Physicochemical Parameters Affecting the Distribution and Diversity of the Water Column Microbial Community in the High-Altitude Andean Lake System of La Brava and La Punta

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
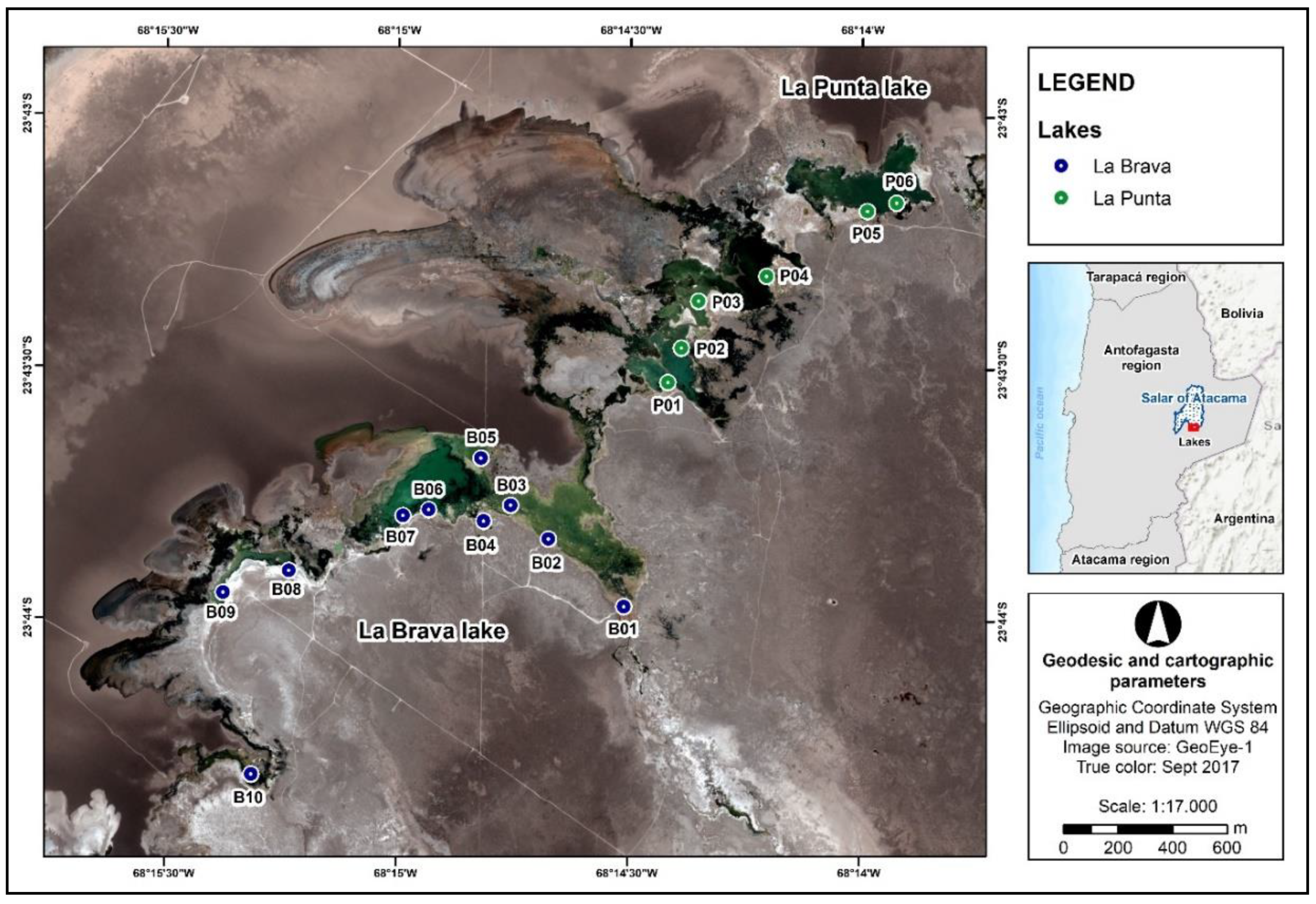
2.1. Area of Study and Sample Collection
2.2. Physicochemical Parameters of the Water Columns
2.3. DNA Extraction
2.4. Sequencing and Bioinformatic Analysis
2.5. Taxonomic Analysis
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Parameters of the Water Columns
3.2. Next-Generation Sequencing Data
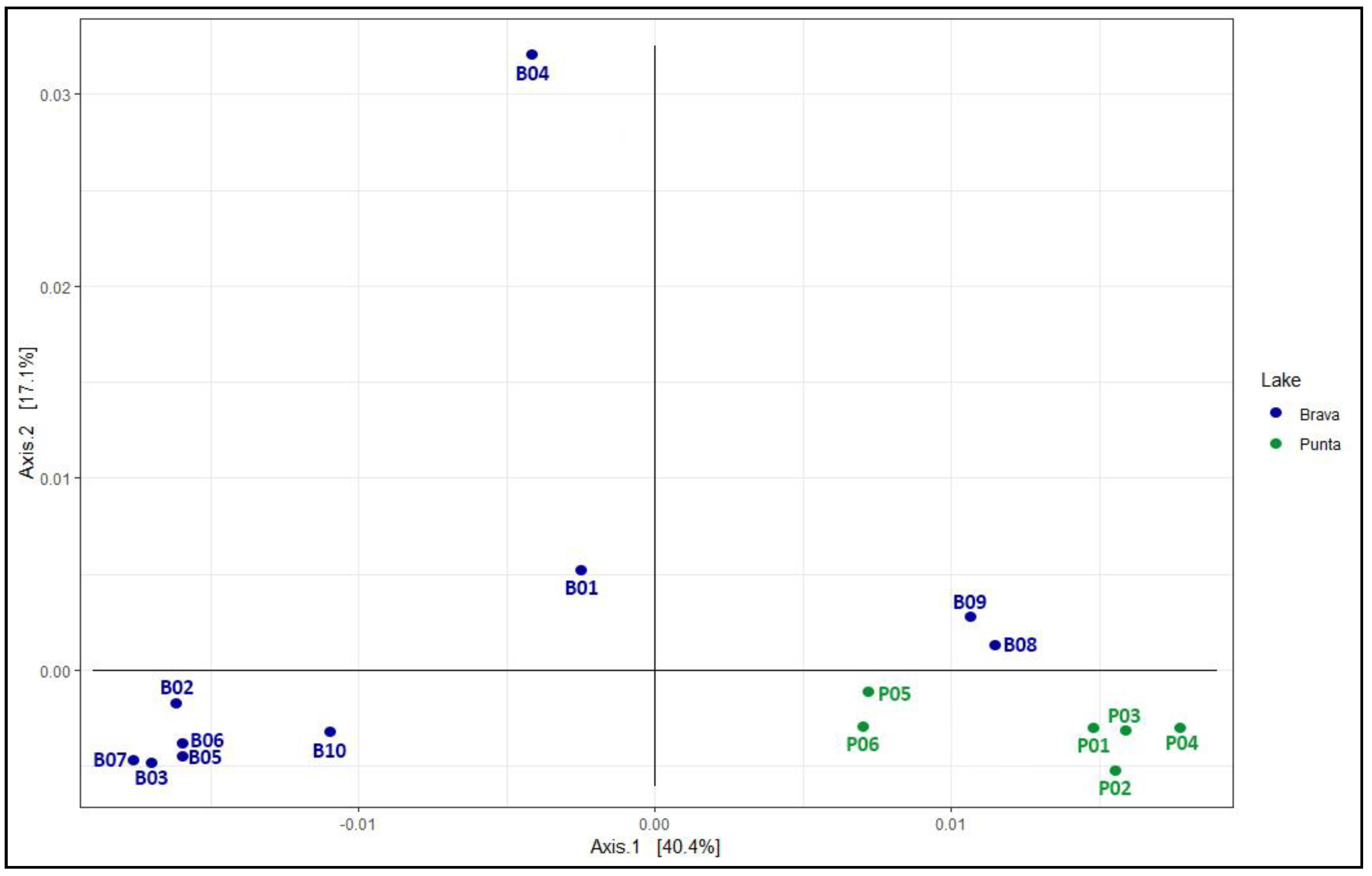
3.3. Diversity Comparisons
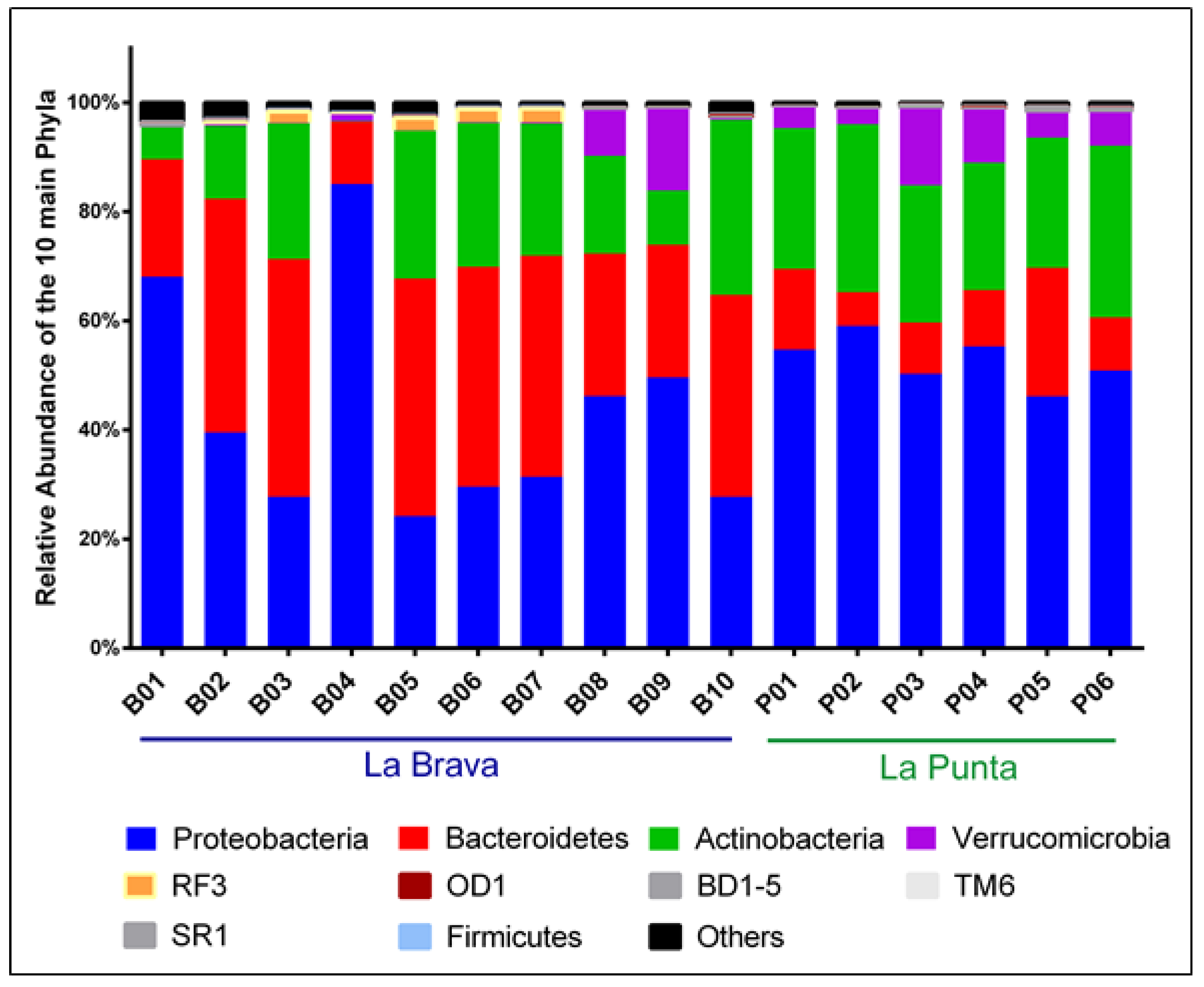
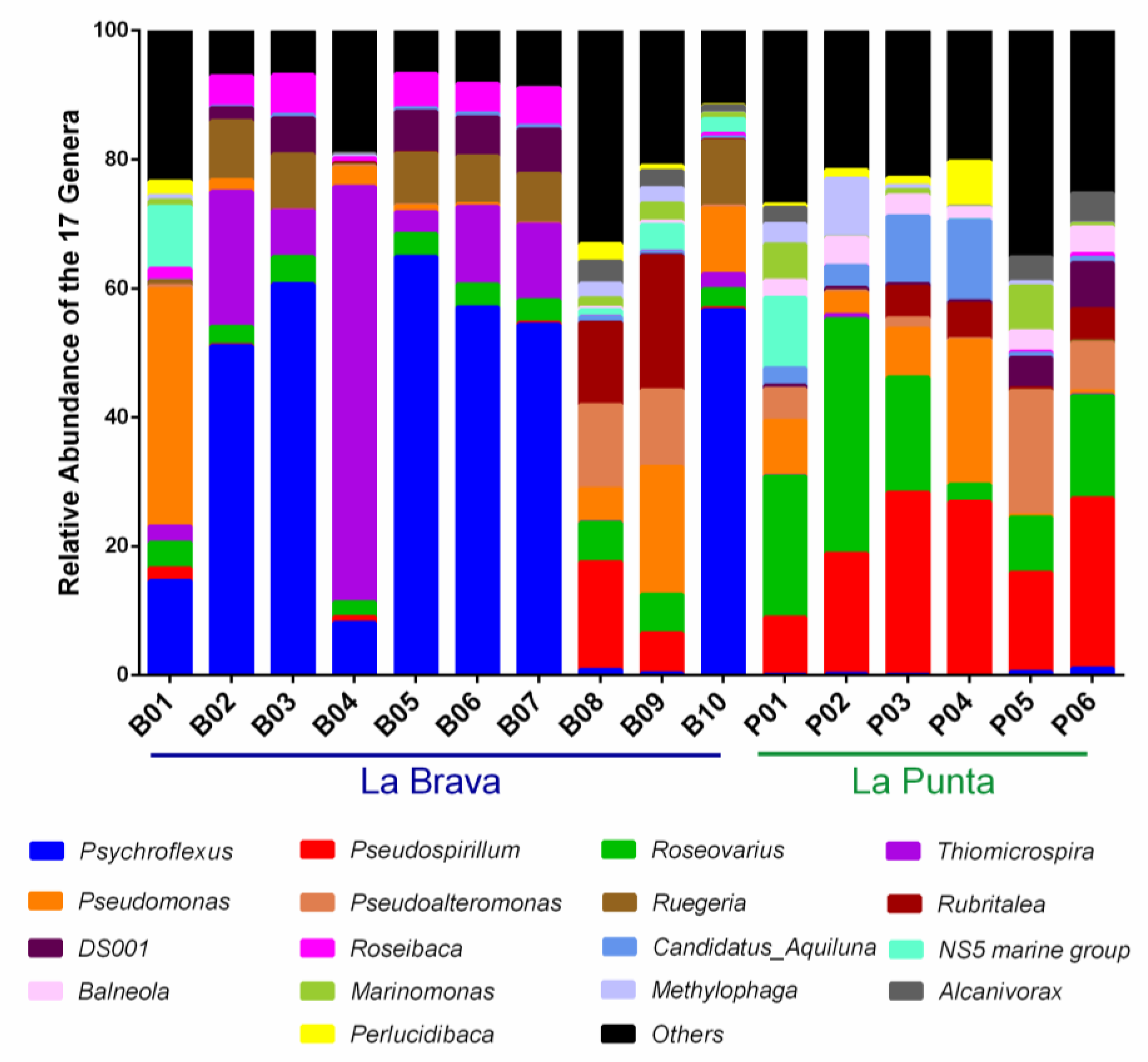
3.4. Comparison between Lakes at the Genus Level
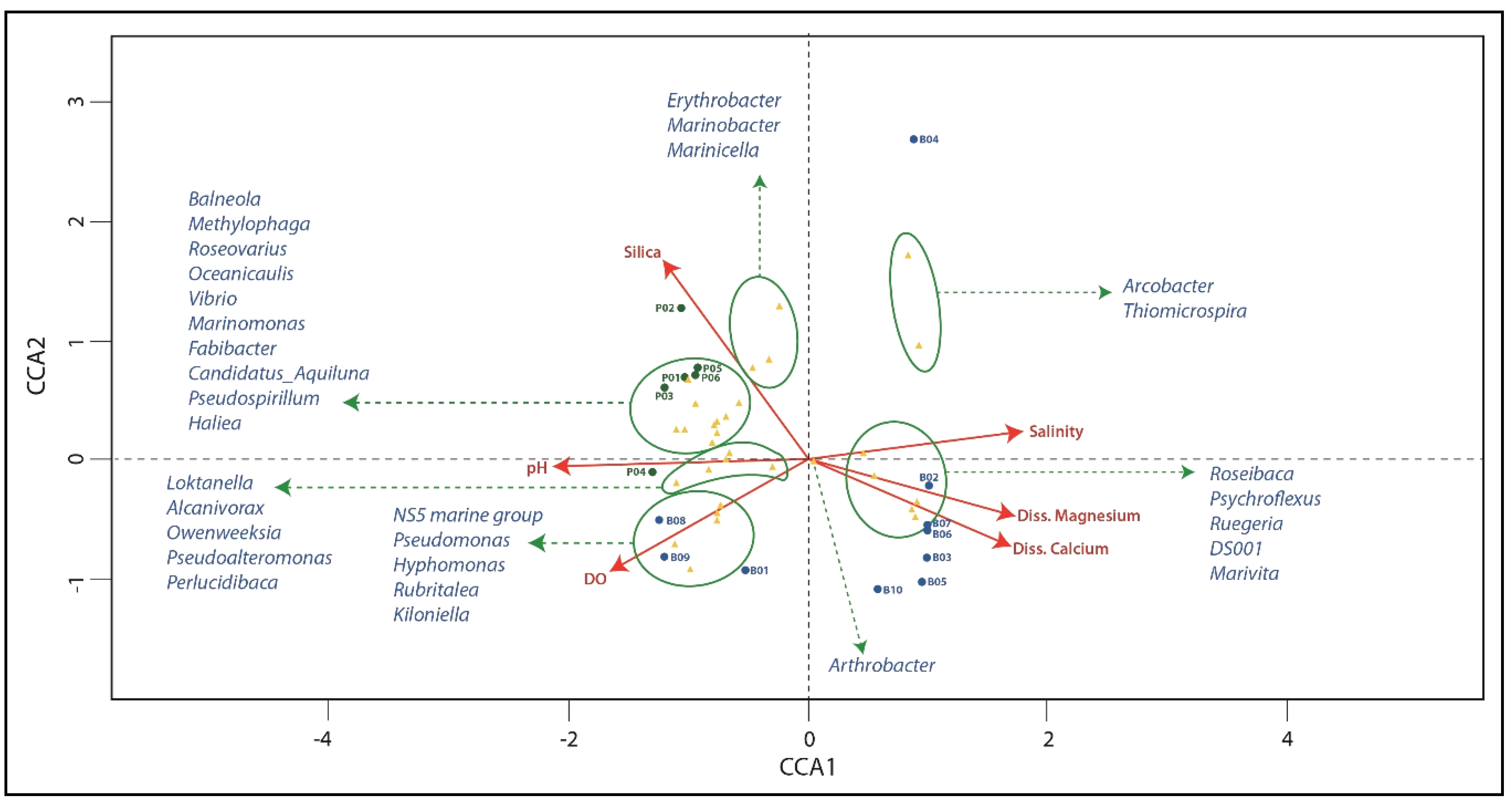
3.5. Distribution of Genera with Respect to Environmental Variables
4. Discussion
4.1. Physicochemical Parameters of Water and Microbial Diversity
4.2. Distribution of Genera with Respect to Environmental Variables
4.3. Role of pH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Albarracín, V.H.; Kurth, D.; Ordoñez, O.F.; Belfiore, C.; Luccini, E.; Salum, G.M.; Piacentini, R.D.; Farías, M.E. High-up: A Remote Reservoir of Microbial Extremophiles in Central Andean Wetlands. Front. Microbiol. 2015, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bull, A.T.; Andrews, B.A.; Dorador, C.; Goodfellow, M. Introducing the Atacama Desert. Antonie Van Leeuwenhoek. 2018, 111, 1269–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risacher, F.; Alonso, H.; Salazar, C. The Origin of Brines and Salts in Chilean Salars: A Hydrochemical Review. Earth-Sci. Rev. 2003, 63, 249–293. [Google Scholar] [CrossRef]
- Anderson, M.; Low, R.; Foot, S. Sustainable Groundwater Development in Arid, High Andean Basins. Geol. Soc. London Spec. Publ. 2002, 193, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Soto, J.; Román-Figueroa, C.; Paneque, M. A Model for Estimating the Vegetation Cover in the High-Altitude Wetlands of the Andes (HAWA). Land 2019, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.; Low, R.; Foot, S. Sustainable Groundwater Development in Arid, High Andean Basins. In Sustainable Groundwater Development, 1st ed.; Hiscock, K., Rivett, M., Davison, R., Eds.; Geological Society: London, UK, 2017; pp. 133–144. ISBN 1862390975. [Google Scholar]
- Colburn, E.A. Temporary Waters. In Ecosystem Ecology, 1st ed.; Jørgensen, S.E., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2008; pp. 427–439. ISBN 9780444534668. [Google Scholar]
- Harris, G. Lagoons. In Ecosystem Ecology, 1st ed.; Jørgensen, S.E., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2008; pp. 296–303. ISBN 9780444534668. [Google Scholar]
