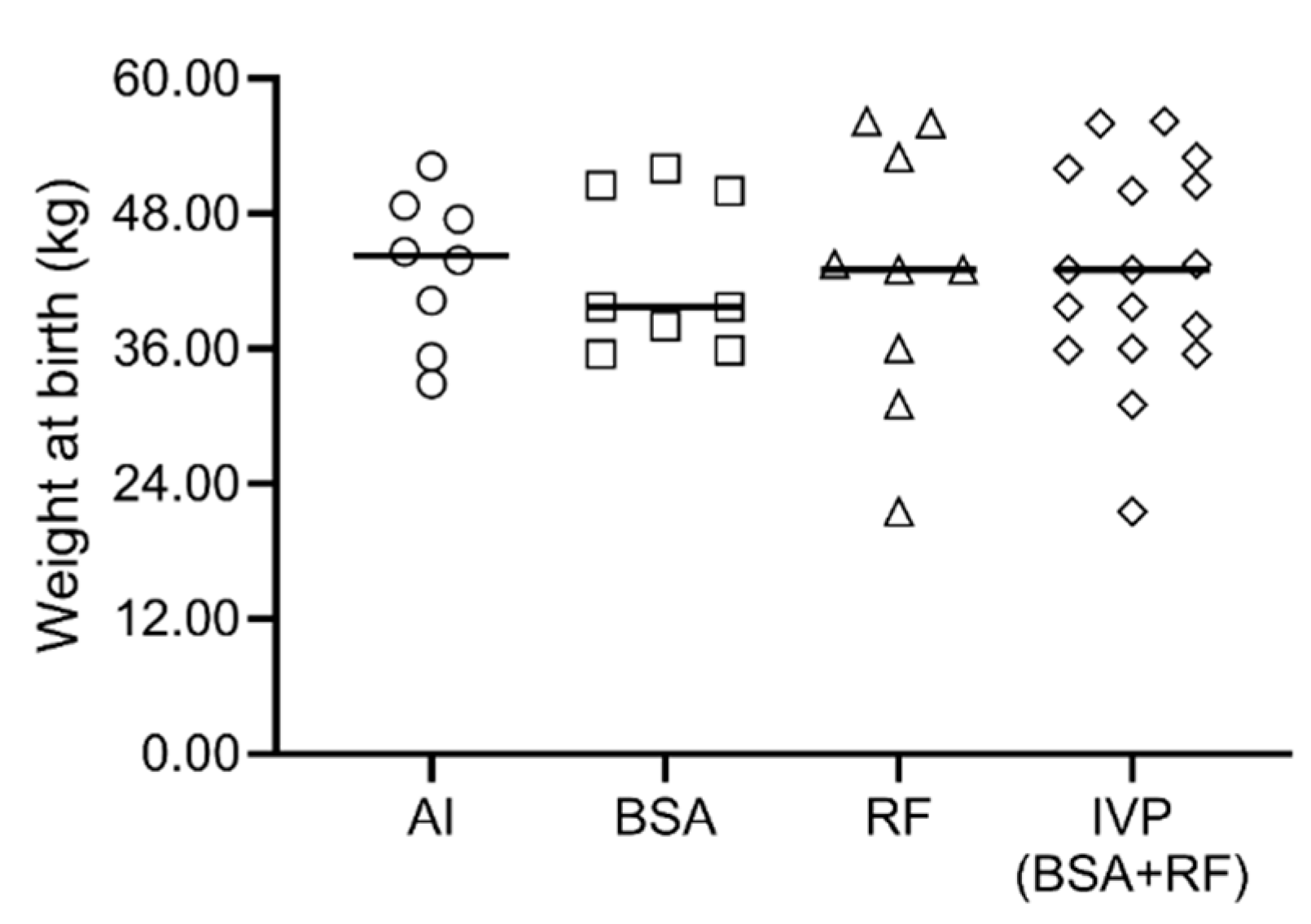
3.3. Gestation Length, Parturitions and Neonatal Period
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Viana, J. IETS Data Retrieval Committee. 2018 statistics of embryo production and transfer in domestic farm animals. Embryo Technol. Newsl. 2019, 37, 7–25. [Google Scholar]
- Drost, M.; Ambrose, J.D.; Thatcher, M.-J.; Cantrell, C.K.; Wolfsdorf, K.E.; Hasler, J.F.; Thatcher, W.W. Conception rates after artificial insemination or embryo transfer in lactating dairy cows during summer in florida. Theriogenology 1999, 52, 1161–1167. [Google Scholar] [CrossRef]
- Pontes, J.H.F.; Nonato-Junior, I.; Sanches, B.V.; Ereno-Junior, J.C.; Uvo, S.; Barreiros, T.R.R.; Oliveira, J.A.; Hasler, J.F.; Seneda, M.M. Comparison of embryo yield and pregnancy rate between in vivo and in vitro methods in the same Nelore (Bos indicus) donor cows. Theriogenology 2009, 71, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Taverne, M.; Breukelman, S.; Perényi, Z.; Dieleman, S.; Vos, P.; Jonker, H.; de Ruigh, L.; van Wagtendonk-de Leeuw, J.M.; Beckers, J.-F. The monitoring of bovine pregnancies derived from transfer of in vitro produced embryos. Reprod. Nutr. Dev. 2002, 42, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.M.; Block, J.; Morelli, P.; Navarette, A.E.; Amstalden, M.; Bonilla, L.; Hansen, P.J.; Bilby, T.R. Efficacy of embryo transfer in lactating dairy cows during summer using fresh or vitrified embryos produced in vitro with sex-sorted semen. J. Dairy Sci. 2011, 94, 3437–3445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dochi, O.; Takahashi, K.; Hirai, T.; Hayakawa, H.; Tanisawa, M.; Yamamoto, Y.; Koyama, H. The use of embryo transfer to produce pregnancies in repeat-breeding dairy cattle. Theriogenology 2008, 69, 124–128. [Google Scholar] [CrossRef]
- Holm, P.; Booth, P.J.; Callesen, H. Kinetics of early in vitro development of bovine in vivo-and in vitro-derived zygotes produced and/or cultured in chemically defined or serum-containing media. Reproduction 2002, 123, 553–565. [Google Scholar] [CrossRef]
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; De La Fuente, J.; Lonergan, P.; Gutiérrez-Adán, A. Consequences of in vitro culture conditions on embryo development and quality. Reprod. Domest. Anim. 2008, 43, 44–50. [Google Scholar] [CrossRef]
- Sudano, M.J.; Paschoal, D.M.; da Silva Rascado, T.; Magalhães, L.C.O.; Crocomo, L.F.; de Lima-Neto, J.F.; da Cruz Landim-Alvarenga, F. Lipid content and apoptosis of in vitro-produced bovine embryos as determinants of susceptibility to vitrification. Theriogenology 2011, 75, 1211–1220. [Google Scholar] [CrossRef] [Green Version]
- Vajta, G.; Rienzi, L.; Cobo, A.; Yovich, J. Embryo culture: Can we perform better than nature? Reprod. Biomed. Online 2010, 20, 453–469. [Google Scholar] [CrossRef] [Green Version]
