
Abstract
High grain yields of upland rice (Oryza sativa L.) can be achieved in no-tillage systems. However, managing nitrogen (N) fertilization for rice in succession to forage grasses is a challenge because forage residues change N cycling and increase microbial immobilization of N, thereby reducing N availability to the subsequent cash crop. In the present study, two field experiments were conducted to determine if applying all or part of the N fertilizer on preceding palisade grass (Urochloa brizantha) and ruzigrass (Urochloa ruziziensis) or their desiccated residues immediately before rice seeding can supply N to the subsequent rice crop. Forage biomass yield (8–16 Mg ha− 1), N accumulation, and N supply to the subsequent upland rice were highest when all of the N fertilizer was applied on forage grasses at 50, 40 or 35 days before rice seeding (DBS), as opposed to the conventional split application at rice seeding and at tillering. On average, the grain yield of upland rice was 54% higher in succession to palisade grass compared with ruzigrass. The grain yield of rice was higher when N was applied to palisade grass at 35 DBS and ruzigrass at 50 DBS, reaching 5.0 Mg ha− 1 and 3.7 Mg ha− 1, respectively. However, applying N to ruzigrass was less effective for increasing upland rice yields since the yields did not differ from the treatments with the conventional split application. Adjusting the time of N application to forage grasses to increase the grain yields of subsequent upland rice is a sustainable alternative that can promote the economic viability of upland rice production.
Similar content being viewed by others


Introduction
Rice is a staple food for more than half of the global population, and world rice production is approximately 750 Tg per year (Faostat 2019). Most of the world’s rice is cultivated in flooded fields, but flooding rice consumes large amounts of water. In addition, the potential emission of greenhouse gases from flooded rice is more than three times higher than from upland crops (Bouman et al. 2007; Carlson et al. 2017). Upland rice cultivation has lower water consumption requirements and is compatible with current agricultural production practices in subsistence crop systems, with few purchased inputs to high-technology systems needed (Kumar and Ladha 2011; Crusciol et al. 2013). Studies have reported grain yields of rainfed upland rice of greater than 3 Mg ha− 1 (Moro et al. 2013; Nascente and Stone 2018). Upland rice cultivation has been used to convert raw land to agriculture, to quickly renew degraded pasture in one growing season, and in rotation with soybean [Glycine max (L.) Merr.] in South America (Santos et al. 2006; Bouman et al. 2007). Moreover, upland rice is a cash crop option for cultivation in crop rotation/succession with maize and soybean that avoids the pathogen cycles associated with other crops. Given these benefits, upland rice cultivation has been gaining in popularity globally but is still considered to provide low economic returns compared with less risky and more profitable crops. The Brazilian grain yield of upland rice is ~ 2.0 Mg ha− 1, approximately 31.5% of lowland rice yield (CONAB 2017).
To make upland rice production viable, alternative approaches such as no-tillage are required. Conservation of residues on the soil surface plays an important role in nutrient cycling in no-tillage systems (Pacheco et al. 2011, 2017; Crusciol et al. 2015). In these systems, the biomass of cover crops cultivated in fall-winter is left to cover and protect the soil and to recycle nutrients back to the soil for the next crop (Calegari et al. 2013; Crusciol et al. 2015; Costa et al. 2016). Forage grasses of the genus Urochloa used as cover crops in no-tillage systems grow fast, produce high biomass yields and have relatively high C/N ratios (Pacheco et al. 2011; Soratto 2011; Costa et al. 2016). These traits help preserve soil moisture and supply nutrients to the following crop over a longer period due to the slow rate of residue decomposition by microorganisms (Ceretta et al. 2002; Bani et al. 2018). Urochloa species have great tolerance to water deficit stress and high capacity to regrow due to their deep root systems (Felismino et al. 2012; Pacheco et al. 2017), thus making them more resistant to adverse conditions during fall and winter than other crops. In addition to improving soil fertility and increasing the yield of the subsequent crop (Crusciol et al. 2015), these perennial forage grasses reduce potential losses from leaching due to their deep roots (Sullivan et al. 2017). Moro et al. (2013) obtained maximum grain yields of 4.8–6.8 Mg ha− 1 for rainfed upland rice grown in succession to Urochloa grasses.
Nitrogen is a yield-limiting nutrient in upland rice production. N increases shoot dry matter, the grain harvest index, and components that are positively related to grain yield (Fageria 2007; Fageria et al. 2010). Recent studies have demonstrated that upland rice grain yield responds to N fertilization rates higher than 100 kg N ha− 1, especially when grown in no-tillage systems following forage grasses (Guimarães and Stone 2003; Lopes et al. 2013). However, forage grasses can also compromise the synchronization between N available in the soil and upland rice N uptake and yields by increasing N immobilization before the cash crop is planted, especially in systems with forage grasses with high C/N ratios (Guimarães and Stone 2003; Cantarella 2007; Rosolem et al. 2017).
The conventional recommendation for N fertilization of upland rice is application of 20% of N at seeding plus 80% of N sidedressed at tillering (Cantarella et al. 1997). These recommendations are based on conventional soil tillage and excessive N supply at the beginning of the upland rice cycle (seeding to tillering), which can cause N lodging in the plant. However, little is known about efficient N management for upland rice in no-tillage systems with large residues (Kluthcouski and Stone 2003; Nascente et al. 2013). Previous studies have found no clear tendency of the response to N application in the early growth stage of rice, as both increases in yield (Guimarães and Stones 2003) and no effects on yield (Nascente et al. 2011) have been reported for N application at upland rice seeding in succession to pasture in no-tillage systems.
Studies of N application after seeding in the early growth stages of the upland rice cycle in different regions have shown both grain yield and economic gains (Kluthcouski et al. 2006). Applying N fertilizer to the preceding forage grasses may be an efficient method for providing a much-needed increase in cover crop biomass in no-tillage systems. In addition, N application on forage grasses may facilitate the synchronization of N release from forage decomposition with the period of highest demand by the succeeding upland rice crop (Rosolem et al. 2004, 2017). Because of their deep root systems, forage grasses may improve N cycling from the soil in agricultural systems and thus reduce excessive N losses in complex plant-straw-soil systems (Crusciol et al. 2014; Galdos et al. 2020). In addition, applying N fertilizer to the previous forage grass could avoid damage to the cash crop during the growth stage due to mechanical operations and save fuel and labor costs (Kluthcouski et al. 2006; Soratto 2011).
