Selection and Validation of Reference Genes for Quantitative Real-Time PCR in White Clover (Trifolium repens L.) Involved in Five Abiotic Stresses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Abiotic Stress Treatments
2.2. RNA Isolation and cDNA Synthesis
2.3. Selection of Reference Genes and PCR Primer Design
2.4. qRT-PCR Analysis
2.5. Stability Ranking of Candidate Reference Genes
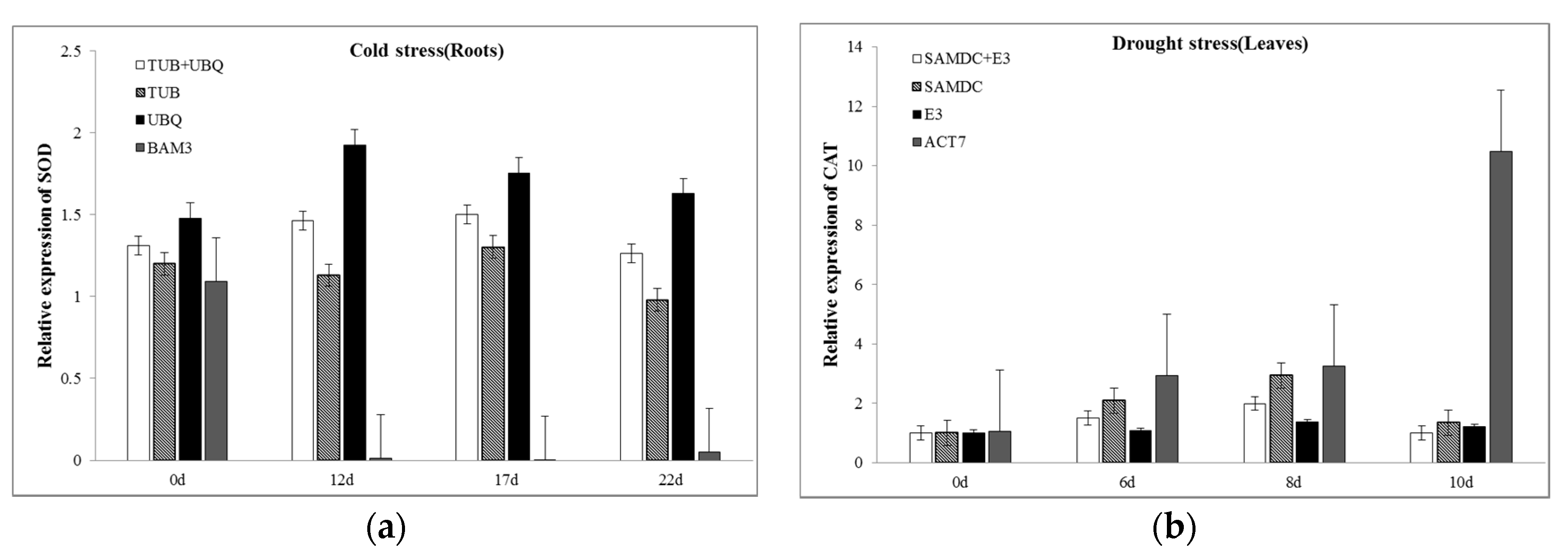
2.6. Validation of Reference Genes by Expression Analysis of Cyt-Cu/Zn SOD and CAT Under Abiotic Stresses
3. Results
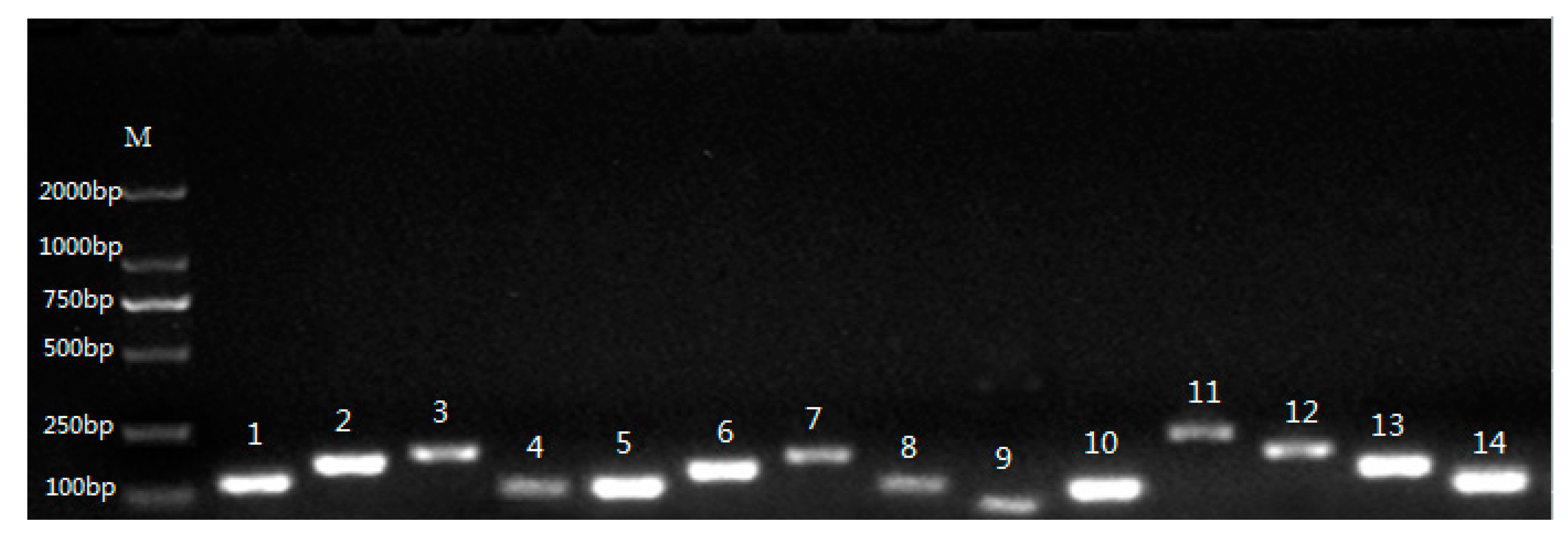
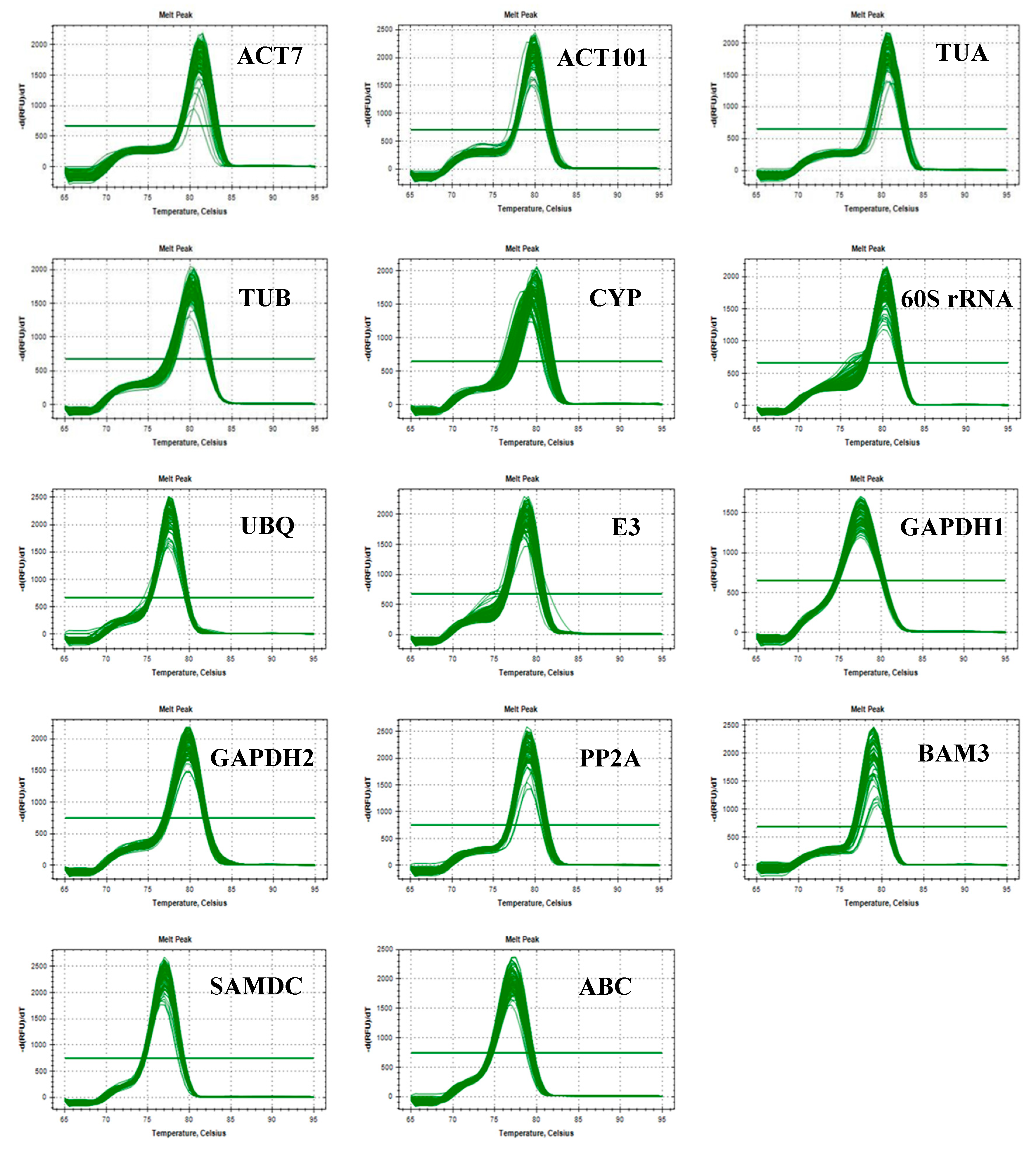
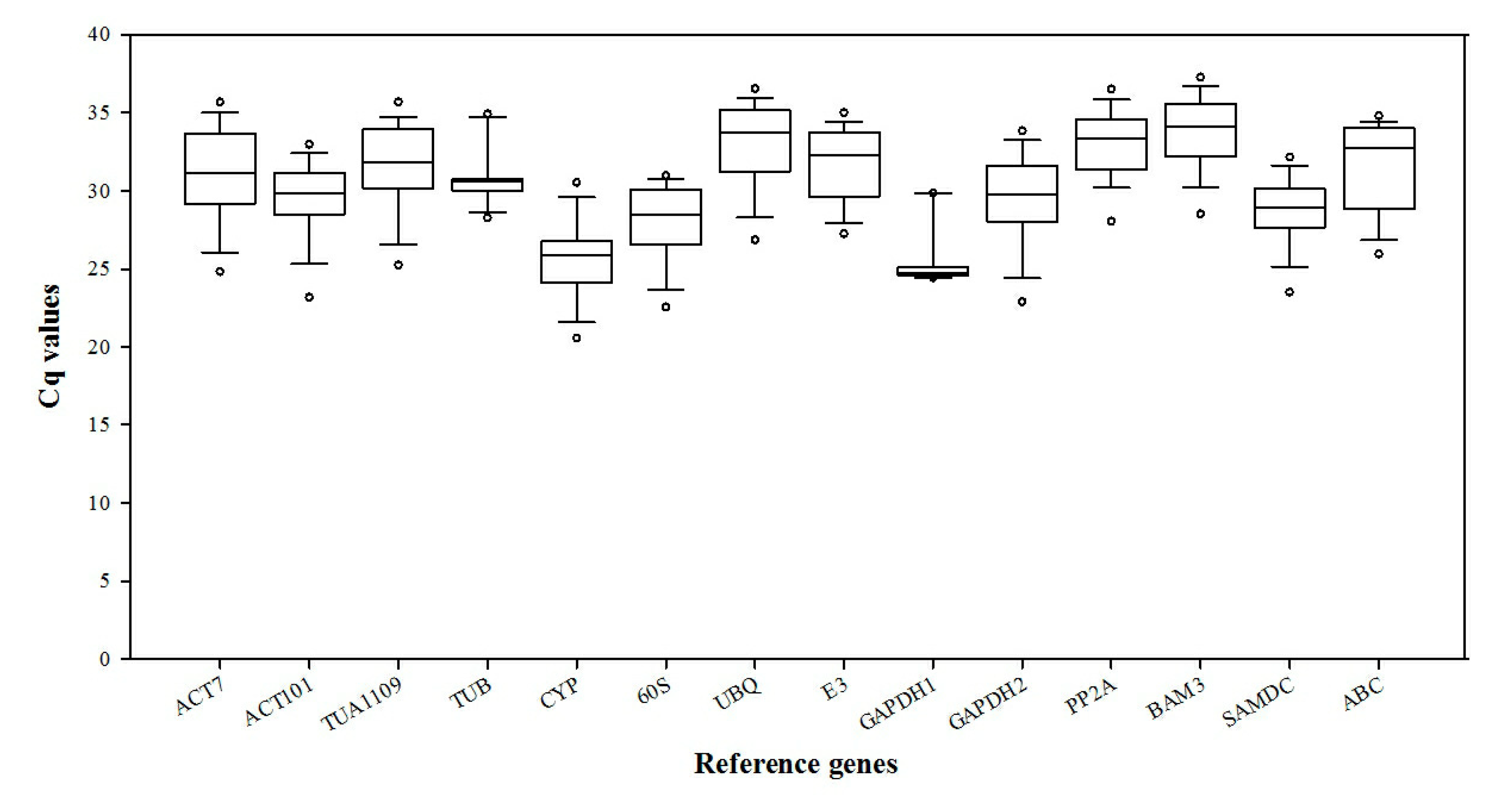
3.1. Verification of PCR Amplicons, Primer Specificity, and Gene-Specific PCR Amplification Efficiency
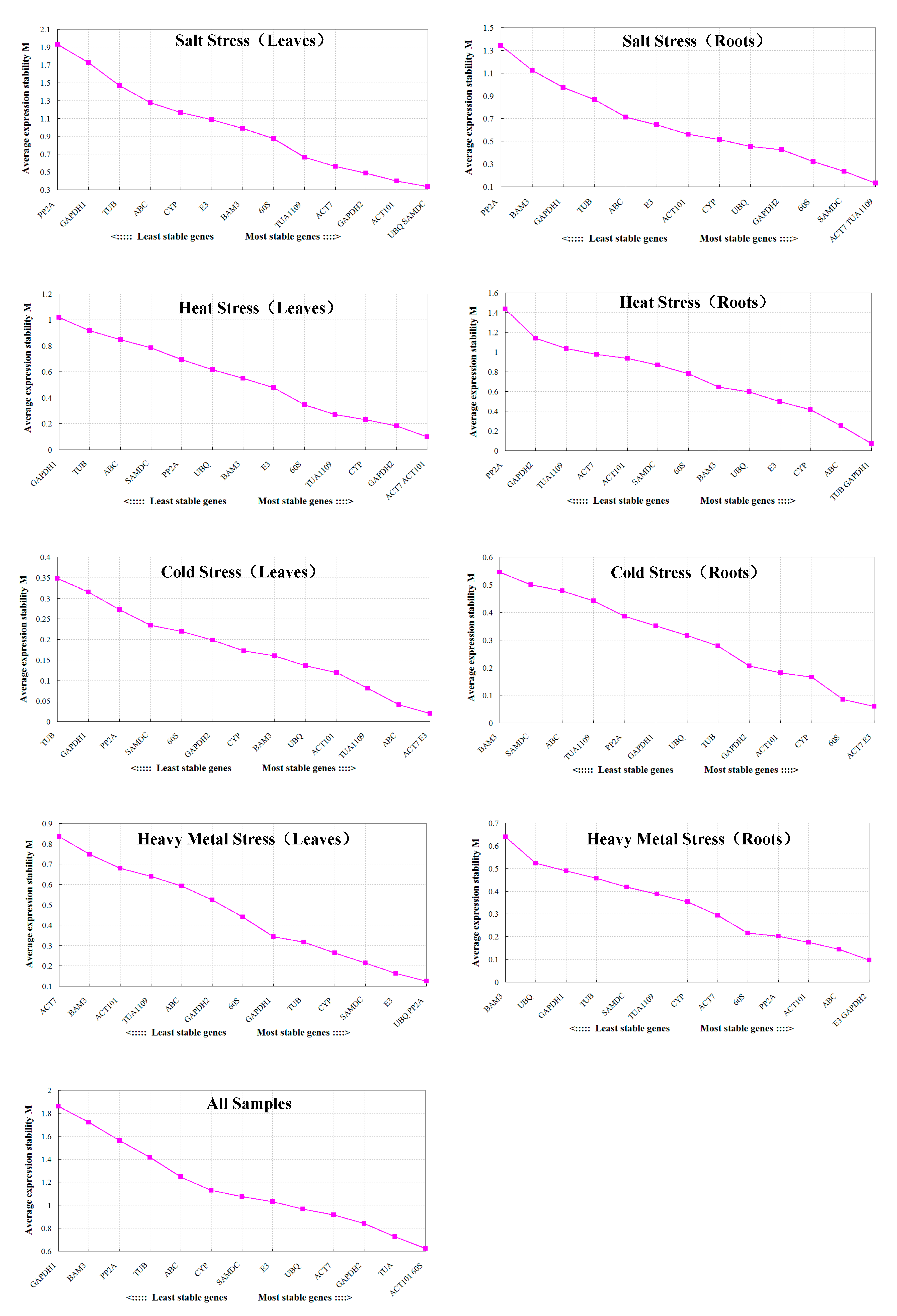
3.2. Stability Ranking of Candidate Reference Genes
3.2.1. GeNorm Analysis
3.2.2. NormFinder Analysis
3.2.3. BestKeeper Analysis
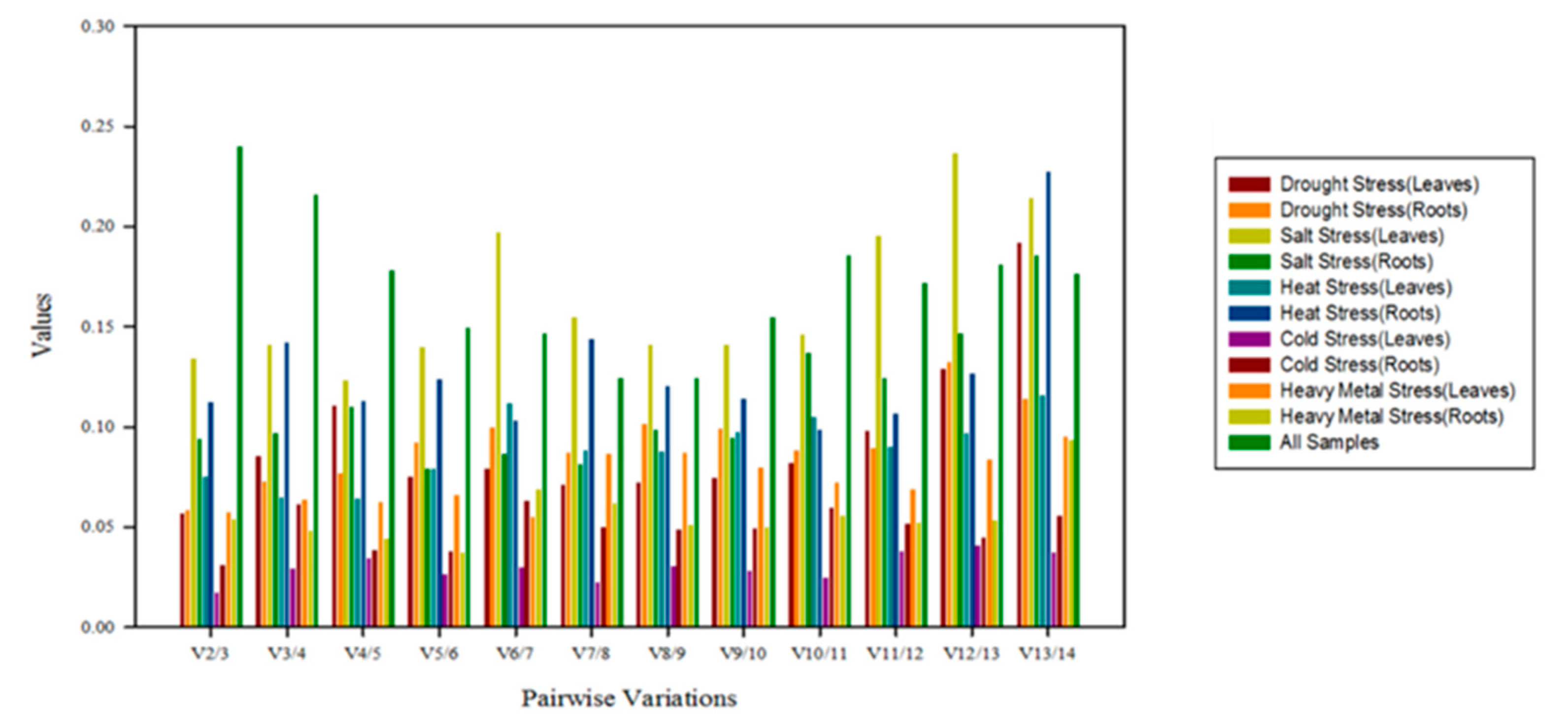
3.2.4. RefFinder Analysis
3.3. Validation of the Reference Genes Identified from this Study
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, L.; Yan, H.; Jiang, X.; Zhang, X.; Zhang, Y.; Huang, X.; Zhang, Y.; Miao, J.; Xu, B.; Frazier, Y. Evaluation of candidate reference genes for normalization of quantitative RT-PCR in switchgrass under various abiotic stress conditions. BioEnergy Res. 2014, 7, 1201–1211. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef]
- Liu, S.; Cai, P.; Hou, N.; Piao, X.; Wang, H.; Hung, T.; Chen, Q. Genome-wide identification and characterization of a panel of house-keeping genes in Schistosoma japonicum. Mol. Biochem. Parasitol. 2012, 182, 75–82. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef]
- Weyrich, A.; Axtner, J.; Sommer, S. Selection and validation of reference genes for real-time RT-PCR studies in the non-model species Delomys sublineatus, an endemic Brazilian rodent. Biochem. Biophys. Res. Commun. 2010, 392, 145–149. [Google Scholar] [CrossRef]
- Zhu, X.; Li, X.; Chen, W.; Chen, J.; Lu, W.; Chen, L.; Fu, D. Evaluation of new reference genes in papaya for accurate transcript normalization under different experimental conditions. PLoS ONE 2012, 7, e44405. [Google Scholar] [CrossRef]
- Li, Q.F.; Sun, S.; Yuan, D.Y.; Yu, H.X.; Gu, M.H.; Liu, Q.Q. Validation of candidate reference genes for the accurate normalization of real-time quantitative RT-PCR data in rice during seed development. Plant Mol. Biol. Rep. 2010, 28, 49–57. [Google Scholar] [CrossRef]