- Melack, G. Saline and Soda Lakes. In Ecosystem Ecology, 1st ed.; Jørgensen, S.E., Ed.; Elsevier B.V: Amsterdam, The Netherlands, 2008; pp. 380–384. ISBN 9780444534668. [Google Scholar]
- Soluciones en Gestión Ambiental S.A. (SGA). Línea Base: Ecosistemas acuáticos continentales. Continuidad Operacional Compañía Minera Zaldívar. Capítulo 3-11; Servicio de Evaluación Ambiental: Santiago, Chile, 2018; p. 171. (In Spanish) [Google Scholar]
- Bryanskaya, A.V.; Malup, T.K.; Lazareva, E.V.; Taran, O.P.; Rozanov, A.S.; Efimov, V.M.; Peltek, S.E. The Role of Environmental Factors in the Composition of Microbial Communities in Saline Lakes in the Novosibirsk Region (Russia). BMC Microbiol. 2016, 16 (Suppl. 1), 4. [Google Scholar] [CrossRef] [Green Version]
- Bull, A.T.; Asenjo, J.A.; Goodfellow, M.; Gomez-Silva, B. The Atacama Desert: Technical Resources and the Growing Importance of Novel Microbial Diversity. Annu. Rev. Microbiol. 2016, 70, 215–234. [Google Scholar] [CrossRef] [Green Version]
- Rampelotto, P.H. Extremophiles and Extreme Environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Ferrer, G.C.; Moreno, J.R.; Farías, M.E.; Albarracín, V.H. The Diversity of Microbial Extremophiles. In Molecular Diversity of Environmental Prokaryotes, 1st ed.; Rodriguez, T.B., Silva, A.E., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 87–126. ISBN 9781482233223. [Google Scholar]
- Demergasso, C.; Casamayor, E.O.; Chong, G.; Galleguillos, P.; Escudero, L.; Pedrós-Alió, C. Distribution of Prokaryotic Genetic Diversity in Athalassohaline Lakes of the Atacama Desert, Northern Chile. FEMS Microbiol. Ecol. 2004, 48, 57–69. [Google Scholar] [CrossRef]
- Demergasso, C.; Dorador, C.; Meneses, D.; Blamey, J.; Cabrol, N.; Escudero, L.; Chong, G. Prokaryotic Diversity Pattern in High-Altitude Ecosystems of the Chilean Altiplano. J. Geophys. Res. 2010, 115, 1–14. [Google Scholar] [CrossRef]
- Demergasso, C.; Escudero, L.; Casamayor, E.O.; Chong, G.; Balagué, V.; Pedrós-Alió, C. Novelty and Spatio–Temporal Heterogeneity in the Bacterial Diversity of Hypersaline Lake Tebenquiche (Salar de Atacama). Extremophiles 2008, 12, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Dorador, C.; Busekow, A.; Vila, I.; Imhoff, J.F.; Witzel, K.-P. Molecular Analysis of Enrichment Cultures of Ammonia Oxidizers from the Salar de Huasco, a High Altitude Saline Wetland in Northern Chile. Extremophiles 2008, 12, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorador, C.; Vila, I.; Imhoff, J.F.; Witzel, K.