A previous study showed that applying N fertilizer to forage grasses grown before maize is impracticable because the resulting N immobilization compromises N supply early in the maize crop cycle (Momesso et al. 2019). However, the N uptake and N-use efficiency of upland rice differ from those of maize due to the lower N fertilizer rate and N demand of the former (Makino et al. 2003). Thus, this study tests two hypotheses in successive experiments: (1) applying N on forage grasses preceding the rice crop is an alternative system of food production that enhances rice yields; (2) applying part of the N fertilizer on forage grasses and part at rice seeding can avoid N immobilization early in the rice cycle and enhance rice yields. To test the above hypotheses, the effects of applying all or part of the N fertilizer on preceding palisade grass (Urochloa brizantha) and ruzigrass (Urochloa ruziziensis) or on their desiccated residues immediately before rice seeding on N supply and upland rice yields were compared with those of conventional N fertilization.
Materials and methods
Site description, experimental design and crop management
Two field experiments were conducted in Botucatu in São Paulo State, southeastern Brazil (48° 26′ W, 22° 51′ S, 740 m). Experiment 1 (E1) was carried out in two growing seasons (2014–2015 and 2015–2016), and experiment 2 (E2) was carried out in the 2015–2016 growing season (Supplementary Fig. 1). The climate is Cwa according to the Köppen climate classification system, i.e., tropical with dry winters and hot, rainy summers (Alvares et al. 2013). Precipitation and average temperatures were measured during the experiments (Supplementary Fig. 2). The soil was a clayey, kaolinitic, thermic Typic Haplorthox (USDA 2014) with 630, 90, and 280 g kg− 1 of clay, silt, and sand, respectively. The chemical characteristics of the topsoil (0–0.20 m) were determined before initiating the experiments in each of the growing seasons and are shown in Supplementary Table 1.
The areas were managed in a no-tillage system for seven years prior to the start of the experiments. Both experiments were set up in areas previously cultivated with palisade grass (U. brizantha cv. Marandu) and ruzigrass (U. ruziziensis) under no-tillage before upland rice seeding. The historical crop rotation is shown in Supplementary Table 2. Palisade grass and ruzigrass were seeded at a density of 10 kg seed ha− 1 (34% viable seed) and were not previously fertilized, i.e., they grew only with residual fertilization from a previous crop. The forage grasses were cultivated for approximately 20 months before upland rice seeding, and the pastures were managed without weed control and mowed three times a year. In 2014 and 2015, plant material 0.30 m above soil level was mechanically mowed before experiment installation in order to stimulate growth and N uptake by the forage grasses in both experiments.
We set up two experiments to evaluate the effect of applying all (E1) or part (E2) of the N rate on forage grasses or their residues (Supplementary Fig. 1). The experimental designs were a completely randomized block with four replications. The field plots consisted of 10 rows with a length of 6 m and spacing of 0.34 m and a total plot area of 20.4 m2. A schematic diagram of N application timing in E1 and E2 is presented in Supplementary Fig. 1.
E1 was arranged in a 2 × 6 factorial scheme over two growing seasons. The plots were planted with a combination of two forage species {{palisade grass [(Urochloa brizantha (Hochst. Ex A. Rich.) R.D. Webster] and ruzigrass (U. ruziziensis R. Germ. and C.M. Evrard) Morrone and Zuloaga}} with six N management strategies (Supplementary Fig. 1): (1) control (no N application), (2) conventional method 1 (20 kg N ha− 1 applied in the upland rice seeding furrow plus 80 kg N ha− 1 sidedressed at the rice tillering stage), (3) N applied on live forage grass 50 days before seeding (DBS) of upland rice, (4) N applied on live forage grass 40 DBS of upland rice, (5) N applied on live forage grass 35 DBS of upland rice, and (6) N applied on forage grass residue 1 DBS of upland rice (pre-seeding of upland rice). The total N fertilizer rate was 100 kg N ha− 1 in all treatments except for the control. In all treatments, ammonium nitrate was used as the N source.
In E2, a 2 × 4 factorial scheme was used. The plots were composed of the same two forage grasses as in E1 and four N management strategies (Supplementary Fig. 1): (1) control (no N application), (2) conventional method 2 (100 kg N ha− 1 sidedressed at upland rice tillering stage), (3) N applied on live forage grass 35 DBS of upland rice, and (4) N applied on live forage grass residue 1 DBS of upland rice (pre-seeding of upland rice)]. Except for the control, the treatments received 100 kg N ha− 1 at the different application times plus 20 kg N ha− 1 at rice seeding, giving a total rate of 120 kg N ha− 1. Ammonium sulfate was used as the N source for all N treatments.
In all treatments, forage species terminations was performed at 30 DBS of upland rice by spraying 1.56 kg glyphosate ha− 1 (active ingredient). For the treatments with N application 1 DBS of upland rice (pre-seeding), N was applied on the desiccated forage grass residue in December.
The upland rice was seeded using a no-tillage drill at a density of 70 seeds m− 1. In E1, the rice cultivar BRS Monarca was used in both growing seasons. This cultivar has an intermediate cycle (112 days) and long grains and performs well in several management systems. The recently released upland rice cultivar ANa 5015 was used in E2. This cultivar has a shorter cycle (92 days), is classified as an intermediate type with long grains, and has the potential to produce 5 Mg ha− 1. The basic fertilization in the rice seeding furrows consisted of 50 kg P2O5 ha− 1 as triple superphosphate and 50 kg K2O ha− 1 as potassium chloride.
For the conventional N fertilization, in E1 20 kg N ha− 1 was band-applied 0.05 to 0.10 m from the seed line on the day of seeding and complemented with 80 kg N ha− 1 sidedressed at the rice tillering stage. In E2, all N treatments received 20 kg N ha− 1 band-applied between 0.05 and 0.10 m from the seed line on the day of upland rice seeding. For the conventional method in E2, an additional 100 kg N ha− 1 was band-applied at the tillering stage. Therefore, the treatments in E1 and E2 differed in (1) time of N application; (2) the application of 20 kg N ha− 1 at seeding in all treatments in E2 (only the conventional methods received N at seeding in E1); (3) the N rate, which was 100 kg ha− 1 in E1 and 120 kg ha− 1 in E2; (4) the use of ammonium nitrate as the N source in E1 but ammonium sulfate in E2.