- Cook, N.L.; Robert, V.; Donkin, J.J.; Corinna, V.D.H. Validation of reference genes for normalization of real-time quantitative RT-PCR data in traumatic brain injury. J. Neurosci. Res. 2010, 87, 34–41. [Google Scholar] [CrossRef]
- Zheng, W.J.; Sun, L. Evaluation of housekeeping genes as references for quantitative real time RT-PCR analysis of gene expression in Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 30, 638–645. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Li, Y.P.; Zhang, X.Q.; Ma, X.; Huang, L.K.; Yan, Y.H.; Peng, Y. Chitosan and spermine enhance drought resistance in white clover, associated with changes in endogenous phytohormones and polyamines, and antioxidant metabolism. Funct. Plant Biol. 2018, 45, 1205–1222. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, Y.; He, X.; Yong, B.; Peng, Y.; Zhang, X.; Ma, X.; Yan, Y.; Huang, L.; Nie, G. The hydrogen sulfide, a downstream signaling molecule of hydrogen peroxide and nitric oxide, involves spermidine-regulated transcription factors and antioxidant defense in white clover in response to dehydration. Environ. Exp. Bot. 2019, 161, 255–264. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X.; Huang, L.; Yan, Y. Metabolic pathways regulated by chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef] [PubMed]
- Tony, R.; Karen, S.; Kelly, O.; Dennis, M.; Jaco, V.; Ann, C. Normalisation of real-time RT-PCR gene expression measurements in Arabidopsis thaliana exposed to increased metal concentrations. Planta 2008, 227, 1343–1349. [Google Scholar]
- Narancio, R.; John, U.; Mason, J.; Spangenberg, G. Selection of optimal reference genes for quantitative RT-PCR transcript abundance analysis in white clover (Trifolium repens L.). Funct. Plant Biol. 2018, 45, 737–744. [Google Scholar] [CrossRef]
- Mukesh, J.; Aashima, N.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar]
- Mori, R.; Wang, Q.; Danenberg, K.D.; Pinski, J.K.; Danenberg, P.V. Both beta-actin and GAPDH are useful reference genes for normalization of quantitative RT-PCR in human FFPE tissue samples of prostate cancer. Prostate 2008, 68, 1555–1560. [Google Scholar] [CrossRef]