- Wydooghe, E.; Heras, S.; Dewulf, J.; Piepers, S.; Van Den Abbeel, E.; De Sutter, P.; Vandaele, L.; Van Soom, A. Replacing serum in culture medium with albumin and insulin, transferrin and selenium is the key to successful bovine embryo development in individual culture. Reprod. Fertil. Dev. 2014, 26, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, M.; Lopera-vasquez, R.; Maillo, V.; Sanchez-Calabuig, M.J.; Núnez, C.; Gutierrez-Adan, A.; Rizos, D. Bovine oviductal and uterine fluid support in vitro embryo development. Reprod. Fertil. Dev. 2018, 30, 935–9345. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, S.; Sánchez Hurtado, M.A.; Gutiérrez, H.; Sánchez Margallo, F.M.; Romar, R.; Latorre, R.; Coy, P.; López Albors, O. Mimicking physiological O2 tension in the female reproductive tract improves assisted reproduction outcomes in pig. Mol. Hum. Reprod. 2018, 24, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, M.; Henning, H.; Costa, P.F.; Malda, J.; Melchels, F.P.; Wubbolts, R.; Stout, T.A.E.; Vos, P.L.A.M.; Gadella, B.M. Improved bovine embryo production in an oviduct-on-a-chip system: Prevention of poly-spermic fertilization and parthenogenic activation. Lab Chip 2017, 17, 905–916. [Google Scholar] [CrossRef] [Green Version]
- Hasler, J.F. In-vitro production of cattle embryos: Problems with pregnancies and parturition. Hum. Reprod. 2000, 15, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschoal, D.M.; Sudano, M.J.; Schwarz, K.R.L.; Maziero, R.R.D.; Guastali, M.D.; Crocomo, L.F.; Magalhães, L.C.O.; Martins, A.; Leal, C.L.V.; da Cruz Landim-Alvarenga, F. Cell apoptosis and lipid content of in vitro–produced, vitrified bovine embryos treated with forskolin. Theriogenology 2017, 87, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef]
- Ferraz, P.A.; Burnley, C.; Karanja, J.; Viera-Neto, A.; Santos, J.E.P.; Chebel, R.C.; Galvão, K.N. Factors affecting the success of a large embryo transfer program in Holstein cattle in a commercial herd in the southeast region of the United States. Theriogenology 2016, 86, 1834–1841. [Google Scholar] [CrossRef]
- Sanches, B.V.; Lunardelli, P.A.; Tannura, J.H.; Cardoso, B.L.; Colombo Pereira, M.H.; Gaitkoski, D.; Basso, A.C.; Arnold, D.R.; Seneda, M.M. A new direct transfer protocol for cryopreserved IVF embryos. Theriogenology 2016, 85, 1147–1151. [Google Scholar] [CrossRef]
- Parrish, J.J. Bovine in vitro fertilization: In vitro oocyte maturation and sperm capacitation with heparin. Theriogenology 2014, 81, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Lopes, J.S.; Canha-Gouveia, A.; París-Oller, E.; Coy, P. Supplementation of bovine follicular fluid during in vitro maturation increases oocyte cumulus expansion, blastocyst developmental kinetics, and blastocyst cell number. Theriogenology 2019, 126, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Bó, G.A.; Mapletoft, R.J. Evaluation and classification of bovine embryos. Anim. Reprod. 2013, 10, 344–348. [Google Scholar]
- Lonergan, P.; Fair, T. The ART of studying early embryo development: Progress and challenges in ruminant embryo culture. Theriogenology 2014, 81, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Sunde, A.; Brison, D.; Dumoulin, J.; Harper, J.; Lundin, K.; Magli, M.C.; Van den Abbeel, E.; Veiga, A. Time to take human embryo culture seriously. Hum. Reprod. 2016, 31, 2174–2182. [Google Scholar] [CrossRef] [PubMed]