Sampling, measurement, and analyses
The dry mass yield of the forage grasses was evaluated on the day of termination and 90 days later in order to determine the initial and remaining amounts of dry matter biomass above the soil surface. Three samples per plot of aboveground plant material were collected using a wooden frame with an internal area of 0.25 m2 on each date to obtain a composite sample. Sampling was performed within the plots at random points along diagonal crosswise lines. The collected biomass was dried by forced-air circulation at 65 ºC for 72 h and weighed, and the data were extrapolated to Mg ha− 1 for dry matter determination. The plant samples were ground to pass through a 0.85-mm stainless-steel sieve and analyzed to determine the N concentration.
Rice leaf samples were collected for nutrient concentration determination when 50% of the panicles in each plot were at the flowering stage. The flag leaves of 20 randomly chosen plants per plot were collected following the procedure described by Cantarella et al. (1997). The leaf samples were dried by forced-air circulation at 65 °C for 72 h, ground to pass through a 0.85-mm stainless-steel sieve, and analyzed to determine nutrient (N, P, K, Ca, Mg, and S) concentrations.
The samples of forage grass residues and upland rice leaves were digested with sulfuric acid for N determination and with nitro-perchloric solution for the other nutrients. The N, P, and S concentrations were determined from the digested solutions by semi-micro-Kjeldahl distillation, colorimetry, and turbidimetry methods, respectively. The K, Ca, and Mg concentrations were determined by atomic absorption spectrophotometry (Malavolta et al. 1997).
On the same dates as rice leaf sampling, shoots of plants from 1.0-m lengths of the two central rows were cut at the soil level to determine aboveground dry matter. The plant shoots were dried by forced-air circulation at 65 °C for 72 h and weighed, and the data were extrapolated to Mg ha− 1. The following parameters were evaluated 85 and 108 days after emergence in E2 and E1, respectively: plant height, number of panicles per m2, total number of spikelets per panicles, spikelet fertility, and 1000-grain weight. The plants were manually harvested, and the panicles from the usable areas of the plots were collected and dried in the sun for 1 to 2 days before mechanical threshing using a research plot thresher. The grain yield (unhulled grain weight, moisture content corrected to 130 g kg− 1 and conversion to Mg ha− 1) was determined. The N-use efficiency (NUE) was defined as the increase in grain yield per unit of N applied (Fageria and Baligar 2005) and was determined by dividing the difference between the grain yield (kg ha− 1) in each N treatment and the grain yield of the control (no N application) by the applied N rate (kg ha− 1).
Statistical analyses
For all variables, analysis of variance and F probability tests were performed using the SISVAR statistical software package (Ferreira 2011). First, all data were initially tested for normality using the Shapiro-Wilk test procedure and were distributed normally (W ≥ 0.90). Then, the data were submitted to ANOVA to determine the effect of forage grass and N application timing on biomass and N accumulated of palisade grass and ruzigrass, and rice compounds, grain yields and NUE of rice. For E1, forage grass species, N application timing, and growing season were considered fixed effects. For E2, forage grass species and N application timing were considered fixed effects. The blocks and all block interactions were considered random effects. A comparison of means was performed with the LSD test (P ≤ 0.05) when the F-test was significant. To estimate the relationship between rice yield and forage grass biomass (on the day of termination), the linear components were tested with the whole data sets of E1 and E2; to estimate the relationship between rice yield and N released by grass residues from 0 to 90 days after termination, the linear components were tested with the data sets of E1 and E2 split according to forage grass species (Mead and Drasgow 1993).
Results
Forage grass biomass yield and N release
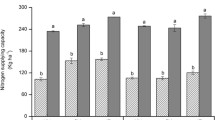
The interaction of forage grass species and time of N application significantly influenced dry matter yield and N accumulation by forage grasses in both E1 and E2 (Figs. 1 and 2). On the day of termination, forage grass dry matter was highest for early N application on live palisade grass (50 DBS and 35 DBS in E1 and E2, respectively) (Figs. 1 and 2a). In E1, compared with the control, palisade grass dry matter was 39% higher when N was applied 50 DBS (11.5 Mg ha− 1) and decreased in the treatments in which N was applied to live forage grasses closer to rice seeding (40 DBS and 35 DBS). The dry matter results in E2 followed similar trends. N application to live palisade grass 35 DBS resulted in an increase in yield compared with the treatments where N was applied over forage residues; at the other times of N application (1 DBS and conventional), the yields did not differ from the control (Fig. 2a). However, even for the unfertilized forage grasses (controls), the dry matter yield of palisade grass at termination (9 Mg ha− 1) was 33% greater than that of ruzigrass in both experiments.

Forage grass × N application timing interaction effect on the amount of dry matter (a, b) and N (c, d) in shoot residues of forage grasses at 0 (a,c) and 90 (b, d) days after termination (DAT) in experiment 1. Average of 2014–2015 and 2015–2016 growing seasons. Treatments are described in Table 1. Error bars are one standard error from the mean. Different lowercase letters denote significant differences between forage grasses and different uppercase letters denote significant differences among N application systems (LSD, P ≤ 0.05)

Forage grass × N application timing interaction effect on the amount of dry matter (a, b) and N (c, d) in shoot residues of forage grasses at 0 (a, c) and 90 (b, d) days after termination (DAT) in experiment 2 (2015–2016 growing season). Treatments are described in Table 1. Error bars are one standard error from the mean. Different lowercase letters denote significant differences between forage grasses and different uppercase letters denote significant differences among N application systems (LSD, P ≤ 0.05)
At 90 DAT, the remaining dry matter of the forage grasses decreased sharply but followed the trends of the sampling performed immediately after termination (0 DAT). For palisade grass, the values of remaining biomass were highest when N was applied early on live forage (50 DBS and 35 DBS in E1 and E2, respectively) (Figs. 1b and 2b). In addition, in E1, remaining dry matter was higher when palisade grass received N 10 days before forage termination (40 DBS) than when N was applied 5 days before termination (35 DBS). For ruzigrass in both experiments, all treatments in which N fertilizer was applied to live forage grasses resulted in similar remaining dry matter (3.5 Mg ha− 1 average), while remaining dry matter was lower in the treatments in which N was applied over ruzigrass residues (Fig. 1b).