- Glare, E.M.; Divjak, M.; Bailey, M.J.; Walters, E.H. Beta-Actin and GAPDH housekeeping gene expression in asthmatic airways is variable and not suitable for normalising mRNA levels. Thorax 2002, 57, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Yuan, J.; Niu, P.; Xie, J.; Jiang, W.; Huang, Y.; Bie, Z. Screening suitable reference genes for normalization in reverse transcription quantitative real-time PCR analysis in melon. PLoS ONE 2014, 9, e87197. [Google Scholar] [CrossRef]
- Xiao, Z.; Sun, X.; Liu, X.; Li, C.; He, L.; Chen, S.; Su, J. Selection of reliable reference genes for gene expression studies on rhododendron molle G. Don. Front. Plant Sci. 2016, 7, 1547. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, K.D.; Fernandes de Melo, D.; Morais, V.D.; Vasconcelos, I.M.; Costa, J.H. Selection of suitable soybean EF1α genes as internal controls for real-time PCR analyses of tissues during plant development and under stress conditions. Plant Cell Rep. 2014, 33, 1453. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Reddy, R.A.; Ponnuvel, K.M.; Rohela, G.K.; Shabnam, A.A.; Ghosh, M.; Mishra, R.K. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Mulberry (Morus alba L.) under different abiotic stresses. Mol. Biol. Rep. 2019, 46, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, L.; Wang, X.; Liu, H.; Tian, H.; Wang, J. Selection of reliable reference genes for gene expression analysis in seeds at different developmental stages and across various tissues in Paeonia ostii. Mol. Biol. Rep. 2019, 46, 6003–6011. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Leg. Med. 2012, 126, 943–952. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Andreas, U.; Ioana, C.; Triinu, K.; Jian, Y.; Faircloth, B.C.; Maido, R.; Steven, G.R. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar]
- Chen, Y.; Hu, B.; Tan, Z.; Liu, J.; Yang, Z.; Li, Z.; Huang, B.R. Selection of reference genes for quantitative real-time PCR normalization in creeping bentgrass involved in four abiotic stresses. Plant Cell Rep. 2015, 34, 1825–1834. [Google Scholar] [CrossRef]
- Jacinta, G.; Nicholas, E.; Allen, V.D.; Eduardo, B. Selection and validation of reference genes for gene expression analysis in switchgrass (Panicum virgatum) using quantitative real-time RT-PCR. PLoS ONE 2014, 9, e91474. [Google Scholar]
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Kwon, S.Y.; Kim, S.R. Ectopic expression of a cold-responsive CuZn superoxide dismutase gene, SodCc1, in transgenic rice(Oryza sativa L.). J. Plant Biol. 2009, 52, 154–160. [Google Scholar] [CrossRef]