- Canovas, S.; Ivanova, E.; Romar, R.; García-Martínez, S.; Soriano-Úbeda, C.; García-Vázquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. eLife 2017, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Block, J.; Bonilla, L.; Hansen, P.J. Efficacy of in vitro embryo transfer in lactating dairy cows using fresh or vitrified embryos produced in a novel embryo culture medium. J. Dairy Sci. 2010, 93, 5234–5242. [Google Scholar] [CrossRef]
- Chebel, R.C.; Demétrio, D.G.B.; Metzger, J. Factors affecting success of embryo collection and transfer in large dairy herds. Theriogenology 2008, 69, 98–106. [Google Scholar] [CrossRef]
- Do, V.H.; Catt, S.; Amaya, G.; Batsiokis, M.; Walton, S.; Taylor-Robinson, A.W. Comparison of pregnancy in cattle when non-vitrified and vitrified in vitro-derived embryos are transferred into recipients. Theriogenology 2018, 120, 105–110. [Google Scholar] [CrossRef]
- Bonilla, L.; Block, J.; Denicol, A.C.; Hansen, P.J. Consequences of transfer of an in vitro-produced embryo for the dam and resultant calf. J. Dairy Sci. 2014, 97, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Lazzari, G.; Wrenzycki, C.; Herrmann, D.; Duchi, R.; Kruip, T.; Niemann, H.; Galli, C. Cellular and molecular deviations in bovine in vitro-produced embryos are related to the large offspring syndrome. Biol. Reprod. 2002, 67, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Stronge, A.J.H.; Sreenan, J.M.; Diskin, M.G.; Mee, J.F.; Kenny, D.A.; Morris, D.G. Post-insemination milk progesterone concentration and embryo survival in dairy cows. Theriogenology 2005, 64, 1212–1224. [Google Scholar] [CrossRef]
- Schrick, F.N.; Inskeep, E.K.; Butcher, R.L. Pregnancy rates for embryos transferred from early postpartum beef cows into recipients with normal estrous cycles. Biol. Reprod. 1993, 49, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Costa, L.L.; Silva, J.R. Plasma progesterone profiles and factors affecting embryo-fetal mortality following embryo transfer in dairy cattle. Theriogenology 2002, 58, 51–59. [Google Scholar] [CrossRef]
- Breukelman, S.P.; Perényi, Z.; Taverne, M.A.M.; Jonker, H.; van der Weijden, G.C.; Vos, P.L.A.M.; de Ruigh, L.; Dieleman, S.J.; Beckers, J.F.; Szenci, O. Characterisation of pregnancy losses after embryo transfer by measuring plasma progesterone and bovine pregnancy-associated glycoprotein-1 concentrations. Vet. J. 2012, 194, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Shorten, P.R.; Ledgard, A.M.; Donnison, M.; Pfeffer, P.L.; McDonald, R.M.; Berg, D.K. A mathematical model of the interaction between bovine blastocyst developmental stage and progesterone-stimulated uterine factors on differential embryonic development observed on Day 15 of gestation. J. Dairy Sci. 2018, 101, 736–751. [Google Scholar] [CrossRef]
- O’Hara, L.; Scully, S.; Maillo, V.; Kelly, A.K.; Duffy, P.; Carter, F.; Forde, N.; Rizos, D.; Lonergan, P. Effect of follicular aspiration just before ovulation on corpus luteum characteristics, circulating progesterone concentrations and uterine receptivity in single-ovulating and superstimulated heifers. Reproduction 2012, 143, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Forde, N.; Lonergan, P. The role of progesterone and conceptus-derived factors in uterine biology during early pregnancy in ruminants. J. Dairy Sci. 2016, 99, 5941–5950. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, L.; Forde, N.; Kelly, A.K.; Lonergan, P. Effect of bovine blastocyst size at embryo transfer on day 7 on conceptus length on day 14: Can supplementary progesterone rescue small embryos? Theriogenology 2014, 81, 1123–1128. [Google Scholar] [CrossRef]