In general, the earlier that N was applied to palisade grass, the higher the value of N accumulation at termination in both experiments (Figs. 1 and 2c). In E1, N accumulation in palisade grass and ruzigrass was highest with the earliest N application (50 DBS), with values of 288 and 131 kg ha− 1, respectively (Fig. 1c). In E2, N accumulation was highest when N was applied at 35 DBS (Fig. 2c). At 90 DAT, remaining N in the forage grass residue was highest in the treatment receiving N on palisade grass 50 DBS in E1 and 35 DBS in E2 (Figs. 1d and 2d), similar to the trends for remaining dry matter.
Nutrient concentration in upland rice leaves
Forage grass had little influence on the leaf N concentration of upland rice (Supplementary Table 3). In both experiments, the leaf N concentrations in upland rice leaves were higher in succession to palisade grass compared with ruzigrass and in all treatments with N application compared to the control. No significant influence of the treatments on leaf concentrations of P, K, Ca, Mg, and S was observed in either E1 or E2 (Supplementary Table 3).
Plant growth, yield components, and grain yield of upland rice in experiment 1
In E1, the timing of N application significantly influenced the plant height of upland rice (Table 1). The plant height increased from 0.70 m in the control to an average of 0.80 m in the 40 DBS, 35 DBS, and 1 DBS treatments.
The interaction of forage grass species and N application timing had a significant influence on shoot dry matter, number of panicles per m2 and spikelets per panicle (Table 1; Fig. 3a). On average, the shoot dry matter yield of rice in succession to palisade grass was 55% higher when N was applied closer to the day of forage termination (35 DBS and 40 DBS) compared with the control. For rice in succession to ruzigrass, shoot dry matter yield was higher in the treatment with N applied at 50 DBS (7.8 Mg ha− 1) than in the other N treatments (4.0 Mg ha− 1). Nitrogen application on live forage grasses also increased the number of panicles per m2 (Fig. 3b). Rice in succession to palisade grass and ruzigrass receiving N fertilizer at 50, 40, 35 and 1 DBS had a higher number of panicles per m2 than rice in the treatments with conventional N application; however, the increase in the number of panicles per m2 of rice was highest for rice in succession to palisade grass. The trend for the number of spikelets per panicle across all N applications was similar to that for shoot dry matter yield of upland rice, i.e., higher when N fertilizer was applied on palisade grass at 35 DBS and 40 DBS (Fig. 3c).

Forage grass × N application timing interaction effect on rice shoot dry matter (a), number of panicles per m2 (b), number of spikelets per panicle (c), grain yield (d), and N-use efficiency (e) of upland rice in experiment 1. Average of 2014–2015 and 2015–2016 growing seasons. Treatments are described in Table 1. Error bars are one standard error from the mean. Different lowercase letters denote significant differences between forage grasses and different uppercase letters denote significant differences among N application systems (LSD, P ≤ 0.05)
Both the percentage of spikelet fertility and 1000-grain weight were lower in rice in succession to ruzigrass than in rice in succession to palisade grass, with respective values of 77.7% and 80.7% and 21.5 and 23.1 g (Table 1). Regardless of the forage grass, zero-N application resulted in 5% lower spikelet fertility compared with N fertilizer application on forage grass and its residues (50 DBS, 40 DBS, 35 DBS, and 1 DBS). A similar trend was observed for 1000-grain weight. Zero-N application (control) decreased 1000-grain weight by 8.7% compared with the treatments receiving N fertilizer.
The interaction of forage grass species and N application timing also significantly influenced rice grain yield and NUE (Table 1; Fig. 3). Rice grain yields were higher in succession to palisade grass compared with ruzigrass under every N application timing treatment, but the NUE of rice in succession to palisade grass was higher only when N was applied at 40, 35 and 1 DBS (Fig. 3d, e). For rice in succession to palisade grass, the rice grain yield was highest when N was applied 35 DBS and 40 DBS and decreased in the other treatments in the following order: 50 DBS, 1 DBS, conventional and control (Figs. 3 and 5). However, the NUE of rice was highest when cultivated in succession to palisade grass receiving N fertilizer at 35 DBS. For rice in succession to ruzigrass, the highest rice grain yield (3.7 Mg ha− 1) was obtained when N was applied at 50 DBS and was 131% higher than that in the control (1.6 Mg ha− 1). In addition, the grain yields of rice in succession to palisade grass and ruzigrass were higher under the conventional method of N application (in the seeding furrow plus sidedressing at tillering of upland rice) than in the control.
Plant growth, yield components, and grain yield of upland rice in experiment 2
In E2, the forage grass species affected rice plant height (Table 1). Upland rice plants were 17% taller when cultivated in succession to palisade grass compared to ruzigrass. In addition, all treatments with N fertilizer resulted in taller plants than the control, but there was no difference in plant height between N application strategies.
The shoot dry matter yield, yield components, grain yield, and NUE of upland rice were affected by the forage grass species and N application interaction (Table 1; Fig. 4). The shoot dry matter and yield components (panicle per m2, spikelets per panicle and 1000-grain weights) of rice were highest when N fertilizer was applied on live palisade grass (35 DBS) and its residues (1 DBS), followed by the treatments with conventional N application and the controls without N fertilizer (Fig. 4a). By contrast, the shoot dry matter and yield components of rice cultivated in succession to ruzigrass were similar among the treatments that received N fertilizer (35 DBS, 1 DBS and conventional). Thus, while N fertilization of both palisade grass and ruzigrass increased the rice response compared with the control, cultivation in succession to palisade grass resulted in the greatest shoot dry matter and yield components.