- Jackson, C.; Dench, J.; Moore, A.L.; Halliwell, B.; Foyer, C.H.; Hall, D.O. Subcellular localisation and identification of superoxide dismutase in the leaves of higher plants. Eur. J. Biochem. 1978, 91, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.S.; Heinen, J.L.; Holaday, A.S.; Burke, J.J.; Allen, R.D. Increased resistance to oxidative stress in transgenic plants that overexpress chloroplastic Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci. USA 1993, 90, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Kornyeyev, D.; Logan, B.P.; Allen, R.D.; Holaday, A.S. Enhanced photochemical light utilization and decreased chilling-inducedphotoinhibition of photosystem II in cotton overexpressing genes encodingchloroplast-targeted antioxidant enzymes. Physiol. Plant 2010, 113, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kirkham, M.B. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Huang, T.; Long, J.; Liu, S.W.; Yang, Z.W.; Zhu, Q.J.; Zhao, X.L.; Peng, C.C. Selection and validation of reference genes for mRNA expression by quantitative real-time PCR analysis in neolamarckia cadamba. Sci. Rep. 2018, 8, 9311. [Google Scholar] [CrossRef]
- Dheda, K.; Huggett, J.F.; Chang, J.S.; Kim, L.U.; Bustin, S.A.; Johnson, M.A.; Rook, G.A.W.; Zumla, A. The implications of using an inappropriate reference gene for real-time reverse transcription PCR data normalization. Anal. Biochem. 2005, 344, 141–143. [Google Scholar] [CrossRef]
- Lü, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Wu, J.; Yang, C.; Pan, H. Selection and validation of reference genes for RT-qPCR analysis of the ladybird beetle Henosepilachna vigintioctomaculata. Front. Physiol. 2018, 9, e01614. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Wang, X.; Chen, J.; Huang, Z.; Huo, H.; Jiang, C.; Huang, H.; Zhang, C.; Wei, S. Selection of reference genes for qPCR normalization in buffalobur (Solanum rostratum Dunal). Sci. Rep. 2019, 9, 6948. [Google Scholar] [CrossRef] [Green Version]
- Artico, S.; Nardeli, S.M.; Brilhante, O.; Grossi-De-Sa, M.F.; Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutum for accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol. 2010, 10, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Abbreviation | Gene Name | Primer Sequence Forward and Reverse | Amplicon Length (bp) | Tm (°C) | Accession Number |
|---|---|---|---|---|---|
| ACT7 | Actin 7 | GGCAGACGCTGAGGATATTCAACC ATGACGTGGTCGGCCAACAATAC | 124 | 60.3 | MT822509 |