- Clemente, M.; de La Fuente, J.; Fair, T.; Al Naib, A.; Gutierrez-Adan, A.; Roche, J.F.; Rizos, D.; Lonergan, P. Progesterone and conceptus elongation in cattle: A direct effect on the embryo or an indirect effect via the endometrium? Reproduction 2009, 138, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Majewska, M.; Lee, H.Y.; Tasaki, Y.; Acosta, T.J.; Szostek, A.Z.; Siemieniuch, M.; Okuda, K.; Skarzynski, D.J. Is cortisol a modulator of interferon tau action in the endometrium during early pregnancy in cattle? J. Reprod. Immunol. 2012, 93, 82–93. [Google Scholar] [CrossRef]
- Michael, A.E.; Papageorghiou, A.T. Potential significance of physiological and pharmacological glucocorticoids in early pregnancy. Hum. Reprod. Update 2008, 14, 497–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ealy, A.D.; Wooldridge, L.K.; Mccoski, S.R. Post-transfer consequences of in vitro-produced embryos in cattle. J. Anim. Sci. 2019, 30, 2555–2568. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.S.; Bisinotto, R.S.; Lima, F.S.; Greco, L.F.; Morrison, A.; Kumar, A.; Thatcher, W.W.; Santos, J.E.P. Plasma anti-Müllerian hormone in adult dairy cows and associations with fertility. J. Dairy Sci. 2014, 97, 6888–6900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.Y.; Jimenez-Krassel, F.; Steibel, J.P.; Lu, Y.; Baktula, A.; Vukasinovic, N.; Neuder, L.; Ireland, J.L.H.; Ireland, J.J.; Tempelman, R.J. Genomic heritability and genome-wide association analysis of anti-Müllerian hormone in Holstein dairy heifers. J. Dairy Sci. 2018, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stojsin-Carter, A.; Costa, N.N.; De Morais, R.; De Bem, T.H.; Costa, M.P.; Carter, T.F.; Gillis, D.J.; Neal, M.S.; Ohashi, O.M.; Miranda, M.S.; et al. Fetal sex alters maternal anti-Mullerian hormone during pregnancy in cattle. Anim. Reprod. Sci. 2017, 186, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.-R.; Helmer, H.; Behrens, O.; Liu, H.-C.; Antonian, L.; Chang, S.M.; Fields, M.J. Oxytocin and Bovine Parturition: A Steep Rise in Endometrial Oxytocin Receptors Precedes Onset of Labor1. Biol. Reprod. 2005, 47, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Kruip, T.A.M.; Den Daas, J.H.G. In vitro produced and cloned embryos: Effects on pregnancy, parturition and offspring. Theriogenology 1997, 47, 43–52. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Broadbent, P.J.; Dolman, D.F. In vitro produced embryos as a means of achieving pregnancy and improving productivity in beef cows. Anim. Sci. 1995, 60, 55–64. [Google Scholar] [CrossRef]
- Yang, B.; Im, G.; Park, S. Characteristics of Korean native, Hanwoo, calves produced by transfer of in vitro produced embryos. Anim. Reprod. Sci. 2001, 67, 153–158. [Google Scholar] [CrossRef]
- Park, Y.-S.; Kim, S.-S.; Kim, J.-M.; Park, H.-D.; Byun, M.-D. The effects of duration of in vitro maturation of bovine oocytes on subsequent development, quality and transfer of embryos. Theriogenology 2005, 64, 123–134. [Google Scholar] [CrossRef]