-P. Cyanobacterial Diversity in Salar de Huasco, A High Altitude Saline Wetland in Northern Chile: An Example of Geographical Dispersion? FEMS Microbiol. Ecol. 2008, 64, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Dorador, C.; Vila, I.; Remonsellez, F.; Imhoff, J.F.; Witzel, K.-P. Unique Clusters of Aarchaea in Salar de Huasco, An Athalassohaline Evaporitic Basin of the Chilean Altiplano. FEMS Microbiol. Ecol. 2010, 73, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, A.B.; Rasuk, M.C.; Visscher, P.T.; Contreras, M.; Novoa, F.; Poire, D.G.; Patterson, M.M.; Ventosa, A.; Farías, M.E. Microbial Diversity in Sediment Ecosystems (Evaporites Domes, Microbial Mats, and Crusts) of Hypersaline Laguna Tebenquiche, Salar de Atacama, Chile. Front. Microbiol. 2016, 7, 1284. [Google Scholar] [CrossRef] [Green Version]
- Farías, M.E.; Contreras, M.; Rasuk, M.C.; Kurth, D.; Flores, M.R.; Poiré, D.G.; Novoa, F.; Visscher, P.T. Characterization of Bacterial Diversity Associated With Microbial Mats, Gypsum Evaporites and Carbonate Microbialites in Thalassic Wetlands: Tebenquiche and La Brava, Salar de Atacama, Chile. Extremophiles 2014, 18, 311–329. [Google Scholar] [CrossRef]
- Farías, M.E.; Rasuk, M.C.; Gallagher, K.L.; Contreras, M.; Kurth, D.; Fernández, A.B.; Poiré, D.; Novoa, F.; Pieter, T.V. Prokaryotic Diversity and Biogeochemical Characteristics of Benthic Microbial Ecosystems at La Brava, a Hypersaline Lake at Salar De Atacama, Chile. PLoS ONE 2017, 12, 0186867. [Google Scholar] [CrossRef] [Green Version]
- Vierheilig, J.; Savio, D.; Ley, R.E.; Mach, R.L.; Farnleitner, A.H.; Reischer, G.H. Potential Applications of Next Generation DNA Sequencing of 16S rRNA Gene Amplicons in Microbial Water Quality Monitoring. Water Sci. Technol. 2015, 72, 1962–1972. [Google Scholar] [CrossRef] [Green Version]
- Naghoni, A.; Emtiazi, G.; Amoozegar, M. Microbial diversity in the hypersaline Lake Meyghan, Iran. Sci. Rep. 2017, 7, 11522. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Zhang, X.; Liu, J.; Long, Q.; Chen, L.; Liu, D.; Zhu, D. Microbial community structure and diversity within hypersaline Keke Salt Lake environments. Can. J. Microbiol. 2017, 63, 895–908. [Google Scholar] [CrossRef]
- Zhu, D.; Han, R.; Long, Q.; Gao, X.; Xing, J.; Shen, G.; Li, Y.; Wang, R. An evaluation of the core bacterial communities associated with hypersaline environments in the Qaidam Basin, China. Arch. Microbiol. 2020, 3. [Google Scholar] [CrossRef] [PubMed]
- Rathour, R.; Gupta, J.; Mishra, A.; Rajeev, A.; Dupont, C.; Thakur, I. A comparative metagenomic study reveals microbial diversity and their role in the biogeochemical cycling of Pangong lake. Sci. Total Environ. 2020, 731, 139074. [Google Scholar] [CrossRef] [PubMed]