| ACT101 | Actin 101 | TGCTTGATTCCGGTGATGGTGTG TTCTCGGCAGAGGTACTGAAGGAG | 163 | 60.3 | MT822510 |
| TUA | Alpha tubulin | TGGAGGAACTGGATCTGGTCTTGG AACAGGACAGCAACATCGGTGTG | 186 | 60.6 | MT822511 |
| TUB | Beta tubulin | CCAGCAGTACCGCAACTTGTCTG ACGACCGTGGCGTGGATCTG | 94 | 62.3 | MT822512 |
| CYP | Cyclophilin | ACGTTGTGTTCGGTCAAGTTGTTG GGCGACGACAACAGGCTTAGAG | 101 | 59.6 | MT822513 |
| 60S rRNA | 60S ribosomal RNA | AACGGTGCTGTGGAGACAATGTAC TTGTGGAACTGCTTAGTGCTCTCC | 134 | 59.5 | MT822514 |
| UBQ | Ubiquitin | ACTGCGTGCAACCAAGGATGATAG TGCCTCGTCTGAAGACTGACCAG | 163 | 60.0 | MT822515 |
| E3 | Ubiquitin | ATTGCCTGCTGATCCTGATCTGC ACCACTGCAACCACACCAAGC | 95 | 60.7 | MT822516 |
| GAPDH1 | Glyceraldehyde 3-phosphate dehydrogenase 1 | GCGTGAACGAGGCTGACTACAAG CCTTGACGATGCCGAACTTCTCC | 117 | 60.8 | MT822517 |
| GAPDH2 | Glyceraldehyde 3-phosphate dehydrogenase 2 | CCATCACTGCCACTCAGAAGACTG AATGTTGAATGAGGCGGCTCTTCC | 80 | 60.1 | MT822518 |
| PP2A | Protein phosphatase 2A | CGGAGCCGGTGTTGTGACAAG AGGCGTGCTCTGTAGGAACTCC | 199 | 61.9 | MT822519 |
| BAM3 | Beta-amylase 3 | TGTTGGTGACTCATGCAGCATTCC GTGGTGTCCTTCCGGCAAGAAC | 158 | 60.8 | MT822520 |
| SAMDC | S-adenosylmethionine decarboxylase | TCAGCAGCCAAGATGACCAACAAC ACAGCAGCACCTTCAACAGAGTTC | 119 | 60.0 | MT822521 |
| ABC | ATP-binding | AAGGATGTACCGCGCCTTCTTATG ATCTCCGCATCTTCCGCACAATAC | 82 | 59.5 | MT822522 |
| Rank | Drought Stress | Salt Stress | Heat Stress | Cold Stress | Heavy metal Stress | All Samples | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | ||
| 1 | E3 (0.029) | UBQ (0.164) | 60S (0.154) | SAMDC (0.136) | 60S (0.106) | UBQ (0.063) | TUA1109 (0.032) | UBQ (0.061) | UBQ (0.156) | ACT101 (0.101) | UBQ (0.049) |
| 2 | SAMDC (0.029) | CYP (0.180) | UBQ (0.286) | CYP (0.136) | TUA1109 (0.156) | BAM3 (0.063) | ACT101 (0.053) | TUB (0.072) | CYP (0.177) | TUA1109 (0.124) | TUB (0.125) |
| 3 | UBQ (0.043) | 60S (0.195) | CYP (0.444) | 60S (0.203) | GAPDH2 (0.171) | CYP (0.136) | CYP (0.081) | PP2A (0.185) | 60S (0.189) | CYP (0.140) | CYP (0.130) |
| 4 | 60S (0.124) | ABC (0.299) | SAMDC (0.487) | ACT101 (0.267) | ACT7 (0.303) | ABC (0.498) | UBQ (0.100) | GAPDH1 (0.226) | PP2A (0.266) | SAMDC (0.198) | GAPDH2 (0.171) |
| 5 | CYP (0.287) | PP2A (0.431) | BAM3 (0.526) | ACT7 (0.350) | E3 (0.316) | 60S (0.512) | ACT7 (0.108) | ACT101 (0.227) | E3 (0.270) | E3 (0.224) | ACT101 (0.349) |
| 6 | GAPDH2 (0.307) | TUB (0.432) | E3 (0.564) | TUA1109 (0.451) | CYP (0.326) | SAMDC (0.544) | E3 (0.111) | ACT7 (0.255) | GAPDH2 (0.372) | PP2A (0.229) | TUA (0.414) |