- Pimenta-Oliveira, A.; Oliveira-Filho, J.P.; Dias, A.; Gonçalves, R.C. Morbidity-mortality and performance evaluation of Brahman calves from in vitro embryo production. BMC Vet. Res. 2011, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wagtendonk-De Leeuw, A.M.; Aerts, B.J.G.; Den Daas, J.H.G. Abnormal offspring following in vitro production of bovine preimplantation embryos: A field study. Theriogenology 1998, 49, 883–894. [Google Scholar] [CrossRef]
- Siqueira, L.G.B.; Dikmen, S.; Ortega, M.S.; Hansen, P.J. Postnatal phenotype of dairy cows is altered by in vitro embryo production using reverse X-sorted semen. J. Dairy Sci. 2017, 100, 5899–5908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, G.M.; Amer, P.; Stachowicz, K.; Meier, S. Phenotypic associations between gestation length and production, fertility, survival, and calf traits. J. Dairy Sci. 2016, 99, 418–426. [Google Scholar] [CrossRef]
- Goyache, F.; Fernandez, I.; Alvarez, I.; Royo, L.J.; Gutierrez, J.P. Gestation length in the asturiana de los valles beef cattle breed and its relationship with birth weight and calving ease. Arch. Zootec. 2002, 51, 431–439. [Google Scholar]
- Van Wagtendonk-de Leeuw, A.M.; Mullaart, E.; de Roos, A.P.W.; Merton, J.S.; den Daas, J.H.G.; Kemp, B.; de Ruigh, L. Effects of different reproduction techniques: AI, moet or IVP, on health and welfare of bovine offspring. Theriogenology 2000, 53, 575–597. [Google Scholar] [CrossRef]
- Behboodi, E.; Anderson, G.B.; BonDurant, R.H.; Cargill, S.L.; Kreuscher, B.R.; Medrano, J.F.; Murray, J.D. Birth of large calves that developed from in vitro-derived bovine embryos. Theriogenology 1995, 44, 227–232. [Google Scholar] [CrossRef]
- Schmidt, M.; Greve, T.; Avery, B.; Beckers, J.F.; Sulon, J.; Hansen, H.B. Pregnancies, calves and calf viability after transfer of in vitro produced bovine embryos. Theriogenology 1996, 46, 527–539. [Google Scholar] [CrossRef]
- Jacobsen, H.; Schmidt, M.; Holm, P.; Sangild, P.T.; Vajta, G.; Greve, T.; Callesen, H. Body dimensions and birth and organ weights of calves derived from in vitro produced embryos cultured with or without serum and oviduct epithelium cells. Theriogenology 2000, 53, 1761–1769. [Google Scholar] [CrossRef]
- Merton, J.S.; Knijn, H.M.; Flapper, H.; Dotinga, F.; Roelen, B.A.J.; Vos, P.L.A.M.; Mullaart, E. Cysteamine supplementation during invitro maturation of slaughterhouse-and opu-derived bovine oocytes improves embryonic development without affecting cryotolerance, pregnancy rate, and calf characteristics. Theriogenology 2013, 80, 365–371. [Google Scholar] [CrossRef]
- Hasler, J.F.; Henderson, W.B.; Hurtgen, P.J.; Jin, Z.Q.; McCauley, A.D.; Mower, S.A.; Neely, B.; Shuey, L.S.; Stokes, J.E.; Trimmer, S.A. Production, freezing and transfer of bovine IVF embryos and subsequent calving results. Theriogenology 1995, 43, 141–152. [Google Scholar] [CrossRef]