Forage grass × N application timing interaction effect on the shoot dry matter (a), number of panicles per m2 (b), number of spikelets per panicle (c), spikelet fertility (d), 1000-grain weight (e), grain yield (f), and N-use efficiency (g) of upland rice in experiment 2 (2015–2016 growing season). Treatments are described in Table 1. Different lowercase letters denote significant differences between forage grasses and different uppercase letters denote significant differences among N application systems (LSD, P ≤ 0.05)

Relationships between rice yield and forage grass biomass (n = 128) on the day of termination (a), and rice yield and N released by grass residues (n = 64) for N application timing on each forage grass (b) in experiment 1 (E1) and experiment 2 (E2). The black solid line in (a) means linear model for both forage grasses. The blue spots and solid line mean palisade grass (Urochloa brizantha) values, and the black spots and dashed line mean ruzigrass (U. ruziziensis) values in (b). The description of treatments is in Table 1
Similar to the trend of shoot dry matter of rice following ruzigrass, N fertilization increased spikelet fertility compared with the control in succession to both forages (Fig. 4d); however, when no N was applied (control), spikelet fertility was more than double in succession to palisade grass compared to ruzigrass.
The grain yield and NUE of rice differed significantly among the N treatments in succession to palisade grass (Figs. 4f and 5). The grain yields following palisade grass were 4.6, 4.1, 3.6, and 2.3 Mg ha− 1 in the 35 DBS, 1 DBS, conventional, and control treatments, respectively; by contrast, the grain yield of rice following ruzigrass with N fertilization was similar in all N management strategies (2.4–2.6 Mg ha− 1) and higher than in the control (1.1 Mg ha− 1) (Figs. 4 and 5). The NUE of upland rice was larger the earlier that N was applied for rice in succession to palisade grass but was similar for all times of N application for rice in succession to ruzigrass (Fig. 4 g). Regardless of N treatment, higher upland rice grain yields and NUE values were obtained in succession to palisade grass (3.6 Mg ha− 1 and 19 kg grain kg− 1 N) relative to ruzigrass (2.1 Mg ha− 1 and 6 kg grain kg− 1 N).
Discussion
Effect of nitrogen application on forage grasses
In this study of two growing seasons, earlier N application on forage species increased forage grass residues. The highest biomass yields of forage grass were achieved with N application at 50 and 35 DBS of upland rice in E1 and E2, respectively. Early N application on forage grasses stimulated the growth of the grasses in the period between N fertilization and forage grass termination when the weather conditions were favorable. However, at termination, there was no significant difference in biomass yield between N application on forage grasses at 40 and 35 DBS, probably because the intervening period of ~ 5 days was too short to affect plant growth. When N was not applied, palisade grass and ruzigrass produced at least 11.5 and 6.9 Mg ha− 1 of dry matter biomass in E1 and 12.2 and 8.5 Mg ha− 1 of dry matter biomass in E2, which is considered appropriate to provide mulch for a no-tillage system (Kluthcouski and Stone 2003). High biomass yields of grass cover crops benefit the agricultural system by suppressing weeds and allowing herbicide application to be reduced or eliminated (Fageria et al. 2005; Sao Miguel et al. 2018). In addition, greater cover crop biomass helps to preserve soil moisture, which increases microbial activity and improves plant nutrition (Chavarria et al. 2016; Verzeaux et al. 2017). The high yields in this study reflect the high fertility of the soil, which has been cultivated under no-tillage for many years.
The biomass yield of palisade grass was higher than that of ruzigrass in all N treatments due to differences in the characteristics of the two species. Some studies have shown that, compared to ruzigrass, palisade grass has higher tolerance to soil acidy and high levels of soil aluminum; higher biomass yield (Alvim et al. 1990; Timossi et al. 2007); and a higher capacity of regrowth and straw persistence in the soil (Felismino et al. 2012). The higher biomass yield of palisade grass led to greater N accumulation at termination due to the faster growth and higher NUE of palisade grass. Contrary to the results of the present study, Pacheco et al. (2011) and Momesso et al. (2019) found similar N accumulation in the two species despite the higher shoot dry matter yield of palisade grass.
Based on the initial biomass and N accumulated in the forage at termination, we measured the loss of mass and of N in the plant dry matter after 90 days. Sharp decreases in both biomass and N content were observed. Part or most of the N loss from the grasses was probably transferred to the soil. During the 90 days after termination, on average 6–12 Mg ha− 1 of biomass and 60–200 kg ha− 1 N accumulated in forage grasses were released from the cover crops to the system. The release of N by the grasses may have increased the soil N available to the crop in succession, especially in the plots with forage grasses fertilized with N (from 120 to 200 kg N ha− 1 released). These results indicate that palisade grass and ruzigrass have the potential to recycle and release a high amount of N into the agricultural system.
In addition to the loss of plant mass and N, palisade grass residue released greater amounts of N than ruzigrass residue during the 90 days after termination. In the treatments with N applied on live palisade grass residue (50 DBS, 40 DBS, and 35 DBS), an average of 130 kg N ha− 1 was released from the residues. Applying nitrogen fertilizer to forage grasses before termination may affect biomass yields as well as chemical composition, including the proportions of cellulose, hemicellulose, and lignin. The chemical composition of forage grass influences the rate of decomposition, which is carried out by microorganisms in two main steps: degradation of cellulose and hemicellulose followed by degradation of lignin (Bani et al. 2018). Ruzigrass undergoes a period of fast decomposition followed by a second period of slow decomposition (Pacheco et al. 2017). Hence, the much lower biomass of ruzigrass at the two sampling times can be explained by the differences in the characteristics of the two plant species.
Higher yields of upland rice related to nitrogen management and forage grasses
The macronutrient concentrations in upland rice leaves were within adequate ranges for rice crops (Cantarella et al. 1997). We aimed to increase forage grass biomass yields in order to release N for the subsequent upland rice crop. Nutrient cycling by forage grasses can help supply the nutrients required by upland rice, as observed by Soratto (2011), Nascente and Stone (2018) and Momesso et al. (2019). In both experiments, leaf N concentrations in upland rice leaves were lower in the control than in all treatments with N application; however, even in the control (no N application), the upland rice plants were properly nourished, probably due to the nutrients released by the decomposition of the forage grass residues, since the biomass yields of the control treatments were also high.
Applying N fertilizer at rice seeding resulted in lower rice yields than earlier N applications on forage grasses. When N fertilizer was applied on palisade grass at 40 and 35 DBS in E1, the forage grass produced approximately 14 Mg ha− 1 of shoot dry matter and 200 kg ha− 1 of accumulated N. These residues and N accumulation promoted higher shoot dry matter and yields of upland rice compared with the conventional method of N application and the control. In contrast to our findings, a study on the same farm in which all N fertilizer was applied on grasses reported a lower grain yield of subsequent maize compared with the conventional recommended N application at seeding and in the V4 to V6 growth stage of maize (Momesso et al. 2019). Apparently, the response of the subsequent crop to N fertilization on forage grass is dependent on the crop species and its response to N.