- Boutaiba, S.; Hacene, H.; Bidle, K.; Maupin-Furlow, J. Microbial diversity of the hypersaline Sidi Ameur and Himalatt Salt Lakes of the Algerian Sahara. J. Arid. Environ. 2011, 75, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casamayor, E.; Triadó-Margarit, X.; Castañeda, C. Microbial biodiversity in saline shallow lakes of the Monegros Desert, Spain. FEMS Microbiol. Ecol. 2013, 85, 503–518. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, P.; Acosta, E.; Dorador, C.; Sommaruga, R. Large Differences in Bacterial Community Composition Among Three Nearby Extreme Waterbodies of the High Andean Plateau. Front. Microbiol. 2016, 7, 976. [Google Scholar] [CrossRef]
- Aguilar, P.; Dorador, C.; Vila, I.; Sommaruga, R. Bacterioplankton composition in tropical high-elevation lakes of the Andean plateau. FEMS Microbiol. Ecol. 2018, 94, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.L.; Zwart, G.; Schauer, M.; Kamst-Van Agterveld, M.P.; Hahn, M.W. Bacterioplankton Community Composition Along a Salinity Gradient of Sixteen High-Mountain Lakes Located on the Tibetan Plateau, China. Appl. Environ. Microbiol. 2006, 72, 5478–5485. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yao, T.; Jiao, N.; Zhu, L.; Hu, A.; Liu, X. Salinity Impacts the Bacterial Community Composition in Five High-Altitude Lakes from the Tibetan Plateau in Western China. Geomicrobiol. J. 2013, 30, 462–469. [Google Scholar] [CrossRef]
- Tang, X.; Xie, G.; Shao, K.; Sai, B.; Chen, Y.; Gao, G. Influence of Salinity on the Bacterial Community Composition in Lake Bosten, A Large Oligosaline Lake in Arid Northwestern China. Appl. Environ. Microbiol. 2012, 78, 4748–4751. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.-P.; Liu, Y.; Miao, L.-L.; Wang, F.; Chu, L.-M.; Wang, J.-L.; Liu, Z.-P. Prokaryotic Community Structure Driven by Salinity and Ionic Concentrations in Plateau Lakes of the Tibetan Plateau. Appl. Environ. Microbiol. 2016, 82, 1846–1858. [Google Scholar] [CrossRef] [Green Version]
- Babidge, S. Contested Value and an Ethics of Resources: Water, Mining and Indigenous People in the Atacama Desert, Chile. Aust. J. Anthropol. 2015, 27, 84–103. [Google Scholar] [CrossRef]
- Alonso, H.; Risacher, F. Geoquimica del Salar de Atacama, Part 1: Origen de Los Componentes y Balance Salino. Rev. Geol. Chile 1996, 23, 113–122. [Google Scholar]
- Dirección General de Aguas (DGA). Diagnóstico y sistematización de información de planes de alerta temprana vigentes con condicionamiento de derechos; Ministerio de Obras Públicas, Gobierno de Chile: Santiago, Chile, 2014; p. 534. (In Spanish) [Google Scholar]