| 7 | ACT101 (0.477) | GAPDH2 (0.514) | ACT101 (0.686) | GAPDH2 (0.576) | ACT101 (0.384) | E3 (0.553) | ABC (0.115) | CYP (0.272) | GAPDH1 (0.388) | GAPDH2 (0.291) | GAPDH1 (0.487) |
| 8 | PP2A (0.506) | ACT101 (0.536) | GAPDH2 (0.919) | UBQ (0.644) | BAM3 (0.401) | GAPDH1 (0.595) | BAM3 (0.137) | 60S (0.291) | TUB (0.405) | ACT7 (0.295) | PP2A (0.526) |
| 9 | TUA1109 (0.538) | E3 (0.565) | ACT7 (1.029) | TUB (0.657) | UBQ (0.444) | TUB (0.646) | GAPDH2 (0.164) | E3 (0.300) | SAMDC (0.418) | ABC (0.295) | SAMDC (0.527) |
| 10 | ABC (0.584) | TUA1109 (0.662) | TUB (1.257) | GAPDH1 (0.754) | PP2A (0.656) | ACT101 (0.656) | 60S (0.248) | TUA1109 (0.342) | ABC (0.430) | 60S (0.328) | ABC (0.607) |
| 11 | BAM3 (0.604) | SAMDC (0.700) | TUA1109 (1.279) | E3 (0.816) | TUB (0.765) | ACT7 (0.688) | PP2A (0.256) | ABC (0.349) | TUA1109 (0.464) | TUB (0.335) | E3 (0.649) |
| 12 | ACT7 (0.891) | ACT7 (0.765) | ABC (1.288) | ABC (1.025) | SAMDC (0.859) | TUA1109 (0.919) | SAMDC (0.257) | SAMDC (0.362) | ACT101 (0.547) | GAPDH1 (0.386) | ACT7 (0.708) |
| 13 | TUB (1.008) | GAPDH1 (1.103) | GAPDH1 (1.979) | BAM3 (1.182) | ABC (0.873) | GAPDH2 (1.283) | GAPDH1 (0.335) | GAPDH2 (0.399) | BAM3 (0.813) | UBQ (0.532) | 60S (0.773) |
| 14 | GAPDH1 (1.871) | BAM3 (1.112) | PP2A (2.064) | PP2A (1.794) | GAPDH1 (1.117) | PP2A (2.200) | TUB (0.358) | BAM3 (0.537) | ACT7 (0.914) | BAM3 (0.906) | BAM3 (2.841) |
| Rank | Drought Stress | Salt Stress | Heat Stress | Cold Stress | Heavy Metal Stress | All Samples | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | ||
| 1 | GAPDH1 (1.25 ± 0.31) | GAPDH1 (0.73 ± 0.18) | GAPDH1 (0.25 ± 0.06) | GAPDH1 (0.01 ± 0.00) | GAPDH1 (0.20 ± 0.05) | TUB (0.20 ± 0.06) | GAPDH1 (0.03 ± 0.01) | TUB (0.19 ± 0.07) | TUB (0.02 ± 0.01) | TUB (0.06 ± 0.02) | TUB (4.73 ± 1.47) |
| 2 | TUB (2.28 ± 0.67) | BAM3 (1.33 ± 0.46) | PP2A (1.55 ± 4.81) | TUB (0.35 ± 0.11) | TUB (1.06 ± 0.32) | GAPDH1 (0.38 ± 0.09) | TUB (0.18 ± 0.06) | GAPDH1 (0.24 ± 0.07) | GAPDH1 (0.32 ± 0.08) | GAPDH1 (0.18 ± 0.04) | PP2A (5.39 ± 1.78) |
| 3 | E3 (5.89 ± 1.74) | TUB (2.27 ± 0.67) | BAM3 (2.26 ± 6.73) | BAM3 (1.40 ± 0.48) | ACT101 (2.42 ± 0.71) | ABC (0.89 ± 0.30) | PP2A (0.48 ± 0.16) | PP2A (0.48 ± 0.16) | CYP (0.76 ± 0.20) | SAMDC (0.50 ± 0.14) | BAM3 (5.44 ± 1.84) |