- McEvoy, T.G.; Sinclair, K.D.; Broadbent, P.J.; Goodhand, K.L.; Robinson, J.J. Post-natal growth and development of Simmental calves derived from in vivo or in vitro embryos. Reprod. Fertil. Dev. 1998, 10, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Chavatte-Palmer, P.; Heyman, Y.; Richard, C.; Monget, P.; LeBourhis, D.; Kann, G.; Chilliard, Y.; Vignon, X.; Renard, J.P. Clinical, hormonal, and hematologic characteristics of bovine calves derived FROM nuclei from somatic cells. Biol. Reprod. 2002, 66, 1596–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numabe, T.; Oikawa, T.; Kikuchi, T.; Horiuchi, T. Production efficiency of Japanese black calves by transfer of bovine embryos produced in vitro. Theriogenology 2000, 54, 1409–1420. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Total Presumptive Zygotes | Cleavage Rate | Blastocyst Yield | ||
|---|---|---|---|---|---|
| n | % | n | % | n | |
| BSA | 360 | 85.6 ± 1.9 | 308 | 26.7 ± 2.3 | 96 |
| RF | 429 | 85.6 ± 1.7 | 367 | 25.9 ± 2.1 | 111 |
| Group | Recipients | Day 30 | Day 60 | Day 90 | Day 150 | Day 210 | Parturition | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | n | % | n | % | n | |
| AI | 35 | 22.9 | 8 | 22.9 | 8 | 22.9 | 8 | 22.9 | 8 | 22.9 | 8 | 22.9 | 8 |
| BSA | 45 | 22.2 | 10 | 17.8 | 8 | 17.8 | 8 | 17.8 | 8 | 17.8 | 8 | 17.8 | 8 |
| RF | 54 | 22.2 | 12 | 18.5 | 10 | 18.5 | 10 | 16.7 | 9 | 16.7 | 9 | 16.7 | 9 |
| IVP (BSA + RF) | 99 | 22.2 | 22 | 18.2 | 18 | 18.2 | 18 | 17.3 | 17 | 17.3 | 17 | 17.3 | 17 |
| Group | n | Gestation Length | Minimum | Maximum |
|---|---|---|---|---|
| AI | 8 | 281.1 ± 0.7 | 277 | 284 |
| BSA | 5 | 280.4 ± 1.4 | 275 | 282 |
| RF | 5 | 273.6 ± 9.7 | 240 | 298 * |
| IVP (BSA + RF) | 10 | 277.0 ± 4.8 | 240 | 298 * |
| Group | Parturitions | Induced Parturition | Calving Ease | Male Calves | Neonatal Mortality | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Easy | Difficult | ||||||||||
| n | % | n | % | n | % | n | % | n | % | n | |
| AI | 8 | 0 † | 0 | 100 | 8 | 0 | 0 | 75.0 | 6 | 0 * | 0 |
| BSA | 8 | 37.5 | 3 | 87.5 | 7 | 12.5 | 1 | 62.5 | 5 | 12.5 | 1 |
| RF | 9 | 44.4 | 4 | 77.8 | 7 | 22.2 | 2 | 55.6 | 5 | 44.4 * | 4 |
| IVP (BSA+RF) | 17 | 41.2 † | 7 | 82.4 | 14 | 17.6 | 3 | 58.8 | 10 | 29.4 | 5 |
| Characteristic | Case 1 | Case 2 | Case 3 | Case 4 | Case 5 |
|---|---|---|---|---|---|
| Age at death (days) | 12 | 0 | 2 | 13 | 0 |
| Sex | Female | Male | Female | Male | Female |
| Birth weight (kg) | 21.5 | 56.0 | 43.0 | 52.0 | 56.2 |
| Group | RF | RF | RF | BSA | RF |
| Cause of death | Premature calf ND | Dystocia | Septicaemia | Diarrhoea | Dystocia, spine fracture |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, J.S.; Alcázar-Triviño, E.; Soriano-Úbeda, C.; Hamdi, M.; Cánovas, S.; Rizos, D.; Coy, P. Reproductive Outcomes and Endocrine Profile in Artificially Inseminated versus Embryo Transferred Cows. Animals 2020, 10, 1359. https://doi.org/10.3390/ani10081359
Lopes JS, Alcázar-Triviño E, Soriano-Úbeda C, Hamdi M, Cánovas S, Rizos D, Coy P. Reproductive Outcomes and Endocrine Profile in Artificially Inseminated versus Embryo Transferred Cows. Animals. 2020; 10(8):1359. https://doi.org/10.3390/ani10081359
Chicago/Turabian StyleLopes, Jordana S., Estefanía Alcázar-Triviño, Cristina Soriano-Úbeda, Meriem Hamdi, Sebastian Cánovas, Dimitrios Rizos, and Pilar Coy. 2020. "Reproductive Outcomes and Endocrine Profile in Artificially Inseminated versus Embryo Transferred Cows" Animals 10, no. 8: 1359. https://doi.org/10.3390/ani10081359