The aim of applying N on forage grasses or its residues prior to rice seeding was to increase the flexibility of fertilization timing and permit N release to the subsequent crop by straw decomposition. Indeed, the strategy of applying N to forage grasses was efficient for achieving high grain yields of upland rice. A possible explanation for this positive effect is that forage grasses can efficiently absorb N from fertilizer and gradually release N via the slow decomposition of biomass, thus avoiding N losses. Studies have shown that N fertilization accelerates the rate of decomposition of plant residues (Parton et al. 1988; Rosolem et al. 2004, 2017) by reducing the C/N ratio of the cover crop. Therefore, the more gradual release of the N retained by the grasses when N was applied earlier may have benefited rice nutrition compared to the pulse of N supplied by the conventional method of fertilization. The synchronization between soil inorganic N supply and crop demand may be attributable to the microbial decomposition of straw residues (Cantarella 2007; Guo et al. 2018; Fan et al. 2019).
In E2, we also tested the effect of start fertilization by applying 20 kg N ha− 1 in the treatments in which most of the N fertilizer was applied to the forage before rice sowing. The premise was that start fertilization would decrease the risk of N immobilization and competition between plants and microorganisms (Kuzyakov and Xu 2013; Bani et al. 2018). However, no effect of start fertilization was observed, as there was no difference in rice yields between the treatments with N applied at 35 DBS and 1 DBS.
Our finding of reduced rice grain yield under the conventional method of application was somewhat unexpected since the conventional split application of N aims to supply N in the rice growth stage with high N demand, thereby increasing nutrient uptake by plants and reducing losses by NO3− leaching, NH3 volatilization and N2O emissions (Rao and Prasad 1980; Cantarella 2007). However, the second split application occurred at the time of highest rainfall, which may increase the risk of losses of N to leaching. Our data support a role of NO3− leaching by rainfall in the insufficient N for grain yields since applying N fertilizer at rice tillering (conventional method) resulted in lower nitrogen recovery by rice. In addition, these data highlight that early N application can also be advantageous by increasing the flexibility of N application timing to increase rice yields and NUE.
In addition, the effect of mulching on rice plants may be more relevant than previously thought. In our study, all treatments comparing the time of N application were fertilized with 100 or 120 kg N ha− 1; thus, even if some losses occurred in some treatments, the N applied should be enough to sustain yields of 4 to 5 Mg ha− 1 (Cantarella et al. 1997). There was a significant relationship between the cover crop biomass and rice grain yield; for the same amounts of N released by grass decomposition, yields were higher in succession to palisade grass than ruzigrass. In addition, rice grain yields were higher when N was applied earlier to the grass, consistent with the increased grass and hence mulch production.
Rice has a shallow root system compared with crops such as maize (Fageria and Moreira 2011) and may be more responsive to mulching, which helps to preserve soil water, reduce soil temperature close to the soil surface, and stimulate microbial activity, improving conditions for root growth. Mulching may also help alleviate stress caused by poor soil physical conditions. Rice responds to N fertilizer application under both no-tillage and conventional tillage management, but yields tend to be lower under no-tillage (Stone et al. 1980; Kluthcouski et al. 2000; Nascente et al. 2011), possibly due to the greater effect of soil compaction on crops with shallow root systems.
Conclusions
Here we showed that the strategy of N application in advance of upland rice seeding does not compromise rice yield. Nitrogen application on palisade grass 35 days before rice seeding and on its residues just before rice seeding represents an alternative N application timing for enhancing grain yields in tropical no-tillage systems. Furthermore, it is clear that mulching plays an important role in the response of upland rice to N. These are new options for managing fertilization to sustainably increase rice yields in tropical environments, where soil cover and no-tillage are fundamental for soil preservation. Relevant questions about which microbial processes are the main drivers of available N in soil and why a seemingly risky fertilization practice, such as applying N before seeding the main crop, worked well in a region with high precipitation deserve further investigations.
Abbreviations
- C:
-
Carbon
- Ca:
-
Calcium
- DBS:
-
Days before seeding
- K:
-
Potassium
- Mg:
-
Magnesium
- N:
-
Nitrogen
- NUE:
-
N-use efficiency
- P:
-
Phosphorus
- SOM:
-
Soil organic matter
- S:
-
Sulfur
References
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorol Z 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
Alvim MJ, Botrel MA, Verneque RS, Salvati JA (1990) Nitrogen application in access to Brachiaria. 1- Dry matter production effect. Pasturas Tropicales 2:2–6
Bani A, Pioli S, Ventura M, Panzacchi P, Borrusa L, Tognetti R, Tonon G, Brusetti L (2018) The role of microbial community in the decomposition of leaf litter and deadwood. Appl Soil Ecol 126:75–84. https://doi.org/10.1016/j.apsoil.2018.02.017
Bouman BAM, Lampayan RM, Tuong TP (2007) Water management in irrigated rice: coping with water scarcity. International Rice Research Institute, Los Baños