| 4 | BAM3 (5.91 ± 1.75) | E3 (2.50 ± 0.69) | SAMDC (2.48 ± 8.60) | ACT101 (2.82 ± 0.83) | ACT7 (2.61 ± 0.78) | E3 (1.71 ± 0.56) | UBQ (0.87 ± 0.29) | UBQ (0.55 ± 0.20) | E3 (0.78 ± 0.26) | TUA1109 (0.64 ± 0.20) | SAMDC (6.17 ± 1.77) |
| 5 | 60S (6.15 ± 1.56) | ABC (2.61 ± 0.73) | ABC (2.78 ± 9.20) | SAMDC (3.20 ± 0.94) | TUA1109 (2.99 ± 0.92) | UBQ (1.99 ± 0.68) | ACT101 (0.92 ± 0.29) | ACT101 (0.90 ± 0.28) | UBQ (0.97 ± 0.35) | CYP (0.91 ± 0.23) | E3 (6.20 ± 1.97) |
| 6 | UBQ (6.20 ± 1..80) | UBQ (3.86 ± 1.08) | TUB (2.80 ± 0.84) | PP2A (3.49 ± 1.15) | GAPDH2 (3.03 ± 0.88) | CYP (2.16 ± 0.57) | TUA1109 (1.01 ± 0.34) | CYP (1.03 ± 0.31) | SAMDC (1.05 ± 0.32) | BAM3 (1.54 ± 0.52) | GAPDH1 (6.32 ± 1.63) |
| 7 | CYP (6.21 ± 1.38) | CYP (4.73 ± 1.03) | E3 (6.16 ± 1.93) | 60S (3.53 ± 0.98) | CYP (3.36 ± 0.84) | BAM3 (2.23 ± 0.81) | ACT7 (1.30 ± 0.43) | ABC (1.11 ± 0.38) | PP2A (1.06 ± 0.38) | ACT7 (1.66 ± 0.51) | ACT101 (6.85 ± 2.01) |
| 8 | PP2A (6.71 ± 2.09) | PP2A (5.12 ± 1.52) | CYP (6.59 ± 1.61) | ACT7 (3.57 ± 1.11) | 60S (4.13 ± 1.16) | SAMDC (3.78 ± 1.16) | E3 (1.34 ± 0.43) | ACT7 (1.16 ± 0.41) | BAM3 (1.21 ± 0.41) | PP2A (1.72 ± 0.60) | UBQ (7.14 ± 2.35) |
| 9 | SAMDC (6.91 ± 1.81) | 60S (5.48 ± 1.28) | 60S (6.65 ± 1.86) | CYP (3.72 ± 0.94) | BAM3 (4.26 ± 1.43) | ACT7 (4.07 ± 1.34) | GAPDH2 (1.36 ± 0.41) | BAM3 (1.25 ± 0.46) | ABC (2.04 ± 0.69) | ACT101 (1.78 ± 0.52) | 60S (7.21 ± 2.02) |
| 10 | TUA1109 (7.99 ± 2.25) | TUA1109 (5.90 ± 1.58) | UBQ (6.80 ± 2.24) | TUA1109 (3.76 ± 1.19) | E3 (4.79 ± 1.48) | 60S (4.37 ± 1.29) | BAM3 (1.38 ± 0.43) | E3 (1.28 ± 0.45) | 60S (2.06 ± 0.63) | E3 (1.85 ± 0.61) | TUA1109 (7.35 ± 2.31) |
| 11 | ABC (8.02 ± 2.21) | GAPDH2 (6.18 ± 1.52) | ACT101 (8.65 ± 2.57) | UBQ (3.92 ± 1.32) | UBQ (4.80 ± 1.54) | ACT101 (4.55 ± 1.40) | CYP (1.40 ± 0.38) | GAPDH2 (1.29 ± 0.42) | GAPDH2 (2.66 ± 0.83) | ABC (2.03 ± 0.69) | CYP (7.48 ± 1.90) |
| 12 | GAPDH2 (8.09 ± 2.06) | ACT101 (6.40 ± 1.60) | ACT7 (8.84 ± 2.72) | GAPDH2 (4.32 ± 1.27) | PP2A (5.02 ± 1.70) | TUA1109 (4.62 ± 1.50) | ABC (1.43 ± 0.43) | SAMDC (1.31 ± 0.40) | TUA1109 (2.68 ± 0.90) | GAPDH2 (2.23 ± 0.65) | ACT7 (7.70 ± 2.40) |
| 13 | ACT101 (8.37 ± 2.19) | ACT7 (6.59 ± 1.79) | GAPDH2 (9.52 ± 2.82) | E3 (4.97 ± 1.58) | ABC (6.66 ± 2.02) | PP2A (4.74 ± 1.55) | 60S (1.79 ± 0.53) | TUA1109 (1.33 ± 0.46) | ACT101 (3.15 ± 1.00) | 60S (2.52 ± 0.70) | GAPDH2 (7.84 ± 2.30) |
| 14 | ACT7 (9.56 ± 2.63) | SAMDC (7.12 ± 1.79) | TUA1109 (9.72 ± 3.11) | ABC (5.23 ± 1.71) | SAMDC (6.89 ± 1.98) | GAPDH2 (6.33 ± 2.05) | SAMDC (1.93 ± 0.55) | 60S (1.53 ± 0.46) | ACT7 (3.91 ± 1.31) | UBQ (2.71 ± 0.92) | ABC (8.33 ± 2.61) |