- Cáceres, L.; Gómez-Silva, B.; Garró, X.; Rodríguez, V.; Monardesm, V.; McKay, C.P. Relative Humidity Patterns and Fog Water Precipitation in the Atacama Desert and Biological Implications. J. Geophys. Res. 2007, 112, G04. [Google Scholar] [CrossRef] [Green Version]
- Lara, J.; Escudero-González, L.; Ferrero, M.; Chong-Díaz, G.; Pedrós-Alió, C.; Demergasso, C. Enrichment of Arsenic Transforming and Resistant Heterotrophic Bacteria from Sediments of Two Salt Lakes in Northern Chile. Extremophiles 2012, 16, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Covacevich, F.; Silva, R.I.; Cumino, A.; Caló, G.; Negri, R.M.; Salerno, G.L. First Archaeal rDNA Sequences from Argentine Coastal Waters: Unexpected PCR Characterization Using Eukaryotic Primers. Ciencias Mar. 2012, 38, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 22rd ed.; American Public Health Association American Water Works Association Water Environment Federation: Washington, DC, USA, 2012; ISBN 9780875530130. [Google Scholar]
- Williams, W.D. Conductivity and Salinity of Australian Salt Lakes. Aust. J. Mar. Freshw. Res. 1986, 37, 177–182. [Google Scholar] [CrossRef]
- Gottler, R.A. 3010 Introduction. In Standard Methods for the Examination of Water and Wastewater, 23rd ed.; Rice, E., Baird, R., Eaton, A., Eds.; American Public Health Association: Washington, DC, USA, 2017; pp. 1–3. ISBN 9780875532875. [Google Scholar]
- Baxter, T.E. 2340 Hardness. In Standard Methods for the Examination of Water and Wastewater, 23rd ed.; Rice, E., Baird, R., Eaton, A., Eds.; American Public Health Association: Washington, DC, USA, 2017; pp. 1–3. ISBN 9780875532875. [Google Scholar]
- Nakatsu, C.H.; Byappanahalli, M.N.; Nevers, M.B. Bacterial Community 16S rRNA Gene Sequencing Characterizes Riverine Microbial Impact on Lake Michigan. Front. Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; González, A.; Goodrich, J.; Gordon, J.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Quince, C.; Lanzen, A.; Davenport, R.J.; Turnbaugh, P.J. Removing Noise from Pyrosequenced Amplicons. BMC Bioinform. 2011, 12, 38. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster Than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A Flexible Tool for Aligning Sequences to a Template Alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Lee, S. Estimating the Number of Classes Via Sample Coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Hejcmanová-Nezerková, P.; Hejcman, M. A canonical correspondence analysis (CCA) of the vegetation–environment relationships in Sudanese savannah, Senegal. S. Afr. J. Bot. 2006, 72, 256–262. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: https://www.r-project.org (accessed on 15 May 2020).