Calegari A, Tiecher T, Hargrove WL, Ralisch R, Tessier D, Tourdonnet S, Guimarães F, Santos DR (2013) Long-term effect of different soil management systems and winter crops on soil acidity and vertical distribution of nutrients in a Brazilian Oxisol. Soil Till Res 133:32–39. https://doi.org/10.1016/j.still.2013.05.009
Cantarella H, van Raij B, Camargo CEO (1997) Cereals. In: van Raij B, Cantarella H, Quaggio JA, Furlani ACM (eds) Lime and fertilizer recommendations for the State of Sao Paulo, 2nd edn. Agronomic Institute of Campinas, Sao Paulo
Cantarella H (2007) Nitrogen. In: Novais RF, Alvarez VH, Barros NF, Fontes RL, Cantarutti RB, Neves JCL (eds) Soil fertility. Sociedade Brasileira de Ciência do Solo, Viçosa
Carlson KM, Gerber JS, Mueller ND, Herrero M, MacDonald GK, Brauman KA, Havlik P, O’Connell CS, Johnson JA, Saatchi S, West PC (2017) Greenhouse gas emissions intensity of global croplands. Nat Clim Change 7:63–68. https://doi.org/10.1038/nclimate3158
Castro AP, de Morais OP, Breseghello F (2007) BRS Monarca: Monarca: upland rice grown with excellence in grain quality. http://agris.fao.org/agris-search/search.do?recordID=BR20071216445. Accessed 11 Nov 2018
Ceretta CA, Basso CJ, Herbes MG, Poletto N, Silveira MJ (2002) Winter cover crops and corn biomass production and decomposition, under different nitrogen fertilization managements. Cienc Rural 32:49–54. https://doi.org/10.1590/S0103-84782002000100009
Chavarria DN, Verdenelli RA, Serri DL, Restovich SB, Andriulo AE, Meriles JM, Vargas-Gil S (2016) Effect of cover crops on microbial community structure and related enzyme activities and macronutrient availability. Eur J Soil Biol 76:74–82. https://doi.org/10.1016/j.ejsobi.2016.07.002
Companhia Nacional de Abastecimento (CONAB) (2017) 8th Survey – 2015–16 harvest. Harvest surveys. (In Portuguese.) Conselho Nacl. Abastecimento, Brasilia, Distrito Federal, Brazil. http://www.conab.gov.br/OlalaCMS/uploads/arquivos/17_04_17_17_52420_55_boletim_graos_abr2017.pdf. Accessed 3 Feb 2018
Costa CHM, Crusciol CAC, Soratto RP, Ferrari Neto J, Moro E (2016) Nitrogen fertilization on palisadegrass: phytomass decomposition and nutrients release. Pesqui Agropecu Trop 46:159–168. https://doi.org/10.1590/1983-40632016v4639297
Crusciol CAC, Nascente AS, Borghi E, Soratto RP, Martins PO (2015) Improving soil fertility and crop yield in a tropical region with palisadegrass cover crops. Agron J 107:2271–2280. https://doi.org/10.2134/agronj14.0603
Crusciol CAC, Nascente AS, Mateus GP, Pariz CM, Martins PO, Borghi E (2014) Intercropping soybean and palisade grass for enhanced land use efficiency and revenue in a no till system. Eur J Agron 58:53–62. https://doi.org/10.1016/j.eja.2014.05.001
Crusciol CAC, Soratto RP, Nascente AS, Arf O (2013) Root distribution, nutrient uptake, and yield of two upland rice cultivars under two water regimes. Agron J 105:237–247. https://doi.org/10.2134/agronj2012.0298
Fageria NK, Baligar VC (2005) Enhancing nitrogen use efficiency in crop plants. Adv Agron 88:97–185. https://doi.org/10.1016/S0065-2113(05)88004-6
Fageria NK (2007) Yield physiology of rice. J Plant Nutr 30:843–879. https://doi.org/10.1080/15226510701374831
Fageria NK, Morais OP, Santos AB (2010) Nitrogen use efficiency in upland rice genotypes. J Plant Nutr 33:1–11. https://doi.org/10.1080/01904167.2010.496892
Fageria NK, Baligar VC, Bailey BA (2005) Role of cover crops in improving soil and row crop productivity. Commun Soil Sci Plan 36:2733–2757. https://doi.org/10.1080/00103620500303939
Fageria NK, Moreira A (2011) The role of mineral nutrition on root growth of crop plants. Adv Agron 110:251–331. https://doi.org/10.1016/B978-0-12-385531-2.00004-9
Fan F, Yu B, Wang B, George TS, Yi H, Xu D, Li D, Song A (2019) Microbial mechanisms of the contrast residue decomposition and priming effect in soils with different organic and chemical fertilization histories. Soil Bio Biochem 135:213–221. https://doi.org/10.1016/j.soilbio.2019.05.001
FAOSTAT (2019) Production: crops. Food and Agriculture Organization of the United Nations, Rome, Italy. http://www.fao.org/faostat/en/#data/QC. Accessed 14 Jan 2019
Felismino MF, Pagliarini MS, Valle CB, Resende RM (2012) Meiotic stability in two valuable interspecific hybrids of Brachiaria. Plant Breed 131:402–408. https://doi.org/10.1111/j.1439-0523.2011.01942.x
Ferreira DF (2011) Sisvar: a computer statistical analysis system. Cienc Agrotech 35:1039–1042
Galdos MV, Brown E, Rosolem CA, Pires LF, Hallett PD, Mooney SJ (2020) Brachiaria species infuence nitrate transport in soil by modifying soil structure with their root system. Sci Rep 10:5072. https://doi.org/10.1038/s41598-020-61986-0
Guimarães CM, Stone LF (2003) Nitrogen fertilization of upland rice under no-tillage system. Rev Bras Eng Agr Amb 7:210–214. https://doi.org/10.1590/S1415-43662003000200004
Guo T, Zhang Q, Ai C, Liang G, He L, Zhou, Wei (2018) Nitrogen enrichment regulates straw decomposition and its associated microbial community in a double-rice cropping system. Sci Rep 8:1847. https://doi.org/10.1038/s41598-018-20293-5
Kluthcouski J, Fancelli AL, Dourado-Neto D, Ribeiro CM, Ferraro LA (2000) Yield of soybean, corn, common bean and rice under no-tillage management. Sci Agr 57:97–104. https://doi.org/10.1590/S0103-90162000000100016
Kluthcouski J, Stone LF (2003) Sustainable management in the soils of Cerrados. In: Kluthcouski J, Stone LF, Aidar H (eds) Crop-livestock system. Embrapa Arroz e Feijão, Santo Antonio do Goias
Kluthcouski J, Aidar H, Thung M, Oliveira FRA, Cobucci T (2006) Early management of nitrogen in main annual crops. Embrapa Arroz e Feijão, Santo Antônio de Goiás
Kumar V, Ladha JK (2011) Chapter six: direct seeding of rice—recent developments and future research needs. Adv Agron 111:297–413. https://doi.org/10.1016/B978-0-12-387689-8.00001-1