| Treatment | Ranking order | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| All samples | 60S | ACT101 | E3 | SAMDC | CYP | TUA1109 | TUB | UBQ | GAPDH2 | ACT7 | GAPDH1 | PP2A | BAM3 | ABC |
| Drought stress (Leaves) | SAMDC | E3 | UBQ | 60S | GAPDH2 | CYP | GAPDH1 | ACT101 | TUB | TUA1109 | PP2A | BAM3 | ABC | ACT7 |
| Drought stress (Roots) | UBQ | 60S | PP2A | CYP | ACT101 | ABC | GAPDH2 | TUB | GAPDH1 | E3 | BAM3 | SAMDC | TUA1109 | ACT7 |
| Salt stress (Leaves) | UBQ | SAMDC | 60S | ACT101 | CYP | BAM3 | GAPDH1 | E3 | TUB | GAPDH2 | ACT7 | PP2A | TUA1109 | ABC |
| Salt stress (Roots) | SAMDC | 60S | ACT7 | TUA1109 | CYP | ACT101 | GAPDH1 | TUB | GAPDH2 | UBQ | BAM3 | E3 | ABC | PP2A |
| Heat stress (Leaves) | 60S | ACT7 | GAPDH2 | ACT101 | TUA1109 | CYP | E3 | GAPDH1 | TUB | BAM3 | UBQ | PP2A | SAMDC | ABC |
| Heat stress (Roots) | UBQ | BAM3 | TUB | GAPDH1 | CYP | ABC | E3 | 60S | SAMDC | ACT101 | ACT7 | TUA1109 | GAPDH2 | PP2A |
| Cold stress (Leaves) | TUA1109 | ACT7 | ACT101 | E3 | CYP | UBQ | ABC | GAPDH1 | BAM3 | PP2A | TUB | GAPDH2 | 60S | SAMDC |
| Cold stress (Roots) | TUB | UBQ | ACT7 | ACT101 | PP2A | GAPDH1 | E3 | CYP | 60S | ABC | GAPDH2 | SAMDC | TUA1109 | BAM3 |
| Heavy metal stress (Leaves) | UBQ | CYP | PP2A | E3 | TUB | GAPDH1 | 60S | SAMDC | GAPDH2 | ABC | TUA1109 | BAM3 | ACT101 | ACT7 |
| Heavy metal stress (Roots) | ACT101 | E3 | TUA1109 | CYP | GAPDH2 | SAMDC | TUB | PP2A | ABC | GAPDH1 | ACT7 | 60S | BAM3 | UBQ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pu, Q.; Li, Z.; Nie, G.; Zhou, J.; Liu, L.; Peng, Y. Selection and Validation of Reference Genes for Quantitative Real-Time PCR in White Clover (Trifolium repens L.) Involved in Five Abiotic Stresses. Plants 2020, 9, 996. https://doi.org/10.3390/plants9080996
Pu Q, Li Z, Nie G, Zhou J, Liu L, Peng Y. Selection and Validation of Reference Genes for Quantitative Real-Time PCR in White Clover (Trifolium repens L.) Involved in Five Abiotic Stresses. Plants. 2020; 9(8):996. https://doi.org/10.3390/plants9080996
Chicago/Turabian StylePu, Qi, Zhou Li, Gang Nie, Jiqiong Zhou, Lin Liu, and Yan Peng. 2020. "Selection and Validation of Reference Genes for Quantitative Real-Time PCR in White Clover (Trifolium repens L.) Involved in Five Abiotic Stresses" Plants 9, no. 8: 996. https://doi.org/10.3390/plants9080996