- Mandakovic, D.; Rojas Alvarado, C.; Maldonado, J.; Latorre, M.; Travisany, D.; Delage, E.; Bihouée, A.; Jean, G.; Díaz, F.; Fernández-Gómez, B.; et al. Structure and Co-Occurrence Patterns in Microbial Communities Under Acute Environmental Stress Reveal Ecological Factors Fostering Resilience. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Fernández, A.B.; Kurth, D.; Contreras, M.; Novoa, F.; Poiré, D.; Farías, M. Bacterial Diversity in Microbial Mats and Sediments from the Atacama Desert. Microb. Ecol. 2016, 71, 44–56. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Kurth, D.; Flores, M.R.; Contreras, M.; Novoa, F.; Poire, D.; Alfarías, M. Microbial Characterization of Microbial Ecosystems Associated to Evaporites Domes of Gypsum in Salar de Llamara in Atacama Desert. Microb. Ecol. 2014, 68, 483–494. [Google Scholar] [CrossRef]
- Salas, J.; Guimerá, J.; Cornellá, O.; Aravena, R.; Guzmán, E.; Tore, C.; von Igel, W.; Moreno, R. Hidrogeología del Sistema lagunar del margen este del Salar de Atacama (Chile). Boletín Geológico Min. 2010, 121, 357–372. [Google Scholar]
- Pop Ristova, P.; Pichler, T.; Friedrich, M.; Bühring, S. Bacterial Diversity and Biogeochemistry of Two Marine Shallow-Water Hydrothermal Systems off Dominica (Lesser Antilles). Front. Microbiol. 2017, 8, 2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; Liu, Y.; Lin, X.; Zhang, H.; Zeng, J.; Hou, J.; Yang, Y.; Yao, T.; Knight, R.; Chu, H. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ. Microbiol. 2012, 14, 2457–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.P. Psychroflexus. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–6. [Google Scholar]
- Brinkhoff, T.; Kuever, J.; Muyzer, G.; Jannasch, H.W. Thiomicrospira. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–10. [Google Scholar]
- Palleroni, N.J. Pseudomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 323–378. [Google Scholar]
- Zhong, Z.; Liu, Y.; Wang, F.; Zhou, Y.-G.; Liu, H.-C.; Liu, Z.-P. Psychroflexus salis sp. nov. and Psychroflexus planctonicus sp. nov., isolated from a salt lake. Int. J. Syst. Evol. Microbiol. 2015, 66, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauro, F.; Cappelletti, M.; Ghezzi, D.; Columbu, A.; Hong, P.; Zowawi, H.; Carbone, C.; Piccini, L.; Vergara, F.; Zannoni, D.; et al. Microbial diversity and biosignatures of amorphous silica deposits in orthoquartzite caves. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Vasanthi, N.; Saleena, L.; Raj, S. Silica Solubilization Potential of Certain Bacterial Species in the Presence of Different Silicate Minerals. Silicon 2016, 10, 267–275. [Google Scholar] [CrossRef]
- Yoon, J.; Kang, S.; Lee, J. Marivita geojedonensis sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2012, 63, 1568–1575. [Google Scholar] [CrossRef]
- Budinoff, C.; Dunlap, J.; Hadden, M.; Buchan, A. Marivita roseacus sp. nov., of the family Rhodobacteraceae, isolated from a temperate estuary and an emended description of the genus Marivita. J. Gen. Appl. Microbiol. 2011, 57, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Liu, Y.; Hou, T.; Liu, H.; Zhou, Y.; Wang, F.; Liu, Z. Marivita lacus sp. nov., isolated from a saline lake. Int. J. Syst. Evol. Microbiol. 2015, 65, 1889–1894. [Google Scholar] [CrossRef]
- Hedlund, B.P.; Yoon, J.; Kasai, H. Rubritalea. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–7. [Google Scholar]
- Wiese, J.; Thiel, V.; Gärtner, A.; Schmaljohann, R.; Imhoff, J.F. Kiloniella laminariae gen. nov., sp. nov., an alphaproteobacterium from the marine macroalga Laminaria saccharina. Int. J. Syst. Evol. Microbiol. 2009, 59, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Weiner, R.M. Hyphomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–13. [Google Scholar]
- Vandamme, P.; Dewhirst, F.E.; Paster, B.J.; On, S.L. Arcobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–8. [Google Scholar]
- Holmer, M.; Storkholm, P. Sulphate reduction and sulphur cycling in lake sediments: A review. Freshw. Biol. 2001, 46, 431–451. [Google Scholar] [CrossRef]
- Fillinger, L.; Hug, K.; Griebler, C. Selection imposed by local environmental conditions drives differences in microbial community composition across geographically distinct groundwater aquifers. FEMS Microbiol. Ecol. 2019, 95, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Podell, S.; Emerson, J.B.; Jones, C.M.; Ugalde, J.A.; Welch, S.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. Seasonal fluctuations in ionic concentrations drive microbial succession in a hypersaline lake community. ISME J. 2014, 8, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Pan, L.; Song, M.; Tian, C.; Gao, S. Microbiota assemblages of water, sediment, and intestine and their associations with environmental factors and shrimp physiological health. Appl. Microbiol. Biotechnol. 2014, 102, 8585–8598. [Google Scholar] [CrossRef] [PubMed]
- Labrenz, M.; Lawson, P.A.; Tindall, B.J.; Hirsch, P. Roseibaca ekhonensis gen. nov., sp. nov., an alkalitolerant and aerobic bacteriochlorophyll a-producing alphaproteobacterium from hypersaline Ekho Lake. Int. J. Syst. Evol. Microbiol. 2009, 59, 1935–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, J.S.; Whitman, W.B. Ruegeria. In Bergey’s Manual of Systematics of Archaea and Bacteria; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 1–25. [Google Scholar]