Kuzyakov Y, Xu XL (2013) Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytol 198:656–669. https://doi.org/10.1111/nph.12235
Lopes RA, Buzetti S, Teixeira Filho MCM, Benett CGS, Arf MV (2013) Doses, sources and application times of nitrogen on rice grown in no till-age system. Rev Caatinga 26:79–87. https://doi.org/10.1590/S0100-06832013000600023
Makino A, Sakuma H, Sudo E, Mae T (2003) Differences between maize and rice in N-use efficiency for photosysntehsis and protein allocation. Plant Cell Physiol 44:952–956. https://doi.org/10.1093/pcp/pcg113
Malavolta E, Vitti GC, Oliveira SA (1997) Evaluation of nutritional status of plants: principles and applications. Potafós, Piracicaba
Marschner H (2012) Mineral nutrition of higher plants, 3rd edn. Academic Press, London
Mead AD, Drasgow F (1993) Equivalence of computerized and paper-and-pencil cognitive ability tests: a meta-analysis. Psychol Bull 114:449–458
Momesso L, Crusciol CAC, Soratto RP, Vyn TJ, Tanaka KS, Costa CHM, Ferrari Neto J, Cantarella H (2019) Impacts of nitrogen management on no-till maize production following forage cover crops. Agron J 111:639–649. https://doi.org/10.2134/agronj2018.03.0201
Moro E, Crusciol CAC, Cantarella H, Nascente AS (2013) Upland rice under no-tillage preceded by crops for soil cover and nitrogen fertilization. Rev Cienc Solo 37:1669–1677. https://doi.org/10.1590/S0100-06832013000600023
Nascente AS, Crusciol CAC, Cobucci T (2013) The no-tillage system and cover crops: alternatives to increase upland rice yields. Eur J Agron 45:124–131. https://doi.org/10.1016/j.eja.2012.09.004
Nascente AS, Kluthkouski J, Rabelo RR, Oliveira P, Cobucci T, Crusciol CAC (2011) Upland rice cultivars development and yield regarding soil management. Pesqui Agropec Trop 42:186–192. https://doi.org/10.5216/pat.v41i2.6980
Nascente AS, Stone LF (2018) Cover crops as affecting soil chemical and physical properties and development of upland rice and soybean cultivated in rotation. Rice Sci 25:340–349. https://doi.org/10.1016/j.rsci.2018.10.004
Pacheco LP, Monteiro MMS, Petter FA, Nobrega JC, Santos AS (2017) Biomass and nutrient cycling by cover crops in Brazilian cerrado in the State of Piauí. Rev Caatinga 30:13–23. https://doi.org/10.1590/1983-21252017v30n102rc
Pacheco LP, Leandro WM, Machado PLOA, Assis RL, Cobucci T, Madari BE, Petter FA (2011) Biomass production and nutrient accumulation and release by cover crops in the off-season. Pesquis Agropec Bras 46:17–25. https://doi.org/10.1590/S0100-204X2011000100003
Parton WJ, Stewart JWB, Cole CV (1988) Dynamics of C, N, P and S in grassland soils: a model. Biogeochemistry 5:109–131. https://doi.org/10.1007/BF02180320
Rao EVS, Prasad R (1980) Nitrogen leaching losses from conventional and new nitrogenous fertilizers in low-land rice culture. Plant Soil 57:383–392. https://doi.org/10.1007/BF02211695
Rosolem CA, Ritz K, Cantarella H, Galdos MV, Hawkesford MJ, Whalley WR, Mooney SJ (2017) Enhanced plant rooting and crop system management for improved N use efficiency. Adv Agron 146:205–239. https://doi.org/10.1016/bs.agron.2017.07.002
Rosolem CA, Pace L, Crusciol CAC (2004) Nitrogen management in maize cover crop rotations. Plant Soil 264:261–271. https://doi.org/10.1023/B:PLSO.0000047761.50641.a3
Santos AB, Stone LF, Vieira NRA (2006) A cultura do arroz no Brasil. Embrapa Arroz e Feijão, Santo Antônio de Goiás
Sao Miguel ASDC, Pacheco LP, Souza ED, Silva CMR, Carvalho IC (2018) Cover crops in the weed management in soybean culture. Planta Daninha 36:e018172534. https://doi.org/10.1590/s0100-83582018360100072
Soratto RP (2011) Nitrogen early application and sources for common bean in succession to forage grasses in no-tillage system. Thesis (Livre-Docente), Universidade Estadual Paulista, Faculdade de Ciências Agronômicas, Botucatu
Sullivan CAO, Whisson K, Treble K, Roper MM, Micin SF, Ward PR (2017) Biological nitrification inhibitin by weeds: wild radish, brome grass, wild oats and annual ryegrass decrease nitrification rates in the rhizospheres. Crop Pasture Sci 68:798–804. https://doi.org/10.1071/CP17243
Stone LF, Santos AB, Steinmetz S (1980) Effect of cultural practices on water retention capacity of the soil and on yield of upland rice. Pesqui Agropec Bras 15:63–68
Timossi PC, Durigan JC, Leite GJ (2007) Straw formation by brachiaria species to adoption of no-tillage system. Bragantia 66:617–622. https://doi.org/10.1590/S0006-87052007000400012
USDA, United States Department of Agriculture (2014) Keys to soil taxonomy, 12th edn. USDA-Natural Resources Conservation Service, Washington
van Raij B, Andrade JC, Cantarella H, Quaggio JA (2001) Chemical analysis to evaluate soil fertility in tropical soil. Instituto Agronômico de Campinas, Campinas
Verzeaux J, Hirel B, Dubois F, Lea PJ, Tetu T (2017) Agricultural practices to improve nitrogen use efficiency through the use of arbuscular mycorrhizae: basic and agronomic aspects. Plant Sci 264:48–56. https://doi.org/10.1016/j.plantsci.2017.08.004
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38. https://doi.org/10.1097/00010694-193401000-00003
Acknowledgements
This study is supported by the São Paulo Research Foundation (FAPESP; 2015/17953-6) and National Council for Scientific and Technological Development (CNPq) for an award for excellence in research to the second, third, and sixth authors. Publication number 7013 of the Netherlands Institute of Ecology (NIOO-KNAW).
Author information
Authors and Affiliations
Corresponding authors
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Momesso, L., Crusciol, C.A.C., Soratto, R.P. et al. Upland rice yield enhanced by early nitrogen fertilization on previous palisade grass. Nutr Cycl Agroecosyst 118, 115–131 (2020). https://doi.org/10.1007/s10705-020-10088-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-020-10088-4