- Silva-Castro, G.; Uad, I.; Gonzalez-Martinez, A.; Rivadeneyra, A.; Gonzalez-Lopez, J.; Rivadeneyra, M. Bioprecipitation of calcium carbonate crystals by bacteria isolated from saline environments grown in culture media amended with seawater and real brine. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Arias, D.; Cisternas, L.A.; Miranda, C.; Rivas, M. Biomineralization of calcium and magnesium crystals from seawater by halotolerant bacteria isolated from Atacama Salar (Chile). Desalination 2017, 405, 1–9. [Google Scholar] [CrossRef]
- Arias, D.; Cisternas, L.; Miranda, C.; Rivas, M. Bioprospecting of ureolytic bacteria from Laguna Salada for biomineralization Applications. Front. Bioeng. Biotechnol. 2019, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Al Disi, Z.; Jaoua, S.; Bontognali, T.; Attia, E.; Al-Kuwari, H.; Zouari, N. Evidence of a Role for Aerobic Bacteria in High Magnesium Carbonate Formation in the Evaporitic Environment of Dohat Faishakh Sabkha in Qatar. Front. Environ. Sci. 2017, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yan, H.; Zhou, J.; Tucker, M.E.; Han, M.; Zhao, H.; Mao, G.; Zhao, Y.; Han, Z. Bio-Precipitation of calcium and magnesium ions through extracellular and intracellular process induced by Bacillus licheniformis SRB2. Minerals 2019, 9, 526. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wu, L.; Liu, Y.; Zhang, P.; Li, Q.; Zhou, J.; Hess, N.J.; Hazen, T.C.; Yang, W.; Chakraborty, R. Microbial Interactions with Dissolved Organic Matter Drive Carbon Dynamics and Community Succession. Front. Microbiol. 2018, 9, 1234. [Google Scholar] [CrossRef] [PubMed]
- Zamarreño, D.V.; Inkpen, R.; May, E. Carbonate Crystals Precipitated by Freshwater Bacteria and Their Use as a Limestone Consolidant. Appl. Environ. Microbiol. 2009, 75, 5981–5990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Thiotrichalesord. nov. In Bergey’s Manual of Systematics Bacteriology, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J.R., Eds.; Springer: New York, NY, USA, 2005; Volume 2, Part B; pp. 131–210. [Google Scholar] [CrossRef]
- Thiel, V.; Tank, M.; Neulinger, S.C.; Gehrmann, L.; Dorador, C.; Imhoff, J.F. Unique Communities of Anoxygenic Phototrophic Bacteria in Saline Lakes of Salar de Atacama (Chile): Evidence for a New Phylogenetic Lineage of Phototrophic Gammaproteobacteria from puf LM Gene Analyses. FEMS Microbiol. Ecol. 2010, 74, 510–522. [Google Scholar] [CrossRef]
- Ren, L.; Jeppesen, E.; He, D.; Wang, J.; Liboriussen, L.; Xing, P.; Wu, Q.L. pH influences the importance of niche-related and neutral processes in lacustrine bacterioplankton assembly. Appl. Environ. Microbiol. 2015, 9, 3104–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratzke, C.; Gore, J. Modifying and reacting to the environmental pH can drive bacterial interactions. PLoS Biol. 2018, 16, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Calviño, D.; Bååth, E. Growth response of the bacterial community to pH in soils differing in pH. FEMS Microbiol. Ecol. 2010, 73, 149–156. [Google Scholar] [CrossRef]
- Jin, Q.; Kirk, M.F. pH as a Primary Control in Environmental Microbiology: 1. Thermodynamic Perspective. Front. Environ. Sci. 2018, 6, 1–15. [Google Scholar] [CrossRef]
- Bethke, C.M.; Sanford, R.A.; Kirk, M.F.; Jin, Q.; Flynn, T.M. The thermodynamic ladder in geomicrobiology. Am. J. Sci. 2011, 311, 183–210. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, H.; Dong, H.; Liu, Y. A comprehensive census of lake microbial diversity on a global scale. Sci. China Life Sci. 2019, 62, 1320–1331. [Google Scholar] [CrossRef]
- Sorokin, D.; Berben, T.; Melton, E.; Overmars, L.; Vavourakis, C.; Muyzer, G. Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Grant, W.; Duckworth, A.; Owenson, G. Microbial diversity of soda lakes. Extremophiles 1998, 2, 191–200. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Sampling Points Characteristics | ||
|---|---|---|---|
| Hydrological Dynamics | Ecological Disaggregation Characteristics * | ||
| Inlet | Main Body | Isolated Body | |
| La Brava | B01 | B01 to B07 | B08 to B10 |
| La Punta | - | P01 to P04 | P05 to P06 |
| Lake | Sampling Points | pH | DISS Oxygen (mg/L) | Salinity (g/L) | EC (mS/cm) | Total Silica (mg/L) | Ammonium (mg/L) | Nitrite (mg/L) | Nitrate (mg/L) | Total Nitrogen (mg/L) | P-PO4 Phosphates (mg/L) | Phosphorus (mg/L) | DISS Calcium (mg/L) | DISS Magnesium (mg/L) | Hardness (CaCO3 mg/L) | Alkalinity Carbonates mg/L CaCO3 | Alkalinity Bicarbonates mg/L CaCO3- | Total Alkalinity mg/L CaCO3 | TOC (mg/L) | Total Solids (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| La Brava | B01 | 8.06 | 3.09 | 19.08 | 30.32 | 68.90 | <0.09 | <0.01 | 0.25 | 1.00 | <0.09 | <0.05 | 219.88 | 633.20 | 3156.60 | <1.00 | 317.40 | 317.40 | 5.00 | 5.00 |
| B02 | 7.64 | 1.73 | 52.84 | 77.32 | 91.80 | <0.09 | <0.01 | 1.03 | 1.70 | <0.09 | <0.05 | 805.00 | 2151.90 | 10,871.60 | <1.00 | 556.90 | 556.90 | 15.20 | 4.00 | |
| B03 | 7.55 | 1.61 | 50.87 | 74.67 | 74.60 | <0.09 | <0.01 | 0.63 | 31.20 | 0.11 | 0.13 | 441.80 | 1608.90 | 7728.60 | <1.00 | 503.40 | 503.40 | 7.66 | 4.00 | |
| B04 | 7.63 | 1.74 | 52.90 | 77.40 | 67.40 | <0.09 | <0.01 | 0.39 | 1.70 | <0.09 | <0.05 | 336.28 | 1340.10 | 6358.20 | <1.00 | 409.00 | 409.00 | 6.25 | 6.00 | |
| B05 | 7.62 | 3.94 | 77.84 | 110.40 | 86.60 | <0.09 | <0.01 | 1.05 | 3.10 | <0.09 | 0.20 | 816.90 | 2347.50 | 11,706.80 | <1.00 | 674.70 | 674.70 | 17.00 | 6.00 | |
| B06 | 7.86 | 1.58 | 77.30 | 109.70 | 87.80 | <0.09 | <0.01 | 1.05 | 3.40 | 0.10 | 0.11 | 835.40 | 2,456.60 | 12,202.30 | <1.00 | 674.70 | 674.70 | 17.40 | 4.00 | |
| B07 | 7.92 | 3.00 | 27.70 | 42.70 | 89.60 | <0.09 | <0.01 | 0.96 | 2.40 | <0.09 | 0.12 | 758.60 | 4545.00 | 20,610.50 | <1.00 | 621.20 | 621.20 | 15.40 | 6.00 | |
| B08 | 7.99 | 5.96 | 14.65 | 23.77 | 83.80 | <0.09 | <0.01 | 0.24 | 1.00 | <0.09 | <0.05 | 255.77 | 549.00 | 2899.40 | 46.90 | 381.30 | 428.20 | 5.53 | 5.00 | |
| B09 | 8.18 | 6.35 | 22.39 | 35.11 | 86.30 | <0.09 | <0.01 | 0.38 | 0.50 | <0.09 | 0.10 | 323.86 | 694.30 | 3667.80 | 89.50 | 379.10 | 468.60 | 7.14 | 5.00 | |
| B10 | 7.64 | 4.82 | 50.87 | 47.28 | 78.00 | <0.09 | <0.01 | 0.44 | 2.10 | <0.09 | 0.07 | 383.70 | 1070.50 | 5366.40 | <1.00 | 385.60 | 385.60 | 5.00 | 4.00 | |
| La Punta | P01 | 8.19 | 4.01 | 23.40 | 36.57 | 127.40 | <0.09 | <0.01 | 0.50 | 1.80 | <0.09 | 0.16 | 336.15 | 724.90 | 3824.50 | 10.70 | 334.40 | 345.10 | 7.64 | 4.00 |
| P02 | 8.21 | 3.71 | 23.46 | 36.66 | 127.60 | <0.09 | <0.01 | 0.53 | 1.40 | <0.09 | <0.05 | 358.44 | 729.70 | 3899.90 | 49.00 | 300.30 | 349.30 | 9.04 | 10.00 | |
| P03 | 8.35 | 4.06 | 17.10 | 27.41 | 110.80 | <0.09 | <0.01 | 0.33 | 1.50 | <0.09 | <0.05 | 284.66 | 462.60 | 2615.80 | 63.90 | 305.70 | 369.60 | 5.84 | 3.00 | |
| P04 | 8.27 | 4.25 | 13.08 | 21.42 | 100.50 | <0.09 | <0.01 | 0.24 | 1.10 | <0.09 | 0.11 | 242.26 | 397.08 | 2240.10 | 36.20 | 317.40 | 353.60 | 14.50 | 5.00 | |
| P05 | 8.07 | 3.14 | 40.10 | 60.00 | 103.80 | <0.09 | <0.01 | 0.85 | 2.80 | <0.09 | <0.05 | 340.20 | 771.35 | 4025.90 | <1.00 | 488.80 | 488.80 | 11.70 | 6.00 | |
| P06 | 8.17 | 3.55 | 40.93 | 61.15 | 107.20 | <0.09 | <0.01 | 0.95 | 3.80 | <0.09 | <0.05 | 484.50 | 1141.20 | 5909.30 | <1.00 | 519.70 | 519.70 | 15.40 | 7.00 | |
 | ||||||||||||||||||||
| Lake | Sample Points | Observed Out | Chao Index | Shannon Index | Simpson Index |
|---|---|---|---|---|---|
| La Brava | B01 | 1183 | 1234.43 | 4.24 | 0.95 |
| B02 | 1006 | 1085.33 | 3.98 | 0.92 | |
| B03 | 935 | 1046.12 | 3.89 | 0.92 | |
| B04 | 924 | 1093.36 | 3.62 | 0.87 | |
| B05 | 985 | 1120.05 | 3.76 | 0.91 | |
| B06 | 983 | 1104.11 | 3.88 | 0.93 | |
| B07 | 896 | 936.67 | 3.90 | 0.93 | |
| B08 | 948 | 1043.00 | 4.15 | 0.96 | |
| B09 | 909 | 970.09 | 4.20 | 0.95 | |
| B10 | 1186 | 1235.10 | 3.43 | 0.87 | |
| La Punta | P01 | 828 | 918.29 | 4.26 | 0.96 |
| P02 | 940 | 1071.26 | 4.04 | 0.95 | |
| P03 | 799 | 901.69 | 4.22 | 0.96 | |
| P04 | 835 | 954.02 | 4.09 | 0.95 | |
| P05 | 887 | 997.53 | 3.98 | 0.94 | |
| P06 | 868 | 992.32 | 4.07 | 0.95 |
| La Brava-La Punta | La Brava | La Punta | Main | Isolated | |||||
|---|---|---|---|---|---|---|---|---|---|
| (Main-Isolated) | (Main-Isolated) | La Brava-La Punta | La Brava-La Punta | ||||||
| Genus | %Cum Sum | Genus | %Cum Sum | Genus | %Cum Sum | Genus | %Cum Sum | Genus | %Cum Sum |
| Psychroflexus | 26.37 | Psychroflexus | 18.48 | Thiomicrospira | 20.51 | Psychroflexus | 23.65 | Psychroflexus | 23.87 |
| Thiomicrospira | 36.34 | Pseudospirillum | 28.07 | Psychroflexus | 40.47 | Thiomicrospira | 44.82 | Pseudomonas | 37.46 |
| Pseudospirillum | 45.83 | Rubritalea | 37.55 | Pseudomonas | 56.69 | Pseudospirillum | 52.17 | Pseudospirillum | 45.93 |
| Pseudomonas | 52.77 | Pseudomonas | 45.51 | NS5 marine group | 60.92 | Roseovarius | 56.67 | Rubritalea | 53.63 |
| Roseovarius | 58.15 | Pseudoalteromonas | 52.08 | Pseudoalteromonas | 64.51 | Pseudomonas | 60.88 | Pseudoalteromonas | 58.76 |
| Ruegeria | 61.85 | Roseovarius | 57.26 | Ruegeria | 68.01 | Pseudoalteromonas | 64.91 | NS5 marine group | 62.27 |
| Pseudoalteromonas | 65.42 | Kiloniella | 60.55 | Pseudospirillum | 71.17 | Ruegeria | 68.62 | Thiomicrospira | 65.42 |
| Rubritalea | 68.74 | Candidatus_Aquiluna | 63.81 | DS001 | 73.16 | DS001 | 71.00 | Ruegeria | 68.34 |
| Candidatus_Aquiluna | 71.28 | Thiomicrospira | 66.80 | Roseibaca | 75.13 | Roseibaca | 73.21 | Roseovarius | 71.05 |
| Roseibaca | 73.41 | Ruegeria | 69.58 | Marinicella | 75.05 | Roseibaca | 73.61 | ||
| DS001 | 75.36 | DS001 | 71.70 | Kiloniella | 76.01 | ||||
| NS5 marine group | 73.48 | ||||||||
| Perlucidibaca | 75.09 | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez Salazar, R.; Aguirre, C.; Soto, J.; Salinas, P.; Salinas, C.; Prieto, H.; Paneque, M. Physicochemical Parameters Affecting the Distribution and Diversity of the Water Column Microbial Community in the High-Altitude Andean Lake System of La Brava and La Punta. Microorganisms 2020, 8, 1181. https://doi.org/10.3390/microorganisms8081181
Núñez Salazar R, Aguirre C, Soto J, Salinas P, Salinas C, Prieto H, Paneque M. Physicochemical Parameters Affecting the Distribution and Diversity of the Water Column Microbial Community in the High-Altitude Andean Lake System of La Brava and La Punta. Microorganisms. 2020; 8(8):1181. https://doi.org/10.3390/microorganisms8081181
Chicago/Turabian StyleNúñez Salazar, Reynaldo, Carlos Aguirre, Jorge Soto, Pamela Salinas, Carlos Salinas, Humberto Prieto, and Manuel Paneque. 2020. "Physicochemical Parameters Affecting the Distribution and Diversity of the Water Column Microbial Community in the High-Altitude Andean Lake System of La Brava and La Punta" Microorganisms 8, no. 8: 1181. https://doi.org/10.3390/microorganisms8081181